Microbial Induced Carbonate Precipitation Using a Native Inland Bacterium for Beach Sand Stabilization in Nearshore Areas

 ,
,
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of a Suitable Urease Active Bacterium
2.1.1. Soil Sampling and Isolation of Ureolytic Bacteria
2.1.2. Identification of Bacteria by 16S rRNA Sequencing
2.1.3. Cultivation of Bacteria and Assessing the Potential for Microbial Induced Carbonate Precipitation
2.1.4. Microbial Cell Growth and Urease Activity Measurement
2.2. Small Scale Sand Solidification Test
2.3. Effect of Mg2+ on Microbial Induced Carbonate Precipitation
2.4. Monitoring Methods
2.4.1. Needle Penetration Device
2.4.2. Scanning Electron Microscopy (SEM) Analyses
2.4.3. X-ray Diffraction (XRD) Analyses
2.4.4. X-ray Fluorescence (XRF) Analyses
3. Results and Discussion
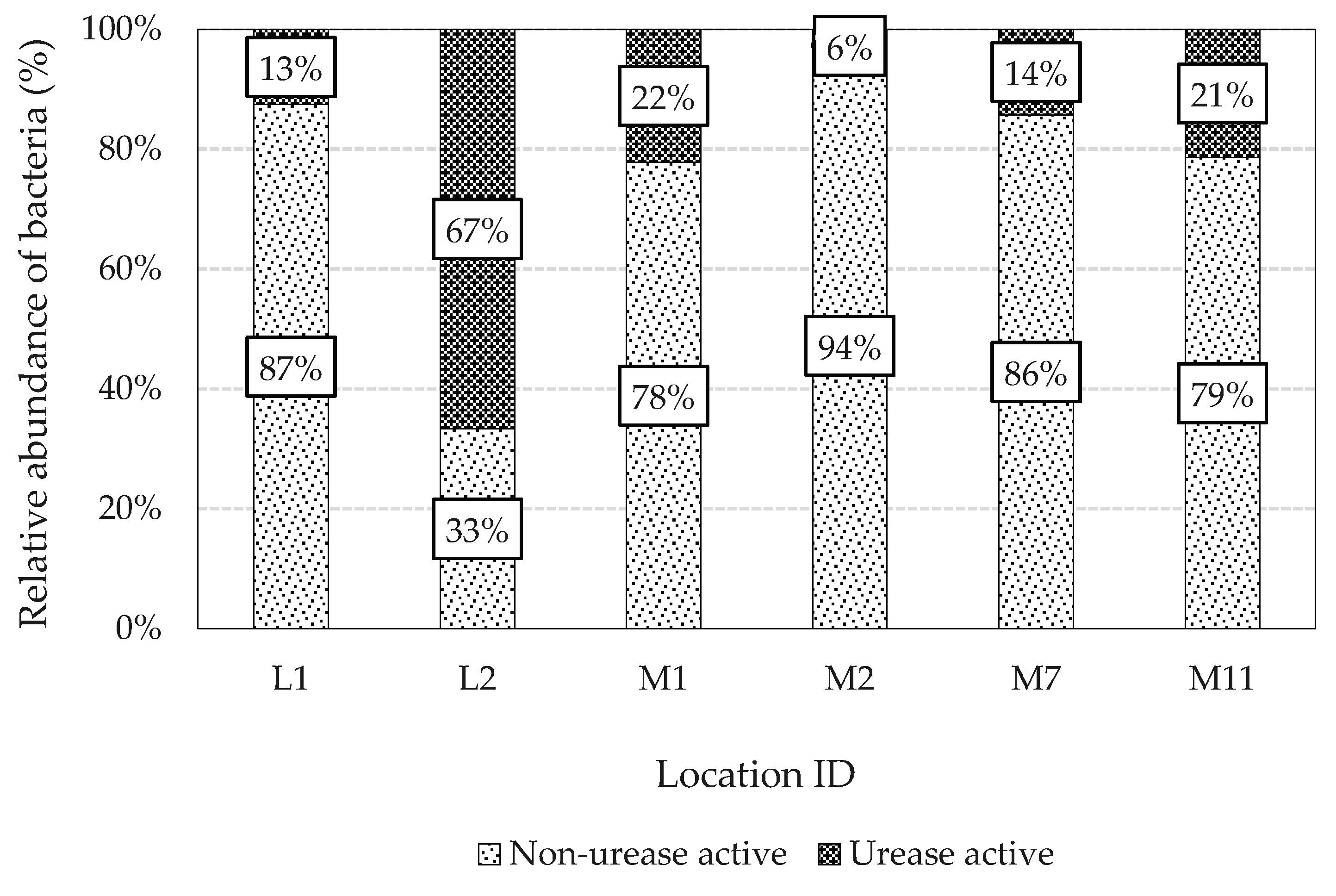
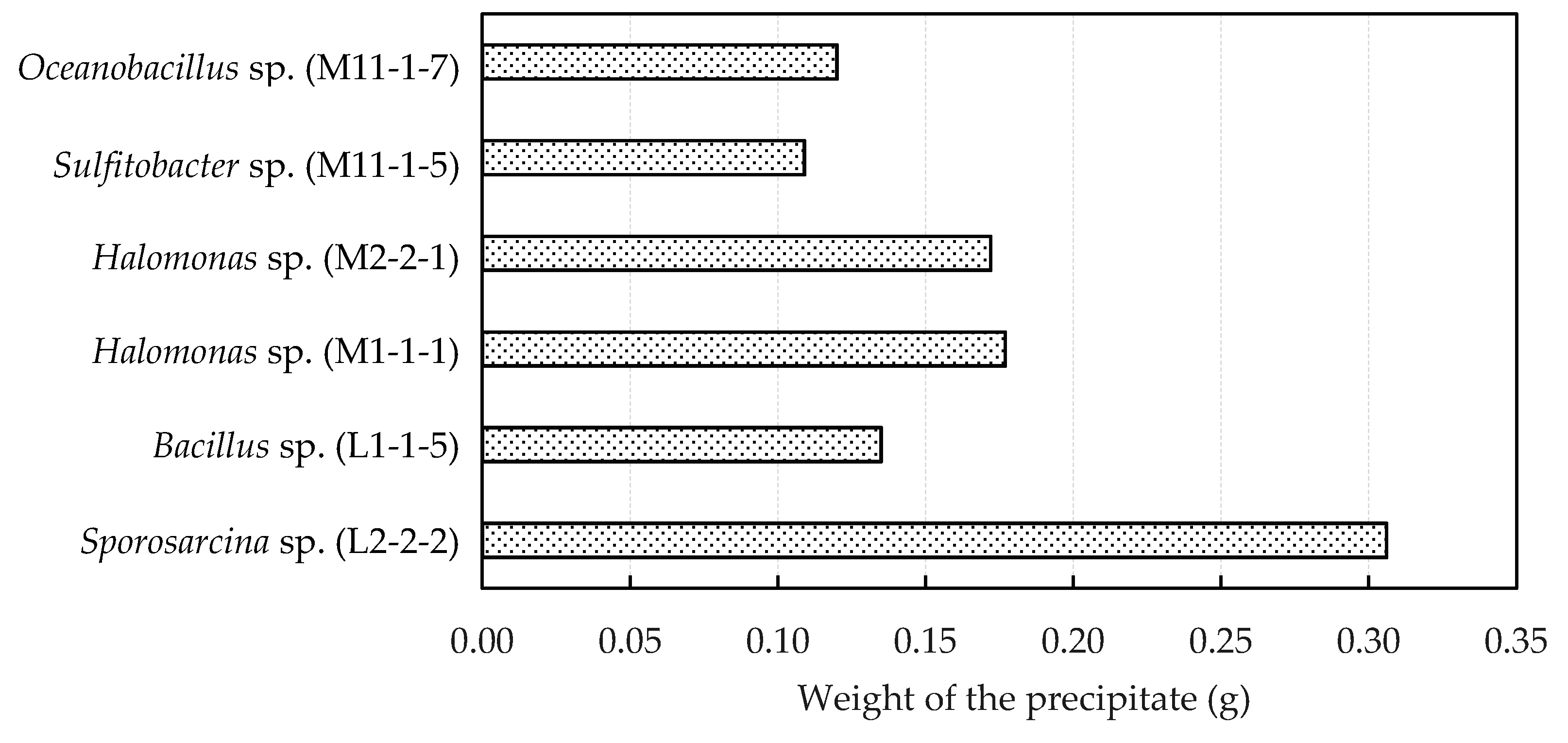
3.1. Isolation and Identification of a Suitable Urease Active Bacterium
3.2. Optimizing the Growth and Enzymatic Activity of the Isolated Sporosarcina sp. Strain
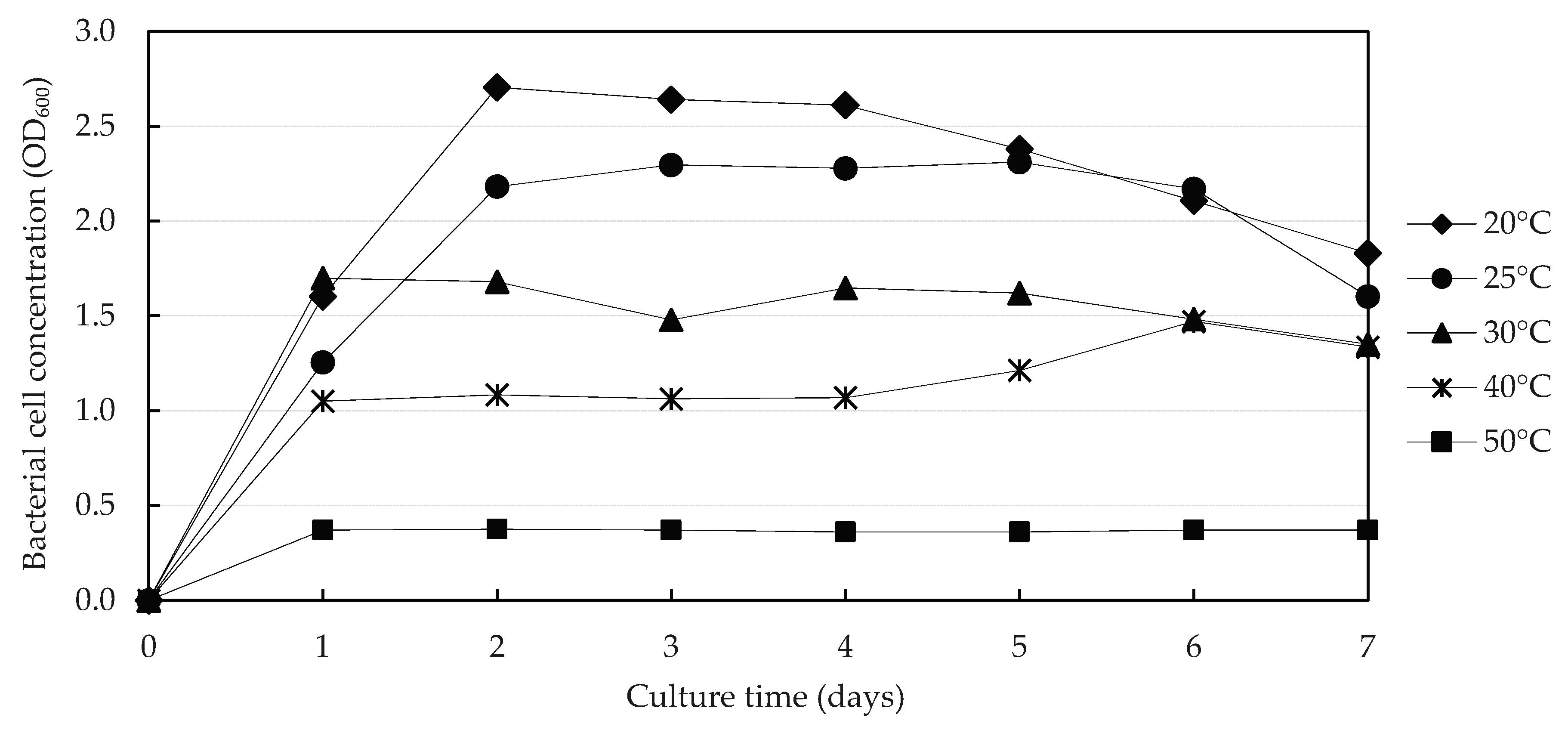
3.2.1. Effect of Temperature
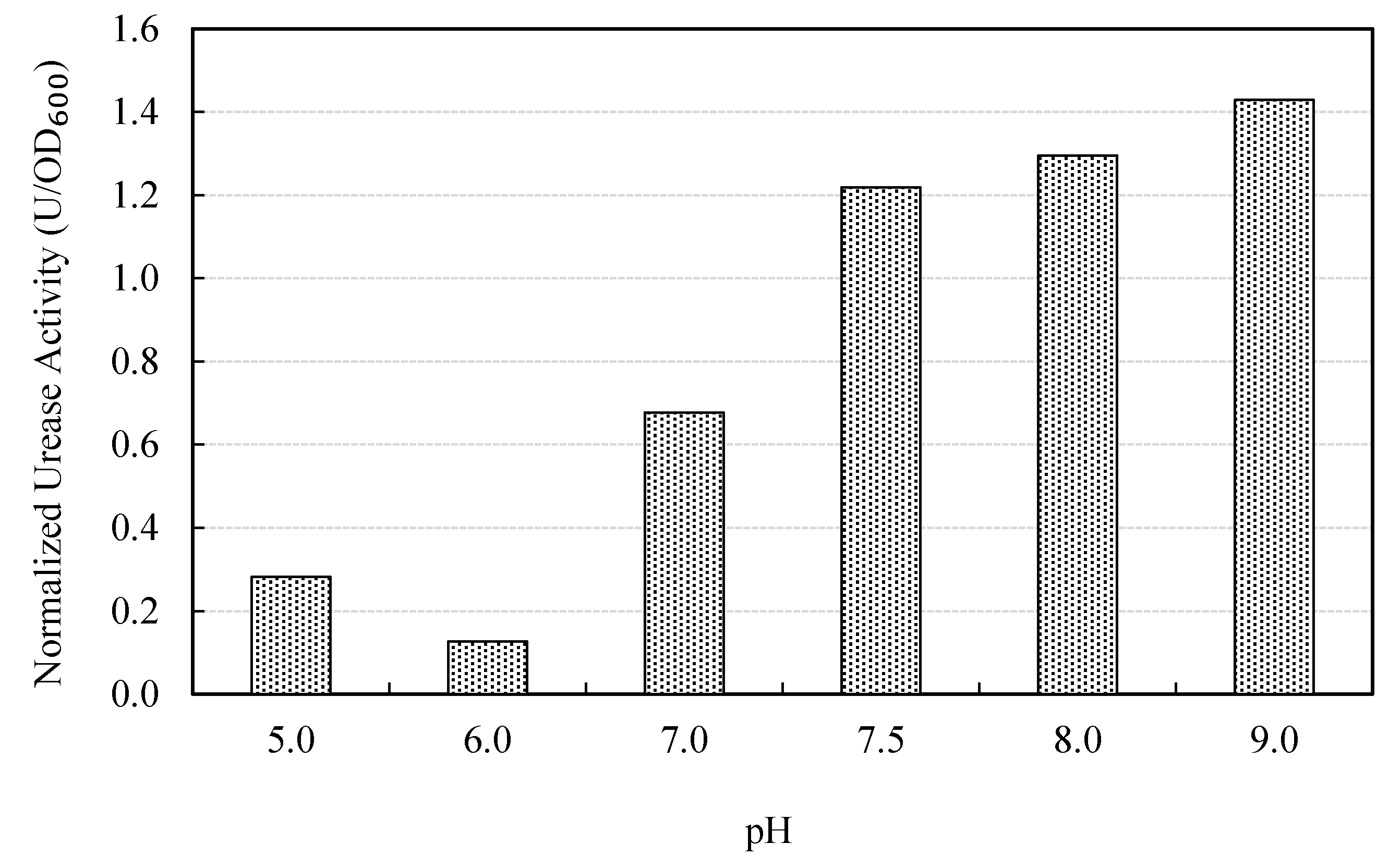
3.2.2. Effect of pH
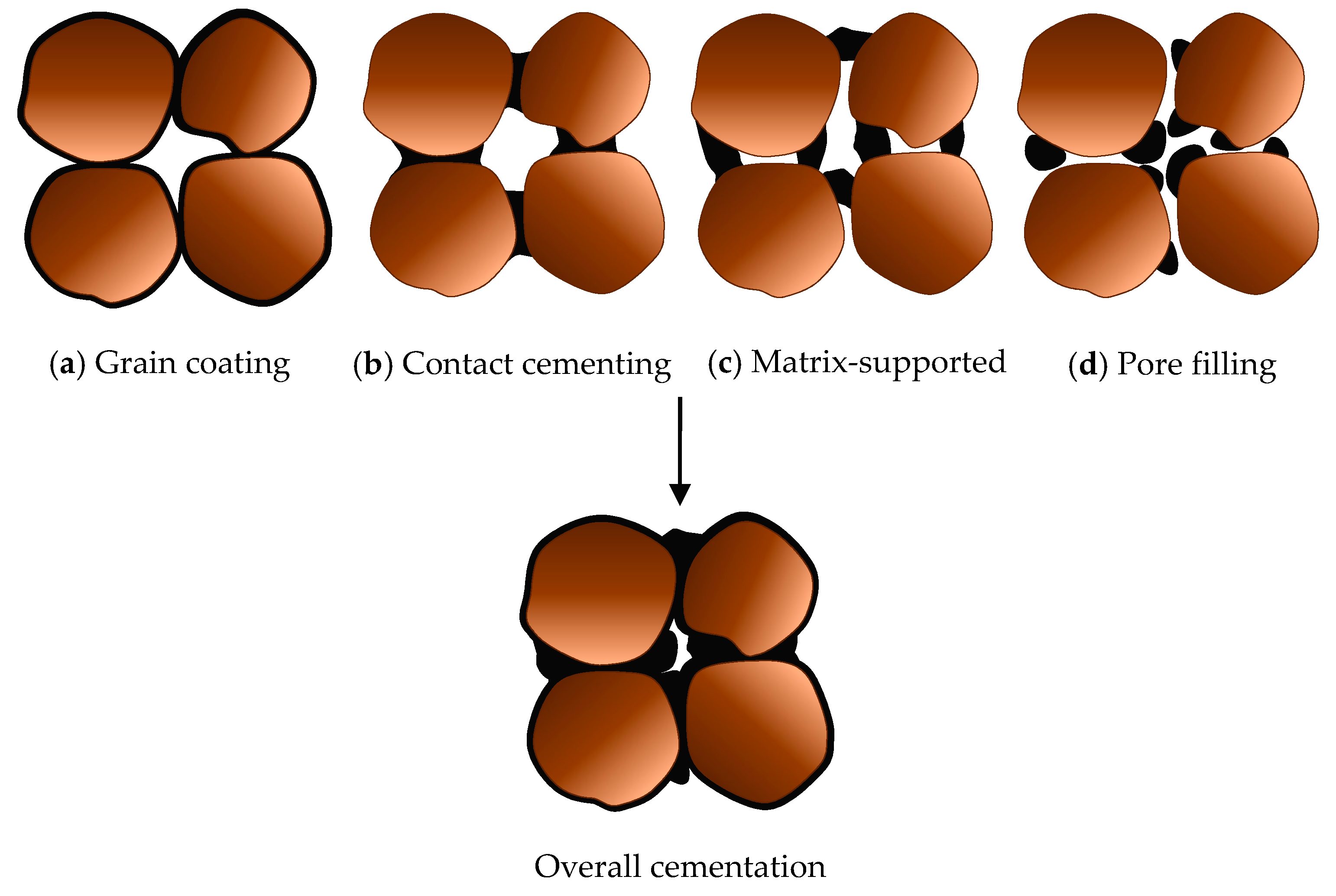
3.3. Potential for Beach Sand Solidification by MICP Treatment
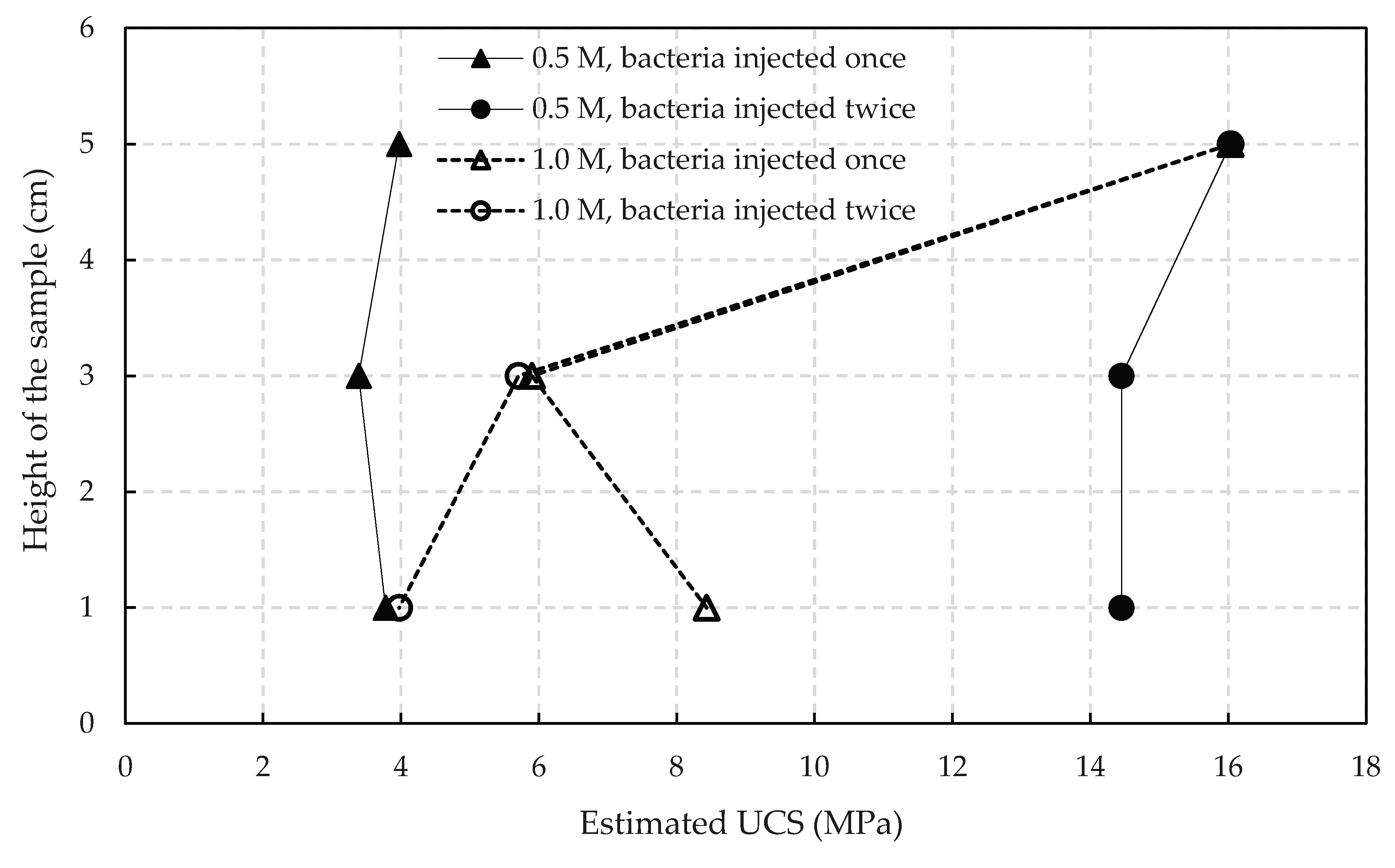
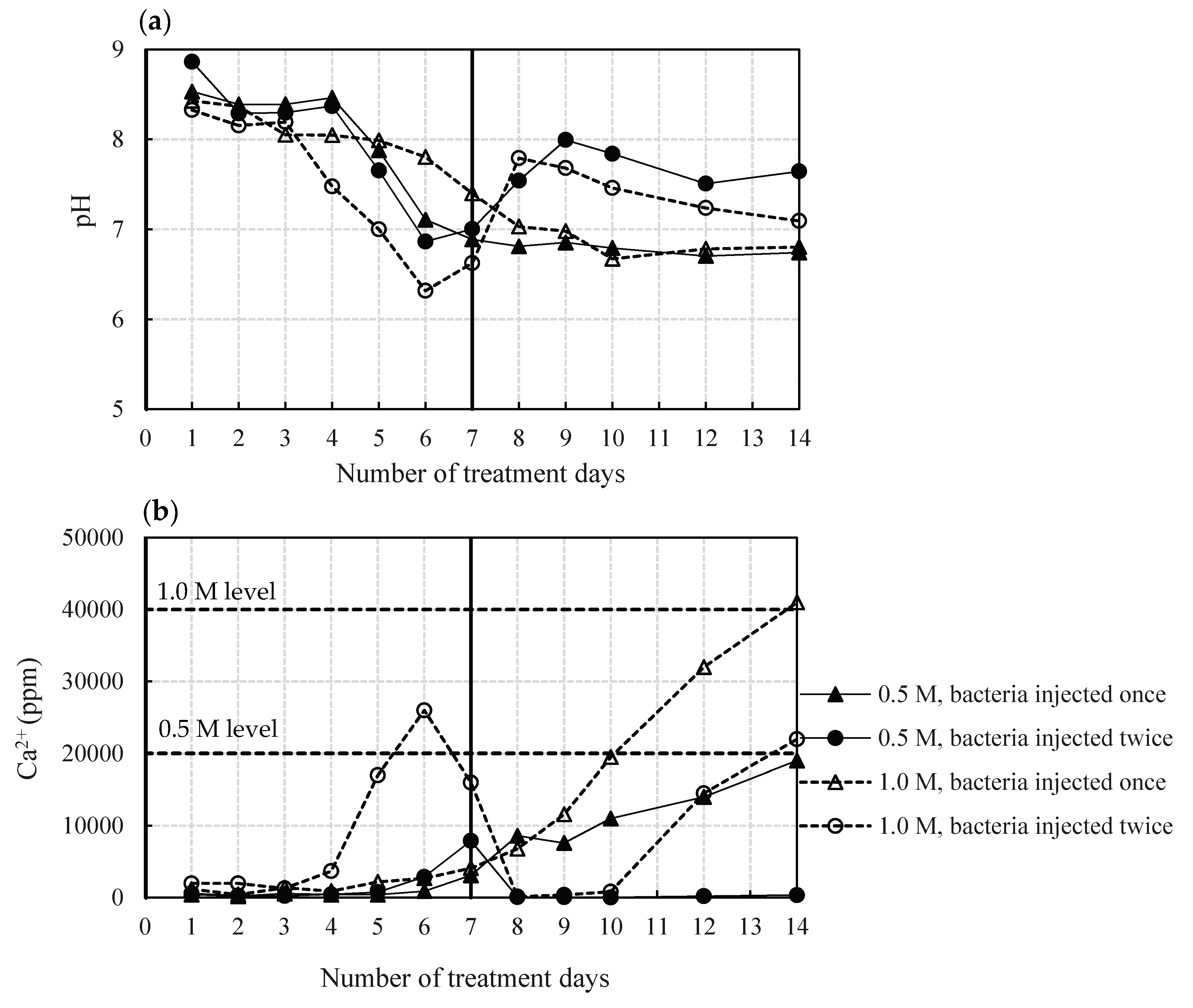
3.3.1. Effect of Bacteria Population and Reactant Concentration
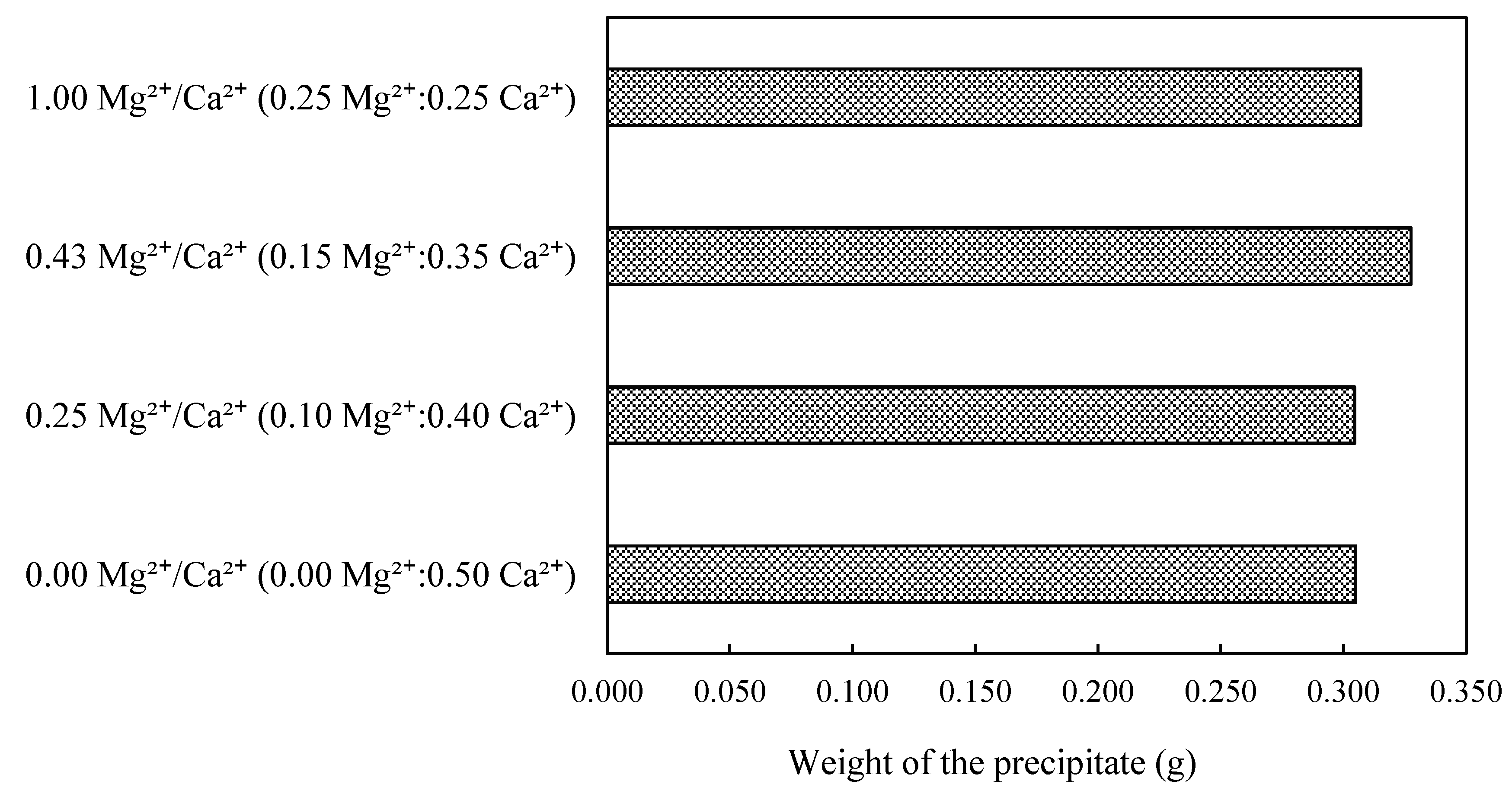
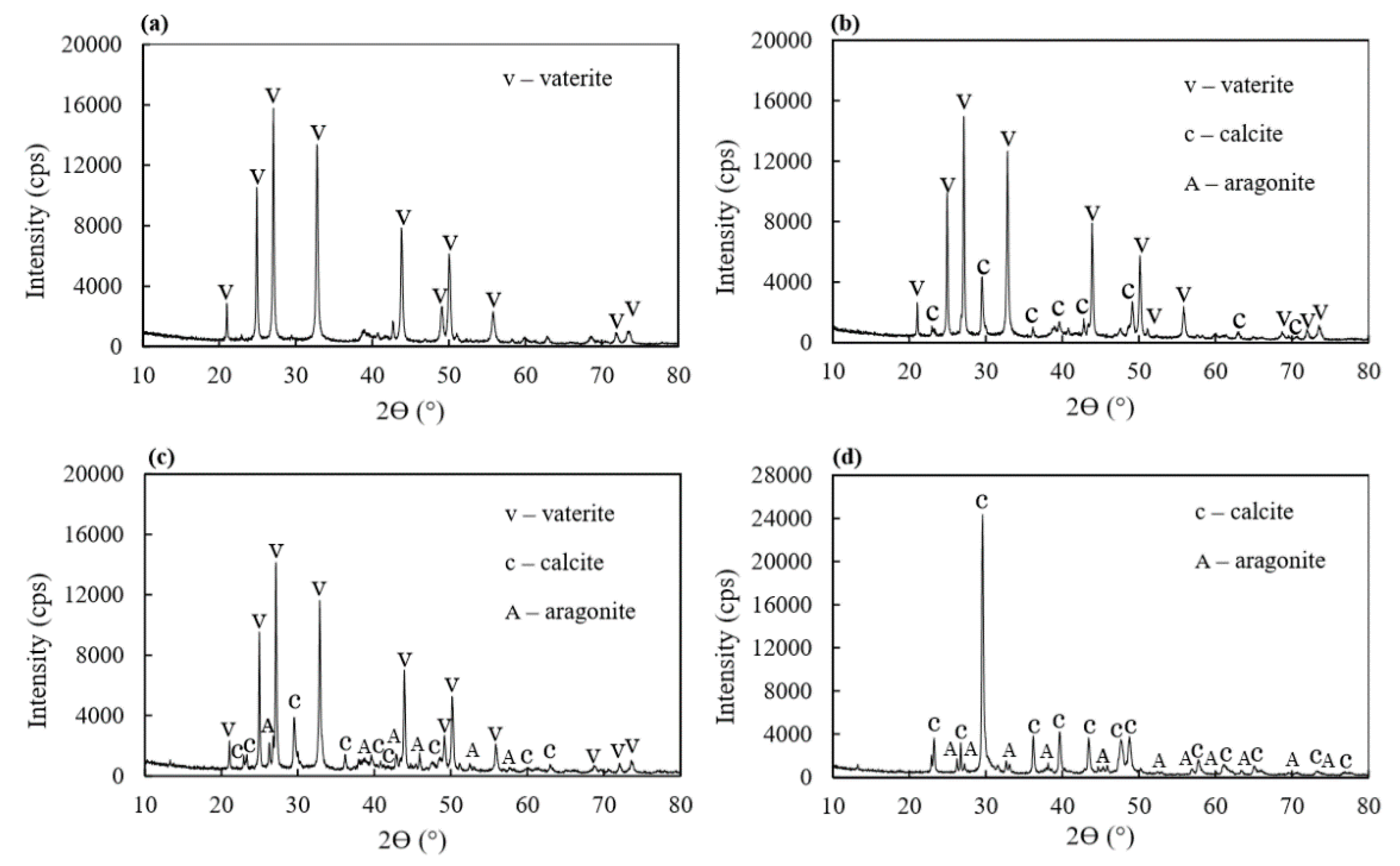
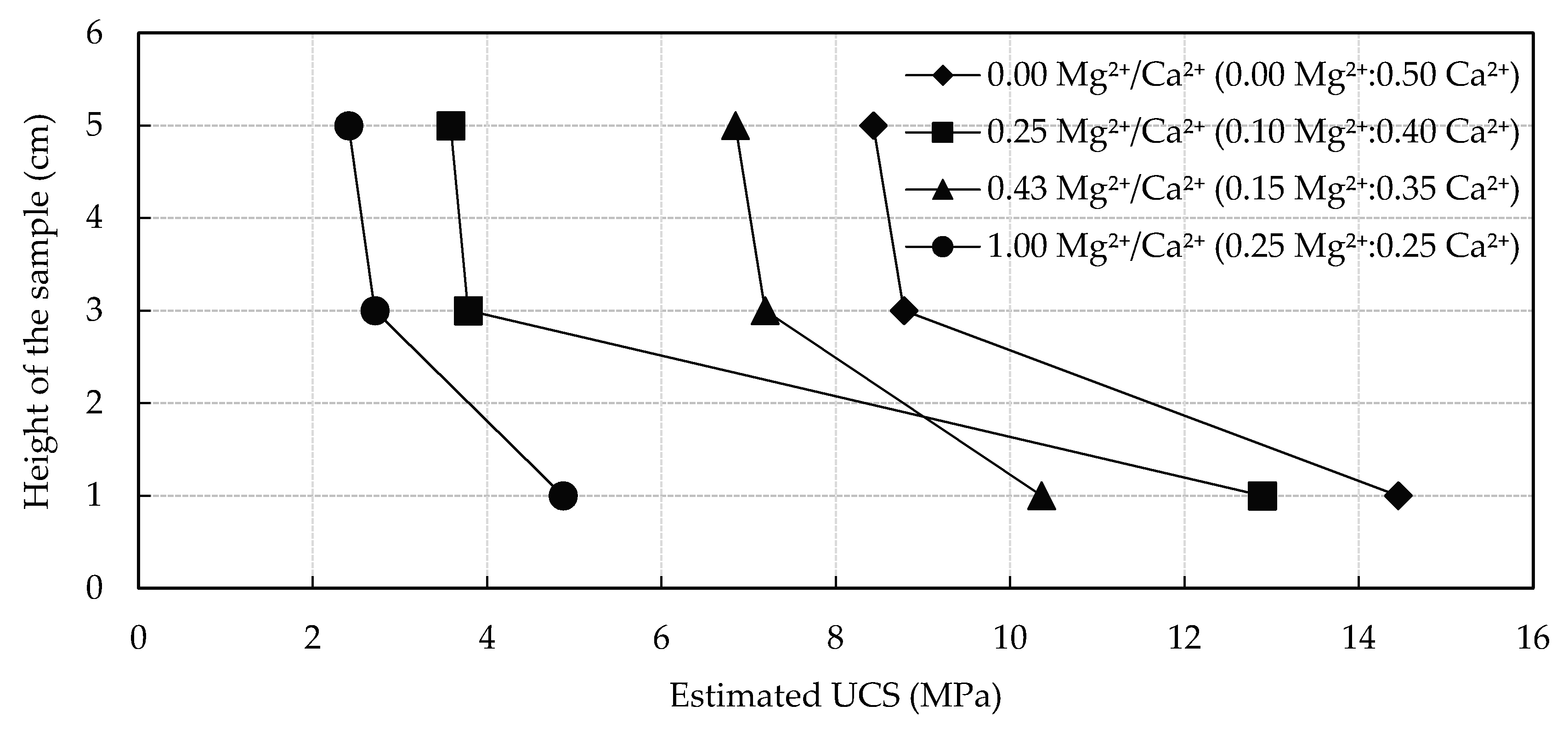
3.3.2. Effect of Mg2+ Addition on Microbial Induced Carbonate Precipitation
4. Conclusions and Implications for the Target Application
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- DeJong, J.T.; Mortensen, B.M.; Martinez, B.C.; Nelson, D.C. Bio-mediated soil improvement. Ecol. Eng. 2010, 36, 197–210. [Google Scholar] [CrossRef]
- Khan, M.N.H.; Shimazaki, S.; Kawasaki, S. Coral Sand Solidification Test Through Microbial Calcium Carbonate Precipitation Using Pararhodobacter sp. Int. J. GEOMATE 2016, 11, 2665–2670. [Google Scholar]
- Gowthaman, S.; Mitsuyama, S.; Nakashima, K.; Komatsu, M.; Kawasaki, S. Microbial Induced Slope Surface Stabilization Using Industrial-Grade Chemicals: A Preliminary Laboratory Study. Int. J. GEOMATE 2019, 17, 110–116. [Google Scholar] [CrossRef]
- Van Paassen, L.A.; Ghose, R.; van der Linden, T.J.M.; van der Star, W.R.L.; van Loosdrecht, M.C.M. Quantifying Biomediated Ground Improvement by Ureolysis: Large-Scale Biogrout Experiment. J. Geotech. Geoenviron. Eng. 2010, 136, 1721–1728. [Google Scholar] [CrossRef]
- Gomez, M.G.; Anderson, C.M.; Graddy, C.M.R.; DeJong, J.T.; Nelson, D.C.; Ginn, T.R. Large-Scale Comparison of Bioaugmentation and Biostimulation Approaches for Biocementation of Sands. J. Geotech. Geoenviron. Eng. 2016, 143, 04016124. [Google Scholar] [CrossRef]
- Canakci, H.; Sidik, W.; Halil Kilic, I. Effect of bacterial calcium carbonate precipitation on compressibility and shear strength of organic soil. Soils Found. 2015, 55, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Cord-Ruwisch, R. In situ soil cementation with ureolytic bacteria by surface percolation. Ecol. Eng. 2012, 42, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yao, J.; Yuan, Z.; Wang, T.; Zhang, Y.; Wang, F. Bioremediation of Cd by strain GZ-22 isolated from mine soil based on biosorption and microbially induced carbonate precipitation. Environ. Sci. Pollut. Res. 2017, 24, 372–380. [Google Scholar] [CrossRef]
- Wu, J.; Wang, X.B.; Wang, H.F.; Zeng, R.J. Microbially induced calcium carbonate precipitation driven by ureolysis to enhance oil recovery. RSC Adv. 2017, 7, 37382–37391. [Google Scholar] [CrossRef] [Green Version]
- Okyay, T.O.; Nguyen, H.N.; Castro, S.L.; Rodrigues, D.F. CO2 sequestration by ureolytic microbial consortia through microbially-induced calcite precipitation. Sci. Total Environ. 2016, 572, 671–680. [Google Scholar] [CrossRef]
- Mwandira, W.; Nakashima, K.; Kawasaki, S.; Ito, M.; Sato, T.; Igarashi, T.; Banda, K.; Chirwa, M.; Nyambe, I.; Nakayama, S.; et al. Efficacy of biocementation of lead mine waste from the Kabwe Mine site evaluated using Pararhodobacter sp. Environ. Sci. Pollut. Res. 2019, 26, 15653–15664. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.S.; Lee, M.-L.; Hii, S.-L. An Overview of the Factors Affecting Microbial-Induced Calcite Precipitation and its Potential Application in Soil Improvement. Int. J. Civ. Environ. Eng. 2012, 6, 188–194. [Google Scholar]
- Stocks-Fischer, S.; Galinat, J.K.; Bang, S.S. Microbiological precipitation of CaCO3. Soil Biol. Biochem. 1999, 31, 1563–1571. [Google Scholar] [CrossRef]
- Harkes, M.P.; van Paassen, L.A.; Booster, J.L.; Whiffin, V.S.; van Loosdrecht, M.C.M. Fixation and distribution of bacterial activity in sand to induce carbonate precipitation for ground reinforcement. Ecol. Eng. 2010, 36, 112–117. [Google Scholar] [CrossRef]
- Cortés, P.; Escrig, C.; Bernat-Maso, E.; Gil, L.; Barbé, J. Effect of Sporosarcina pasteurii on the strength properties of compressed earth specimens. Mater. Construcción 2018, 68, 143. [Google Scholar]
- Whiffin, V.S.; van Paassen, L.A.; Harkes, M.P. Microbial carbonate precipitation as a soil improvement technique. Geomicrobiol. J. 2007, 24, 417–423. [Google Scholar] [CrossRef]
- Temmerman, S.; Meire, P.; Bouma, T.J.; Herman, P.M.J.; Ysebaert, T.; De Vriend, H.J. Ecosystem-based coastal defence in the face of global change. Nature 2013, 504, 79–83. [Google Scholar] [CrossRef]
- Ten Voorde, M.; do Carmo, J.S.A.; Neves, M.G. Designing a Preliminary Multifunctional Artificial Reef to Protect the Portuguese Coast. J. Coast. Res. 2009, 251, 69–79. [Google Scholar] [CrossRef]
- Cheng, L.; Shahin, M.A.; Cord-Ruwisch, R. Bio-cementation of sandy soil using microbially induced carbonate precipitation for marine environments. Géotechnique 2014, 64, 1010–1013. [Google Scholar] [CrossRef]
- Salifu, E.; MacLachlan, E.; Iyer, K.R.; Knapp, C.W.; Tarantino, A. Application of microbially induced calcite precipitation in erosion mitigation and stabilisation of sandy soil foreshore slopes: A preliminary investigation. Eng. Geol. 2016, 201, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Ivanov, V.; Chenghong, G.; Naeimi, M.; Tkalich, P. Microbial geotechnical engineering for disaster mitigation and coastal management. Proc. Earthq. Tsunami 2009, 1–6. [Google Scholar]
- Shanahan, C.; Montoya, B.M. Strengthening Coastal Sand Dunes Using Microbial-Induced Calcite Precipitation; Geo-Congress 2014 Technical Papers; American Society of Civil Engineers (ASCE): Reston, VA, USA, 2014; pp. 1683–1692. [Google Scholar]
- Van der Ruyt, M.; van der Zon, W. Biological in situ reinforcement of sand in near-shore areas. Proc. Inst. Civ. Eng. Geotech. Eng. 2009, 162, 81–83. [Google Scholar] [CrossRef]
- Cheng, L.; Shahin, M.A.; Mujah, D. Influence of Key Environmental Conditions on Microbially Induced Cementation for Soil Stabilization. J. Geotech. Geoenviron. Eng. 2016, 143, 04016083-1–04016083-11. [Google Scholar] [CrossRef]
- Gat, D.; Tsesarsky, M.; Shamir, D.; Ronen, Z. Accelerated microbial-induced CaCO3 precipitation in a defined coculture of ureolytic and non-ureolytic bacteria. Biogeosciences 2014, 11, 2561–2569. [Google Scholar] [CrossRef]
- De Muynck, W.; De Belie, N.; Verstraete, W. Microbial carbonate precipitation in construction materials: A review. Ecol. Eng. 2010, 36, 118–136. [Google Scholar] [CrossRef]
- Lin, H.; Suleiman, M.T.; Brown, D.G.; Kavazanjian, E. Mechanical Behavior of Sands Treated by Microbially Induced Carbonate Precipitation. J. Geotech. Geoenviron. Eng. 2015, 142, 04015066. [Google Scholar] [CrossRef]
- Gat, D.; Ronen, Z.; Tsesarsky, M. Soil Bacteria Population Dynamics Following Stimulation for Ureolytic Microbial-Induced CaCO3 Precipitation. Environ. Sci. Technol. 2016, 50, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Tobler, D.J.; Cuthbert, M.O.; Greswell, R.B.; Riley, M.S.; Renshaw, J.C.; Handley-Sidhu, S.; Phoenix, V.R. Comparison of rates of ureolysis between Sporosarcina pasteurii and an indigenous groundwater community under conditions required to precipitate large volumes of calcite. Geochim. Cosmochim. Acta 2011, 75, 3290–3301. [Google Scholar] [CrossRef]
- Dhami, N.K.; Alsubhi, W.R.; Watkin, E.; Mukherjee, A. Bacterial community dynamics and biocement formation during stimulation and augmentation: Implications for soil consolidation. Front. Microbiol. 2017, 8, 1267. [Google Scholar] [CrossRef]
- Wenderoth, D.F.; Rosenbrock, P.; Abraham, W.R.; Pieper, D.H.; Höfle, M.G. Bacterial community dynamics during biostimulation and bioaugmentation experiments aiming at chlorobenzene degradation in groundwater. Microb. Ecol. 2003, 46, 161–176. [Google Scholar] [CrossRef]
- Nawarathna, T.H.K.; Nakashima, K.; Fujita, M.; Takatsu, M.; Kawasaki, S. Effects of Cationic Polypeptide on CaCO3 Crystallization and Sand Solidification by Microbial-Induced Carbonate Precipitation. ACS Sustain. Chem. Eng. 2018, 6, 10315–10322. [Google Scholar] [CrossRef]
- Graddy, C.M.R.; Gomez, M.G.; Kline, L.M.; Morrill, S.R.; Dejong, J.T.; Nelson, D.C. Diversity of Sporosarcina -like Bacterial Strains Obtained from Meter-Scale Augmented and Stimulated Biocementation Experiments. Environ. Sci. Technol. 2018, 52, 3997–4005. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhao, Q.; Zhi, D.; Wang, Z.; Wang, Y.; Xie, Q.; Wu, Z.; Wang, X.; Li, Y.; Yu, L.; et al. Sporosarcina terrae sp. Nov., isolated from orchard soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2104–2108. [Google Scholar] [CrossRef] [PubMed]
- Dick, J.; De Windt, W.; De Graef, B.; Saveyn, H.; Van Der Meeren, P.; De Belie, N.; Verstraete, W. Bio-deposition of a calcium carbonate layer on degraded limestone by Bacillus species. Biodegradation 2006, 17, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Achal, V.; Mukherjee, A.; Basu, P.C.; Reddy, M.S. Strain improvement of Sporosarcina pasteurii for enhanced urease and calcite production. J. Ind. Microbiol. Biotechnol. 2009, 36, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Hammad, I.A.; Talkhan, F.N.; Zoheir, A.E. Urease activity and induction of calcium carbonate precipitation by Sporosarcina pasteurii NCIMB 8841. J. Appl. Sci. Res. 2013, 9, 1525–1533. [Google Scholar]
- Sun, X.; Miao, L.; Wu, L.; Wang, C. Study of magnesium precipitation based on biocementation. Mar. Georesour. Geotechnol. 2019. [Google Scholar] [CrossRef]
- Kang, C.H.; Choi, J.H.; Noh, J.G.; Kwak, D.Y.; Han, S.H.; So, J.S. Microbially Induced Calcite Precipitation-based Sequestration of Strontium by Sporosarcina pasteurii WJ-2. Appl. Biochem. Biotechnol. 2014, 174, 2482–2491. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.S.; Galinat, J.K.; Ramakrishnan, V. Calcite precipitation induced by polyurethane-immobilized Bacillus pasteurii. Enzyme Microb. Technol. 2001, 28, 404–409. [Google Scholar] [CrossRef]
- Okwadha, G.D.O.; Li, J. Optimum conditions for microbial carbonate precipitation. Chemosphere 2010, 81, 1143–1148. [Google Scholar] [CrossRef]
- Fujita, Y.; Grant Ferris, F.; Daniel Lawson, R.; Colwell, F.S.; Smith, R.W. Calcium carbonate precipitation by ureolytic subsurface bacteria. Geomicrobiol. J. 2000, 17, 305–318. [Google Scholar] [CrossRef]
- Anbu, P.; Kang, C.H.; Shin, Y.J.; So, J.S. Formations of calcium carbonate minerals by bacteria and its multiple applications. Springerplus 2016, 5, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebata-Landa, V. Microbial Activity in Sediments: Effects on Soil Behavior. Ph.D. Thesis, Georgia Institute of Technology, Atlanta, GA, USA, 2007. [Google Scholar]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Synergistic role of bacterial urease and carbonic anhydrase in carbonate mineralization. Appl. Biochem. Biotechnol. 2014, 172, 2552–2561. [Google Scholar] [CrossRef] [PubMed]
- Sahrawat, K.L. Effects of temperature and moisture on urease activity in semi-arid tropical soils. Plant Soil 1984, 8, 401–408. [Google Scholar] [CrossRef]
- Liang, Z.P.; Feng, Y.Q.; Meng, S.X.; Liang, Z.Y. Preparation and properties of urease immobilized onto Glutaraldehyde cross-linked Chitosan beads. Chin. Chem. Lett. 2005, 16, 135–138. [Google Scholar]
- Chen, Y.Y.M.; Anne Clancy, K.; Burne, R.A. Streptococcus salivarius urease: Genetic and biochemical characterization and expression in a dental plaque streptococcus. Infect. Immun. 1996, 64, 585–592. [Google Scholar]
- Fujita, M.; Nakashima, K.; Achal, V.; Kawasaki, S. Whole-cell evaluation of urease activity of Pararhodobacter sp. isolated from peripheral beachrock. Biochem. Eng. J. 2017, 124, 1–5. [Google Scholar] [CrossRef]
- Van Paassen, L.A. Biogrout: Ground Improvement by Microbially Induced Carbonate Precipitation. Ph.D. Thesis, Delft University of Technology, Delft, The Netherlands, 2009. [Google Scholar]
- Mobley, H.L.T.; Island, M.D.; Hausinger, R.P. Molecular biology of microbial ureases. Microbiol. Rev. 1995, 59, 451–480. [Google Scholar] [PubMed]
- Ciurli, S.; Marzadori, C.; Benini, S.; Deiana, S.; Gessa, C. Urease from the soil bacterium Bacillus pasteurii: Immobilization on Ca- polygalacturonate. Soil Biol. Biochem. 1996, 28, 811–817. [Google Scholar] [CrossRef]
- Mahadevan, S.; Sauer, F.D.; Erfle, J.D. Purification and properties of urease from bovine rumen. Biochem. J. 2015, 163, 495–501. [Google Scholar] [CrossRef]
- Ng, W.S.; Lee, L.M.; Khun, T.C.; Ling, H.S. Factors Affecting Improvement in Engineering Properties of Residual Soil through Microbial-Induced Calcite Precipitation. J. Geotech. Geoenviron. Eng. 2014, 140, 04014006. [Google Scholar]
- Zhu, T.; Dittrich, M. Carbonate Precipitation through Microbial Activities in Natural Environment, and Their Potential in Biotechnology: A Review. Front. Bioeng. Biotechnol. 2016, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Al Qabany, A.; Soga, K.; Santamarina, C. Factors Affecting Efficiency of Microbially Induced Calcite Precipitation. J. Geotech. Geoenviron. Eng. 2011, 138, 992–1001. [Google Scholar] [CrossRef]
- Li, W.; Liu, L.P.; Zhou, P.P.; Cao, L.; Yu, L.J.; Jiang, S.Y. Calcite precipitation induced by bacteria and bacterially produced carbonic anhydrase. Curr. Sci. 2011, 100, 502–508. [Google Scholar]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Significant indicators for biomineralisation in sand of varying grain sizes. Constr. Build. Mater. 2016, 104, 198–207. [Google Scholar] [CrossRef]
- Gorospe, C.M.; Han, S.H.; Kim, S.G.; Park, J.Y.; Kang, C.H.; Jeong, J.H.; So, J.S. Effects of different calcium salts on calcium carbonate crystal formation by Sporosarcina pasteurii KCTC 3558. Biotechnol. Bioprocess Eng. 2013, 18, 903–908. [Google Scholar] [CrossRef]
- Gandhi, K.S.; Kumar, R.; Ramkrishna, D. Some Basic Aspects of Reaction Engineering of Precipitation Processes. Ind. Eng. Chem. Res. 1995, 34, 3223–3230. [Google Scholar] [CrossRef]
- Somani, R.S.; Patel, K.S.; Mehta, A.R.; Jasra, R.V. Examination of the polymorphs and particle size of calcium carbonate precipitated using still effluent (i.e., CaCl2 + NaCl solution) of soda ash manufacturing process. Ind. Eng. Chem. Res. 2006, 45, 5223–5230. [Google Scholar] [CrossRef]
- Mujah, D.; Shahin, M.A.; Cheng, L. State-of-the-Art Review of Biocementation by Microbially Induced Calcite Precipitation (MICP) for Soil Stabilization. Geomicrobiol. J. 2017, 34, 524–537. [Google Scholar] [CrossRef]
- Chu, J.; Ivanov, V.; Naeimi, M.; Stabnikov, V.; Liu, H.L. Optimization of calcium-based bioclogging and biocementation of sand. Acta Geotech. 2014, 9, 277–285. [Google Scholar] [CrossRef]
- Mortensen, B.M.; Haber, M.J.; DeJong, J.T.; Caslake, L.F.; Nelson, D.C. Effects of environmental factors on microbial induced calcium carbonate precipitation. J. Appl. Microbiol. 2011, 111, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Fukue, M.; Ono, S.-I.; Sato, Y. Cementation of Sands Due To Microbiologically-Induced Carbonate Precipitation. Soils Found. 2011, 51, 83–93. [Google Scholar] [CrossRef]
- Spanos, N.; Koutsoukos, P.G. The transformation of vaterite to calcite: Effect of the conditions of the solutions in contact with the mineral phase. J. Cryst. Growth 1998, 191, 783–790. [Google Scholar] [CrossRef]
- Bains, A.; Dhami, N.K.; Mukherjee, A.; Reddy, M.S. Influence of Exopolymeric Materials on Bacterially Induced Mineralization of Carbonates. Appl. Biochem. Biotechnol. 2015, 175, 3531–3541. [Google Scholar] [CrossRef] [PubMed]
- Gowthaman, S.; Mitsuyama, S.; Nakashima, K.; Komatsu, M.; Kawasaki, S. Biogeotechnical approach for slope soil stabilization using locally isolated bacteria and inexpensive low-grade chemicals: A feasibility study on Hokkaido expressway soil, Japan. Soils Found. 2019, 59, 484–499. [Google Scholar] [CrossRef]
- Nawarathna, T.H.K.; Nakashima, K.; Kawasaki, S. Chitosan enhances calcium carbonate precipitation and solidification mediated by bacteria. Int. J. Biol. Macromol. 2019, 133, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Putra, H.; Kinoshita, N.; Yasuhara, H.; Lu, C.-W.; Neupane, D. Effect of Magnesium as Substitute Material in Enzyme-Mediated Calcite Precipitation for Soil-Improvement Technique. Front. Bioeng. Biotechnol. 2016, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Mejri, W.; Korchef, A.; Tlili, M.; Ben Amor, M. Effects of temperature on precipitation kinetics and microstructure of calcium carbonate in the presence of magnesium and sulphate ions. Desalin. Water Treat. 2014, 52, 4863–4870. [Google Scholar] [CrossRef]
- Oomori, T.; Kitano, Y. The effect of Magnesium ions on the polymorphic crystallization of calcium carbonate. Bull. Coll. Sci. Univ. Ryukyus 1985, 39, 57–62. [Google Scholar]
- Gawwad, H.A.A.; Mohamed, S.A.E.A.; Mohammed, S.A. Impact of magnesium chloride on the mechanical properties of innovative bio-mortar. Mater. Lett. 2016, 178, 39–43. [Google Scholar] [CrossRef]
- Putra, H.; Yasuhara, H.; Kinoshita, N.; Hirata, A. Optimization of Enzyme-Mediated Calcite Precipitation as a Soil-Improvement Technique: The Effect of Aragonite and Gypsum on the Mechanical Properties of Treated Sand. Crystals 2017, 7, 59. [Google Scholar] [CrossRef]
- Sun, X.; Miao, L. The Comparison of Microbiologically-Induced Calcium Carbonate Precipitation and Magnesium Carbonate Precipitation. In Proceedings of the 8th International Congress on Environmental Geotechnics; Springer Nature Singapore Pte Ltd.: Singapore, 2019; Volume 3, pp. 302–308. [Google Scholar]
- Rong, H.; Qian, C.X.; Li, L.Z. Influence of Magnesium Additive on Mechanical Properties of Microbe Cementitious Materials. Mater. Sci. Forum 2013, 743–744, 275–279. [Google Scholar] [CrossRef]
- Mahawish, A.; Bouazza, A.; Gates, W.P. Effect of particle size distribution on the bio-cementation of coarse aggregates. Acta Geotech. 2018, 13, 1019–1025. [Google Scholar] [CrossRef]
- Sharma, A.; Ramkrishnan, R. Study on effect of Microbial Induced Calcite Precipitates on strength of fine grained soils. Perspect. Sci. 2016, 8, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Wang, Y. Investigating the Factors Affecting the Properties of Coral Sand Treated with Microbially Induced Calcite Precipitation. Adv. Civ. Eng. 2018, 2018, 1–6. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sand Type | ρ | D50 | Cu | Cc | Elemental Composition (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Al2O3 | SiO2 | CaO | TiO2 | Cr2O3 | Fe2O3 | NiO | CuO | SrO | PbO | |||||
| Sand 1 | 2.81 | 0.37 | 3.58 | 1.12 | 3.64 | 75.39 | 4.25 | 8.39 | 0.04 | 8.13 | 0.08 | 0.07 | - | - |
| Sand 2 | 2.68 | 0.61 | 1.51 | 0.93 | - | 24.14 | 71.79 | 0.27 | - | 2.98 | 0.04 | - | 0.61 | 0.06 |
| Test Case | Mg2+ (M) | Ca2+ (M) | Mg2+/Ca2+ Ratio | Sum of Mg2+ and Ca2+ (M) | Urea (M) |
|---|---|---|---|---|---|
| 1 | 0.00 | 0.50 | 0.00 | 0.50 | 0.50 |
| 2 | 0.10 | 0.40 | 0.25 | 0.50 | 0.50 |
| 3 | 0.15 | 0.35 | 0.43 | 0.50 | 0.50 |
| 4 | 0.25 | 0.25 | 1.00 | 0.50 | 0.50 |
| Sample ID | Closest Match to Phylogenetic Grouping | Genbank Accession Number | Similarity |
|---|---|---|---|
| M1-1-1 | Halomonas sp. | AJ306891 | 99.8% |
| M2-2-1 | Halomonas sp. | AJ306888 | 99.6% |
| M11-1-5 | Sulfitobacter sp. | AY180103 | 98.0% |
| M11-1-7 | Oceanobacillus sp. | HQ595230 | 99.9% |
| L2-2-2 | Sporosarcina sp. | HQ676600 | 99.7% |
| L1-1-5 | Bacillus sp. | NR_146034 | 98.7% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nayanthara, P.G.N.; Dassanayake, A.B.N.; Nakashima, K.; Kawasaki, S. Microbial Induced Carbonate Precipitation Using a Native Inland Bacterium for Beach Sand Stabilization in Nearshore Areas. Appl. Sci. 2019, 9, 3201. https://0-doi-org.brum.beds.ac.uk/10.3390/app9153201
Nayanthara PGN, Dassanayake ABN, Nakashima K, Kawasaki S. Microbial Induced Carbonate Precipitation Using a Native Inland Bacterium for Beach Sand Stabilization in Nearshore Areas. Applied Sciences. 2019; 9(15):3201. https://0-doi-org.brum.beds.ac.uk/10.3390/app9153201
Chicago/Turabian StyleNayanthara, Pahala Ge Nishadi, Anjula Buddhika Nayomi Dassanayake, Kazunori Nakashima, and Satoru Kawasaki. 2019. "Microbial Induced Carbonate Precipitation Using a Native Inland Bacterium for Beach Sand Stabilization in Nearshore Areas" Applied Sciences 9, no. 15: 3201. https://0-doi-org.brum.beds.ac.uk/10.3390/app9153201