Biodegradation of Picolinic Acid by Rhodococcus sp. PA18
by

Yanting Zhang
1, 
Junbin Ji
1,
Siqiong Xu
1,
Hongmei Wang
2,
Biao Shen
3,
Jian He
1,
Jiguo Qiu
1,* and
Qing Chen
2,3,* 1
College of Life Sciences, Nanjing Agricultural University, Nanjing 210095, China
2
College of Life Sciences, Zaozhuang University, Zaozhuang 277100, China
3
College of Environmental and Resource Sciences, Nanjing Agricultural University, Nanjing 210095, China
*
Authors to whom correspondence should be addressed.
Appl. Sci. 2019, 9(5), 1006; https://0-doi-org.brum.beds.ac.uk/10.3390/app9051006
Submission received: 7 February 2019
/
Revised: 4 March 2019
/
Accepted: 6 March 2019
/
Published: 11 March 2019
(This article belongs to the Special Issue Environmental Fate of Emerging Organic Micro-Contaminants)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Picolinic acid (PA), a C2-carboxylated pyridine derivative, is a significant intermediate used in industrial production. PA is considered hazardous for the environment and human health. In this study, a Gram-positive bacterium, Rhodococcus sp. PA18, which aerobically utilizes PA as a source of carbon and energy, was isolated. The strain completely degraded 100 mg/L PA within 24 h after induction and formed 6-hydroxypicolinic acid (6HPA), a major PA metabolite, which was identified using ultraviolet-visible spectroscopy, high performance liquid chromatography, and liquid chromatography/time of flight-mass spectrometry analyses. The cell-free extracts converted the PA into 6HPA when phenazine methosulfate was used as an electron acceptor. To our knowledge, this is the first report showing that PA can be metabolized by Rhodococcus. In conclusion, Rhodococcus sp. PA18 may be potentially used for the bioremediation of environments polluted with PA.
1. Introduction
Picolinic acid (PA), a pyridine derivative [1], has emerged as an important intermediate from the industrial syntheses of agricultural chemicals, drugs, dyestuffs, dyes, textiles, and mining [1,2,3,4]. It is also a dead-end product of L-tryptophan biosynthesis in living organisms [5,6,7]. In many common biological processes, PA is produced from the biodegradation of nitrobenzene, catechol, and anthranilic acid [8,9]. Owing to its hydrophilic nature (water solubility of 887 g/L at 20 °C), PA is easily transported to the aquatic environment and to soil [1,10]. Reports show that the pyridine concentration in wastewaters ranges from 20 to 300 mg/L [4]. Nevertheless, the toxicity or disadvantages of the use of PA have also been found. Owing to their relationship with environmental health, the removal of PA from contaminated ecosystems is considered essential for controlling environmental damage. Both physical and chemical methods are too expensive and ineffective in waste disposal management. Biological methods are efficient and cost effective, and hence, play an important role in the clean-up of toxic and hazardous wastes in the contaminated environment. Studies show that PA can be degraded efficiently by microorganisms, and many PA-utilizing bacterial strains, such as Achromobacter sp. JS18 [11], Alcaligenes faecalis JQ135 [12], Arthrobacter picolinophilus DSM 20665 [13], Burkholderia sp. ZD1 [3], and Streptomyces sp. Z2 [14], have been isolated and characterized.
An oxidative attack on the N-heterocyclic aromatic ring of PA is the main step initiating its bacterial degradation [1,3]. However, all of the above strains are Gram-negative bacteria, and whether Gram-positive bacteria can also degrade PA is not known. In this study, a Gram-positive PA-assimilating strain, Rhodococcus sp. PA18, was isolated, identified, and characterized. The aerobic degradation of PA by strain PA18 and the formation of 6-hydroxypicolinic acid (6HPA), the initial product formed after the oxidative attack on the N-heterocyclic aromatic ring, have been discussed.
To the best of our knowledge, this is the first report on the degradation of PA by a Gram-positive bacterium, Rhodococcus sp. PA18. This may be developed as a potentially low cost and environmentally-friendly approach to restore the environments contaminated by PA.
2. Materials and Methods
2.1. Chemicals and Media
PA and 6HPA were acquired from J&K Scientific Ltd. (Shanghai, China). Phenazine methosulfate (PMS), methylene blue (MB), and 2,6-dichloroindophenol (DCIP) were purchased from Sangon Biotech Co., Ltd. (Shanghai, China). All of the other chemicals and solvents used in this study were available commercially.
The Luria Bertani (LB) medium contained (per liter) tryptone (10.0 g), yeast extract (5.0 g), and NaCl (10.0 g). The mineral salts medium (MSM) contained (per liter) (NH4)2SO4 (1.0 g), NaCl (1.0 g), K2HPO4 (1.5 g), KH2PO4 (0.5 g), MgSO4 (0.2 g), and 1 mL trace elements medium stock solution (0.13 g MnSO4·H2O, 0.23 g ZnCl2, 0.03 g CuSO4·H2O, 0.42 g CoCl2·6H2O, 0.15 g Na2MoO4·2H2O, and 0.05 g AlCl3·6H2O). The media were adjusted to pH 7.0 using HCl or NaOH, and were autoclaved at 121 °C for 30 min. For the solid media, 1.5% agar (w/v) was added.
2.2. Isolation and Identification of PA-Degrading Bacteria
Soil samples were collected from a farmland in Nanjing City, China. The enrichment culture was initiated with 5.0 g of soil samples in 100 mL MSM, containing 500 mg/L PA as the sole carbon source. It was incubated under aerobic conditions in a rotary shaker at 30 °C and 150 rpm for one week, and then 5% (v/v) of the culture was transferred to fresh MSM containing PA. The process was repeated four times. The culture was diluted and spread on MSM agar plates containing PA (500 mg/L). The colonies were tested for their PA degradation capability. A strain named PA18, which showed a high PA degrading efficiency, was selected for further studies. The morphological and physiological characteristics of PA18 were characterized, and its 16S rDNA sequence was analyzed according to the method described by Nie et al. [15].
2.3. Biodegradation of PA by a Rhodococcus Strain
The strain Rhodococcus sp. PA18 was cultured in 100 mL of liquid LB medium for 24 h, until the late log phase. The cells were harvested via centrifugation at 5000 rpm for 10 min, washed twice with fresh MSM, and then transferred into MSM containing 100 mg/L PA. After seven days, the Rhodococcus cells were harvested, washed twice with fresh MSM, and then resuspended in fresh MSM. The optical density of the cells at 600 nm (OD600) was adjusted to 2.0. These PA-pre-cultured cells were used for further biodegradation assays. The ability of PA18 to degrade and utilize PA was assessed in 250 mL-conical flasks, with 100 mL of MSM supplemented with a constant initial concentration of PA (100 mg/L), and the cells were incubated in a rotary shaker at 30 °C and 150 rpm. The experiment was repeated in triplicate and the control experiment was set up as above, but without the inoculation of the bacteria. The samples were collected periodically from the cultures. The growth was monitored by measuring the OD600, and the residual PA concentration was measured using HPLC, as described below.
2.4. Effects of Different Factors on PA Biodegradation
In order to determine the optimal conditions for PA biodegradation, different inoculum sizes and initial PA concentrations were tested. Inoculum sizes of 0.005, 0.200, 0.500, and 1.000 at OD600 were used when the initial PA concentration was 100 mg/L. Conversely, the initial concentration of PA varied from 50, 100, 200, 300, 400, to 500 mg/L when the inoculum size was maintained at 0.005 at OD600. All of the above experiments were conducted in 250 mL flasks containing 100 mL MSM in a rotary shaker at 30 °C and 150 rpm. Each experiment was conducted in triplicate, and the residual PA concentration was measured by HPLC, as described below.
2.5. Identification of Metabolites during PA Degradation
To identify the metabolites during PA degradation, the strain PA18 was inoculated in a MSM medium with PA (100 mg/L), and was incubated at 30 °C and 150 rpm. The samples were collected at appropriate intervals for further analysis using UV-VIS spectroscopy, high performance liquid chromatography (HPLC), and liquid chromatography/time of flight-mass spectrometry (LC/TOF-MS).
2.6. Cell-Free Extract Activity Assays
The PA-pre-cultured PA18 cells were grown to the mid-log phase in MSM with 100 mg/L PA. After centrifugation (4°C, 12,000 rpm, 10 min), the harvested cells were resuspended in a 50 mM phosphate buffer (pH 7.0). The cell-free extract was prepared by sonication in an ice-water bath for 30 min (1 s with 2 s intervals). After sonication, the supernatant liquid was separated from the cell debris by centrifugation at 12,000 rpm for 30 min. The supernatant was collected for enzyme assays. The protein concentrations were measured using the Bradford method [16,17]. The picolinic acid hydroxylase activity of the cell-free extract was determined by measuring the increase in the absorbance of the reaction mixture at 310 nm, the absorption maximum of the product 6HPA (ε = 4.45 cm−1 mM−1). The reaction mixture contained 0.1 mM PA and a 0.1 mM electron acceptor in 500 μL of cell extract. The reaction was started by the addition of PA. To evaluate the molar ratio among PA, 6HPA, and PMS, the PMS was maintained at 0.1 mM, while the concentration of PA was varied from 0.00 mM to 0.50 mM. One unit of activity was defined as the amount of enzyme that catalyzed the formation of 1 μmol 6HPA in 1 min. Specific activity refers to the number of units of the enzyme present per milligram protein. The kinetic data were evaluated using nonlinear regression analysis with the Michaelis–Menten equation. All of the data were collected from three independent determinations.
2.7. Analytical Methods
The cell density was estimated spectrophotometrically by detecting the absorbance at 600 nm using an UV-VIS spectrophotometer (Shimadzu, UV-2450, Kyoto, Japan). The concentration of PA and its catabolic intermediates in the supernatants was primarily monitored using an UV-VIS spectrophotometer (Shimadzu, UV-2450, Kyoto, Japan). The spectral data were collected from 400 nm to 200 nm. PA and 6HPA were identified using an HPLC analysis on a Shimadzu AD20 system equipped with a C18 reverse phase column (250 × 4.60 mm, 5 μm; Agilent Technologies, Santa Clara, CA, USA). The concentrations of the compounds were calculated using standard samples. The detection wavelength was set at 260 and 310 nm. The mobile phase was a mixture of methanol–water (10:90, vol/vol) with 0.2% formic acid, and the flow rate was 0.7 mL/min. The column temperature was 30 °C. The LC/TOF-MS analysis was performed in a TripleTOF 5600 (AB SCIEX) mass spectrometer [18], and the conditions were identical to that used for HPLC.
3. Results
3.1. Isolation and Identification of the PA-Degrading Strains
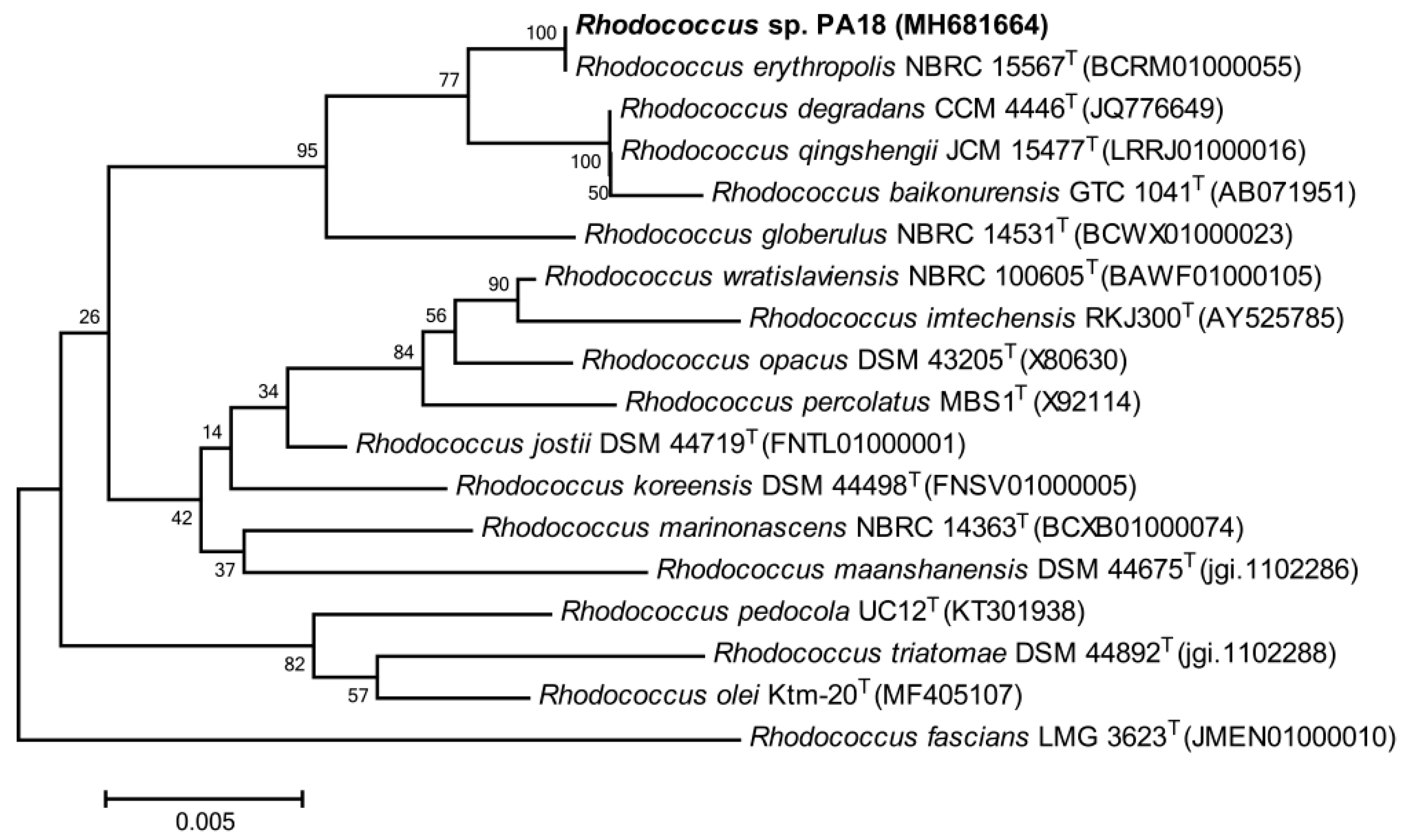
Several strains demonstrated a PA utilization from the enriched soil solution after four weeks. Among these strains, one bacterium, designated as PA18, showed a high PA-degrading efficiency. The strain grew in the presence of 6.5% (w/v) NaCl, and the D-Xylose, cellobiose, lactose, and D-mannitol could be used as the sole source of the carbon and energy. A phylogenetic tree was constructed based on a BLAST analysis of its 16S rDNA sequence (GenBank accession number: MH 681664). The strain PA18 showed a 99%–100.0% sequence similarity to R. erythropolis NBRC 15567T (BCRM01000055), R. degradans CCM 4446T (JQ776649), and R. qingshengii JCM 15477T (LRRJ01000016), and formed a subclade with R. erythropolis NBRC 15567T (Figure 1). Thus, it was identified as a member of Rhodococcus sp. The genus Rhodococcus is considered one of the most promising groups of organisms suitable for the biodegradation of compounds that cannot be easily transformed by other organisms [19]. Members of the genus Rhodococcus exist as abundant indigenous bacterial communities in localities contaminated with various aromatic pollutants [20,21], indicating that Rhodococcus spp. may play an important role in bioremediation. Numerous Rhodococcus have been isolated, for example, R. pyridinivorans PDB9 and Rhodococcus spp. have been reported to degrade pyridine [22,23], Rhodococcus sp. Y22 and R. rhodochrous J1 degrade nicotine or nicotinic acid [24,25], and Rhodococcus sp. B1 degrades quinoline or related-compounds [26]. However, no Rhodococcus species has been shown to degrade PA to date. This study provided new insights regarding PA catabolism and biodegradation by the Rhodococcus species, which may be used for the bioremediation of polluted environments.
3.2. Growth of Strain PA18 and PA Biodegradation
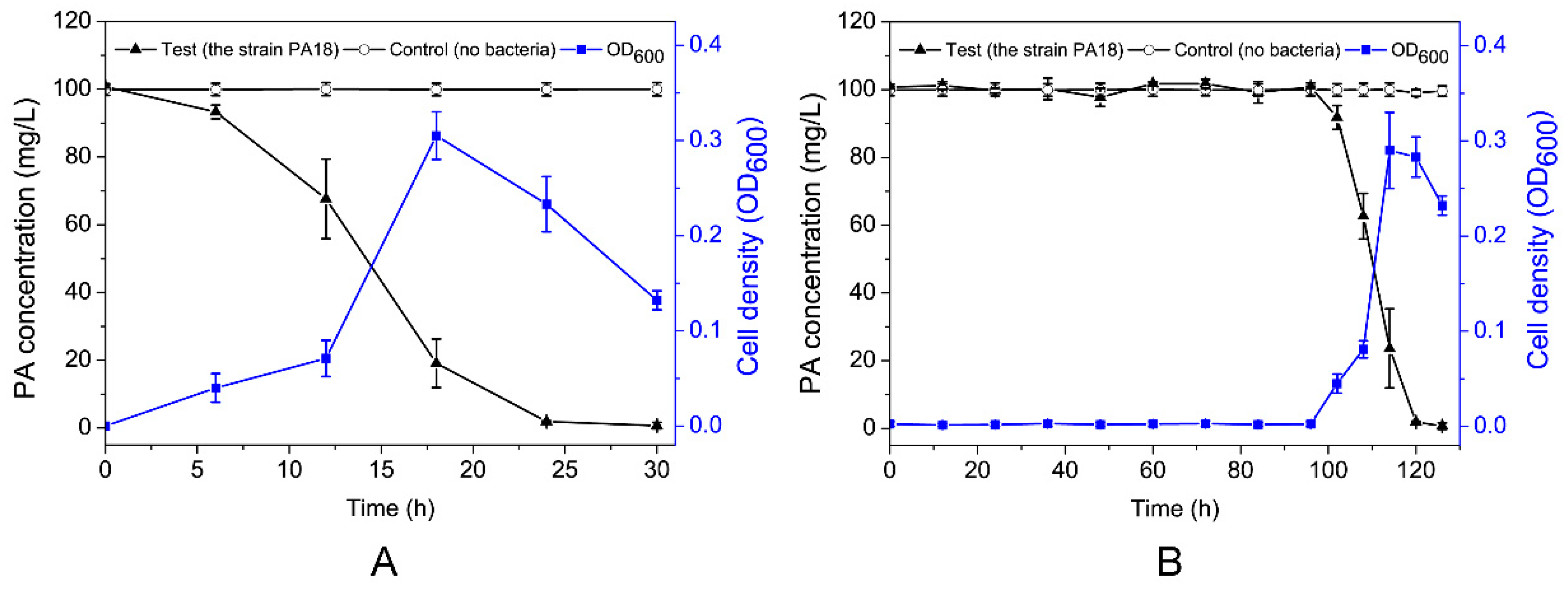
The relationship between the growth of strain PA18 and PA degradation in MSM is indicated by Figure 2. The concentration of PA remained unchanged in the control samples, which did not contain strain PA18. This confirmed that abiotic losses, such as the volatilization and adsorption of PA, were negligible in this study. PA18 degraded 100 mg/L PA at a relatively higher rate within the first 24 h (Figure 2A). The degradation rate decreased substantially from 24 to 28 h, possibly as a result of a lack of substrate, with complete degradation within 28 h. Furthermore, the PA degradation was associated with a concomitant increase in the bacterial cell density. The OD600 value reached 0.31 within 16 h. Interestingly, after a four-day lag phase, the strain PA18 cultured in a LB medium also showed a high-efficiency degradation of PA (Figure 2B). In contrast, the negative control with no inoculated PA18 showed no discernable degradation of PA over the same 120 h. At the same time, the cells grew rapidly with the degradation of PA.
3.3. Effects of Initial PA Concentration and Inoculum of the Strain on PA Biodegradation
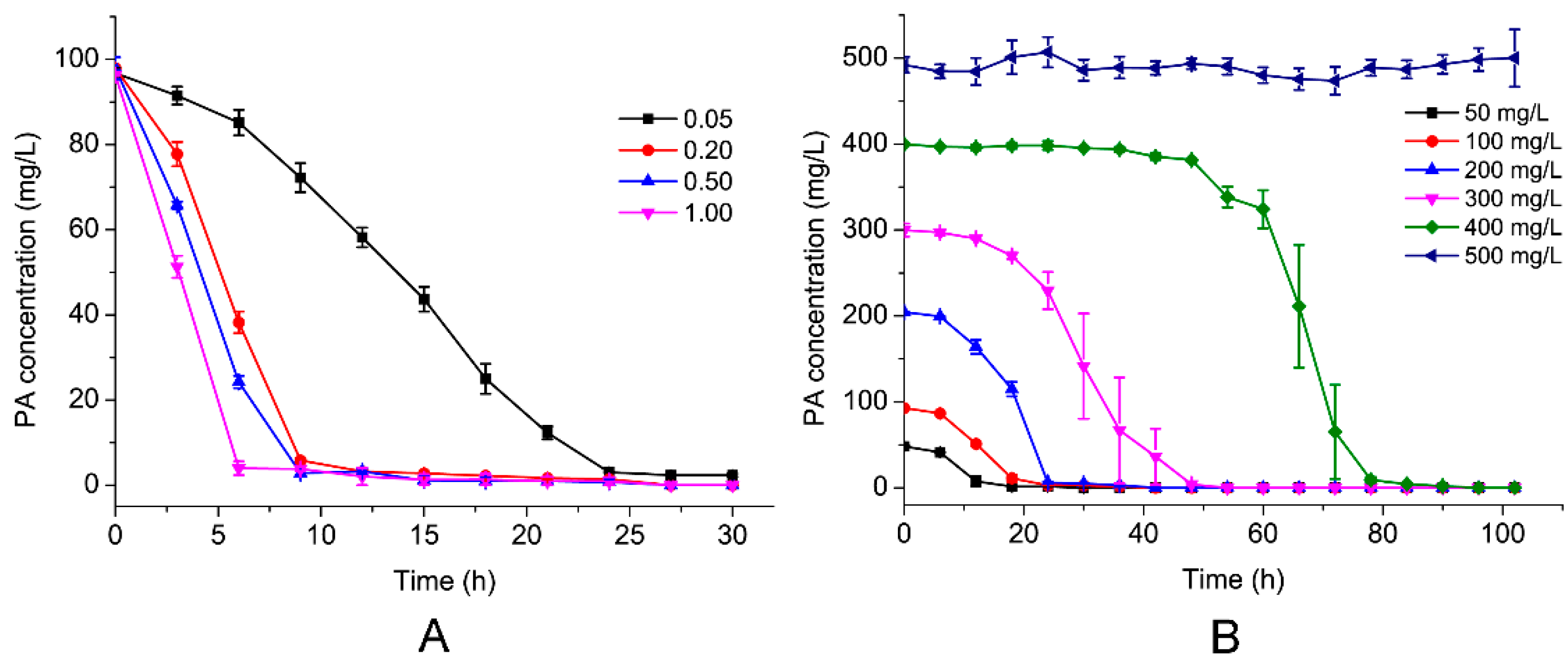
To investigate the effect of the environmental factors on PA biodegradation, the effects of the initial PA concentration and the inoculum of the strain on the PA degradation rate were investigated. As shown in Figure 3A, the PA degradation efficiencies were 10.4%, 28.6%, 68.0%, and 95.9% after 6-h of incubation, from a starting OD600 of 0.05%, 0.20%, 0.50%, and 1.00%, respectively, when the initial PA concentration was 100 mg/L at 30°C, pH 7.0. Furthermore, as shown in Figure 3B, when the initial concentration of PA was increased from 50 mg/L to 300 mg/L, the time required for complete PA removal increased from 18 to 48 h, which increased to 84 h when the initial concentration of PA was increased to 400 mg/L. However, the PA could not be degraded when the concentration was 500 mg/L. One possible reason was that the excessive concentration of PA was toxic for the strain. The delay period was extended as the initial PA concentration increased. Compared with other PA-degrading strains previously reported, such as, Achromobacter sp. JS18 [11], Alcaligenes faecalis JQ135 [12], Arthrobacter picolinophilus DSM 20665 [13], Burkholderia sp. ZD1 [3], and Streptomyces sp. Z2 [14], strain PA18 possessed a very high degradation efficiency.
3.4. Identification of PA Metabolites
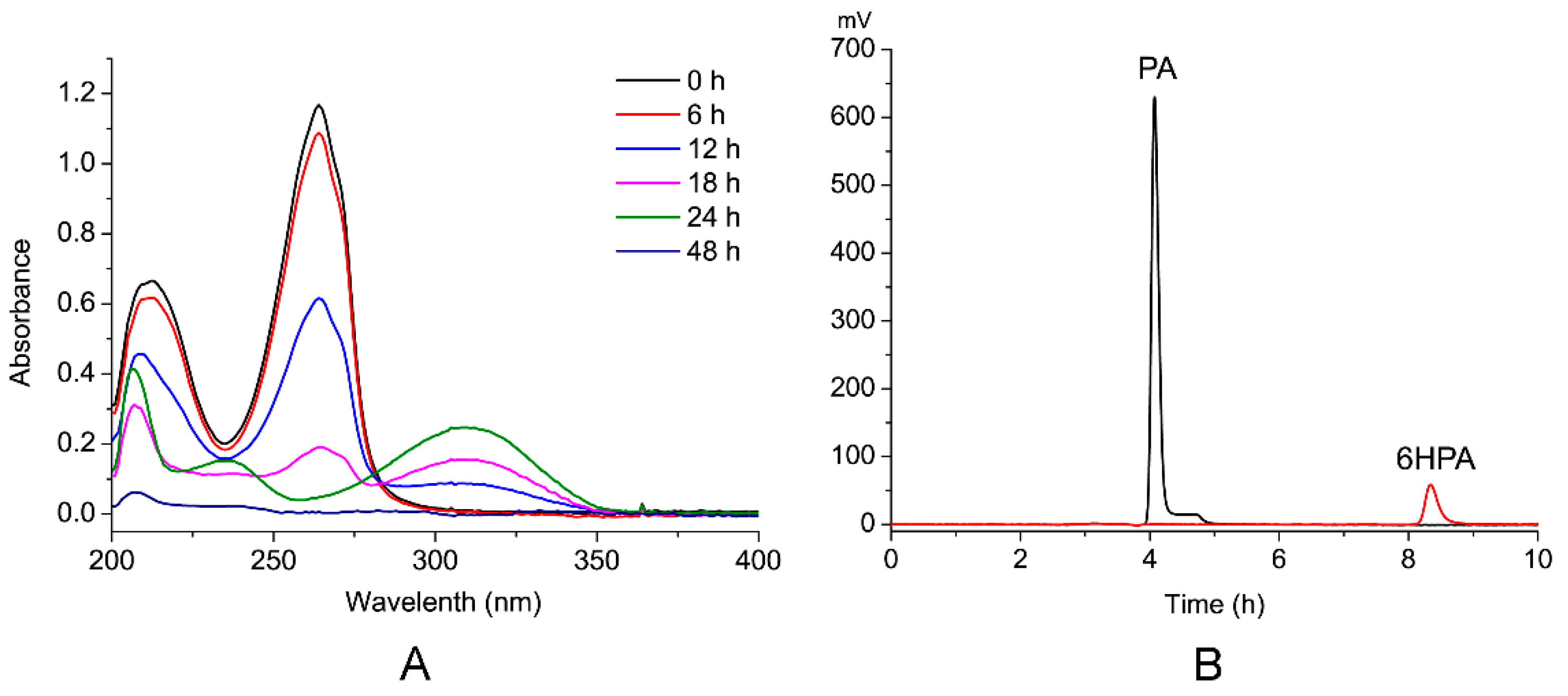
PA degradation by PA18 was analyzed preliminarily using ultraviolet-visible spectroscopy (UV-VIS). In addition to the parent compound, PA (maximum at 260 nm), one major metabolite was detected with an increase in the absorption at 310 nm (Figure 4A). The intermediate product was collected and subjected to high performance liquid chromatography (HPLC). The peak corresponding to PA was detected at a retention time of 4.1 min, and an unknown peak appeared at 8.3 min (Figure 4B).
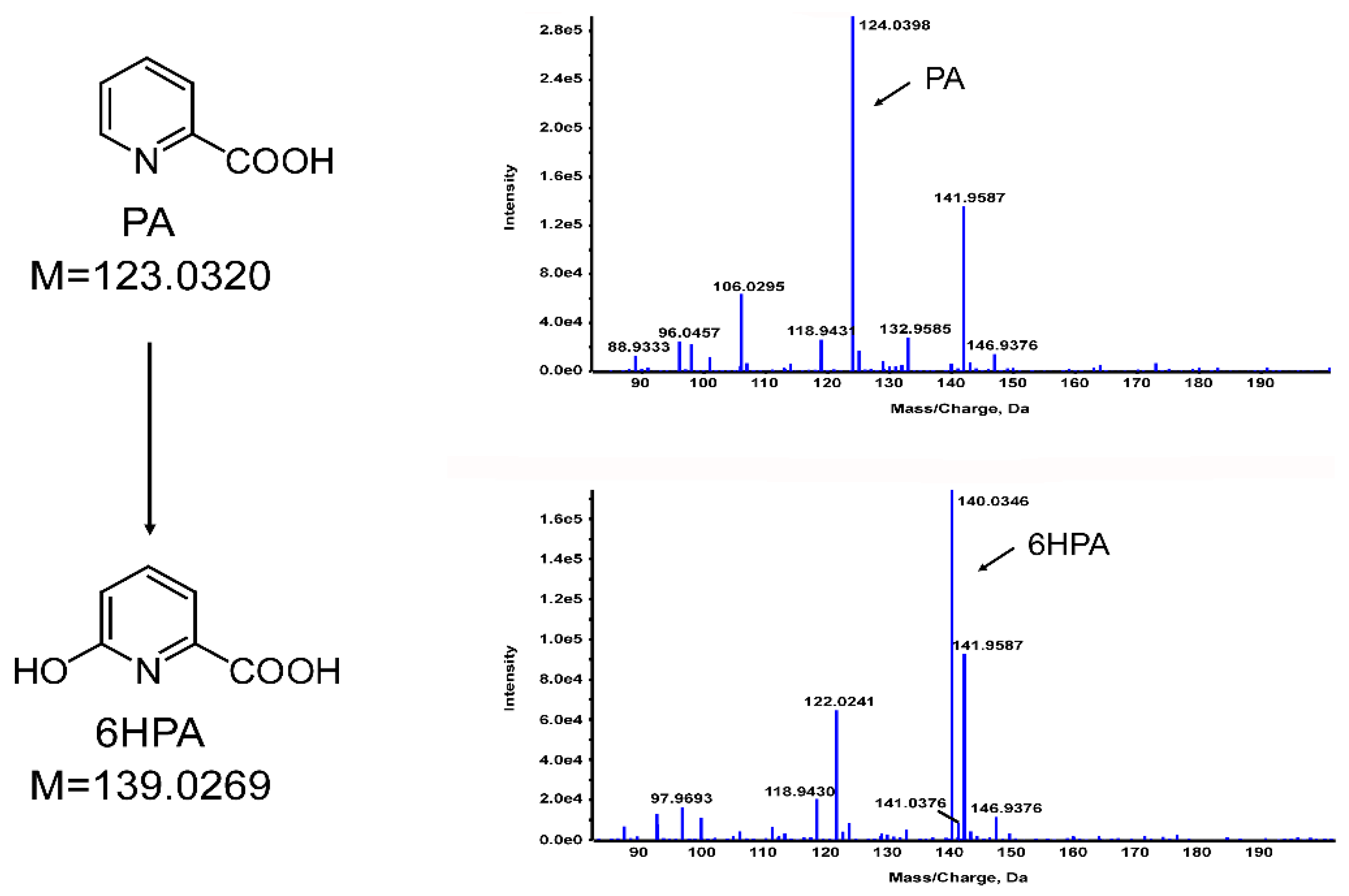
Then, the sample was further analyzed using liquid chromatography/time of flight-mass spectrometry (LC/TOF-MS). The fragment at m/z 124.0398 [M+H]+ conformed to the molecular weight of 124.0354 for PA (Figure 5). In addition, another molecular ion of m/z 140.0346 [M+H]+ was detected. An increment of 16 in the molecular weight indicated that an oxygen atom was added to the PA, which was consistent with the molecular weight of the predicted metabolite 6HPA (C6H5NO3, m/z 140.0303) (Figure 5). Therefore, the PA was possibly hydroxylated by PA18, which was consistent with the results of the previous reports on PA degradation by Burkholderia sp. ZD1 [3] and Alcaligenes faecalis JQ135 [12]. A standard sample of 6HPA was also analyzed using HPLC under the same conditions, and its retention time was identical to that of the unknown peak mentioned above. Therefore, the intermediate metabolite was identified as 6HPA. Throughout the degradation process, no other intermediates were detected.
Usually, the first step in the microbial degradation of the aromatic ring or heterocyclic aromatic compounds is the introduction of a hydroxyl group, and these dehydroxylated substrates can be subsequently decarboxylated [27]. Finally, it can be easily cleaved by dioxygenase or mooxygenase via either an ortho-cleavage pathway or a meta-cleavage pathway, leading to the formation of tricarboxylic acid cycle intermediates [28,29]. The first step, hydroxylation of pyridine or its derivatives, is the key step in biodegradation. Several studies have already shown that pyridine [30], PA [12,31], nicotine [32], and quinolone [33] are hydroxylated at the carbon adjacent to the heteroatom. Pyridine derivatives possessing hydroxyl groups can be used as important starting materials for the synthesis of agrochemicals and pharmaceuticals [34,35]. Future research will be focused on cloning the gene encoding hydroxylase, as well as further study of the enzymatic characteristics.
3.5. Activity of the Enzyme in the Cell Extract
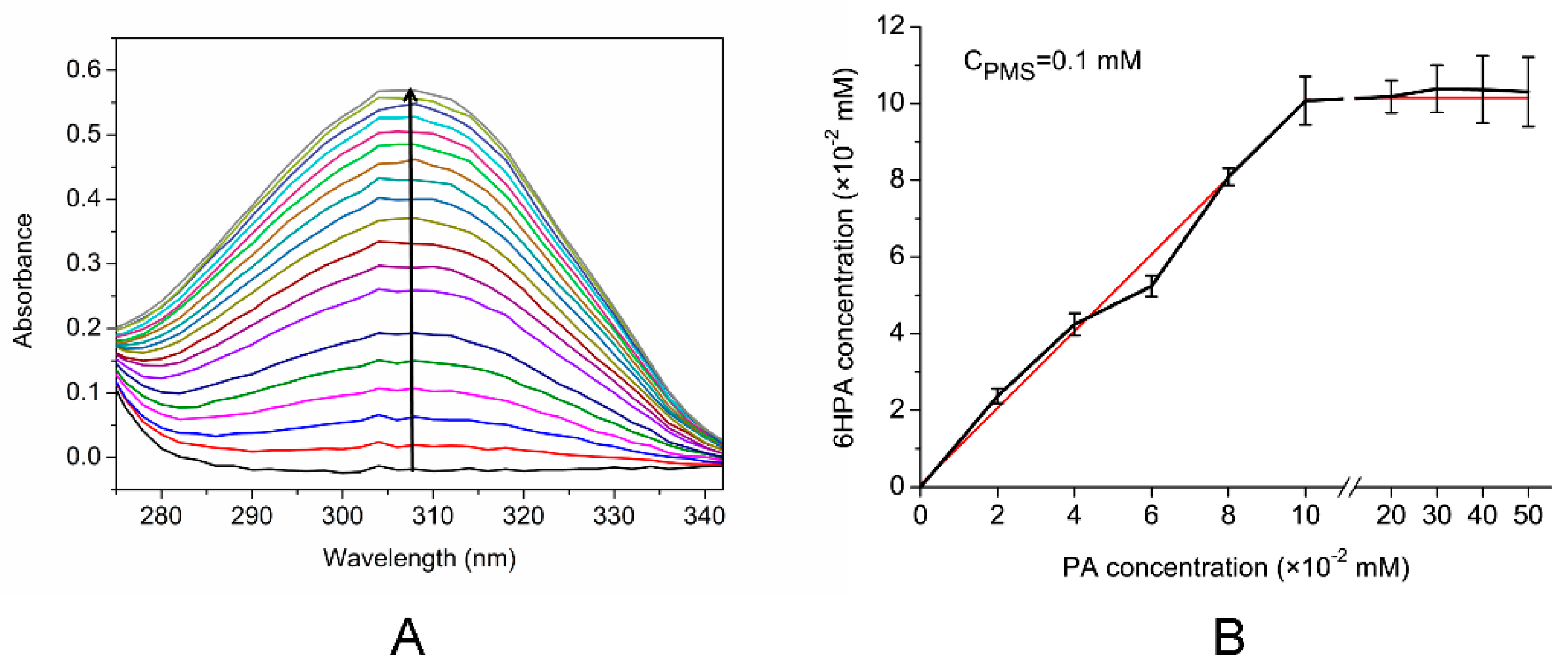
As shown in Figure 6A, a new absorption peak at 310 nm appeared, the intensity of which increased significantly, when PMS acted as an electron acceptor. This corresponded to the absorption maximum of the product 6HPA. However, no 6HPA was produced without the electron acceptor or when the electron acceptor was MB or DCIP (data not shown). These results indicate that the cell extract was able to convert PA into 6HPA, with PMS as the electron acceptor. The Km of the crude enzyme for PA at pH 7.0 and 30°C was 68.86 ± 14.85 μM, and the specific activity was 154.17 U/mg. To verify the quantitative relationship between the PA and PMS, the amount of PA was changed continuously from 0.0 to 0.5 mM, whereas that of PMS was maintained at a constant of 0.1 mM, as shown in Figure 6B. The amount of product 6HPA increased with the concentration of PA, and the PA:6HPA ratio was 1:1. Subsequently, the amount of 6HPA formed did not change, even when the substrate (PA) concentration was increased, indicating that the optimal molar ratio of PA:PMS was 1:1. The hydroxylases of aromatic or N-heterocyclic aromatic compounds that incorporate oxygen into the product are Rieske non-heme iron aromatic ring-hydroxylating oxygenase or cytochrome P450s [29,36]. In strain Pseudomonas putida KT2440, the nicotinic acid can be converted to 6-hydroxynicotinic acid by a two-component hydroxylase (NicAB), whose electron transport chain to the molecular oxygen includes a cytochrome c domain [37]. Interestingly, unlike other studies, the hydroxylase in the strain PA18 cell extract that can convert PA to 6HPA uses PMS as an electron acceptor.
4. Conclusions
In this study, a novel bacterial species, Rhodococcus sp. PA18, was isolated and demonstrated to possess an outstanding PA biodegradation efficiency, even at a low initial inoculum size. This strain completely degraded 100 mg/L PA within 24 h after prior induction. UV-VIS spectroscopy, HPLC, and LC/TOF-MS were used to deduce the biodegradation pathway, and 6HPA was identified as one intermediate. Furthermore, in the crude enzymology experiment, the optimal molar ratio between PA and PMS was 1:1. In addition, the quantitative relationship between the substrate (PA) and the product (6HPA) was 1:1. This is the first study to identify the PA-degrading ability of Rhodococcus. Further studies are required in order to reveal the biochemical process and mechanisms involved in the associated biodegradation pathway.
Author Contributions
conceived and designed the experiments, J.Q., Q.C., and J.H.; performed the experiments, Y.Z., J.J., S.X., and H.M; resources, J.Q., Q.C., and J.H.; writing (original draft preparation), Y.Z. and Q.C.; writing (review and editing), J.Q., B.S., and J.H.; funding acquisition, Q.C. and J.Q.
Funding
This research was funded by the Natural Science Foundation of China (no. 31600080 and no. 31870092) and the Natural Science Foundation of Shandong Province, China (ZR2016CB29).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kaiser, J.P.; Feng, Y.; Bollag, J.M. Microbial metabolism of pyridine, quinoline, acridine, and their derivatives under aerobic and anaerobic conditions. Microbiol. Rev. 1996, 60, 483–498. [Google Scholar] [PubMed]
- Yao, Y.; Tang, H.; Ren, H.; Yu, H.; Wang, L.; Zhang, W.; Behrman, E.J.; Xu, P. Iron(II)-dependent dioxygenase and N-formylamide deformylase catalyze the reactions from 5-hydroxy-2-pyridone to maleamate. Sci. Rep. 2013, 3, 3235. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Wang, Q.; Ning, Y.; Fan, Y.; Feng, S.; He, C.; Zhang, T.C.; Shen, Z. Isolation of a 2-picolinic acid-assimilating bacterium and its proposed degradation pathway. Bioresour. Technol. 2017, 245, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, W.; Wu, Z.L.; Liu, G.M.; Yin, H.; Chen, Y.R.; Hu, N.; Jia, L. Removal of pyridine from its wastewater by using a novel foam fractionation column. Chem. Eng. J. 2017, 321, 151–158. [Google Scholar] [CrossRef]
- Heyes, M.P.; Eugene, O.; Saito, K. Different kynurenine pathway enzymes limit quinolinic acid formation by various human cell types. Biochem. J. 1987, 326, 351–356. [Google Scholar] [CrossRef]
- Bryleva, E.Y.; Brundin, L. Kynurenine pathway metabolites and suicidality. Neuropharmacology 2017, 112, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Esquive, D.G.; Ramirez-Ortega, D.; Pineda, B.; Castro, N.; Rios, C.; de la Cruz, V.P. Kynurenine pathway metabolites and enzymes involved in redox reactions. Neuropharmacology 2017, 112, 331–345. [Google Scholar]
- Asano, Y.; Yamamoto, Y.; Yamada, H. Catechol 2, 3-dioxygenase-catalyzed synthesis of picolinic acids from catechols. Biosci. Biotechnol. Biochem. 1994, 58, 2054–2056. [Google Scholar] [CrossRef]
- Nishino, S.F.; Spain, J.C. Degradation of nitrobenzene by a Pseudomonas pseudoalcaligenes. Appl. Environ. Microbiol. 1993, 59, 2520–2525. [Google Scholar] [PubMed]
- Tian, Y.Q.; Li, W.H. A new process of synthesis of chromium-2-picolinate by chromic anhydride oxidation. Chem. Eng. 2005, 19, 53–54. (In Chinese) [Google Scholar]
- Kutanovas, S.; Karvelis, L.; Vaitekūnas, J.; Stankevičiūtė, J.; Gasparavičiūtė, R.; Meškys, R. Isolation and characterization of novel pyridine dicarboxylic acid-degrading microorganisms. Chemija 2016, 30, 74–83. [Google Scholar]
- Qiu, J.G.; Zhang, J.J.; Zhang, Y.T.; Wang, Y.H.; Tong, L.; Hong, Q.; He, J. Biodegradation of picolinic acid by a newly isolated bacterium Alcaligenes faecalis strain JQ135. Curr. Microbiol. 2017, 74, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, I.; Koenig, K.; Andreesen, J.R. Molybdenum involvement in aerobic degradation of picolinic acid by Arthrobacter picolinophilus. FEMS Microbiol. Lett. 1990, 67, 281–284. [Google Scholar] [CrossRef]
- Zheng, C.; Zhou, J.; Wang, J.; Qu, B.; Lu, H.; Zhao, H. Aerobic degradation of 2-picolinic acid by a nitrobenzene-assimilating strain: Streptomyces sp. Z2. Bioresour. Technol. 2009, 100, 2082–2084. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.J.; Hang, B.J.; Cai, S.; Xie, X.T.; He, J.; Li, S.P. Degradation of cyhalofop-butyl (CyB) by Pseudomonas azotoformans strain QDZ-1 and cloning of a novel gene encoding CyB-hydrolyzing esterase. J. Agric. Food Chem. 2011, 59, 6040–6046. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Qiu, J.G.; Liu, B.; Zhao, L.; Zhang, Y.T.; Cheng, D.; Yan, X.; Jiang, J.D.; Hong, Q.; He, J. A novel degradation mechanism for pyridine derivatives in Alcaligenes faecalis JQ135. Appl. Environ. Microbiol. 2018, 84, e00910-18. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.G.; Jiang, W.K.; Wang, X.H.; Cheng, T.; Zhang, D.S.; Wang, H.; Qiu, J.G.; Cao, L.; Hong, Q. An amidase gene ipaH is responsible for the initial degradation step of iprodione in strain Paenarthrobacter sp. YJN-5. Appl. Environ. Microbiol. 2018, 84, e01150-18. [Google Scholar] [CrossRef] [PubMed]
- Warhurst, A.M.; Fewson, C.A. Biotransformations catalyzed by the genus Rhodococcus. Crit. Rev. Biotechnol. 1994, 14, 29–73. [Google Scholar] [CrossRef]
- Fahy, A.; McGenity, T.J.; Timmis, K.N.; Ball, A.S. Heterogeneous aerobic benzene-degrading communities in oxygen-depleted groundwaters. FEMS Microbiol. Ecol. 2006, 58, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, M.B.; Prouzová, P.; Macková, M.; Macek, T.; Nagle, D.P.; Fletcher, J.S. Polychlorinated biphenyl (PCB)-degrading bacteria associated with trees in a PCB-contaminated site. Appl. Environ. Microbiol. 2006, 72, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.T.; Lee, S.B.; Park, Y.H. Characterization of a pyridine-degrading branched Gram-positive bacterium isolated from the anoxic zone of an oil shale column. Appl. Microbiol. Biotechnol. 1991, 35, 824–829. [Google Scholar] [CrossRef]
- Yoon, J.H.; Kang, S.S.; Cho, Y.G.; Lee, S.T.; Kho, Y.H.; Kim, C.J.; Park, Y.H. Rhodococcus pyridinivorans sp. nov., a pyridine-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2000, 50, 2173–2180. [Google Scholar] [CrossRef] [PubMed]
- Mathew, C.D.; Nagasawa, T.; Kobayashi, M.; Yamada, H. Nitrilase-catalyzed production of nicotinic acid from 3-cyanopyridine in Rhodococcus rhodochrous J1. Appl. Environ. Microbiol. 1988, 54, 1030–1032. [Google Scholar] [PubMed]
- Gong, X.W.; Ma, G.H.; Duan, Y.Q.; Zhu, D.L.; Chen, Y.K.; Zhang, K.Q.; Yang, J.K. Biodegradation and metabolic pathway of nicotine in Rhodococcus sp. Y22. World J. Microbiol. Biotechnol. 2016, 32, 188. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G.; Bauder, R.; Speer, M.; Rommet, T.O.; Lingens, F. Microbial metabolism of quinoline and related compounds. II. Degradation of quinoline by Pseudomonas fluorescens 3, Pseudomonas putida 86 and Rhodococcus spec. B1. Biol. Chem. Hoppe Seyler 1989, 370, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Nagasawa, T. Enzymatic functionalization of aromatic N-heterocycles: Hydroxylation and carboxylation. J. Biosci. Bioeng. 2000, 89, 111–118. [Google Scholar] [CrossRef]
- Mallick, S.; Chakraborty, J.; Dutta, T.K. Role of oxygenases in guiding diverse metabolic pathways in the bacterial degradation of low-molecular-weight polycyclic aromatic hydrocarbons: A review. Crit. Rev. Microbiol. 2011, 37, 64–90. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.K.; Cain, R.B. Microbial metabolism of the pyridine ring. Metabolic pathways of pyridine biodegradation by soil bacteria. Biochem. J. 1975, 146, 157–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orpin, C.G.; Knight, M.; Evans, W.C. The bacterial oxidation of picolinamide, a photolytic product of diquat. Biochem. J. 1972, 127, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.G.; Ma, Y.; Wen, Y.Z.; Chen, L.S.; Wu, L.F.; Liu, W.P. Functional identification of two novel genes from Pseudomonas sp. strain HZN6 involved in the catabolism of nicotine. Appl. Environ. Microbiol. 2012, 78, 2154–2160. [Google Scholar] [CrossRef] [PubMed]
- Shukla, O.P. 8-Hydroxycoumarin: An intermediate in the microbial transformation of quinoline. Curr. Sci. 1984, 53, 1145–1147. [Google Scholar]
- Yasuda, M.; Sakamoto, T.; Sashida, R.; Ueda, M.; Morimoto, Y.; Nagasawa, T. Microbial hydroxyIation of 3-cyanopyridine to 3-cyano-6-hydroxypyridine. Biosci. Biotechnol. Biochem. 1995, 59, 572–575. [Google Scholar] [CrossRef]
- Yu, H.; Tang, H.Z.; Xu, P. Green strategy from waste to value-added-chemical production: Efficient biosynthesis of 6-hydroxy-3-succinoyl-pyridine by an engineered biocatalyst. Sci. Rep. 2014, 4, 5397. [Google Scholar] [CrossRef] [PubMed]
- Hannemann, F.; Bichet, A.; Ewen, K.M.; Bernhardt, R. Cytochrome P450 systems-biological variations of electron transport chains. BBA-Gen Subj. 2007, 1770, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, J.I.; Canales, Á.; Jiménez-Barbero, J.; Ginalski, K.; Rychlewski, L.; García, J.L.; Díaz, E. Deciphering the genetic determinants for aerobic nicotinic acid degradation: The nic cluster from Pseudomonas putida KT2440. Proc. Natl. Acad. Sci. USA 2008, 105, 11329–11334. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Neighbor-joining phylogenetic tree shows the position of strain picolinic acid 18 (PA18) among some members of the genus Rhodococcus, based on the 16S rDNA sequences.
Figure 1.
Neighbor-joining phylogenetic tree shows the position of strain picolinic acid 18 (PA18) among some members of the genus Rhodococcus, based on the 16S rDNA sequences.

Figure 2.
Utilization of PA during the growth of Rhodococcus sp. PA18 in mineral salts medium (MSM) (pH 7.0). The strain was pre-cultured with PA (A). The strain was pre-cultured in a Luria Bertani (LB) medium (B).
Figure 2.
Utilization of PA during the growth of Rhodococcus sp. PA18 in mineral salts medium (MSM) (pH 7.0). The strain was pre-cultured with PA (A). The strain was pre-cultured in a Luria Bertani (LB) medium (B).

Figure 3.
Effects of inocula (A) and the initial PA concentration (B) on the degradation of PA in MSM. The error bars show standard deviations.
Figure 3.
Effects of inocula (A) and the initial PA concentration (B) on the degradation of PA in MSM. The error bars show standard deviations.

Figure 4.
PA degradation analysis by the strain PA18 using UV-VIS and high-performance liquid chromatography (HPLC). (A) The UV-VIS absorption of each compound during PA degradation from 200 to 400 nm. (B) HPLC analysis of the conversion of PA into 6HPA at 12 h. The black line indicates the sample collected at 0 h and the red line indicates at 12 h.
Figure 4.
PA degradation analysis by the strain PA18 using UV-VIS and high-performance liquid chromatography (HPLC). (A) The UV-VIS absorption of each compound during PA degradation from 200 to 400 nm. (B) HPLC analysis of the conversion of PA into 6HPA at 12 h. The black line indicates the sample collected at 0 h and the red line indicates at 12 h.

Figure 5.
LC/TOF-MS profiles of PA and 6HPA.

Figure 6.
Measurement of the enzyme activities in cell-free extracts. (A) Spectrophotometric changes during the transformation of PA. The spectra were recorded every 1 min. The arrow indicates the directions of the spectral changes. (B) Relationship between PA and 6HPA while the concentration of PMS was constant. PA concentration was increased from 0.00 to 0.50 mM whereas the phenazine methosulfate (PMS), concentration was constant at 0.10 mM. The data were calculated from three independent replicates, and error bars indicate standard deviations.
Figure 6.
Measurement of the enzyme activities in cell-free extracts. (A) Spectrophotometric changes during the transformation of PA. The spectra were recorded every 1 min. The arrow indicates the directions of the spectral changes. (B) Relationship between PA and 6HPA while the concentration of PMS was constant. PA concentration was increased from 0.00 to 0.50 mM whereas the phenazine methosulfate (PMS), concentration was constant at 0.10 mM. The data were calculated from three independent replicates, and error bars indicate standard deviations.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Ji, J.; Xu, S.; Wang, H.; Shen, B.; He, J.; Qiu, J.; Chen, Q. Biodegradation of Picolinic Acid by Rhodococcus sp. PA18. Appl. Sci. 2019, 9, 1006. https://0-doi-org.brum.beds.ac.uk/10.3390/app9051006
AMA Style
Zhang Y, Ji J, Xu S, Wang H, Shen B, He J, Qiu J, Chen Q. Biodegradation of Picolinic Acid by Rhodococcus sp. PA18. Applied Sciences. 2019; 9(5):1006. https://0-doi-org.brum.beds.ac.uk/10.3390/app9051006
Chicago/Turabian StyleZhang, Yanting, Junbin Ji, Siqiong Xu, Hongmei Wang, Biao Shen, Jian He, Jiguo Qiu, and Qing Chen. 2019. "Biodegradation of Picolinic Acid by Rhodococcus sp. PA18" Applied Sciences 9, no. 5: 1006. https://0-doi-org.brum.beds.ac.uk/10.3390/app9051006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.