Proliferation, Adult Neuronal Stem Cells and Cells Migration in Pallium during Constitutive Neurogenesis and after Traumatic Injury of Telencephalon of Juvenile Masu Salmon, Oncorhynchus masou

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Animals
2.2. Experimental Damage to the Brain by the Kishimoto Method
2.3. Preparation of Material for Immunohistochemical Studies
2.4. Experimental BrdU Labeling
2.5. Immunohistochemical Detection of Doublecortin, Vimentin and Glial Fibrillar Acidic Protein
2.6. Microscopy
2.7. Statistical Analysis
3. Results
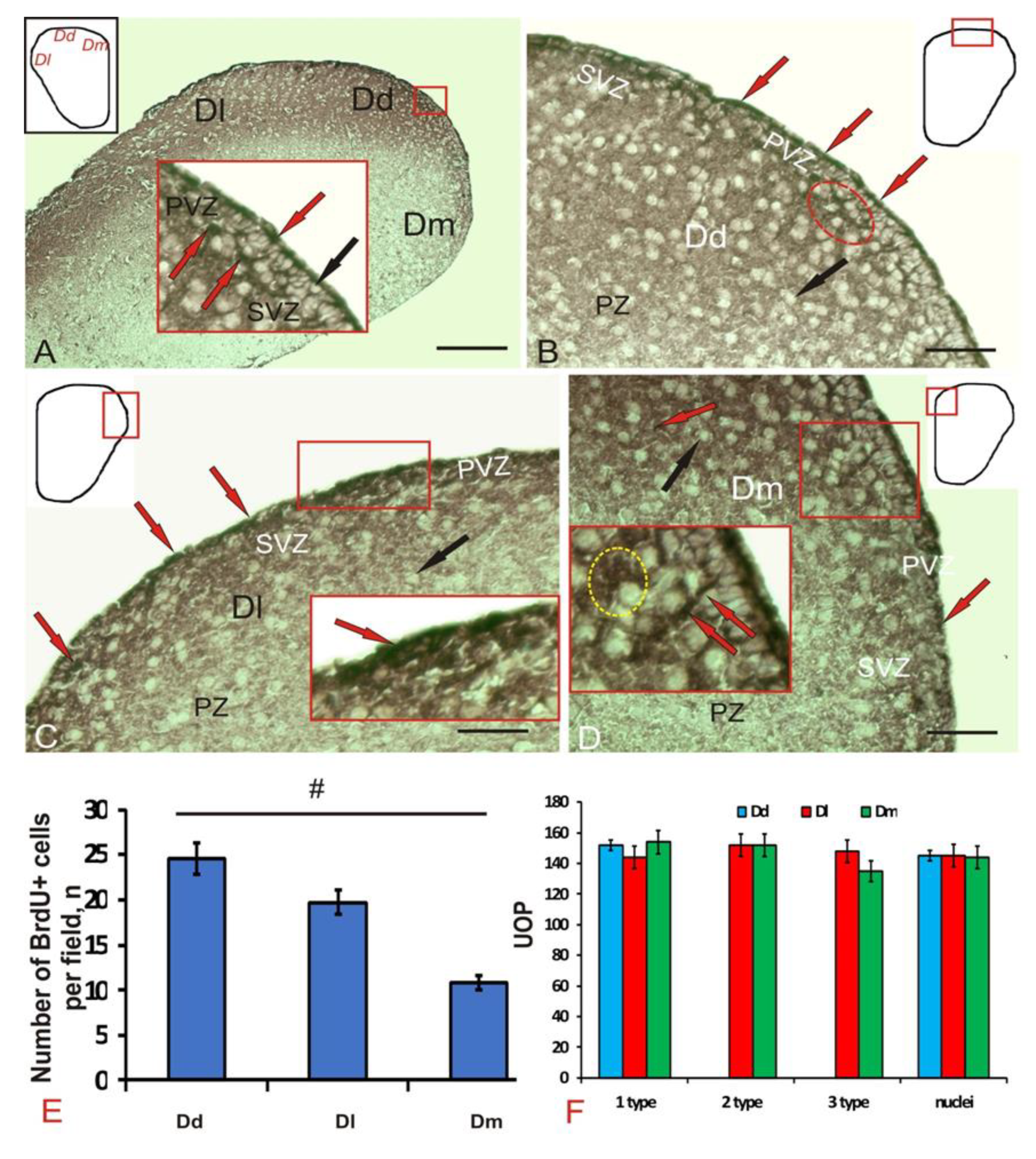
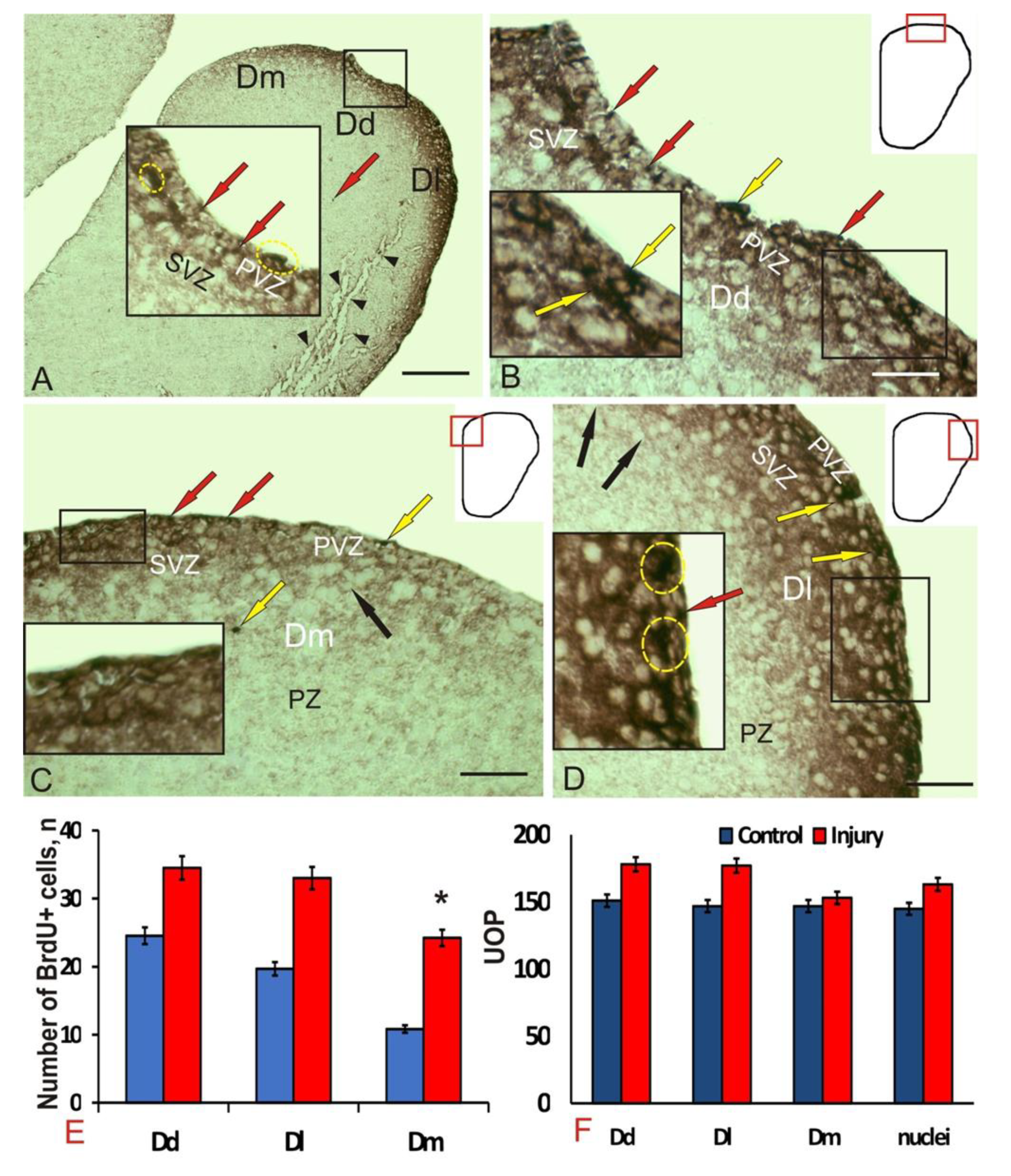
3.1. Experimental Labeling of Brdu in the Intact Pallium of Juvenile O. masou and after a Traumatic Injury
3.2. Immunohistochemical Gfap Labeling in the Intact Juvenile O. masou Pallium and after Traumatic Injury
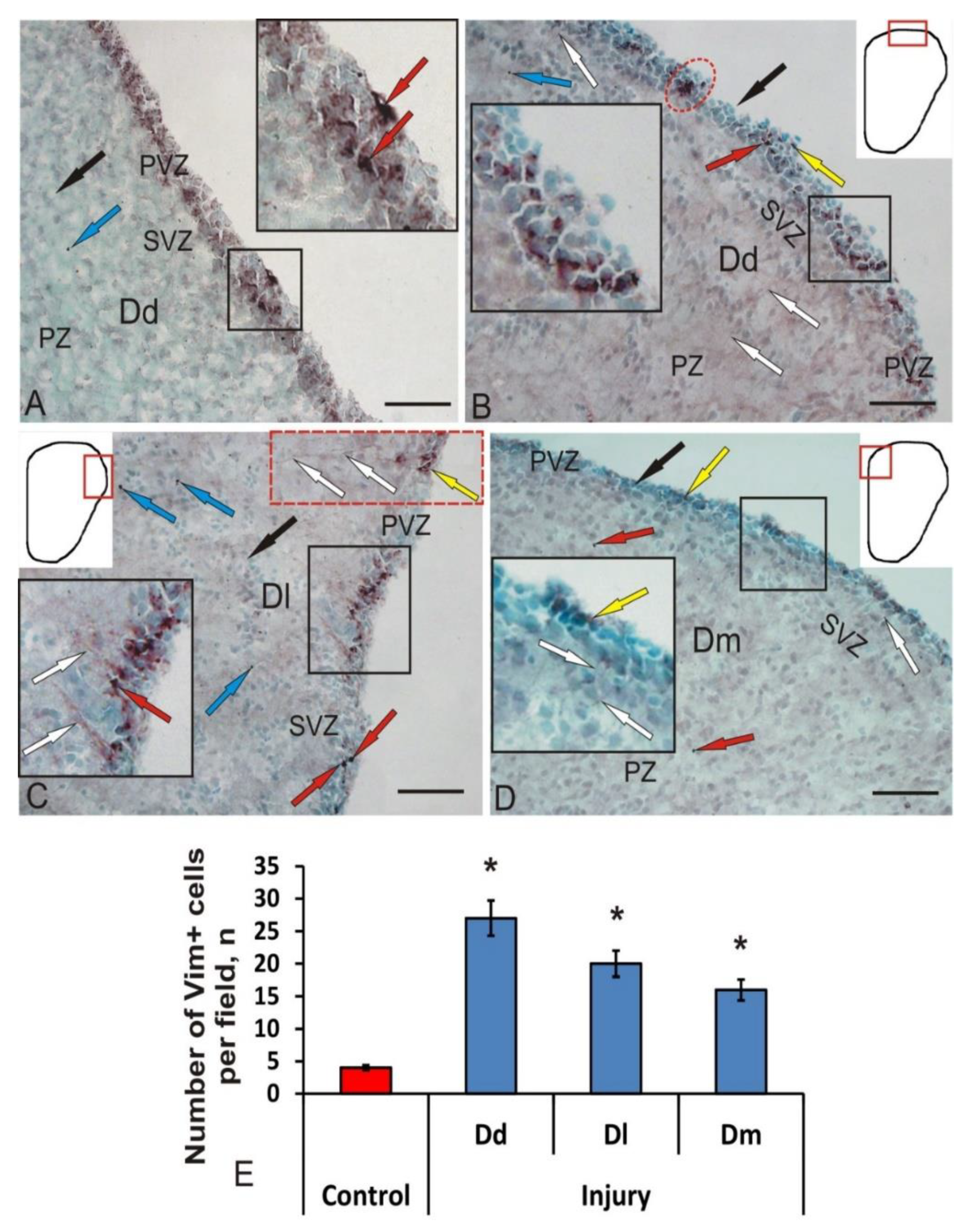
3.3. Immunohistochemical Labeling of Vimentin in the Intact Juvenile O. masou Pallium and after Traumatic Injury
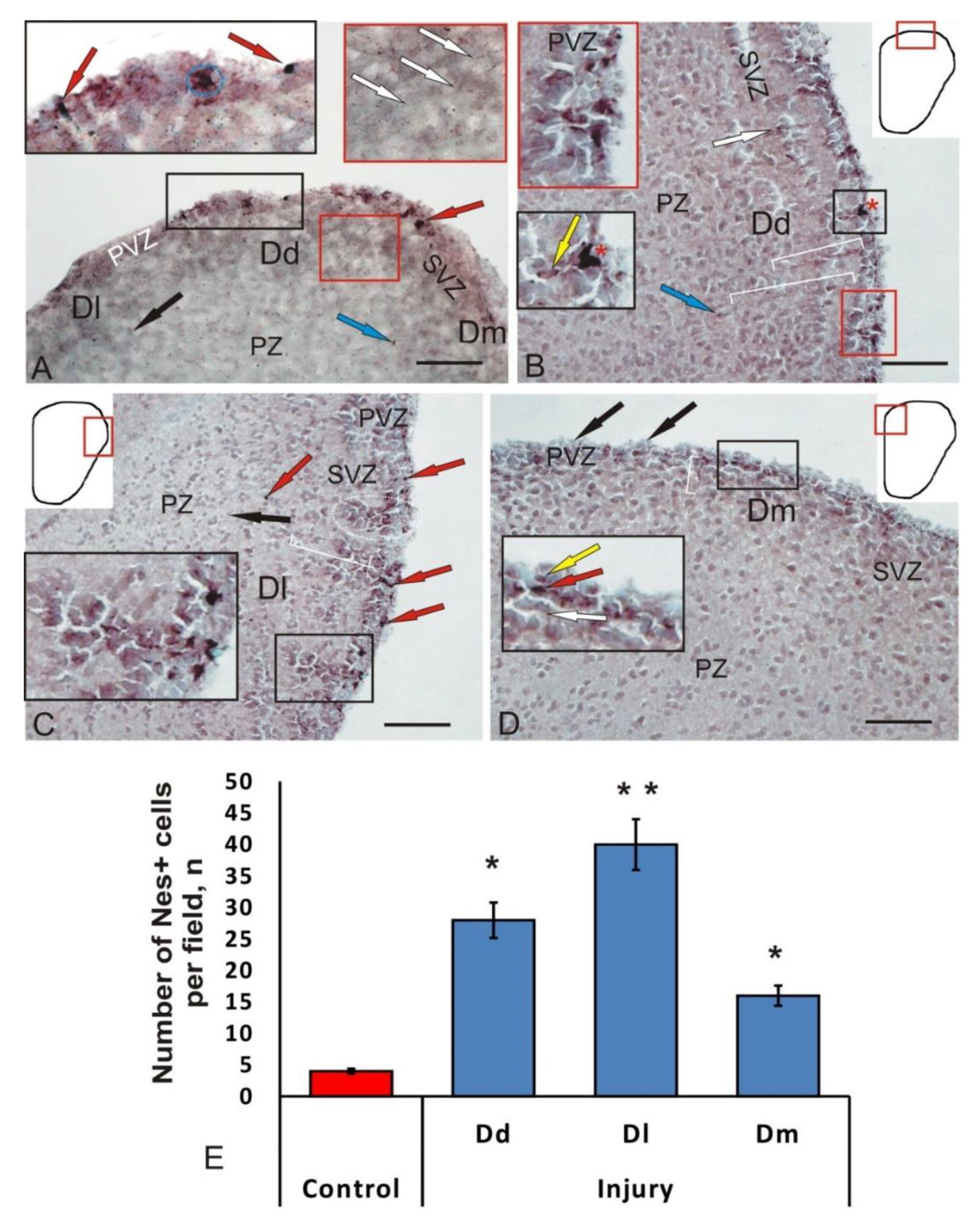
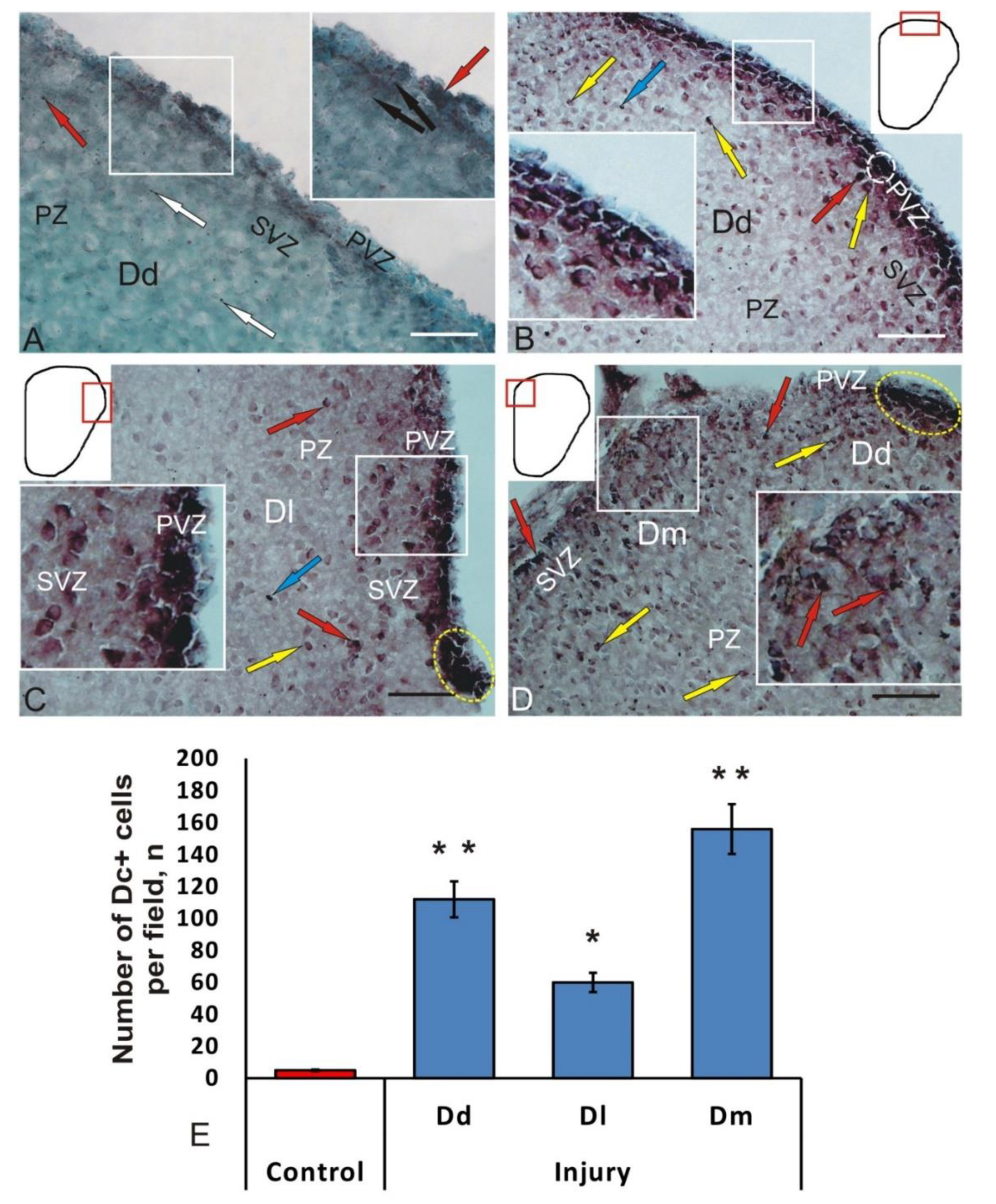
3.4. Immunohistochemical Labeling of Doublecortin in the Intact Juvenile O. masou Pallium and after Traumatic Injury
4. Discussion
4.1. Expression of Brdu in the Juvenile Masu Salmon Pallium under Normal Conditions and after Traumatic Injury
4.2. Expression of Gfap and Vimentin in the Brain of Fish under Normal Conditions and after Traumatic Injury
4.3. Expression of Doublecortin in the Intact Brain of Fish and after Traumatic Injury
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Augusto-Oliveira, M.; Arrifano, G.P.F.; Malva, J.O.; Crespo-Lopez, M.E. Adult hippocampal neurogenesis in different taxonomic groups: Possible functional similarities and striking controversies. Cells 2019, 8, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandel, H.; Brand, M. Comparative aspects of adult neural stem cell activity in vertebrates. Dev. Genes Evol. 2013, 223, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Kroehne, V.; Freudenreich, D.; Hans, S.; Kaslin, J.; Brand, M. Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors. Development 2011, 138, 4831–4841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambroise, E.; Simion, M.; Bourquard, T.; Rizzi, B.; Jaszczyszyn, Y.; Bourge, M.; Affaticati, P.; Heuzé, A.; Jouralet, J. Postembryonic fish brain proliferation zones exhibit neuroepithelial-type gene expression profile. Stem Cells 2017, 35, 1505–1518. [Google Scholar] [CrossRef] [Green Version]
- Lam, C.S.; Marz, M.; Strahle, U. Gfap and nestin reporter lines reveal characteristics of neural progenitors in the adult zebrafish brain. Dev. Dyn. 2009, 238, 475–486. [Google Scholar] [CrossRef]
- Xu, L.; Tang, X.; Wang, Y.; Xu, H.; Fan, X. Radial glia, the keystone of the development of the hippocampal dentate gyrus. Mol. Neurobiol. 2015, 51, 131–141. [Google Scholar] [CrossRef]
- Kriegstein, A.; Alvarez-Buylla, A. The glial nature of embryonic and adult neural stem cells. Ann. Rev. Neurosci. 2009, 32, 149–184. [Google Scholar] [CrossRef] [Green Version]
- Codega, P.; Silva-Vargas, V.; Paul, A.; Maldonado-Soto, A.R.; Deleo, A.M.; Pastrana, E.; Doetsch, F.P. Prospective identification and purification of quiescent adult neural stem cells from their in vivo niche. Neuron 2014, 82, 545–559. [Google Scholar] [CrossRef] [Green Version]
- Giachino, C.; Basak, O.; Lugert, S.; Knuckles, P.; Obernier, K.; Fiorelli, R.; Frank, S.; Raineteau, O.; Alvarez-Buylla, A.; Taylor, V. Molecular diversity subdivides the adult forebrain neural stem cell population. Stem Cells 2014, 32, 70–84. [Google Scholar] [CrossRef] [Green Version]
- Rothenaigner, I.; Krecsmarik, M.; Hayes, J.A.; Bahn, B.; Lepier, A.; Fortin, G.; Götz, M.; Jagasia, R.; Bally-Cuif, L. Clonal analysis by distinct viral vectors identifies bona fide neural stem cells in the adult zebrafish telencephalon and characterizes their division properties and fate. Development 2011, 138, 1459–1469. [Google Scholar] [CrossRef] [Green Version]
- Stigloher, C.; Chapouton, P.; Adolf, B.; Bally-Cuif, L. Identification of neural progenitor pools by E(Spl) factors in the embryonic and adult brain. Brain Res. Bull. 2008, 75, 266–273. [Google Scholar] [CrossRef]
- Olivera-Pasilio, V.; Lasserre, M.; Castelló, M.E. Cell Proliferation, Migration, and Neurogenesis in the Adult Brain of the Pulse Type Weakly Electric Fish, Gymnotus omarorum. Front. Neurosci. 2017, 11, 437. [Google Scholar] [CrossRef] [Green Version]
- Pushchina, E.V.; Stukaneva, M.E.; Varaksin, A.A. Damage-related changes in the cerebellum of juvenile Oncorhynchus masou: Reactivation of neurogenic niches and astrocytic response. J. R. Sci. 2019, 1, 36–54. [Google Scholar]
- Pushchina, E.V.; Kapustyanov, I.A.; Varaksin, A.A. Proliferation and neuro- and gliogenesis in normal and mechanically damaged mesencephalic tegmentum in juvenile chum salmon Oncorhynchus keta. Russ. J. Dev. Biol. 2019, 50, 59–76. [Google Scholar] [CrossRef]
- Pushchina, E.V.; Zharikova, E.I.; Varaksin, A.A. Persistent and reparative neurogenesis in the juvenile masu salmon Oncorhynchus masou telencephalon after mechanical injury. Russ. J. Dev. Biol. 2017, 48, 307–320. [Google Scholar] [CrossRef]
- Alunni, A.; Hermel, J.-M.; Heuze, A.; Bourrat, F.; Jamen, F.; Joly, J.S. Evidence for neural stem cells in the medaka optic tectum proliferation zones. Dev. Neurobiol. 2010, 70, 693–713. [Google Scholar] [CrossRef]
- Kaslin, J.; Ganz, J.; Geffarth, M.; Grandel, H.; Hans, S.; Brand, M. Stem cells in the adult zebrafish cerebellum: Initiation and maintenance of a novel stem cell niche. J. Neurosci. 2009, 29, 6142–6153. [Google Scholar] [CrossRef] [Green Version]
- Dirian, L.; Galant, S.; Coolen, M.; Chen, W.; Bedu, S.; Houart, C.; Bally-Cuif, L.; Foucher, I. Spatial regionalization and heterochrony in the formation of adult pallial neural stem cells. Dev. Cell 2014, 30, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, N.; Shimizu, K.; Sawamoto, K. Neuronal regeneration in a zebrafish model of adult brain injury. Dis. Model. Mech. 2012, 5, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Merkulov, G.A. Kurs Patologogistologicheskoi Tekhniki (A Course in Pathohistological Techniques); Meditsina: Leningrad, Russia, 1969. [Google Scholar]
- Dolbeare, F. Bromodeoxyuridine: Diagnostic toll in biology and medicine. Part I: Historical perspectives, histochemical methods and cell kinetics. Histochem. J. 1995, 27, 339–369. [Google Scholar] [CrossRef]
- Arochena, M.; Anadon, R.; Diaz-Regueira, S.M. Development of vimentin and glial fibrillary acidic protein immunoreactivities in the brain of gray mullet (Chelon Labrosus), an advanced teleost. J. Comp. Neurol. 2004, 469, 413–436. [Google Scholar] [CrossRef] [PubMed]
- Ayanlaja, A.A.; Xiong, Y.; Gao, Y.; Ji, G.; Tang, C.; Abdikani Abdullah, Z.; Gao, D. Distinct features of doublecortin as a marker of neuronal migration and its implications in cancer cell mobility. Front. Mol. Neurosci. 2017, 10, 199. [Google Scholar] [CrossRef] [Green Version]
- Adolf, B.; Chapouton, P.; Lam, C.S.; Topp, S.; Tannhauser, B.; Strahle, U.; Gotz, M.; Bally-Cuif, L. Conserved and acquired features of adult neurogenesis in the zebrafish telencephalon. Dev. Biol. 2006, 295, 278–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, E.; Mouriec, K.; Anglade, I.; Menuet, A.; Le Page, Y.; Gueguen, M.M.; Marmignon, M.H.; Brion, F.; Pakdel, F.; Kah, O. Identification of aromatase-positive radial glial cells as progenitor cells in the ventricular layer of the forebrain in zebrafish. J. Comp. Neurol. 2007, 501, 150–167. [Google Scholar] [CrossRef]
- Fernández, A.S.; Rosillo, J.C.; Casanova, G.; Olivera-Bravo, S. Proliferation zones in the brain of adult fish Austrolebias (Cyprinodontiform: Rivulidae): A comparative study. Neuroscience 2011, 189, 12–24. [Google Scholar] [CrossRef]
- Rosillo, J.; Torres, C.M.; Olivera-Bravo, S.; Casanova, G.; García-Verdugo, J.M.; Fernández, A.S. Telencephalic-olfactory bulb ventricle wall organization in Austrolebias charrua: Cytoarchitecture, proliferation dynamics, neurogenesis and migration. Neuroscience 2016, 12, 63–80. [Google Scholar] [CrossRef]
- Terzibasi, E.T.; Baumgart, M.; Battistoni, G.; Cellerino, A. Adult neurogenesis in the short-lived teleost Nothobranchius furzeri: Localization of neurogenic niches, molecular characterization and effects of aging. Aging Cell. 2012, 11, 241–251. [Google Scholar]
- Teles, M.C.; Sîrbulescu, R.F.; Wellbrock, U.M.; Oliveira, R.F.; Zupanc, G.K.H. Adult neurogenesis in the brain of the Mozambique tilapia, Oreochromis mossambicus. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2012, 198, 427–449. [Google Scholar] [CrossRef]
- Traniello, I.M.; Sîrbulescu, R.F.; Ilieş, I.; Zupanc, G.K. Age-related changes in stem cell dynamics, neurogenesis, apoptosis, and gliosis in the adult brain: A novel teleost fish model of negligible senescence. Dev. Neurobiol. 2014, 74, 514–530. [Google Scholar] [CrossRef]
- Candal, E.; Anadon, R.; Bourrat, F.; Rodriguez-Moldes, I. Cell proliferation in the developing and adult hindbrain and midbrain of trout and medaka (teleosts): A segmental approach. Brain Res. Dev. Brain Res. 2005, 160, 157–175. [Google Scholar] [CrossRef]
- Wullimann, M.; Puelles, L. Postembrionic neural proliferation in the zebrafish forebrain and its relationship to prosomeric domains. Anat. Embryol. 1999, 329, 329–348. [Google Scholar] [CrossRef]
- Bravo, R.; Frank, R.; Blundell, P.A.; Macdonald-Bravo, H. Cyclin/PCNA is the auxiliary protein of DNA polymerase-delta. Nature 1987, 326, 515–517. [Google Scholar] [CrossRef]
- Zupanc, G.K.H.; Sirbulescu, R.F. Adult neurogenesis and neuronal regeneration in the central nervous system of teleost fish. Eur. J. Neurosci. 2011, 34, 917–929. [Google Scholar] [CrossRef]
- Barbosa, J.S.; Sanchez-Gonzalez, R.; Di Giaimo, R.; Baumgart, E.V.; Theis, F.J.; Götz, M.; Ninkovic, J. Neurodevelopment. Live imaging of adult neural stem cell behavior in the intact and injured zebrafish brain. Science 2015, 348, 789–793. [Google Scholar] [CrossRef]
- Ganz, J.; Kaslin, J.; Hochmann, S.; Freudenreich, D.; Brand, M. Heterogeneity and Fgf dependence of adult neural progenitors in the zebrafish telencephalon. Glia 2010, 58, 1345–1363. [Google Scholar] [CrossRef]
- Fishell, G.; Goldman, J.E. A silver lining to stroke: Does ischemia generate new cortical interneurons? Nat. Neurosci. 2010, 13, 145–146. [Google Scholar] [CrossRef]
- Pushchina, E.V.; Obukhov, D.K.; Varaksin, A.A. Features of adult neurogenesis and neurochemical signaling in the Cherry salmon Oncorhynchus masou brain. Neural Regen Res. 2013, 8, 13–23. [Google Scholar]
- Dong, Z.; Yang, N.; Yeo, S.Y.; Chitnis, A.; Guo, S. Intralineage directional notch signaling regulates self-renewal and differentiation of asymmetrically dividing radial glia. Neuron 2012, 74, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Kàlmàn, M. Astroglial architecture of the carp (Cyprinus carpio) brain as revealed by immunohistochemical staining against glial fibrillary acidic protein (GFAP). Anat. Embryol. 1998, 198, 409–433. [Google Scholar]
- Cuoghi, B.; Mola, L. Macroglial cells of the teleost central nervous system: A survey of the main types. Cell Tissue Res. 2009, 338, 319–332. [Google Scholar] [CrossRef]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.G.; Aigner, L. Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 2005, 21, 1–14. [Google Scholar] [CrossRef]
- Ninkovic, J.; Götz, M. Fate specification in the adult brain—Lessons for eliciting neurogenesis from glial cells. Bioessays 2013, 35, 242–252. [Google Scholar] [CrossRef]
- Baumgart, E.V.; Barbosa, J.S.; Bally-Cuif, L.; Götz, M.; Ninkovic, J. Stab wound injury of the zebrafish telencephalon: A model for comparative analysis of reactive gliosis. Glia 2012, 60, 343–357. [Google Scholar] [CrossRef]
- Vitalo, A.G.; Sîrbulescu, R.F.; Ilieş, I.; Zupanc, G.K. Absence of gliosis in a teleost model of spinal cord regeneration. J. Comp. Physiol. A 2016, 202, 445–456. [Google Scholar] [CrossRef]
- Dahl, D.; Rueger, D.C.; Bignami, A.; Weber, K.; Osborn, M. Vimentin, the 57,000 molecular weight protein of fibroblasts filaments, is the major cytoskeletal component in immature glia. Eur. J. Cell Biol. 1981, 24, 191–196. [Google Scholar]
- Schnitzer, J.; Franke, W.W.; Schachner, M. Immunocytochemical demonstration of vimentin in astrocytes and ependymal cells of developing and adult mouse nervous system. J. Cell Biol. 1981, 90, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Kàlmàn, M.; Ari, C. Distribution of GFAP immunoreactive structures in the rhombencephalon of the sterlet (Acipenser ruthenus) and its evolutionary implication. J. Exp. Zool. 2002, 293, 395–406. [Google Scholar] [CrossRef]
- Cerdà, J.; Conrad, M.; Markl, J.; Brand, M.; Herrmann, H. Zebrafish vimentin: Molecular characterization, assembly properties and developmental expression. Eur. J. Cell Biol. 1998, 77, 175–187. [Google Scholar] [CrossRef]
- Herrmann, H.; Munick, M.D.; Brettel, M.; Fouquet, B.; Markl, J. Vimentin in a cold-water fish, the rainbow trout: Highly conserved primary structure but unique assembly properties. J. Cell Sci. 1996, 109, 569–578. [Google Scholar]
- Zupanc, G.K.; Sîrbulescu, R.F. Teleost fish as a model system to study successful regeneration of the central nervous system. Curr. Top. Microbiol. Immunol. 2013, 367, 193–233. [Google Scholar]
- März, M.; Chapouton, P.; Diotel, N.; Vaillant, C.; Hesl, B.; Takamiya, M.; Lam, C.S.; Kah, O.; Bally-Cuif, L.; Strähle, U. Heterogeneity in progenitor cell subtypes in the ventricular zone of the zebrafish adult telencephalon. Glia 2010, 58, 870–888. [Google Scholar] [CrossRef]
- Takeda, A.; Atobe, Y.; Kadota, T.; Goris, R.C.; Funakoshi, K. Axonal regeneration through the fibrous scar in lesioned goldfish spinal cord. Neuroscience 2015, 284, 134–152. [Google Scholar] [CrossRef]
- Schaar, B.T.; Kinoshita, K.; Mcconnell, S.K. Doublecortin microtubule affinity is regulated by a balance of kinase and phosphatase activity at the leading edge of migrating neurons. Neuron 2004, 41, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Manger, P. Presence and Absence of Adult Hippocampal Neurogenesis in Mammals and the Effect of the Environment. In Proceedings of the Adult Neurogenesis: Evolution, Regulation and Function, Dresden, Germany, 6–8 May 2015. [Google Scholar]
- Mazengenya, P.; Manger, P.; Ihunwo, A. Adult Neurogenesis in the Subspecies of Domestic Pigeons Columba Livia Domesticus: The racing Homer and Utility Carneau Pigeons. In Proceedings of the Adult Neurogenesis: Evolution, Regulation and Function, Dresden, Germany, 6–8 May 2015. [Google Scholar]
- Couillard-Despres, S.; Winner, B.; Karl, C.; Lindemann, G.; Schmid, P.; Aigner, R.; Laemke, J.; Bogdahn, U.; Winkler, J.; Bischofberger, J.; et al. Targeted transgene expression in neuronal precursors: Watching young neurons in the old brain. Eur. J. Neurosci. 2006, 24, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.P.; Couillard-Després, S.; Cooper-Kuhn, C.M.; Winkler, J.; Aigner, L.; Kuhn, H.G. Transient expression of doublecortin during adult neurogenesis. J. Comp. Neurol. 2003, 467, 1–10. [Google Scholar] [CrossRef]
- Ambrogini, P.; Lattanzi, D.; Ciuffoli, S.; Agostini, D.; Bertini, L.; Stocchi, V.; Santi, S.; Cuppini, R. Morphofunctional characterization of neuronal cells at different stages of maturation in granule cell layer of adult rat dentate gyrus. Brain Res. 2004, 1017, 21–31. [Google Scholar] [CrossRef]
- Brandt, M.D.; Jessberger, S.; Steiner, B.; Kronenberg, G.; Reuter, K.; Bick-Sander, A.; von der Behrens, W.; Kempermann, G. Transient calretinin-expression defines early postmitotic step of neuronal differentiation in adult hippocampus neurogenesis of mice. Mol. Cell. Neurosci. 2003, 24, 603–613. [Google Scholar] [CrossRef]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional neurogenesis in the adult hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef]
- Kimbrel, E.A.; Kouris, N.A.; Yavanian, G.J.; Chu, J.; Qin, Y.; Chan, A.; Singh, R.P.; McCurdy, D.; Gordon, L.; Levinson, R.D.; et al. Mesenchymal stem cell population derived from human pluripotent stem cells displays potent immunomodulatory and therapeutic properties. Stem Cells Dev. 2014, 23, 1611–1624. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intact Animals | Damaged Telencephalon | ||||
|---|---|---|---|---|---|
| Brain Areas Cell Type | Cell/Nuclei Size, µm | Optical Density, UOD | Cell Type | Cell/Nuclei Size, µm | Optical Density, UOD |
| Dd Undifferentiated cells | 4.5 ± 0.5/2.8 ± 1.0 | 151.7 ± 6.2 | undifferent. oval | 4.3 ± 0.3/2.8 ± 0.4 5.6 ± 0.2/2.8 ± 1.0 | 177.1 ± 5.4 177.6 ± 3.6 |
| nuclei | 3.7 ± 0.2/2.3 ± 0.1 | 145.4 ± 2.3 | nuclei | 3.6 ± 0.3/2.5 ± 0.5 2.8 ± 0.2/2.0 ± 0.3 | 169.2 ± 5.1 168.4 ± 3.5 |
| Dl Undifferentiated cells | 4.6 ± 0.3/3.4 ± 0.5 | 147.8 ± 1.9 | undifferent. | 4.4 ± 0.3/2.7 ± 0.4 | 174.4 ± 5.1 |
| Oval | 5.6 ± 0.4/3.9 ± 0.4 7.6 ± 0.5/5.3 ± 0.9 | 151.9 ± 4.5 144.2 ± 3.3 | oval | 5.9 ± 0.5/4.3 ± 0.8 | 180.5 ± 3.0 |
| nuclei | 3.4 ± 0.4/2.7 ± 0.2 | 145.8 ± 4.6 | nuclei | 3.5 ± 0.3/2.4 ± 0.2 2.4 ± 0.3/1.9 ± 0.4 | 169.5 ± 1.2 165.7 ± 3.1 |
| Dm undifferentiated | 4.2 ± 0.3/2.8 ± 0.2 | 135.4 ± 2 | undifferent. | 4.4 ± 0.2/3.1 ± 0.1 | 152.6 ± 1.8 |
| oval | 5.2 ± 0.2/3.1 ± 0.6 7.2 ± 0.6/3.4 ± 0.5 | 152.2 ± 3.6 153.7 ± 1.1 | oval | - | - |
| nuclei | 3.4 ± 0.3/2.5 ± 0.6 | 144.9 ± 8.6 | nuclei | 3.4 ± 0.2/2.5 ± 0.3 2.7 ± 0.1/2.3 ± 0.3 | 155.3 ± 3.7 151.7 ± 4.8 |
| Intact Animals | Damaged Telencephalon | ||||||
|---|---|---|---|---|---|---|---|
| Cell Size, µm/Localization | Optical Density, UOD | DD | DL | DM | |||
| Cell Size, µm/Localization | Optical Density, UOD | Cell Size, µm/Localization | Optical Density, UOD | Cell Size, µm/Localization | Optical Density, UOD | ||
| GFAP | |||||||
| 10.3 ± 1.3/6.6 ± 1.0 (PVZ) 12.3 ± 0.3/7.9 ± 0.9 (PVZ) 13.4 ± 0.3/9.7 ± 1.0 (PVZ) | +++ +++ +++ | 14.2 ± 1.1/9.3 ± 0.2 (PVZ) 16.4 ± 0.8/8.5 ± 1.3 (PVZ) | +++ +++ | 5.1 ± 0.8/4.5 ± 1.0 (PZ) 9.7 ± 0.2/6.5 ± 0.2 * (PVZ) 11.4 ± 0.3/6.1 ± 1.2 (PVZ) 12.9 ± 0.6/7.1 ± 0.8 (PVZ) | +++ +++ +++ +++ | 9.7 ± 0.2/6.6 ± 0.6 (PVZ) 11.7 ± 1.0/7.1 ± 1.7 (PVZ) 14.8 ± 0.3/9.2 ± 1.1 (PVZ) | +++ +++ +++ |
| 10.8 ± 1.3/6.9 ± 0.2 (PVZ) 12.8 ± 0.2/8.5 ± 1.3 (PVZ) 14.5 ± 0.2/10.1 ± 1.4 (PVZ) | ++ ++ ++ | 11.9 ± 0.5/9.5 ± 0.3 (PVZ) 16.3 ± 0.2/9.7 ± 1.1 (SVZ) 18.8 ± 2.4/10.7 ± 2.1 (SVZ) | ++ ++ ++ | 7.4 ± 0.4/5.8 ± 0.3 (PVZ) 8.1 ± 0.2/5.7 ± 0.5 * (SVZ) 9.8 ± 0.6/5.8 ± 0.5 * (PVZ) 13.0 ± 2.4/6.2 ± 0.4 (PVZ) | ++ ++ ++ ++ | 12.5 ± 0.4/8.5 ± 1.4 (PVZ) 13.5 ± 0.3/8.3 ± 1.9 (PVZ) 14.7 ± 0.7/9.6 ± 1.8 (PVZ) | ++ ++ ++ |
| Vimentin | |||||||
| 9.2 ± 0.5/8.6 ± 0.4 (PVZ) 10.8 ± 0.6/7.4 ± 1.3 (PVZ) 13.2 ± 0.7/7.5 ± 1.7 (PVZ) | +++ ++ +++ | 3.2 ± 0.6/2.1 ± 0.3 (PZ) 8.6 ± 1.3/5.9 ± 0.9 * (PVZ) 10.5 ± 0.7/6.8 ± 1.3 (PVZ) | +++ +++ +++ | 5.5 ± 1.6/3.9 ± 1 (PZ) 11.2 ± 0.4/8.0 ± 0.3 (PVZ) 12.3 ± 0.3/8.2 ± 0.6 (SVZ) 17.3 ± 0.3/8.4 ± 1.0 (PVZ) | +++ +++ +++ +++ | 6.5 ± 0.5/4.0 ± 0.8 (PVZ) 9.3 ± 0.3/5.2 ± 1.1 (PVZ) | ++ ++ |
| 9.1 ± 0.4/7.8 ± 0.5 (PVZ) 10.6 ± 0.2/7.4 ± 0.7 (PVZ) 11.5 ± 0.5/7.7 ± 1.5 (PVZ) | ++ + + | 9.0 ± 0.7/6.5 ± 0.9 (PVZ) 11.7 ± 1.0/8.0 ± 0.5 (PVZ) | ++ ++ | 9.0 ± 0.2/6.6 ± 1.7 * (SVZ) 11.2 ± 0.6/7.3 ± 0.3 (PVZ) 13.1 ± 0.2/8.8 ± 1.1 (PVZ) | ++ ++ ++ | 5.9 ± 0.4/3.7 ± 0.4 (SVZ) 7.7 ± 0.5/6.0 ± 0.7 * (PVZ) 10.1 ± 0.8/6.5 ± 0.9 (SVZ) | + + + |
| Doublecortin | |||||||
| 8.6 ± 1.4/5.0 ± 0.9 (PVZ) 10.8 ± 0.5/6.3 ± 0.4 (PVZ) 13.0 ± 0.6/6.9 ± 2.4 (PVZ) | +++ ++ ++ | 4.6 ± 0.7/3.5 ± 1.1 (PZ) 14.3 ± 0.2/9.9 ± 0.9 (PVZ) 16.8 ± 0.5/10.9 ± 1.5 (PVZ) 21.6 ± 1.4/11.7 ± 2.2 (PVZ) | +++ +++ +++ +++ | 5.4 ± 1.3/4.2 ± 1.0 (PZ) 13.9 ± 1.2/10.1 ± 0.9 (PVZ) 16.2 ± 0.8/11.4 ± 0.8 (PVZ) | +++ +++ +++ | 5.2 ± 0.8/3.7 ± 0.6 (PZ) 9.4 ± 0.6/6.6 ± 0.8 (SVZ) 11.6 ± 0.8/6.7 ± 0.9 (PVZ) | +++ +++ +++ |
| 9.1 ± 1.2/6.9 ± 1.0 * (SVZ) 10.9 ± 0.3/6.6 ± 0.8 (PVZ) 12.3 ± 0.4/7.6 ± 1.6 (PVZ) 16.1 ± 0.8/9.5 ± 2.4 (PVZ) | ++ + + + | 12.0 ± 0.7/9.8 ± 1.8 (PVZ) 13.9 ± 0.4/10.1 ± 2.2 (SVZ) 15.5 ± 0.3/11.7 ± 1.8 (SVZ) 18.4 ± 0.8/13.4 ± 1.7 (SVZ) | ++ ++ ++ ++ | 14.3 ± 0.3/13.1 ± 2.0 (PVZ) 16.5 ± 0.5/11.8 ± 0.9 (SVZ) 19.0 ± 1.0/12.0 ± 0.6 (SVZ) | ++ ++ ++ | 10.3 ± 0.8/8.7 ± 1.5 (SVZ) 12.1 ± 0.4/8.1 ± 1.2 (SVZ) 15.3 ± 0.9/8.1 ± 1.5 (PVZ) | ++ ++ ++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pushchina, E.V.; Zharikova, E.I.; Varaksin, A.A.; Prudnikov, I.M.; Tsyvkin, V.N. Proliferation, Adult Neuronal Stem Cells and Cells Migration in Pallium during Constitutive Neurogenesis and after Traumatic Injury of Telencephalon of Juvenile Masu Salmon, Oncorhynchus masou. Brain Sci. 2020, 10, 222. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10040222
Pushchina EV, Zharikova EI, Varaksin AA, Prudnikov IM, Tsyvkin VN. Proliferation, Adult Neuronal Stem Cells and Cells Migration in Pallium during Constitutive Neurogenesis and after Traumatic Injury of Telencephalon of Juvenile Masu Salmon, Oncorhynchus masou. Brain Sciences. 2020; 10(4):222. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10040222
Chicago/Turabian StylePushchina, Evgeniya V., Eva I. Zharikova, Anatoly A. Varaksin, Igor M. Prudnikov, and Vladimir N. Tsyvkin. 2020. "Proliferation, Adult Neuronal Stem Cells and Cells Migration in Pallium during Constitutive Neurogenesis and after Traumatic Injury of Telencephalon of Juvenile Masu Salmon, Oncorhynchus masou" Brain Sciences 10, no. 4: 222. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10040222