Vacuole and Mitochondria Patch (vCLAMP) Protein Vam6 Is Involved in Maintenance of Mitochondrial and Vacuolar Functions under Oxidative Stress in Candida albicans

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids Construction
2.2. Fluorescence Microscopy
2.3. Flow Cytometry
2.4. Electron Microscopic Observation
2.5. Determination of Intracellular ATP, Reactive Oxygen Species (ROS) and SOD1 Levels
2.6. Hyphal Induction
2.7. Assay of Macrophage-Killing Capacity
2.8. Virulence Assay
3. Results
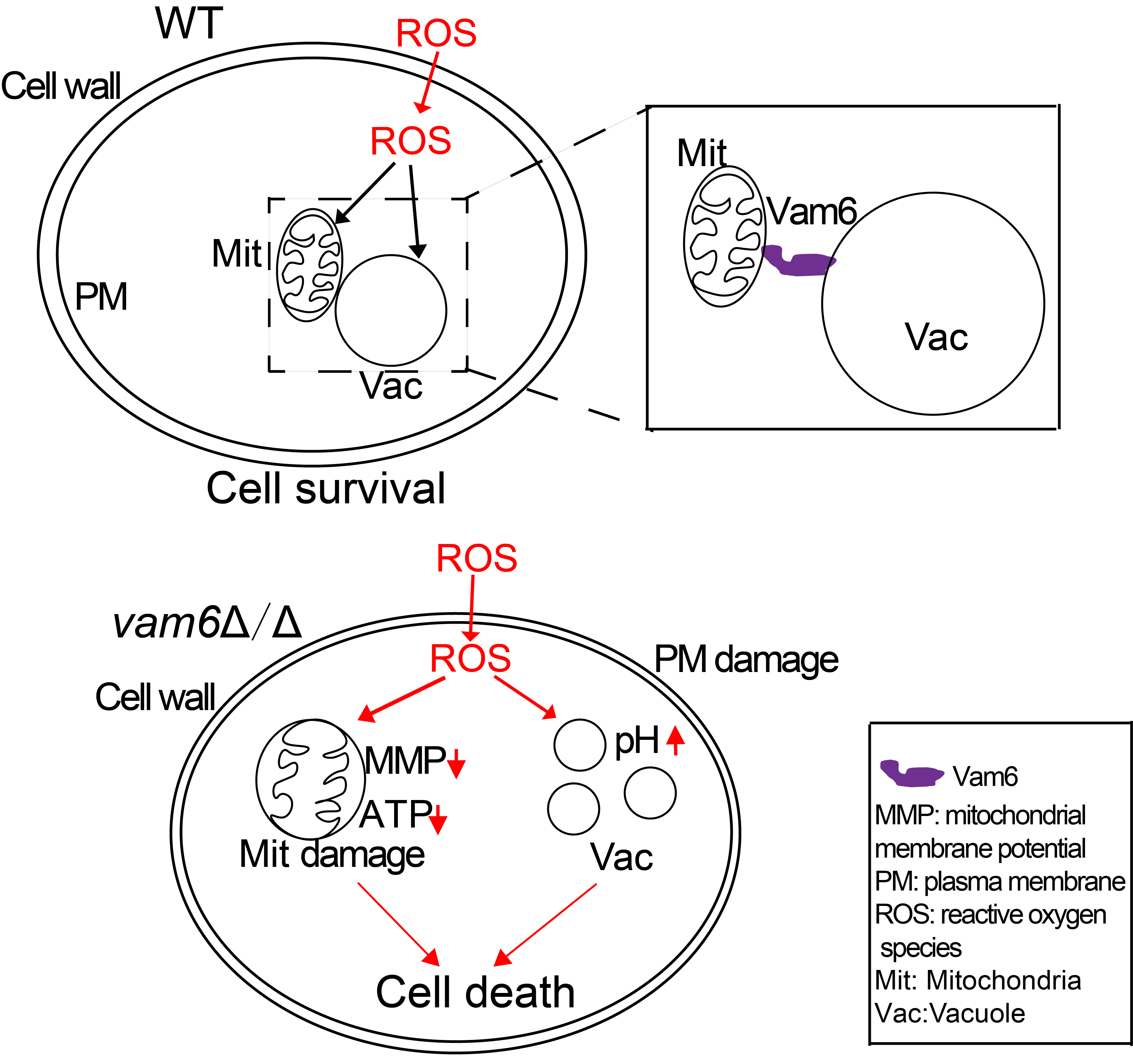
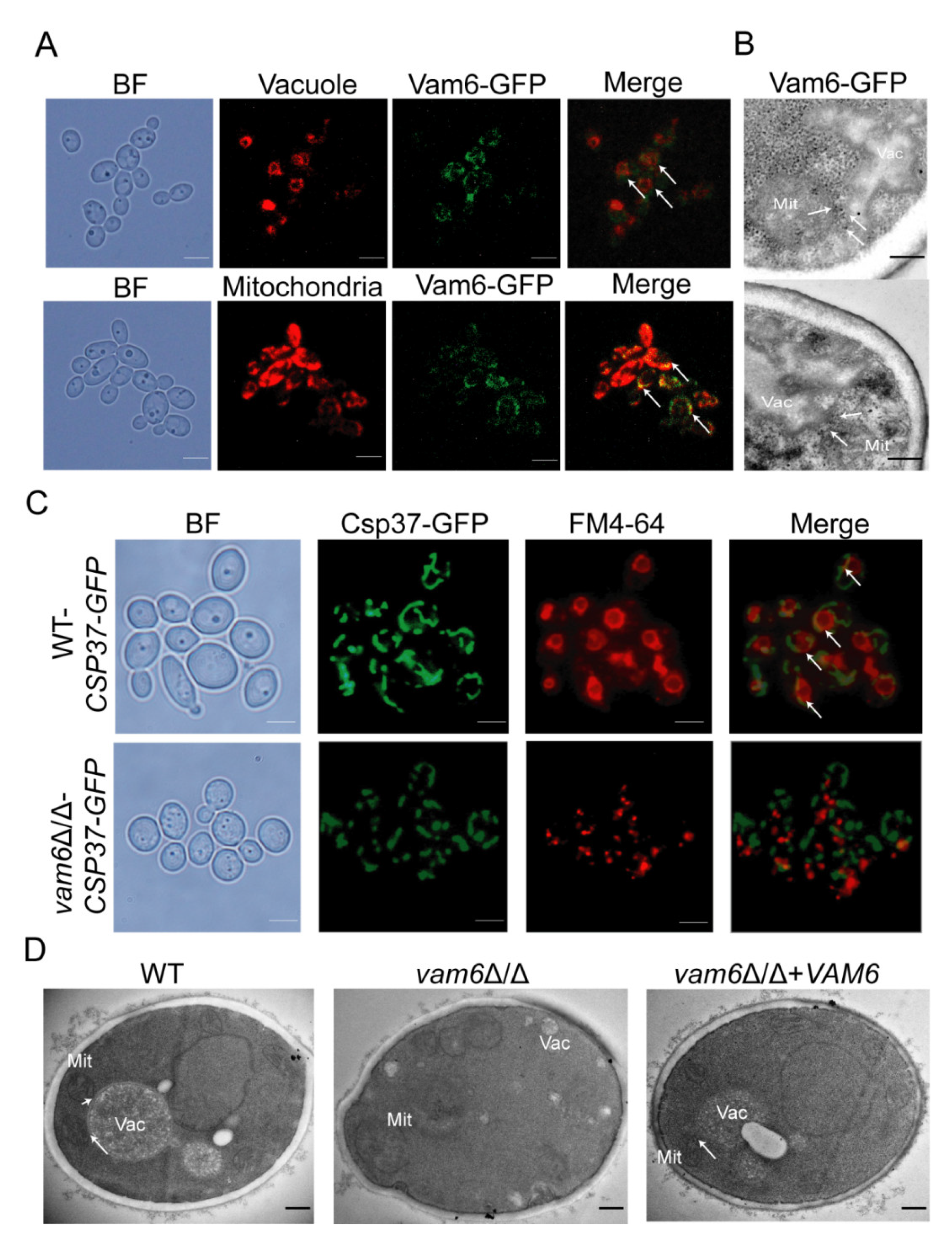
3.1. Vam6 Is Critical for Formation of vCLAMPs
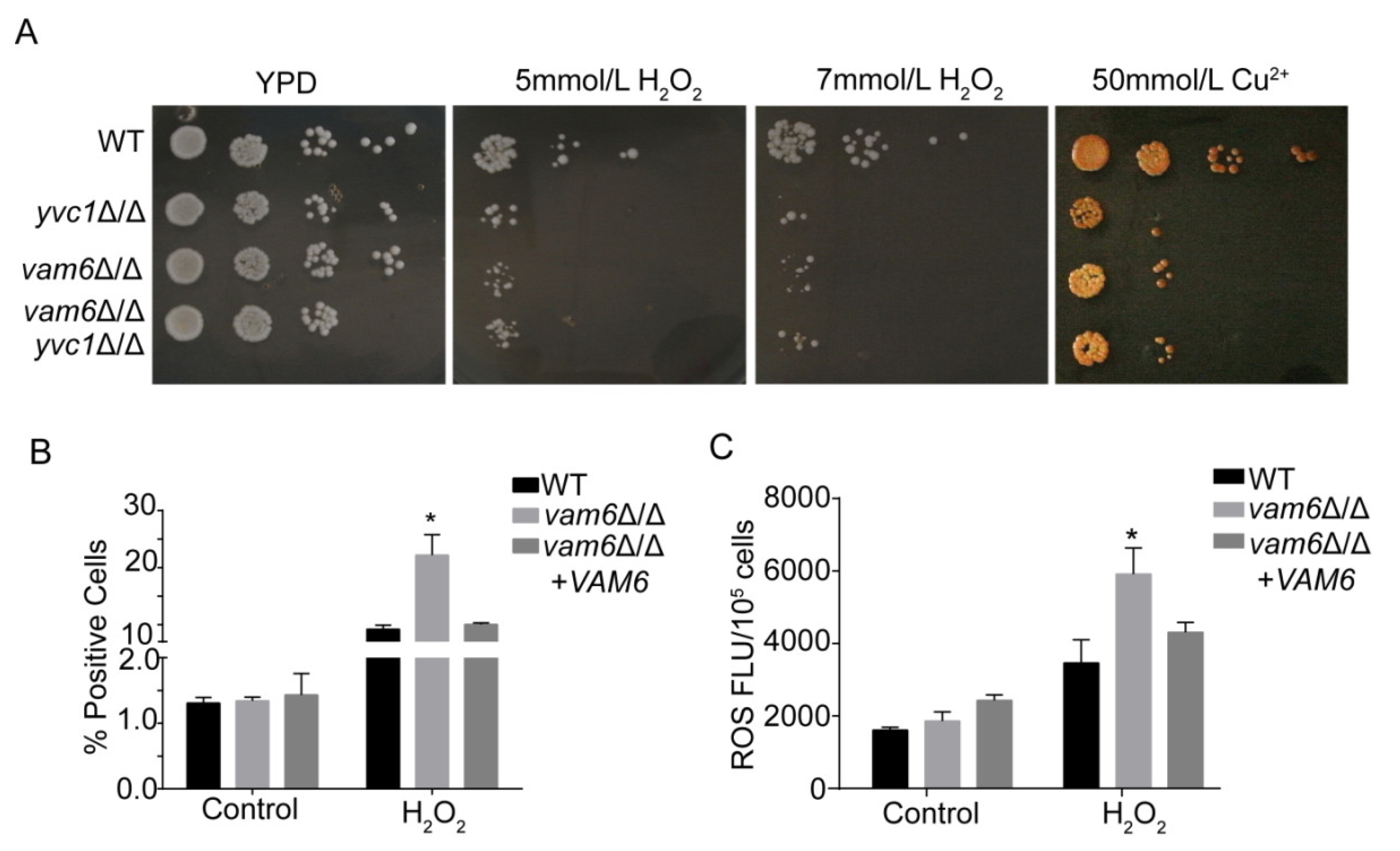
3.2. Deletion of VAM6 Leads to Hypersensitivity to Oxidative Stress
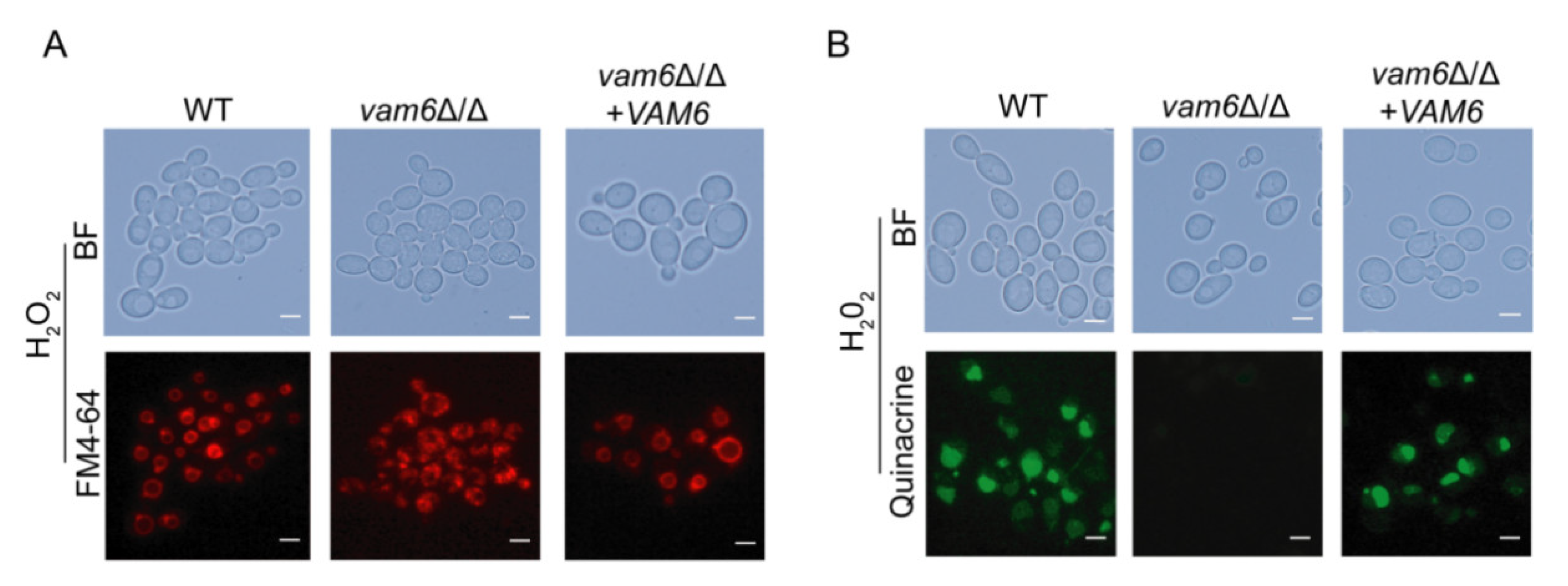
3.3. Deletion of VAM6 Affects Vacuolar Morphology and Function under Oxidative Stress
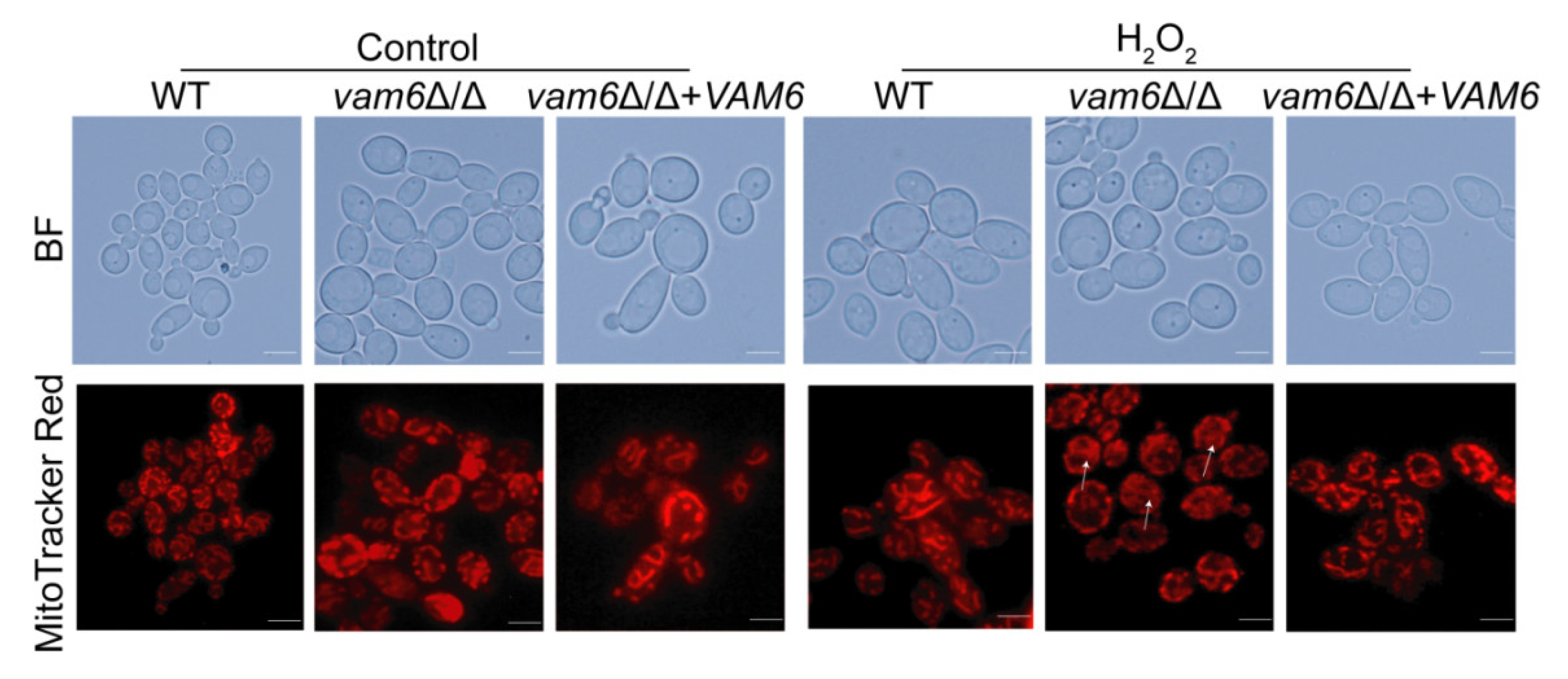
3.4. Vam6 Is Required for Maintaining Mitochondrial Morphology under Oxidative Stress
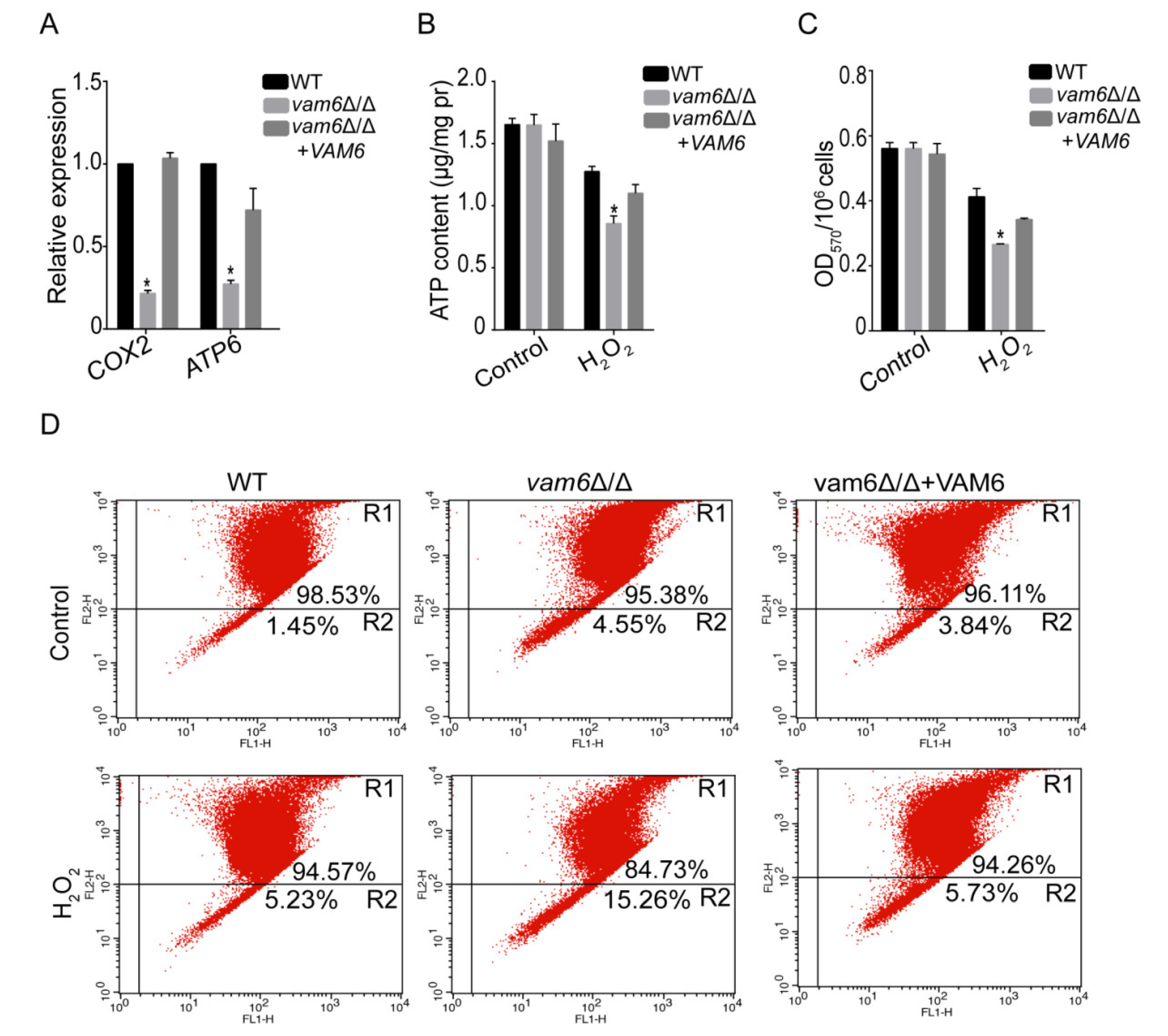
3.5. Deletion of VAM6 Leads to Mitochondrial Dysfunction
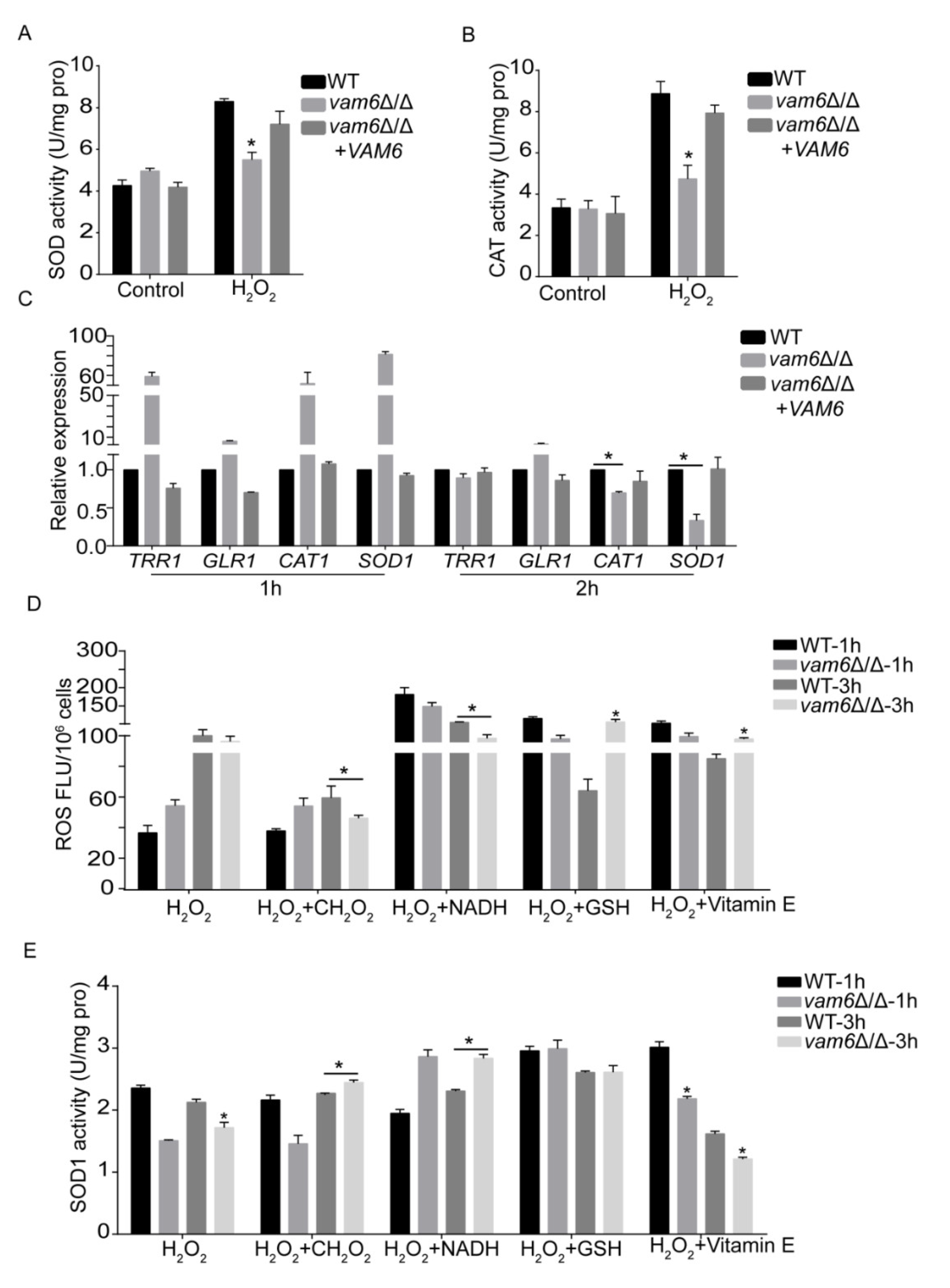
3.6. Effect of Vam6 on Oxidative Stress Response and Antioxidant Capacity
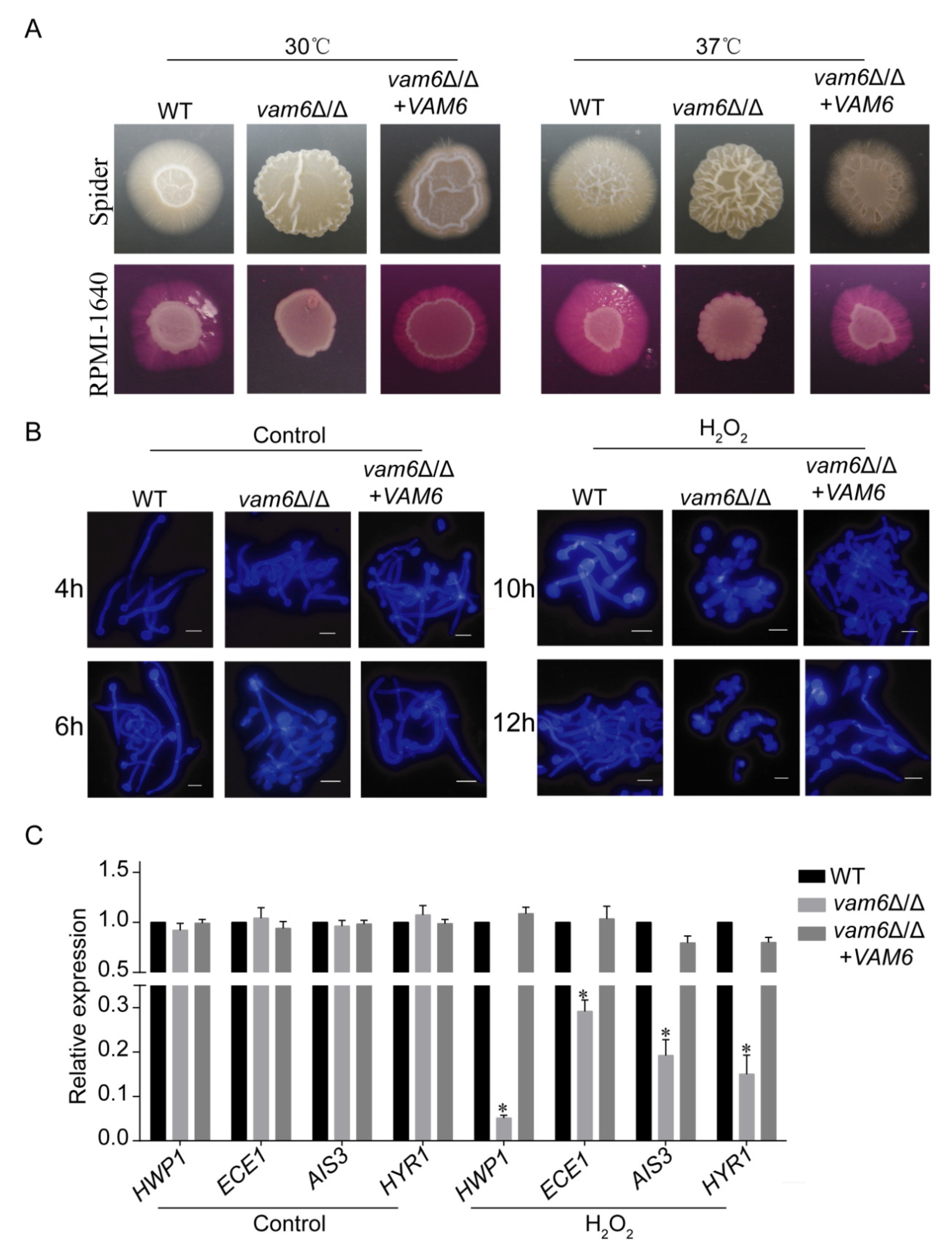
3.7. Deletion of VAM6 Impairs Morphogenesis in C. albicans
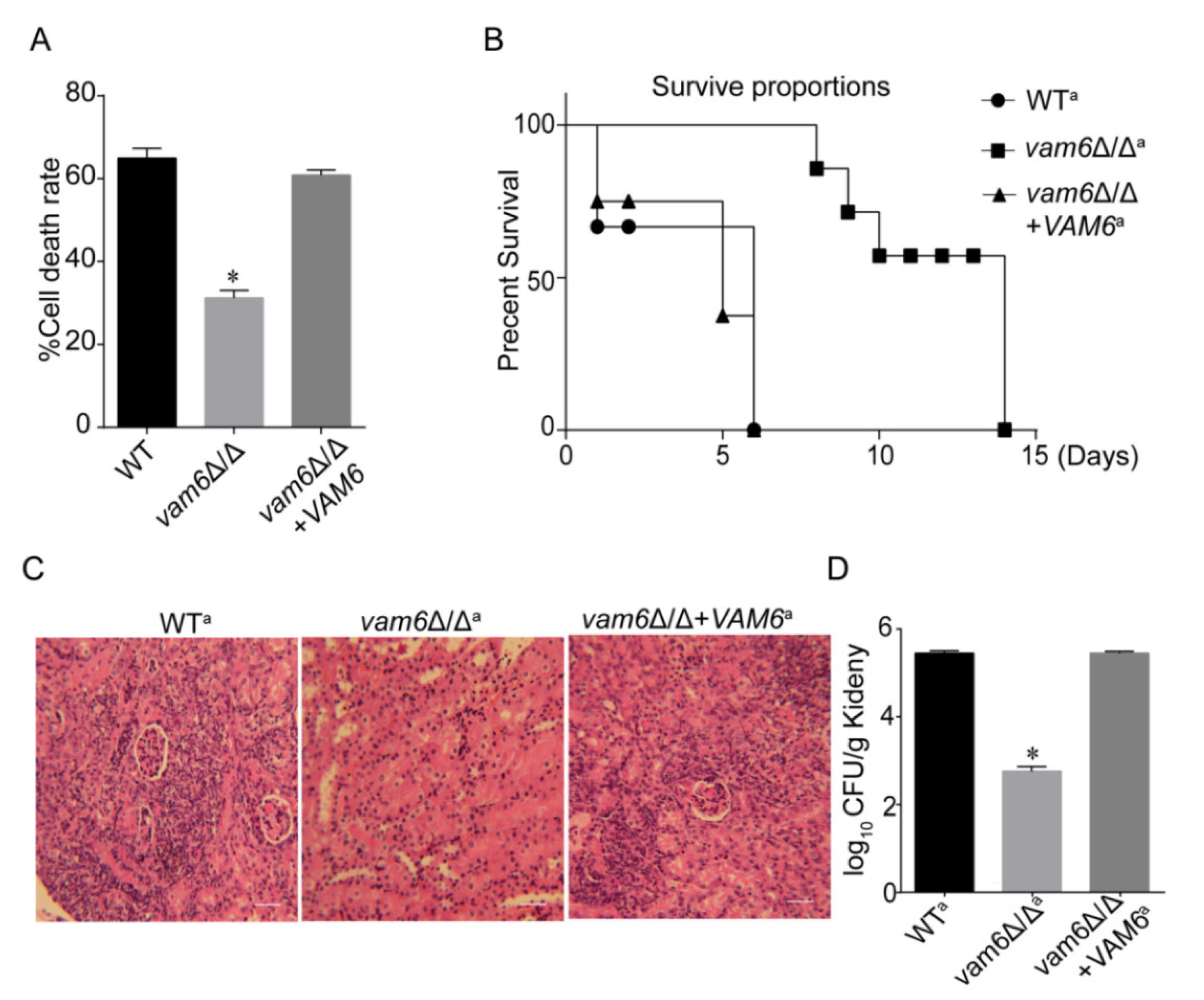
3.8. Deletion of VAM6 Dramatically Reduces the Virulence of C. albicans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, Q.; Zhang, B.; Yang, B.; Chen, J.; Wang, H.; Jia, C.; Ding, X.; Xu, N.; Dong, Y.; Zhang, B.; et al. Interaction among the vacuole, the mitochondria, and the oxidative stress response is governed by the transient receptor potential channel in Candida albicans. Free Radic. Biol. Med. 2014, 77, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Sheevani, P.S.; Aggarwal, A. Nosocomial Candida infection in a rural tertiary care hospital. J. Clin. Diagn. Res. 2013, 7, 405–406. [Google Scholar] [CrossRef] [PubMed]
- Gunsalus, K.T.; Tornberg-Belanger, S.N.; Matthan, N.R.; Lichtenstein, A.H.; Kumamoto, C.A. Manipulation of Host Diet To Reduce Gastrointestinal Colonization by the Opportunistic Pathogen Candida albicans. mSphere 2016, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Yu, Q.; Chen, Y.; Xu, N.; Zhao, Q.; Jia, C.; Zhang, B.; Zhang, K.; Zhang, B.; Xing, L.; et al. The Ccz1 mediates the autophagic clearance of damaged mitochondria in response to oxidative stress in Candida albicans. Int. J. Biochem. Cell Biol. 2015, 69, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.S.; Wilson, D.; Hube, B. Candida albicans iron acquisition within the host. FEMS Yeast Res. 2009, 9, 1000–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enjalbert, B.; MacCallum, D.M.; Odds, F.C.; Brown, A.J. Niche-specific activation of the oxidative stress response by the pathogenic fungus Candida albicans. Infect. Immun. 2007, 75, 2143–2151. [Google Scholar] [CrossRef] [Green Version]
- Palmer, G.E. Vacuolar trafficking and Candida albicans pathogenesis. Commun. Integr. Biol. 2011, 4, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Molero, G.; Diez-Orejas, R.; Navarro-Garcia, F.; Monteoliva, L.; Pla, J.; Gil, C.; Sanchez-Perez, M.; Nombela, C. Candida albicans: Genetics, dimorphism and pathogenicity. Int. Microbiol. 1998, 1, 95–106. [Google Scholar]
- Saville, S.P.; Lazzell, A.L.; Monteagudo, C.; Lopez-Ribot, J.L. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot. Cell 2003, 2, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Bauerova, V.; Hajek, M.; Pichova, I.; Hruskova-Heidingsfeldova, O. Intracellular aspartic proteinase Apr1p of Candida albicans is required for morphological transition under nitrogen-limited conditions but not for macrophage killing. Folia Microbiol. 2014, 59, 485–493. [Google Scholar] [CrossRef]
- Vacca, R.A.; de Pinto, M.C.; Valenti, D.; Passarella, S.; Marra, E.; De Gara, L. Production of reactive oxygen species, alteration of cytosolic ascorbate peroxidase, and impairment of mitochondrial metabolism are early events in heat shock-induced programmed cell death in tobacco Bright-Yellow 2 cells. Plant Physiol. 2004, 134, 1100–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herlan, M.; Vogel, F.; Bornhovd, C.; Neupert, W.; Reichert, A.S. Processing of Mgm1 by the rhomboid-type protease Pcp1 is required for maintenance of mitochondrial morphology and of mitochondrial DNA. J. Biol. Chem. 2003, 278, 27781–27788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.; Zhang, B.; Cui, L.; Li, J.; Yu, Q.; Li, M. Mgm1 is required for maintenance of mitochondrial function and virulence in Candida albicans. Fungal Genet. Biol. 2018, 120, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Ungermann, C. vCLAMPs-an intimate link between vacuoles and mitochondria. Curr. Opin. Cell Biol. 2015, 35, 30–36. [Google Scholar] [CrossRef] [PubMed]
- González Montoro, A.; Auffarth, K.; Hönscher, C.; Bohnert, M.; Becker, T.; Warscheid, B.; Reggiori, F.; van der Laan, M.; Fröhlich, F.; Ungermann, C. Vps39 Interacts with Tom40 to Establish One of Two Functionally Distinct Vacuole-Mitochondria Contact Sites. Dev. Cell 2018, 45, 621–636.e627. [Google Scholar] [CrossRef] [Green Version]
- John Peter, A.T.; Herrmann, B.; Antunes, D.; Rapaport, D.; Dimmer, K.S.; Kornmann, B. Vps13-Mcp1 interact at vacuole-mitochondria interfaces and bypass ER-mitochondria contact sites. J. Cell Biol. 2017, 216, 3219–3229. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N.; Hirata, A.; Ohsumi, Y.; Wada, Y. Vam2/Vps41p and Vam6/Vps39p are components of a protein complex on the vacuolar membranes and involved in the vacuolar assembly in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 1997, 272, 11344–11349. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.B.; Davis, D.; Enloe, B.M.; Mitchell, A.P. A recyclable Candida albicans URA3 cassette for PCR product-directed gene disruptions. Yeast 2000, 16, 65–70. [Google Scholar] [CrossRef]
- Poot, M.; Gibson, L.L.; Singer, V.L. Detection of apoptosis in live cells by MitoTracker red CMXRos and SYTO dye flow cytometry. Cytometry 1997, 27, 358–364. [Google Scholar] [CrossRef]
- Fischer-Parton, S.; Parton, R.M.; Hickey, P.C.; Dijksterhuis, J.; Atkinson, H.A.; Read, N.D. Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J. Microsc. 2000, 198, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Xiao, Y.; Cao, L.L.; Yan, X.; Li, C.; Shi, H.Y.; Wang, J.W.; Ye, Y.H. Cerebroside C increases tolerance to chilling injury and alters lipid composition in wheat roots. PLoS ONE 2013, 8, e73380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanki, T.; Kang, D.; Klionsky, D.J. Monitoring mitophagy in yeast: The Om45-GFP processing assay. Autophagy 2009, 5, 1186–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peralta, E.R.; Martin, B.C.; Edinger, A.L. Differential effects of TBC1D15 and mammalian Vps39 on Rab7 activation state, lysosomal morphology, and growth factor dependence. J. Biol. Chem. 2010, 285, 16814–16821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, I. Attenuation of Candida albicans virulence with focus on disruption of its vacuole functions. J. Oral Microbiol. 2014, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.; Zhang, K.; Zhang, D.; Yu, Q.; Zhao, Q.; Xiao, C.; Dong, Y.; Chu, M.; Li, M. Roles of VPH2 and VMA6 in localization of V-ATPase subunits, cell wall functions and filamentous development in Candida albicans. Fungal Genet. Biol. 2018, 114, 1–11. [Google Scholar] [CrossRef] [PubMed]
- AhYoung, A.P.; Jiang, J.; Zhang, J.; Khoi Dang, X.; Loo, J.A.; Zhou, Z.H.; Egea, P.F. Conserved SMP domains of the ERMES complex bind phospholipids and mediate tether assembly. Proc. Natl. Acad. Sci. USA 2015, 112, E3179–E3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, X.; Wu, D.; Huang, Z.; Hou, T.; Jian, C.; Yu, P.; Lu, F.; Zhang, R.; Sun, T.; et al. Mitochondrial flashes regulate ATP homeostasis in the heart. eLife 2017, 6. [Google Scholar] [CrossRef]
- Bulthuis, E.P.; Adjobo-Hermans, M.J.W.; Willems, P.; Koopman, W.J.H. Mitochondrial Morphofunction in Mammalian Cells. Antioxid. Redox Signal. 2019, 30, 2066–2109. [Google Scholar] [CrossRef] [Green Version]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (deltapsi(m)) in apoptosis; an update. Apoptosis Int. J. Program. Cell Death 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Chekulayeva, L.V.; Shevchuk, I.N.; Chekulayev, V.A.; Ilmarinen, K. Hydrogen peroxide, superoxide, and hydroxyl radicals are involved in the phototoxic action of hematoporphyrin derivative against tumor cells. J. Environ. Pathol. Toxicol. Oncol. 2006, 25, 51–77. [Google Scholar] [CrossRef]
- Kloppel, C.; Michels, C.; Zimmer, J.; Herrmann, J.M.; Riemer, J. In yeast redistribution of Sod1 to the mitochondrial intermembrane space provides protection against respiration derived oxidative stress. Biochem. Biophys. Res. Commun. 2010, 403, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Frohner, I.E.; Bourgeois, C.; Yatsyk, K.; Majer, O.; Kuchler, K. Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance. Mol. Microbiol. 2009, 71, 240–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Yu, Q.; Jia, C.; Wang, Y.; Xiao, C.; Dong, Y.; Xu, N.; Wang, L.; Li, M. The actin-related protein Sac1 is required for morphogenesis and cell wall integrity in Candida albicans. Fungal Genet. Biol. 2015, 81, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; He, H.; Dong, Y.; Pan, H. Hyphae-specific genes HGC1, ALS3, HWP1, and ECE1 and relevant signaling pathways in Candida albicans. Mycopathologia 2013, 176, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Samaranayake, Y.H.; Cheung, B.P.; Yau, J.Y.; Yeung, S.K.; Samaranayake, L.P. Human serum promotes Candida albicans biofilm growth and virulence gene expression on silicone biomaterial. PLoS ONE 2013, 8, e62902. [Google Scholar] [CrossRef] [Green Version]
- Brand, A.; MacCallum, D.M.; Brown, A.J.; Gow, N.A.; Odds, F.C. Ectopic expression of URA3 can influence the virulence phenotypes and proteome of Candida albicans but can be overcome by targeted reintegration of URA3 at the RPS10 locus. Eukaryot. Cell 2004, 3, 900–909. [Google Scholar] [CrossRef] [Green Version]
- Price, A.; Seals, D.; Wickner, W.; Ungermann, C. The docking stage of yeast vacuole fusion requires the transfer of proteins from a cis-SNARE complex to a Rab/Ypt protein. J. Cell Biol. 2000, 148, 1231–1238. [Google Scholar] [CrossRef]
- Oka, M.; Maruyama, J.; Arioka, M.; Nakajima, H.; Kitamoto, K. Molecular cloning and functional characterization of avaB, a gene encoding Vam6p/Vps39p-like protein in Aspergillus nidulans. FEMS Microbiol. Lett. 2004, 232, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Kane, P.M. The long physiological reach of the yeast vacuolar H+-ATPase. J. Bioenergy Biomembr. 2007, 39, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Diebold, L.; Chandel, N.S. Mitochondrial ROS regulation of proliferating cells. Free Radic. Biol. Med. 2016, 100, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, M.; Zhu, Z.; Penka, M.; Helmschrott, C.; Wagener, N.; Wagener, J. Mitochondrial dynamics in the pathogenic mold Aspergillus fumigatus: Therapeutic and evolutionary implications. Mol. Microbiol. 2015, 98, 930–945. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.J.; Price, S.A.; Chilton, L.; Calcutt, N.A.; Tomlinson, D.R.; Verkhratsky, A.; Fernyhough, P. Insulin prevents depolarization of the mitochondrial inner membrane in sensory neurons of type 1 diabetic rats in the presence of sustained hyperglycemia. Diabetes 2003, 52, 2129–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, S.; Dai, C.; Guo, H.; Wang, C.; Hao, Z.; Tang, Q.; Wang, H.; Zhang, Y. Rutin attenuates vancomycin-induced renal tubular cell apoptosis via suppression of apoptosis, mitochondrial dysfunction, and oxidative stress. Phytother. Res. 2019, 33, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Hwang, I.S.; Liu, Q.H.; Woo, E.R.; Lee, D.G. (+)-Medioresinol leads to intracellular ROS accumulation and mitochondria-mediated apoptotic cell death in Candida albicans. Biochimie 2012, 94, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef] [PubMed]
- Albers, D.S.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in aging and neurodegenerative disease. J. Neural Transm. Suppl. 2000, 59, 133–154. [Google Scholar] [PubMed]
- Barelle, C.J.; Priest, C.L.; Maccallum, D.M.; Gow, N.A.; Odds, F.C.; Brown, A.J. Niche-specific regulation of central metabolic pathways in a fungal pathogen. Cell. Microbiol. 2006, 8, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Lima, E.C.; Royer, B.; Vaghetti, J.C.; Simon, N.M.; da Cunha, B.M.; Pavan, F.A.; Benvenutti, E.V.; Cataluna-Veses, R.; Airoldi, C. Application of Brazilian pine-fruit shell as a biosorbent to removal of reactive red 194 textile dye from aqueous solution kinetics and equilibrium study. J. Hazard. Mater. 2008, 155, 536–550. [Google Scholar] [CrossRef]
- Dequard-Chablat, M.; Sellem, C.H.; Golik, P.; Bidard, F.; Martos, A.; Bietenhader, M.; di Rago, J.P.; Sainsard-Chanet, A.; Hermann-Le Denmat, S.; Contamine, V. Two nuclear life cycle-regulated genes encode interchangeable subunits c of mitochondrial ATP synthase in Podospora anserina. Mol. Biol. Evol. 2011, 28, 2063–2075. [Google Scholar] [CrossRef]
- Wilson, R.B.; Davis, D.; Mitchell, A.P. Rapid hypothesis testing with Candida albicans through gene disruption with short homology regions. Journal of bacteriology 1999, 181, 1868–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramón, A.M.; Fonzi, W.A. Diverged binding specificity of Rim101p, the Candida albicans ortholog of PacC. Eukaryotic Cell 2003, 2, 718–728. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, X.; Yang, L.; Liu, Y.; Ma, C.; Ma, T.; Yu, Q.; Li, M. Vacuole and Mitochondria Patch (vCLAMP) Protein Vam6 Is Involved in Maintenance of Mitochondrial and Vacuolar Functions under Oxidative Stress in Candida albicans. Antioxidants 2021, 10, 136. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010136
Mao X, Yang L, Liu Y, Ma C, Ma T, Yu Q, Li M. Vacuole and Mitochondria Patch (vCLAMP) Protein Vam6 Is Involved in Maintenance of Mitochondrial and Vacuolar Functions under Oxidative Stress in Candida albicans. Antioxidants. 2021; 10(1):136. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010136
Chicago/Turabian StyleMao, Xiaolong, Li Yang, Yingzheng Liu, Congcong Ma, Tianyu Ma, Qilin Yu, and Mingchun Li. 2021. "Vacuole and Mitochondria Patch (vCLAMP) Protein Vam6 Is Involved in Maintenance of Mitochondrial and Vacuolar Functions under Oxidative Stress in Candida albicans" Antioxidants 10, no. 1: 136. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010136