The Temporal and Spatial Dynamics of Cortical Emotion Processing in Different Brain Frequencies as Assessed Using the Cluster-Based Permutation Test: An MEG Study

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
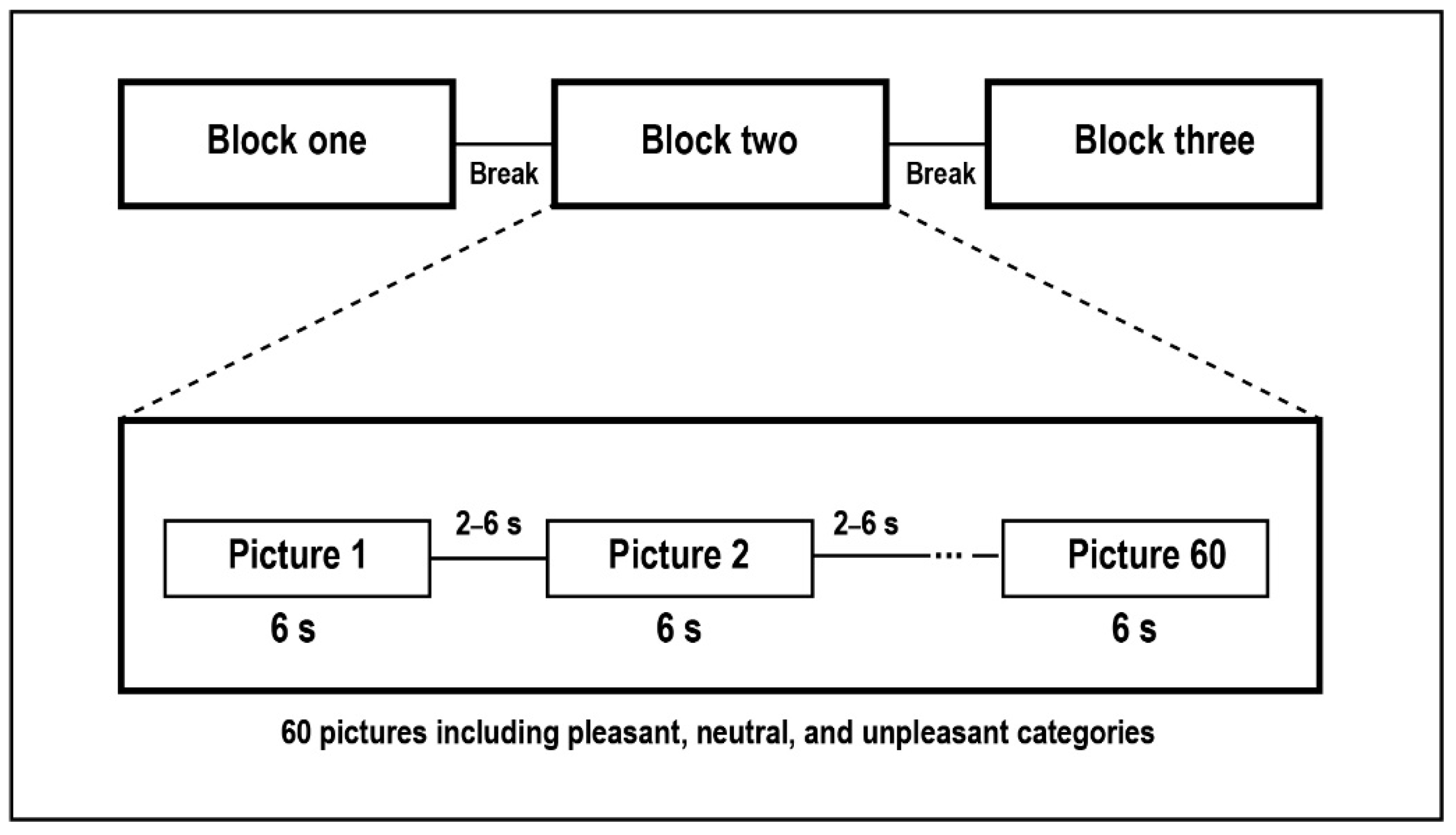
2.2. Stimuli and Design
2.3. Data Acquisition and Analysis
2.4. Time Intervals of Interest
2.5. Applying the Cluster-Based Permutation Test (CBPT)
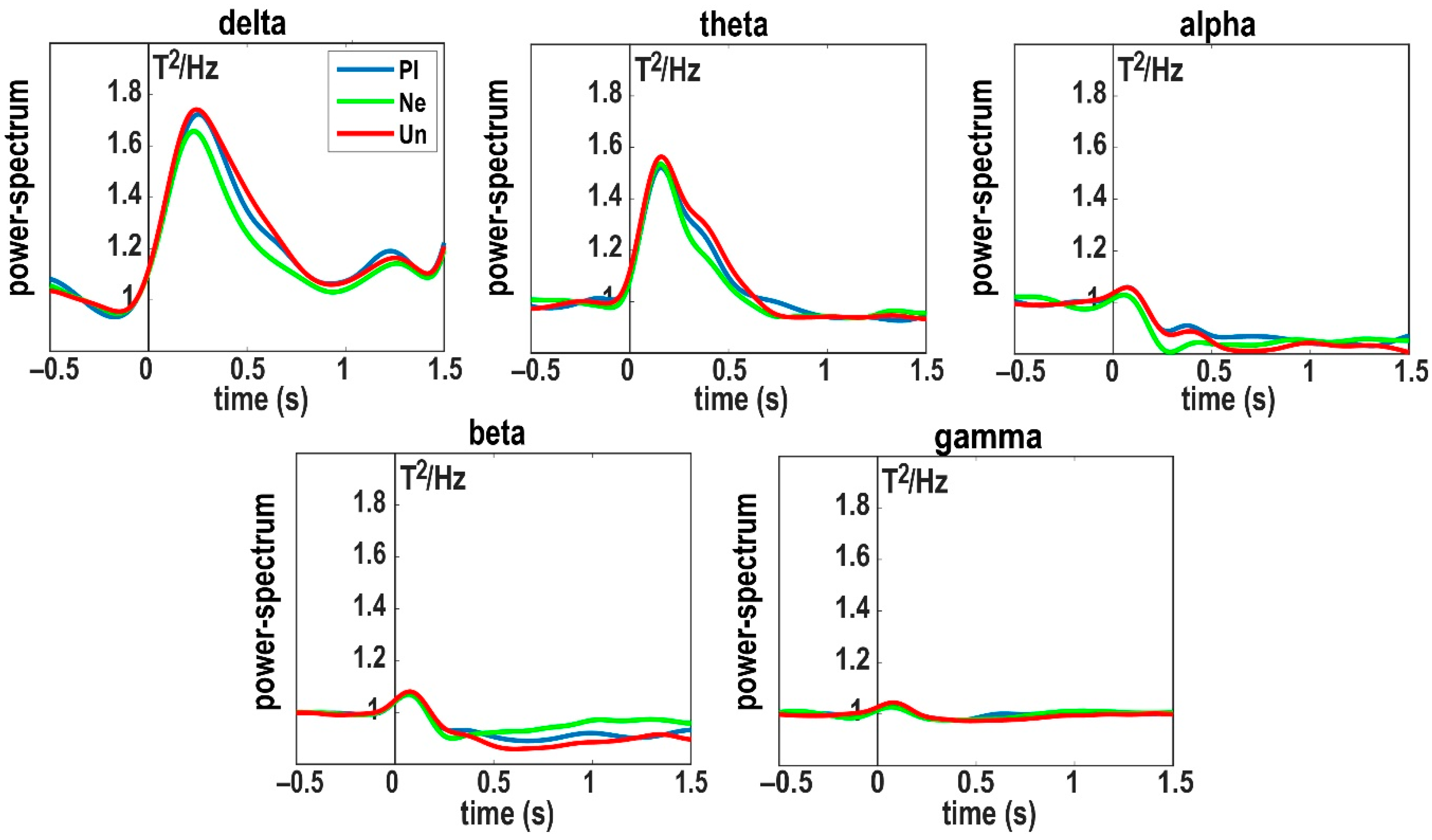
3. Results
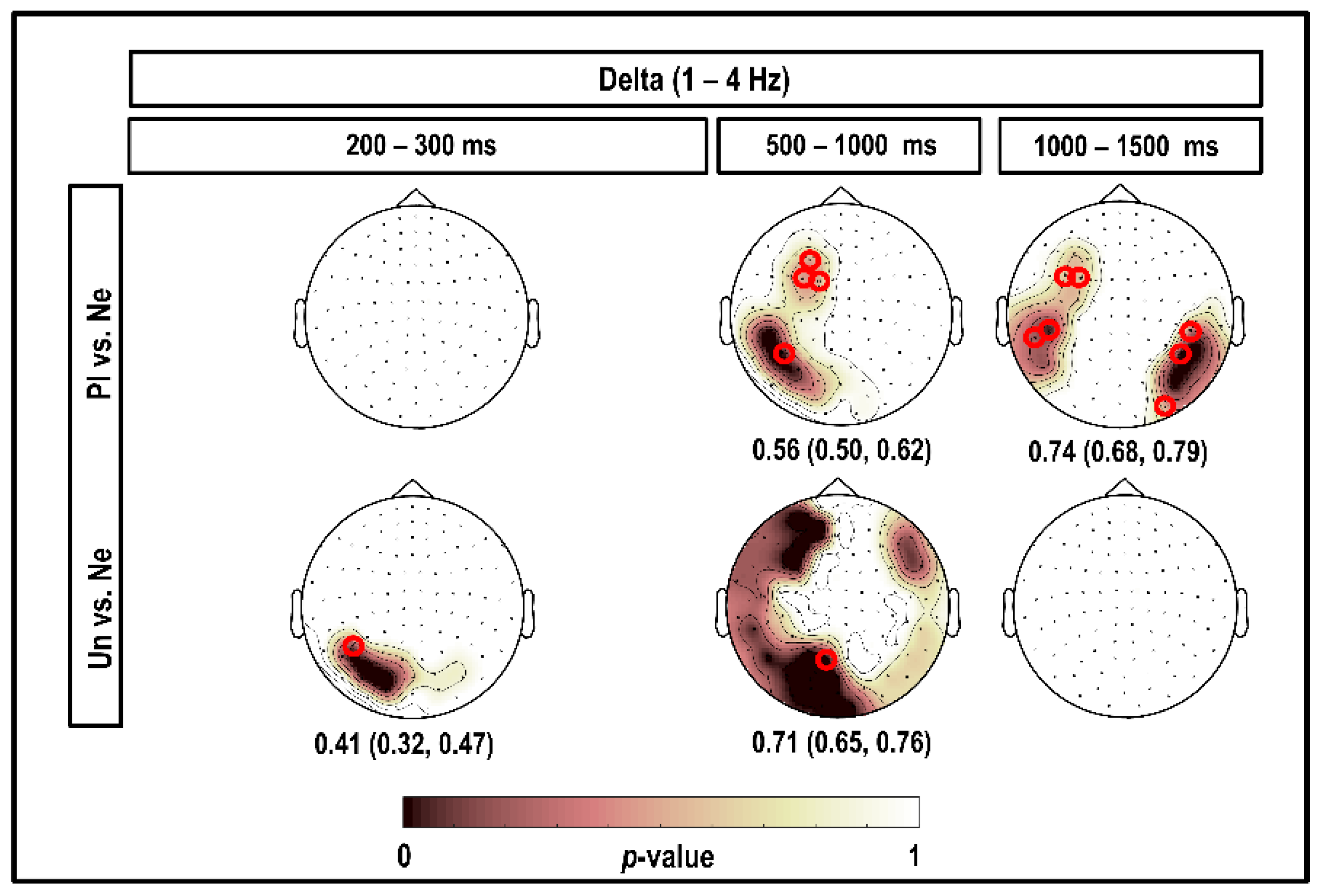
3.1. Delta Band (1–4 Hz)
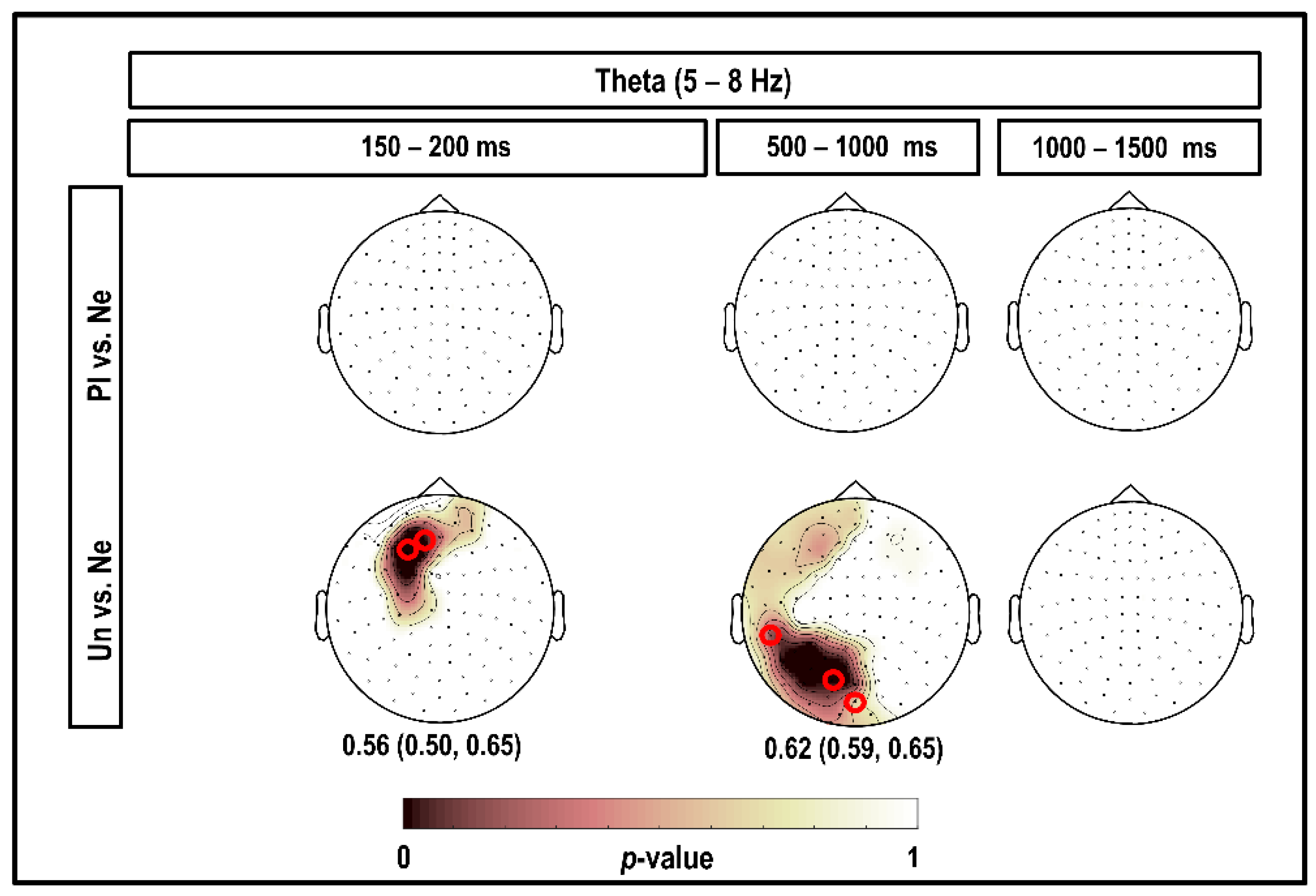
3.2. Theta Band (5–8 Hz)
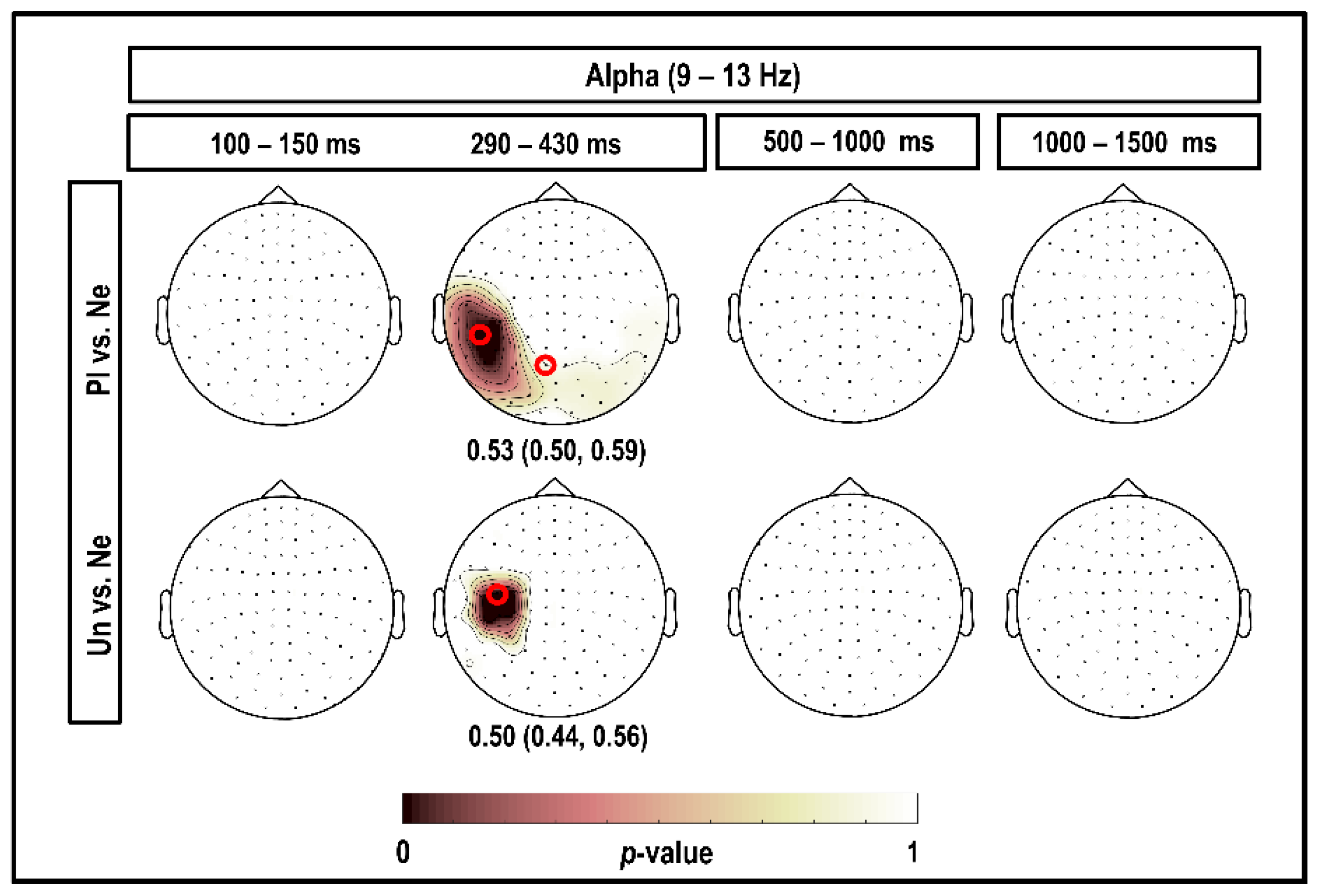
3.3. Alpha Band (9–13 Hz)
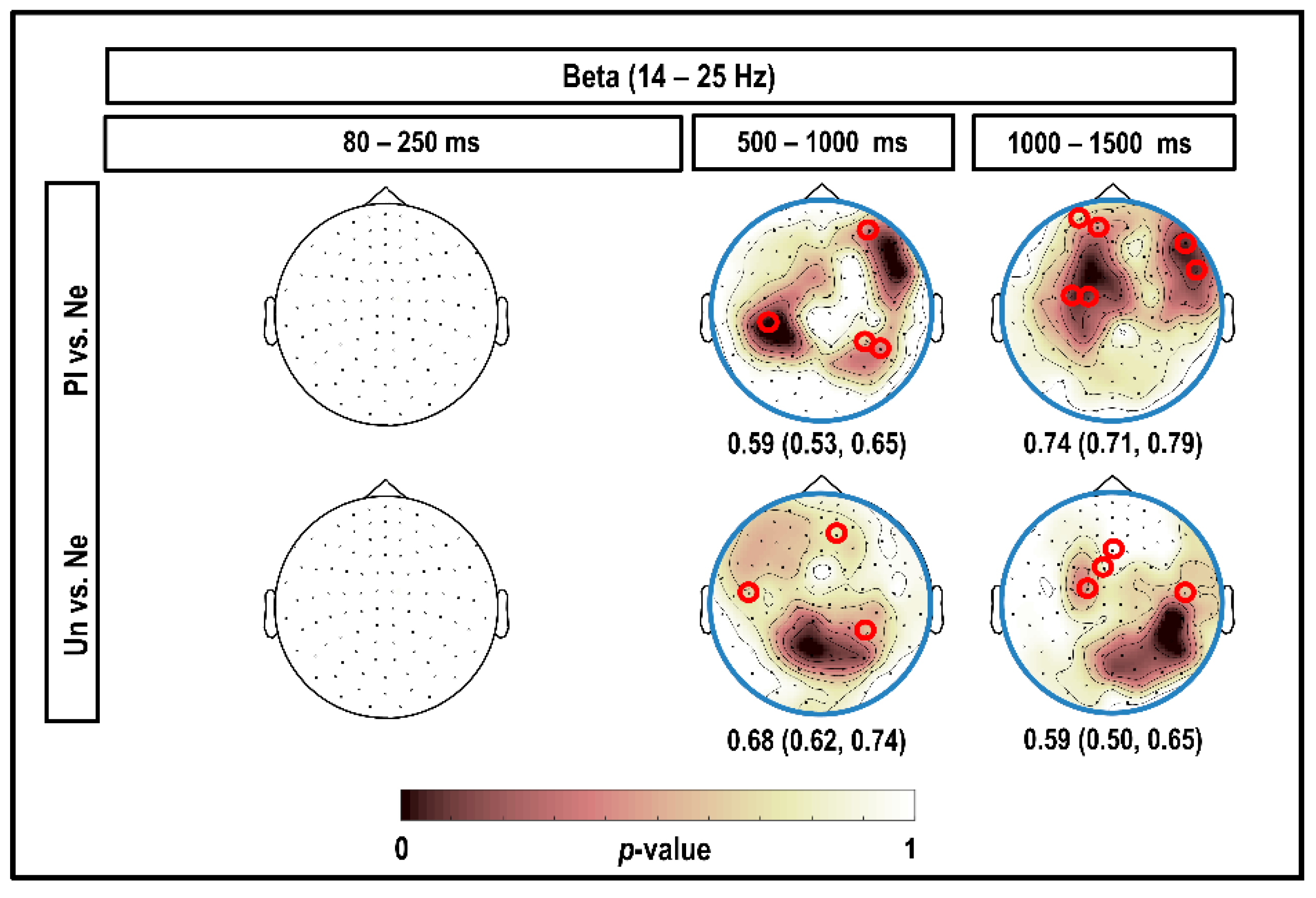
3.4. Beta Band (14–25 Hz)
3.5. Gamma Band (26–45 Hz)
4. Discussion
4.1. Delta
4.2. Theta
4.3. Alpha
4.4. Beta
4.5. Gamma
5. Future Work
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2010, 2011, 1–9. [Google Scholar] [CrossRef]
- Sassenhagen, J.; Draschkow, D. Cluster-based permutation tests of MEG/EEG data do not establish significance of effect latency or location. Psychophysiology 2019, 56. [Google Scholar] [CrossRef]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Frömer, R.; Maier, M.; Rahman, R.A. Group-level EEG-processing pipeline for flexible single trial-based analyses including linear mixed models. Front. Mol. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Maris, E. Randomization tests for ERP topographies and whole spatiotemporal data matrices. Psychophysiology 2004, 41, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Maris, E. Statistical testing in electrophysiological studies. Psychophysiology 2011, 49, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Pratt, M.; Goldstein, A.; Feldman, R. Child brain exhibits a multi-rhythmic response to attachment cues. Soc. Cogn. Affect. Neurosci. 2018, 13, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Styliadis, C.; Ioannides, A.A.; Bamidis, P.D.; Papadelis, C. Mapping the spatiotemporal evolution of emotional processing: An MEG study across arousal and valence dimensions. Front. Hum. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Jabbi, M.; Kohn, P.D.; Nash, T.; Ianni, A.; Coutlee, C.; Holroyd, T.; Carver, F.W.; Chen, Q.; Cropp, B.; Kippenhan, J.S.; et al. Convergent BOLD and beta-band activity in superior temporal sulcus and frontolimbic circuitry underpins human emotion cognition. Cereb. Cortex 2014, 25, 1878–1888. [Google Scholar] [CrossRef]
- Grootswagers, T.; Kennedy, B.L.; Most, S.B.; Carlson, T.A. Neural signatures of dynamic emotion constructs in the human brain. Neuropsychologia 2017. [Google Scholar] [CrossRef]
- Huang, G.; Zhang, Z. Improving sensitivity of cluster-based permutation test for EEG/MEG data. In Proceedings of the 2017 8th International IEEE/EMBS Conference on Neural Engineering (NER), Shanghai, China, 25–28 May 2017. [Google Scholar]
- Pernet, C.; Latinus, M.; Nichols, T.; Rousselet, G. Cluster-based computational methods for mass univariate analyses of event-related brain potentials/fields: A simulation study. J. Neurosci. Methods 2014, 250, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piai, V.; Dahlslätt, K.; Maris, E. Statistically comparing EEG/MEG waveforms through successive significant univariate tests: How bad can it be? Psychophysiology 2014, 52, 440–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urigüen, J.A.; Garcia-Zapirain, B.; Artieda, J.; Iriarte, J.; Valencia, M. Comparison of background EEG activity of different groups of patients with idiopathic epilepsy using Shannon spectral entropy and cluster-based permutation statistical testing. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jessen, S.; Kotz, S. The temporal dynamics of processing emotions from vocal, facial, and bodily expressions. NeuroImage 2011, 58, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Bublatzky, F.; Kavcıoğlu, F.; Guerra, P.; Doll, S.; Junghöfer, M. Contextual information resolves uncertainty about ambiguous facial emotions: Behavioral and magnetoencephalographic correlates. NeuroImage 2020. [Google Scholar] [CrossRef]
- Esslen, M.; Pascual-Marqui, R.D.; Hell, D.; Kochi, K.; Lehmann, D. Brain areas and time course of emotional processing. NeuroImage 2004, 21, 1189–1203. [Google Scholar] [CrossRef]
- Giorgetta, C.; Grecucci, A.; Bonini, N.; Coricelli, G.; Demarchi, G.; Braun, C.; Sanfey, A.G. Waves of regret: A meg study of emotion and decision-making. Neuropsychologia 2013, 51, 38–51. [Google Scholar] [CrossRef]
- Kheirkhah, M.; Brodoehl, S.; Leistritz, L.; Götz, T.; Baumbach, P.; Huonker, R.; Witte, O.W.; Volk, G.F.; Guntinas-Lichius, O.; Klingner, C.; et al. Abnormal emotional processing and emotional experience in patients with peripheral facial nerve paralysis: An MEG study. Brain Sci. 2020, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Li, H.; Luo, G.; Wang, Y.; Tang, H.; Han, L.; Yao, Z. Impaired prefrontal-amygdala effective connectivity is responsible for the dysfunction of emotion process in major depressive disorder: A dynamic causal modeling study on MEG. Neurosci. Lett. 2012, 523, 125–130. [Google Scholar] [CrossRef]
- Peyk, P.; Schupp, H.T.; Elbert, T.; Junghöfer, M. Emotion processing in the visual brain: A MEG analysis. Brain Topogr. 2008, 20, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Abadi, M.K.; Subramanian, R.; Kia, S.M.; Avesani, P.; Patras, I.; Sebe, N. DECAF: MEG-based multimodal database for decoding affective physiological responses. IEEE Trans. Affect. Comput. 2015, 6, 209–222. [Google Scholar] [CrossRef]
- Güntekin, B.; Başar, E. Facial affect manifested by multiple oscillations. Int. J. Psychophysiol. 2009, 71, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Güntekin, B.; Başar, E. A review of brain oscillations in perception of faces and emotional pictures. Neuropsychologia 2014, 58, 33–51. [Google Scholar] [CrossRef]
- Keil, A.; Müller, M.M.; Gruber, T.; Wienbruch, C.; Stolarova, M.; Elbert, T. Effects of emotional arousal in the cerebral hemispheres: A study of oscillatory brain activity and event-related potentials. Clin. Neurophysiol. 2001, 112, 2057–2068. [Google Scholar] [CrossRef] [Green Version]
- Miskovic, V.; Schmidt, L.A. Cross-regional cortical synchronization during affective image viewing. Brain Res. 2010, 1362, 102–111. [Google Scholar] [CrossRef]
- Sakihara, K.; Gunji, A.; Furushima, W.; Inagaki, M. Event-related oscillations in structural and semantic encoding of faces. Clin. Neurophysiol. 2012, 123, 270–277. [Google Scholar] [CrossRef]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Technical Manual and Affective Ratings; NIMH Center for the Study of Emotion and Attention: Gainesville, FL, USA, 1997; pp. 39–58. [Google Scholar]
- Taulu, S.; Simola, J. Spatiotemporal signal space separation method for rejecting nearby interference in MEG measurements. Phys. Med. Biol. 2006, 51, 1759–1768. [Google Scholar] [CrossRef]
- Taulu, S.; Simola, J.; Kajola, M. Applications of the signal space separation method. IEEE Trans. Signal. Process. 2005, 53, 3359–3372. [Google Scholar] [CrossRef] [Green Version]
- Garces, P.; López-Sanz, D.; Maestú, F.; Pereda, E. Choice of magnetometers and gradiometers after signal space separation. Sensors 2017, 17, 2926. [Google Scholar] [CrossRef] [Green Version]
- García, J.P.; Garcés, P.; Del Río, D.; Maestú, F. Tracking the effect of emotional distraction in working memory brain networks: Evidence from an MEG study. Psychophysiology 2017, 54, 1726–1740. [Google Scholar] [CrossRef]
- Popov, T.; Oostenveld, R.; Schoffelen, J.M. FieldTrip made easy: An analysis protocol for group analysis of the auditory steady state brain response in time, frequency, and space. Front. Mol. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FieldTriptoolbox. Cluster-Based Permutation Tests on Time-Frequency Data. Available online: http://www.fieldtriptoolbox.org/tutorial/cluster_permutation_freq/ (accessed on 28 May 2020).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Benjamini, Y.; Yekutieli, D. False discovery rate—Adjusted multiple confidence intervals for selected parameters. J. Am. Stat. Assoc. 2005, 100, 71–81. [Google Scholar] [CrossRef]
- David Groppe. fdr_bh. MATLAB Central File Exchange. 2020. Available online: https://www.mathworks.com/matlabcentral/fileexchange/27418-fdr_bh) (accessed on 6 May 2020).
- Tibshirani, R. Regression shrinkage and selection via the lasso. J. R. Stat. Soc. Ser. Methodol. 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Balconi, M.; Brambilla, E.; Falbo, L. BIS/BAS, cortical oscillations and coherence in response to emotional cues. Brain Res. Bull. 2009, 80, 151–157. [Google Scholar] [CrossRef]
- Balconi, M.; Mazzà, G. Brain oscillations and BIS/BAS (behavioral inhibition/activation system) effects on processing masked emotional cues. Int. J. Psychophysiol. 2009, 74, 158–165. [Google Scholar] [CrossRef]
- Klados, M.; Frantzidis, C.; Vivas, A.B.; Papadelis, C.; Lithari, C.; Pappas, C.; Bamidis, P.D. A framework combining delta Event-Related Oscillations (EROs) and synchronisation effects (ERD/ERS) to study emotional processing. Comput. Intell. Neurosci. 2009, 2009, 1–16. [Google Scholar] [CrossRef]
- Aftanas, L.I.; Varlamov, A.A.; Pavlov, S.V.; Makhnev, V.P.; Reva, N.V. Time-dependent cortical asymmetries induced by emotional arousal: EEG analysis of event-related synchronization and desynchronization in individually defined frequency bands. Int. J. Psychophysiol. 2002, 44, 67–82. [Google Scholar] [CrossRef]
- Balconi, M.; Brambilla, E.; Falbo, L. Appetitive vs. defensive responses to emotional cues. Autonomic measures and brain oscillation modulation. Brain Res. 2009, 1296, 72–84. [Google Scholar] [CrossRef]
- Westphal, K.P.; Grözinger, B.; Diekmann, V.; Scherb, W.; Rees, J.; Leibing, U.; Kornhuber, H.H. Slower theta activity over the midfrontal cortex in schizophrenic patients. Acta Psychiatr. Scand. 1990, 81, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Aftanas, L.I.; Varlamov, A.; Pavlov, S.; Makhnev, V.; Reva, N. Affective picture processing: Event-related synchronization within individually defined human theta band is modulated by valence dimension. Neurosci. Lett. 2001, 303, 115–118. [Google Scholar] [CrossRef]
- Başar, E.; Güntekin, B.; Oniz, A. Principles of oscillatory brain dynamics and a treatise of recognition of faces and facial expressions. Sleep Deprivation Cogn. 2006, 159, 43–62. [Google Scholar] [CrossRef]
- Sun, J.; Sun, B.; Wang, B.; Gong, H. The processing bias for threatening cues revealed by event-related potential and event-related oscillation analyses. Neurosci. 2012, 203, 91–98. [Google Scholar] [CrossRef]
- Güntekin, B.; Başar, E. Emotional face expressions are differentiated with brain oscillations. Int. J. Psychophysiol. 2007, 64, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Güntekin, B.; Başar, E. Gender differences influence brain’s beta oscillatory responses in recognition of facial expressions. Neurosci. Lett. 2007, 424, 94–99. [Google Scholar] [CrossRef]
- Engel, A.K.; Fries, P. Beta-band oscillations—Signalling the status quo? Curr. Opin. Neurobiol. 2010, 20, 156–165. [Google Scholar] [CrossRef]
- Martini, N.; Menicucci, D.; Sebastiani, L.; Bedini, R.; Pingitore, A.; Vanello, N.; Milanesi, M.; Landini, L.; Gemignani, A. The dynamics of EEG gamma responses to unpleasant visual stimuli: From local activity to functional connectivity. NeuroImage 2012, 60, 922–932. [Google Scholar] [CrossRef]
- Müller, M.M.; Keil, A.; Gruber, T.; Elbert, T. Processing of affective pictures modulates right-hemispheric gamma band EEG activity. Clin. Neurophysiol. 1999, 110, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Rouw, R.; Kosslyn, S.M.; Hamel, R. Detecting high-level and low-level properties in visual images and visual percepts. Cognition 1997, 63, 209–226. [Google Scholar] [CrossRef]
- Miskovic, V.; Martinovic, J.; Wieser, M.; Petro, N.M.; Bradley, M.M.; Keil, A. Electrocortical amplification for emotionally arousing natural scenes: The contribution of luminance and chromatic visual channels. Biol. Psychol. 2015, 106, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindler, S.; Schettino, A.; Pourtois, G. Electrophysiological correlates of the interplay between low-level visual features and emotional content during word reading. Sci. Rep. 2018, 8, 12228. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kheirkhah, M.; Baumbach, P.; Leistritz, L.; Brodoehl, S.; Götz, T.; Huonker, R.; Witte, O.W.; Klingner, C.M. The Temporal and Spatial Dynamics of Cortical Emotion Processing in Different Brain Frequencies as Assessed Using the Cluster-Based Permutation Test: An MEG Study. Brain Sci. 2020, 10, 352. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060352
Kheirkhah M, Baumbach P, Leistritz L, Brodoehl S, Götz T, Huonker R, Witte OW, Klingner CM. The Temporal and Spatial Dynamics of Cortical Emotion Processing in Different Brain Frequencies as Assessed Using the Cluster-Based Permutation Test: An MEG Study. Brain Sciences. 2020; 10(6):352. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060352
Chicago/Turabian StyleKheirkhah, Mina, Philipp Baumbach, Lutz Leistritz, Stefan Brodoehl, Theresa Götz, Ralph Huonker, Otto W. Witte, and Carsten M. Klingner. 2020. "The Temporal and Spatial Dynamics of Cortical Emotion Processing in Different Brain Frequencies as Assessed Using the Cluster-Based Permutation Test: An MEG Study" Brain Sciences 10, no. 6: 352. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060352