
EEG Oscillations in Specific Frequency Bands Are Differently Coupled with Angular Joint Angle Kinematics during Rhythmic Passive Elbow Movement

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Hotspot Detection
2.3. Electroencephalography
2.4. Passive Movement
2.5. Data Analysis
3. Results
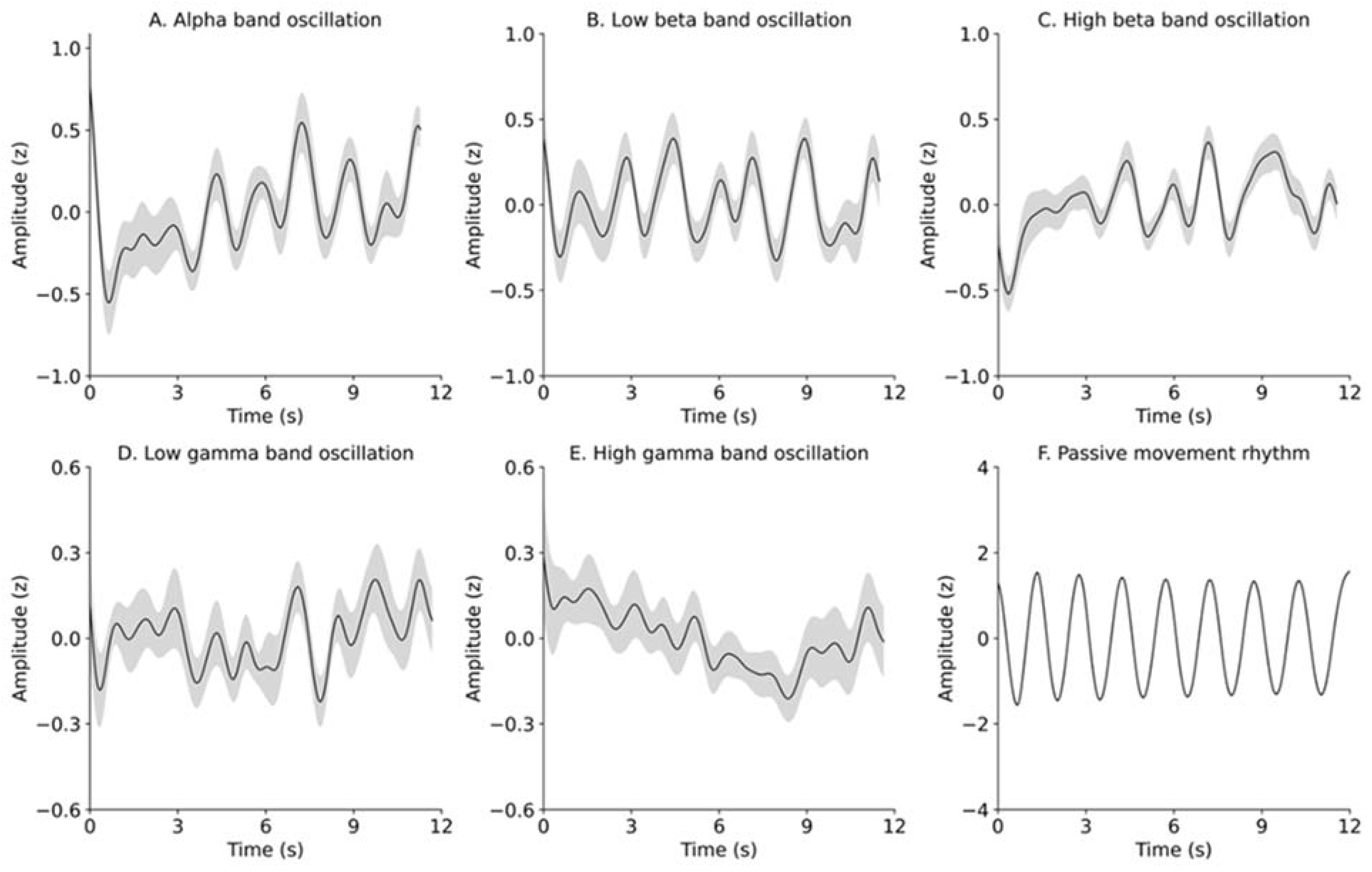
3.1. Consistency in Passive Movement Rhythms
3.2. Time Course of Changes in the EEG Power Spectrum and Elbow Angle
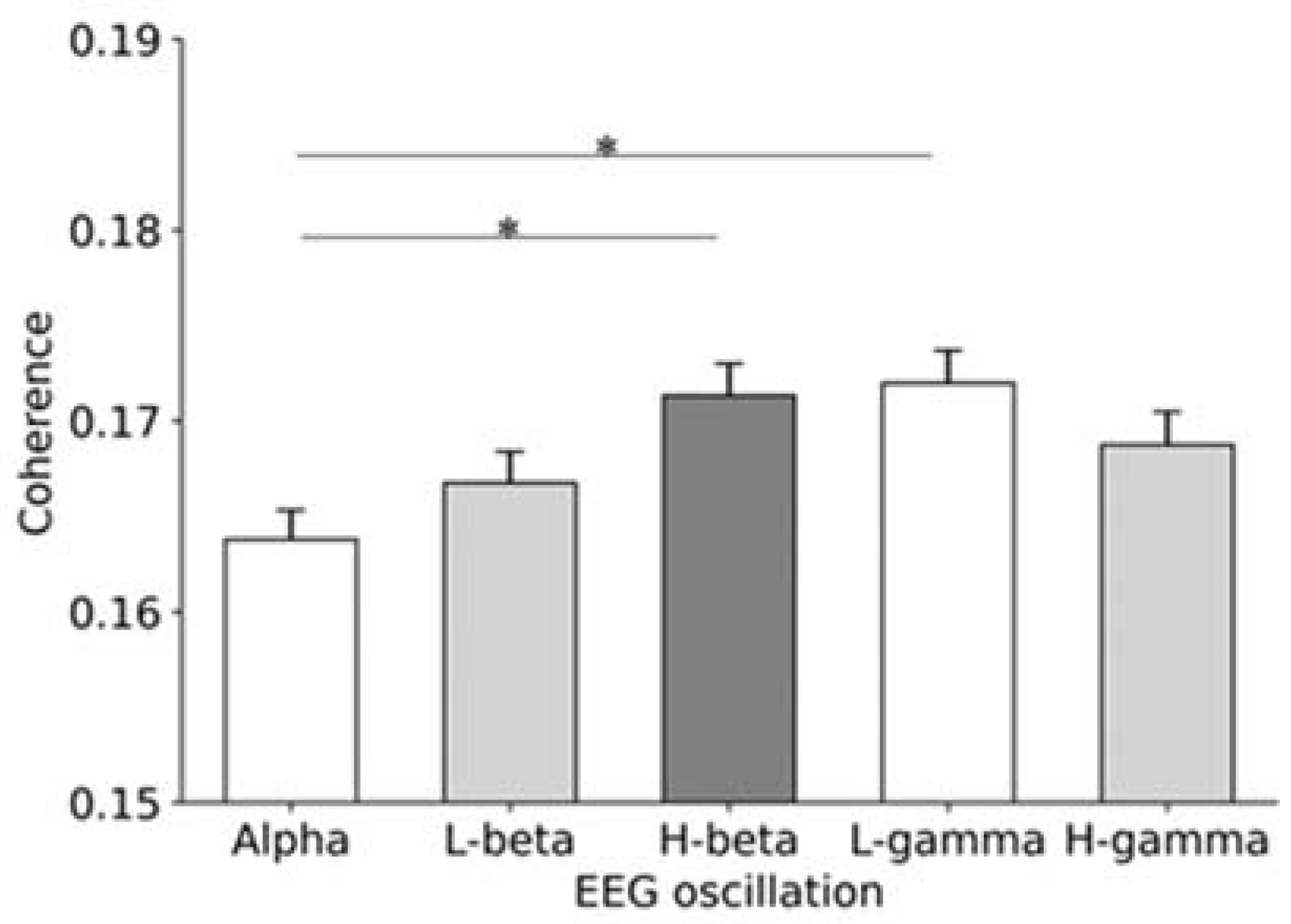
3.3. Coherence between EEG Oscillations and Passive Movement Rhythm
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosseini, Z.S.; Peyrovi, H.; Gohari, M. The Effect of Early Passive Range of Motion Exercise on Motor Function of People with Stroke: A Randomized Controlled Trial. J. Caring Sci. 2019, 8, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Nuyens, G.E.; De Weerdt, W.J.; Spaepen, A.J., Jr.; Kiekens, C.; Feys, H.M. Reduction of spastic hypertonia during repeated passive knee movements in stroke patients. Arch. Phys. Med. Rehabil. 2002, 83, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, Y.Y.; Wang, C.W.; Cheng, H.Y. Effects of active fatiguing movement versus passive repetitive movement on knee proprioception. Clin. Biomech. 2010, 25, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Mima, T.; Sadato, N.; Yazawa, S.; Hanakawa, T.; Fukuyama, H.; Yonekura, Y.; Shibasaki, H. Brain structures related to active and passive finger movements in man. Brain 1999, 122 Pt 10, 1989–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeber, M.; Scherer, R.; Muller-Putz, G.R. EEG Oscillations Are Modulated in Different Behavior-Related Networks during Rhythmic Finger Movements. J. Neurosci. 2016, 36, 11671–11681. [Google Scholar] [CrossRef] [Green Version]
- Pfurtscheller, G.; Lopes da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Formaggio, E.; Storti, S.F.; Boscolo Galazzo, I.; Gandolfi, M.; Geroin, C.; Smania, N.; Fiaschi, A.; Manganotti, P. Time-frequency modulation of ERD and EEG coherence in robot-assisted hand performance. Brain Topogr. 2015, 28, 352–363. [Google Scholar] [CrossRef]
- Alegre, M.; de Gurtubay, I.G.; Labarga, A.; Iriarte, J.; Malanda, A.; Artieda, J. Alpha and beta oscillatory activity during a sequence of two movements. Clin. Neurophysiol. 2004, 115, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Murguialday, A.; Birbaumer, N. Brain oscillatory signatures of motor tasks. J. Neurophysiol. 2015, 113, 3663–3682. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.J.; Leuthardt, E.C.; Schalk, G.; Rao, R.P.; Anderson, N.R.; Moran, D.W.; Miller, J.W.; Ojemann, J.G. Spectral changes in cortical surface potentials during motor movement. J. Neurosci. 2007, 27, 2424–2432. [Google Scholar] [CrossRef]
- Cassim, F.; Monaca, C.; Szurhaj, W.; Bourriez, J.L.; Defebvre, L.; Derambure, P.; Guieu, J.D. Does post-movement beta synchronization reflect an idling motor cortex? Neuroreport 2001, 12, 3859–3863. [Google Scholar] [CrossRef] [PubMed]
- Onishi, H.; Sugawara, K.; Yamashiro, K.; Sato, D.; Suzuki, M.; Kirimoto, H.; Tamaki, H.; Murakami, H.; Kameyama, S. Neuromagnetic activation following active and passive finger movements. Brain Behav. 2013, 3, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Radovanovic, S.; Korotkov, A.; Ljubisavljevic, M.; Lyskov, E.; Thunberg, J.; Kataeva, G.; Danko, S.; Roudas, M.; Pakhomov, S.; Medvedev, S.; et al. Comparison of brain activity during different types of proprioceptive inputs: A positron emission tomography study. Exp. Brain Res. 2002, 143, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Grezes, J.; Decety, J. Functional anatomy of execution, mental simulation, observation, and verb generation of actions: A meta-analysis. Hum. Brain Mapp. 2001, 12, 1–19. [Google Scholar] [CrossRef]
- Jackson, P.L.; Lafleur, M.F.; Malouin, F.; Richards, C.L.; Doyon, J. Functional cerebral reorganization following motor sequence learning through mental practice with motor imagery. NeuroImage 2003, 20, 1171–1180. [Google Scholar] [CrossRef]
- Kasess, C.H.; Windischberger, C.; Cunnington, R.; Lanzenberger, R.; Pezawas, L.; Moser, E. The suppressive influence of SMA on M1 in motor imagery revealed by fMRI and dynamic causal modeling. NeuroImage 2008, 40, 828–837. [Google Scholar] [CrossRef]
- Lacourse, M.G.; Orr, E.L.; Cramer, S.C.; Cohen, M.J. Brain activation during execution and motor imagery of novel and skilled sequential hand movements. NeuroImage 2005, 27, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Alegre, M.; Labarga, A.; Gurtubay, I.G.; Iriarte, J.; Malanda, A.; Artieda, J. Beta electroencephalograph changes during passive movements: Sensory afferences contribute to beta event-related desynchronization in humans. Neurosci. Lett. 2002, 331, 29–32. [Google Scholar] [CrossRef]
- Keinrath, C.; Wriessnegger, S.; Muller-Putz, G.R.; Pfurtscheller, G. Post-movement beta synchronization after kinesthetic illusion, active and passive movements. Int. J. Psychophysiol. 2006, 62, 321–327. [Google Scholar] [CrossRef]
- Hulley, S.B.; Cummings, S.R. Designing Clinical Research; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1988. [Google Scholar]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Cohen, M.X. A better way to define and describe Morlet wavelets for time-frequency analysis. NeuroImage 2019, 199, 81–86. [Google Scholar] [CrossRef]
- van der Spoel, E.; Choi, J.; Roelfsema, F.; Cessie, S.L.; van Heemst, D.; Dekkers, O.M. Comparing Methods for Measurement Error Detection in Serial 24-h Hormonal Data. J. Biol. Rhythm. 2019, 34, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, M.; Piitulainen, H.; De Tiege, X.; Jousmaki, V.; Hari, R. Corticokinematic coherence mainly reflects movement-induced proprioceptive feedback. NeuroImage 2015, 106, 382–390. [Google Scholar] [CrossRef]
- Parkkonen, E.; Laaksonen, K.; Piitulainen, H.; Pekkola, J.; Parkkonen, L.; Tatlisumak, T.; Forss, N. Strength of ~20-Hz Rebound and Motor Recovery After Stroke. Neurorehabil. Neural Repair. 2017, 31, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Tacchino, G.; Gandolla, M.; Coelli, S.; Barbieri, R.; Pedrocchi, A.; Bianchi, A.M. EEG Analysis During Active and Assisted Repetitive Movements: Evidence for Differences in Neural Engagement. IEEE Trans. Neural Syst. Rehabil. Eng. 2017, 25, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Reimer, J.; Penn, R.; Ojakangas, C.L.; Hatsopoulos, N.G. Fast and slow oscillations in human primary motor cortex predict oncoming behaviorally relevant cues. Neuron 2010, 65, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, T.; Trainor, L.J.; Large, E.W.; Ross, B. Internalized timing of isochronous sounds is represented in neuromagnetic beta oscillations. J. Neurosci. 2012, 32, 1791–1802. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.J.; Zanos, S.; Fetz, E.E.; den Nijs, M.; Ojemann, J.G. Decoupling the cortical power spectrum reveals real-time representation of individual finger movements in humans. J. Neurosci. 2009, 29, 3132–3137. [Google Scholar] [CrossRef]
- Gross, J.; Timmermann, L.; Kujala, J.; Dirks, M.; Schmitz, F.; Salmelin, R.; Schnitzler, A. The neural basis of intermittent motor control in humans. Proc. Natl. Acad. Sci. USA 2002, 99, 2299–2302. [Google Scholar] [CrossRef] [Green Version]
- Pollok, B.; Gross, J.; Dirks, M.; Timmermann, L.; Schnitzler, A. The cerebral oscillatory network of voluntary tremor. J. Physiol. 2004, 554, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Seeber, M.; Scherer, R.; Wagner, J.; Solis-Escalante, T.; Muller-Putz, G.R. High and low gamma EEG oscillations in central sensorimotor areas are conversely modulated during the human gait cycle. NeuroImage 2015, 112, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Seeber, M.; Scherer, R.; Wagner, J.; Solis-Escalante, T.; Muller-Putz, G.R. EEG beta suppression and low gamma modulation are different elements of human upright walking. Front. Hum. Neurosci. 2014, 8, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Suzuki, M.; Cho, K.; Iso, N.; Okabe, T.; Hamaguchi, T.; Yamamoto, J.; Kanemura, N. EEG Oscillations in Specific Frequency Bands Are Differently Coupled with Angular Joint Angle Kinematics during Rhythmic Passive Elbow Movement. Brain Sci. 2022, 12, 647. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050647
Suzuki T, Suzuki M, Cho K, Iso N, Okabe T, Hamaguchi T, Yamamoto J, Kanemura N. EEG Oscillations in Specific Frequency Bands Are Differently Coupled with Angular Joint Angle Kinematics during Rhythmic Passive Elbow Movement. Brain Sciences. 2022; 12(5):647. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050647
Chicago/Turabian StyleSuzuki, Takako, Makoto Suzuki, Kilchoon Cho, Naoki Iso, Takuhiro Okabe, Toyohiro Hamaguchi, Junichi Yamamoto, and Naohiko Kanemura. 2022. "EEG Oscillations in Specific Frequency Bands Are Differently Coupled with Angular Joint Angle Kinematics during Rhythmic Passive Elbow Movement" Brain Sciences 12, no. 5: 647. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050647