Supplemental Microalgal DHA and Astaxanthin Affect Astaxanthin Metabolism and Redox Status of Juvenile Rainbow Trout


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Diet Preparation
2.2. Fish Culture and Sampling
2.3. Astaxanthin Analysis
2.4. Determination of Antioxidant/Oxidant Biomarkers
2.5. Gene Expression
2.6. Statistical Analysis
3. Results
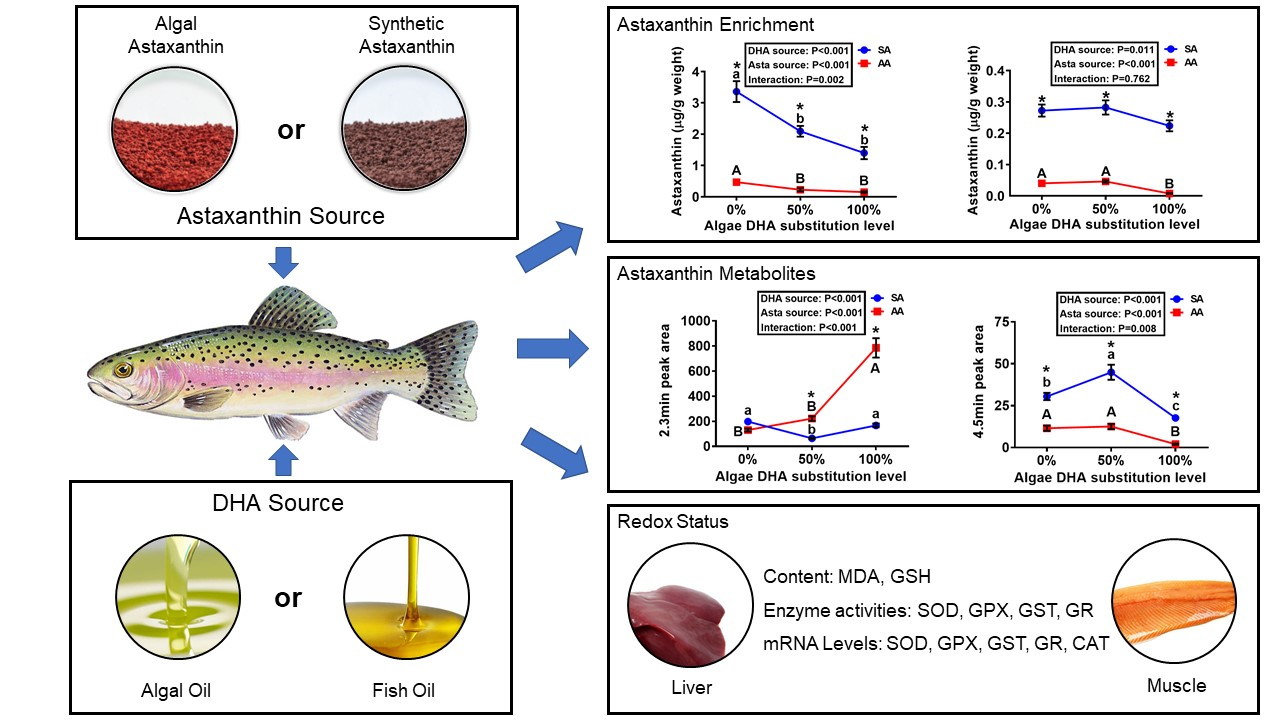
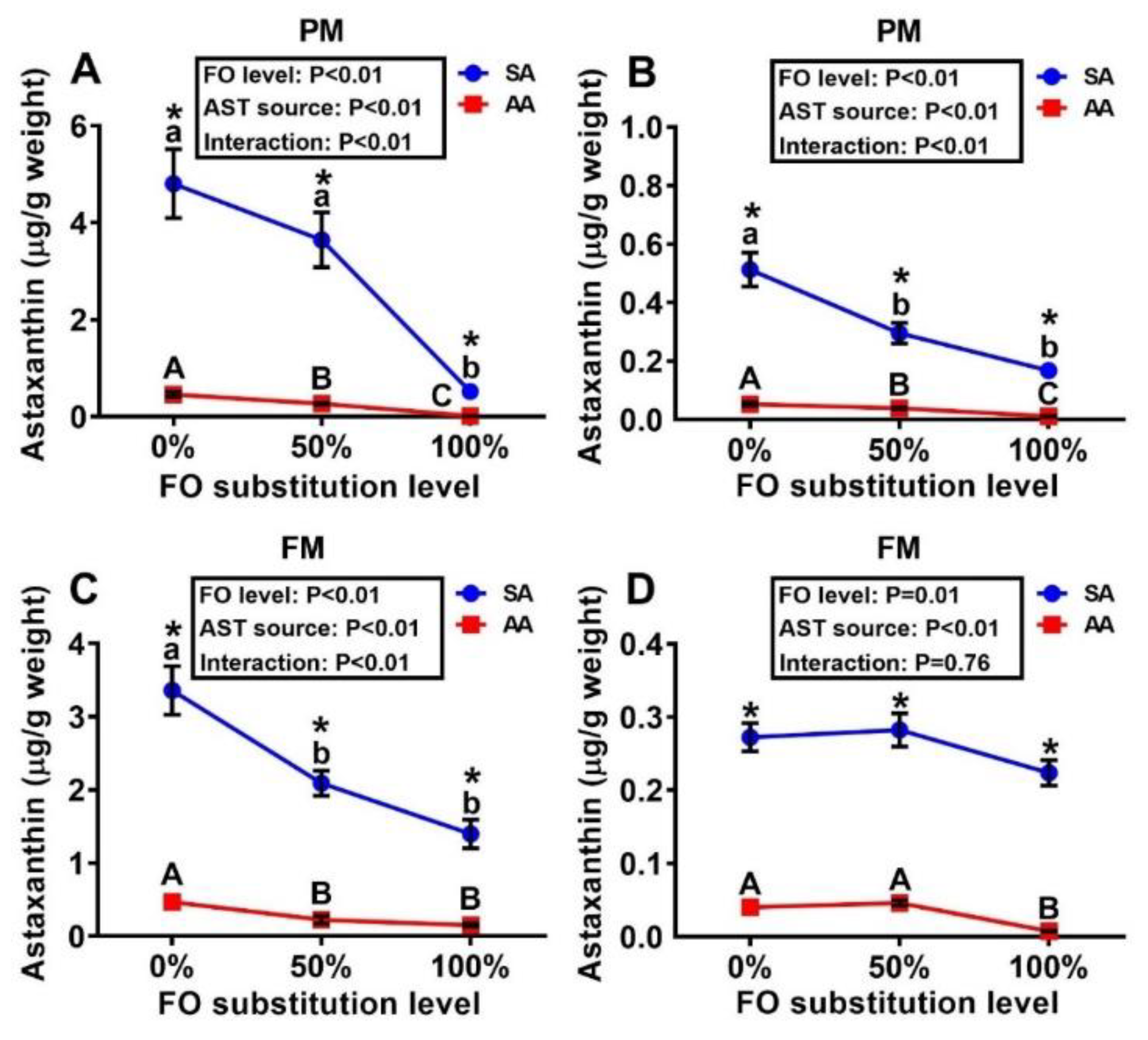
3.1. Concentrations of Astaxanthin in Diets and Tissues
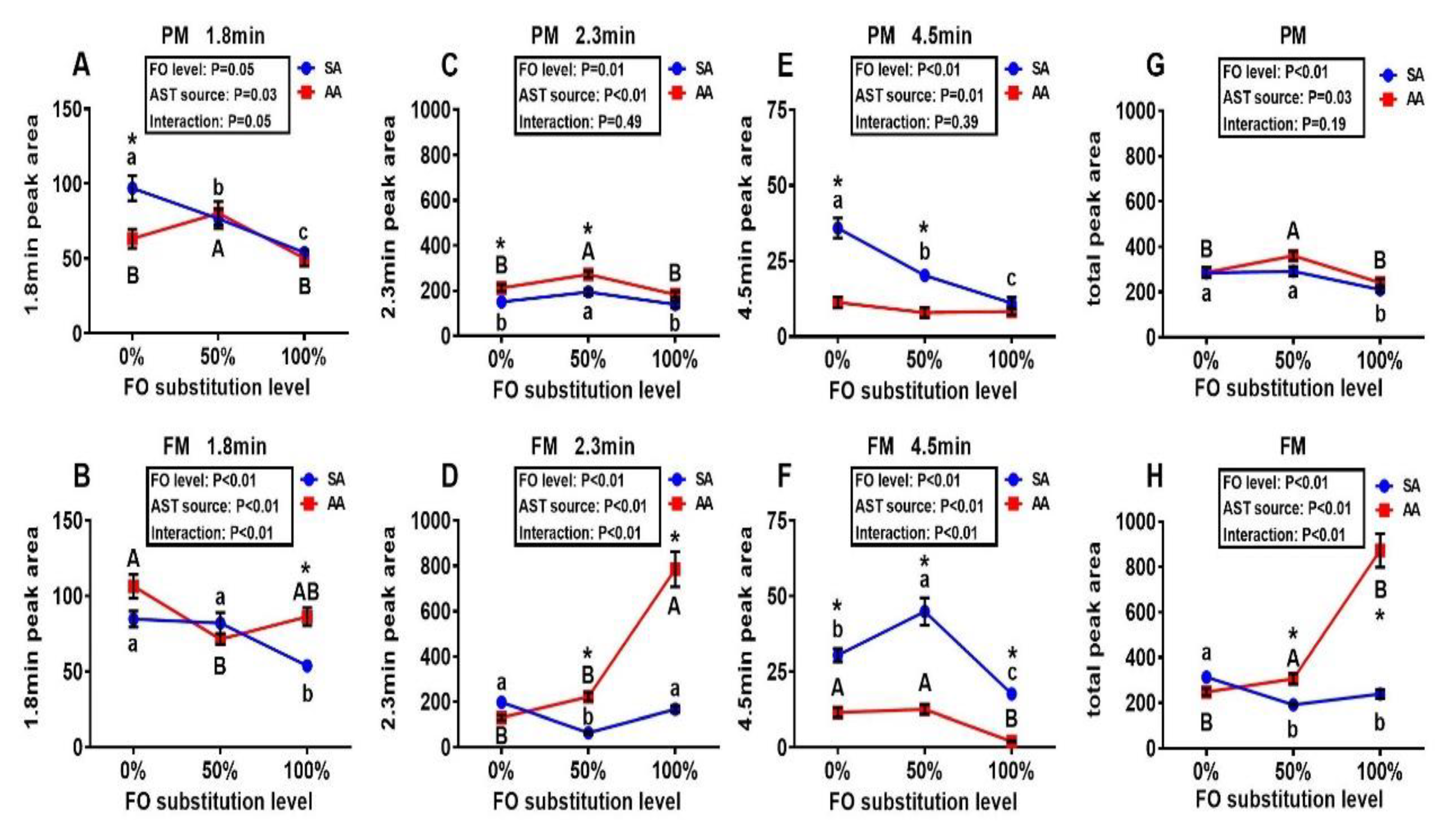
3.2. Metabolites of Astaxanthin in the Liver and Muscle
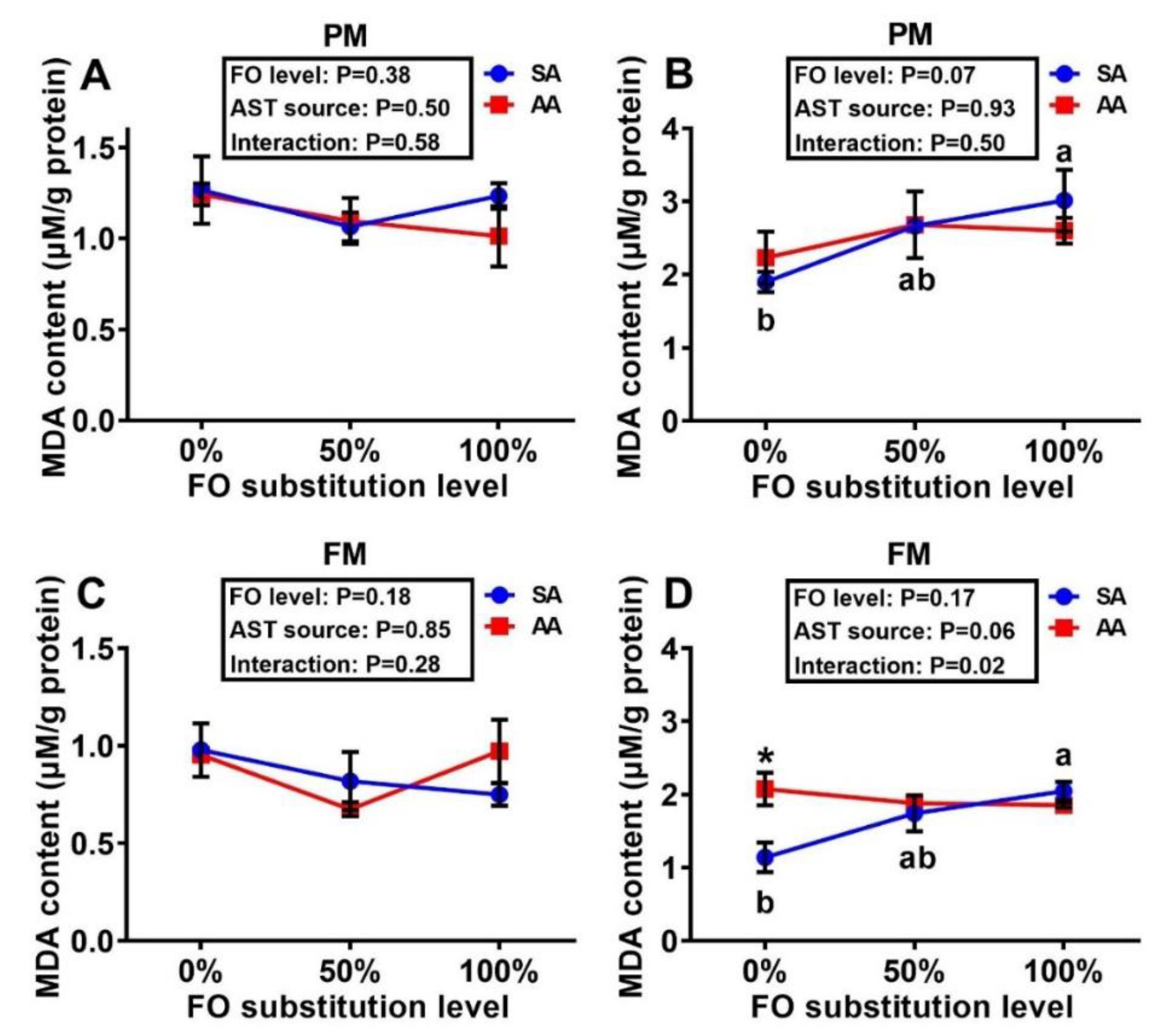
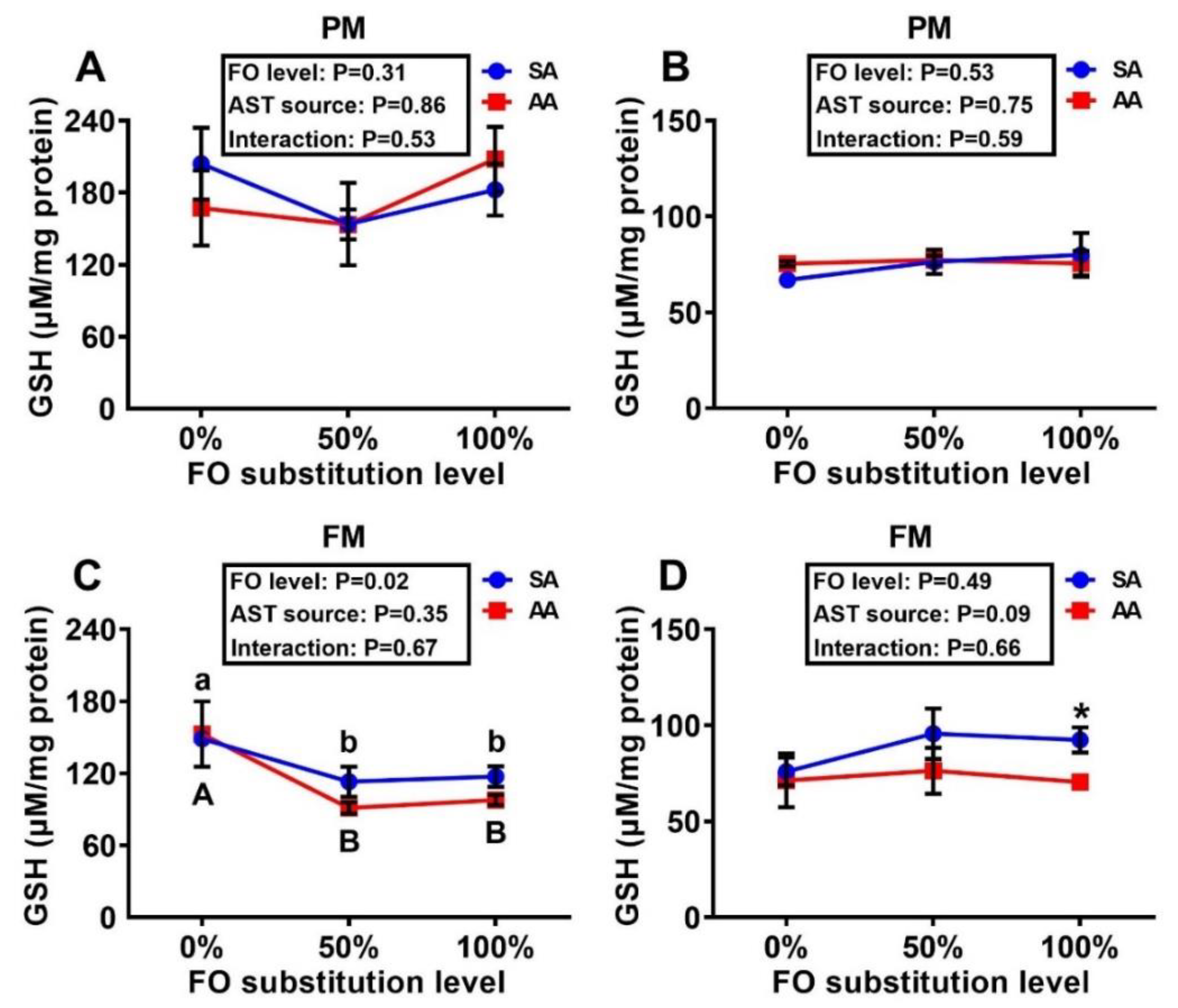
3.3. Tissue MDA and GSH Concentrations and Antioxidant Enzyme Activities
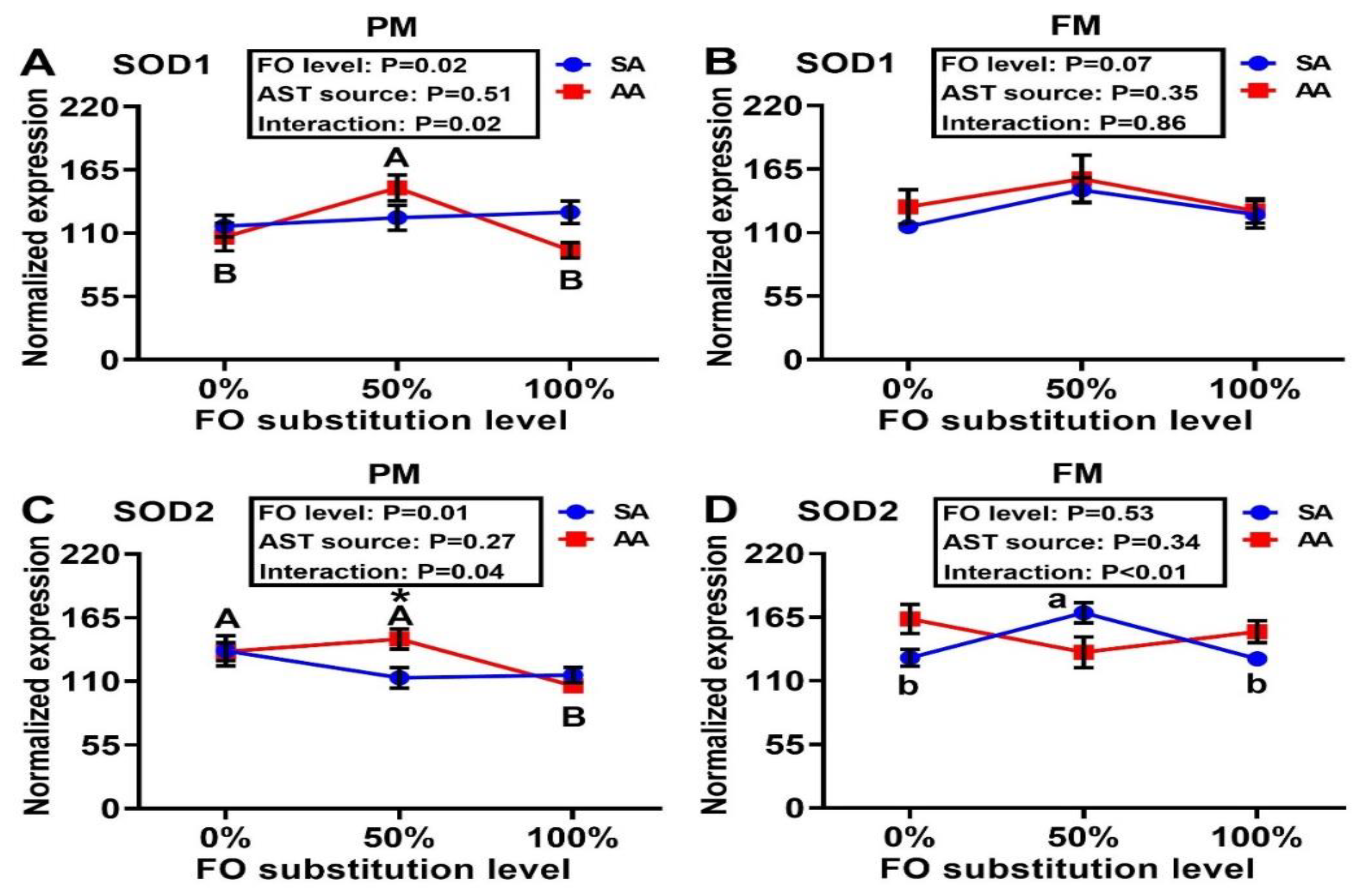
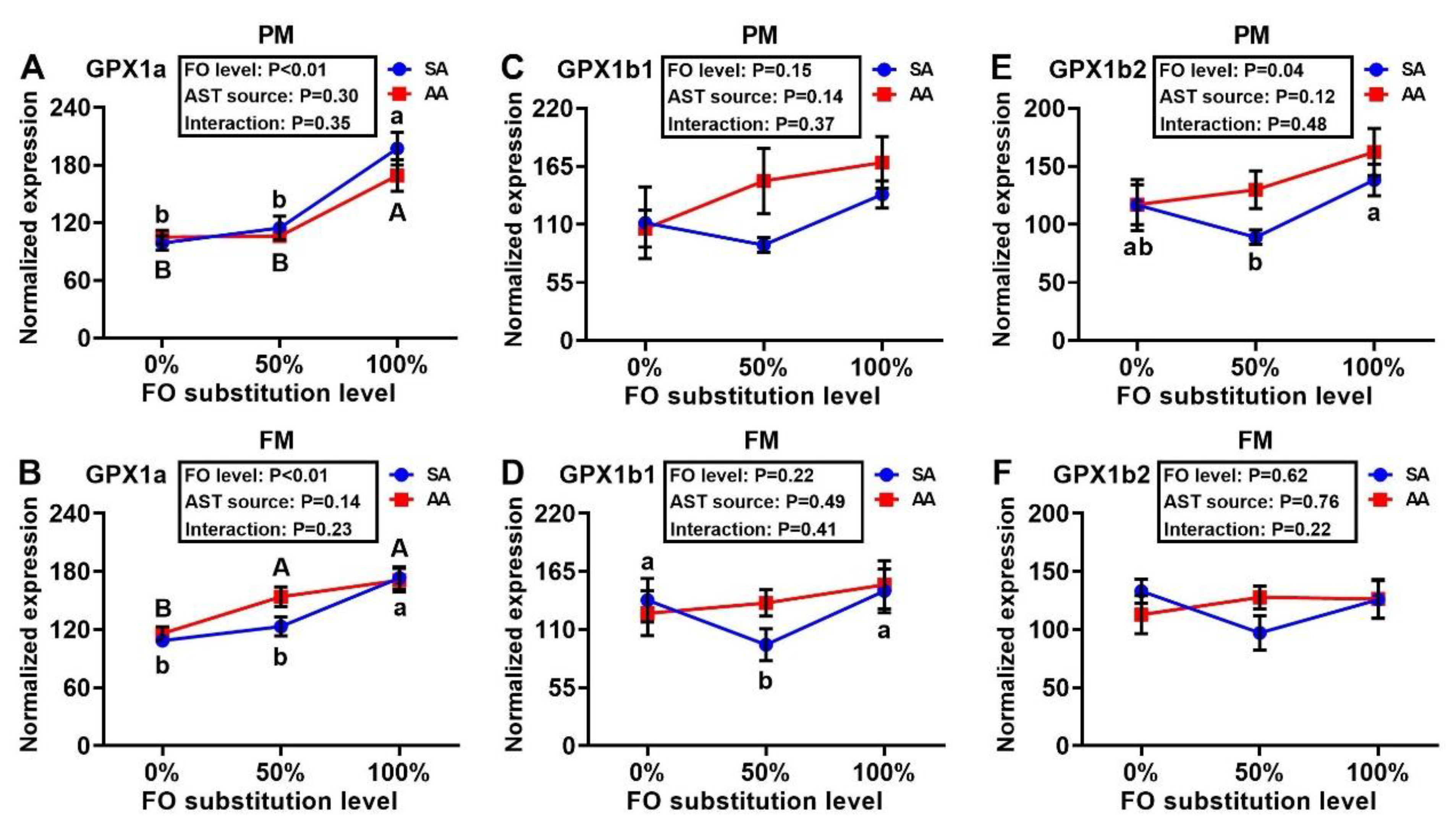
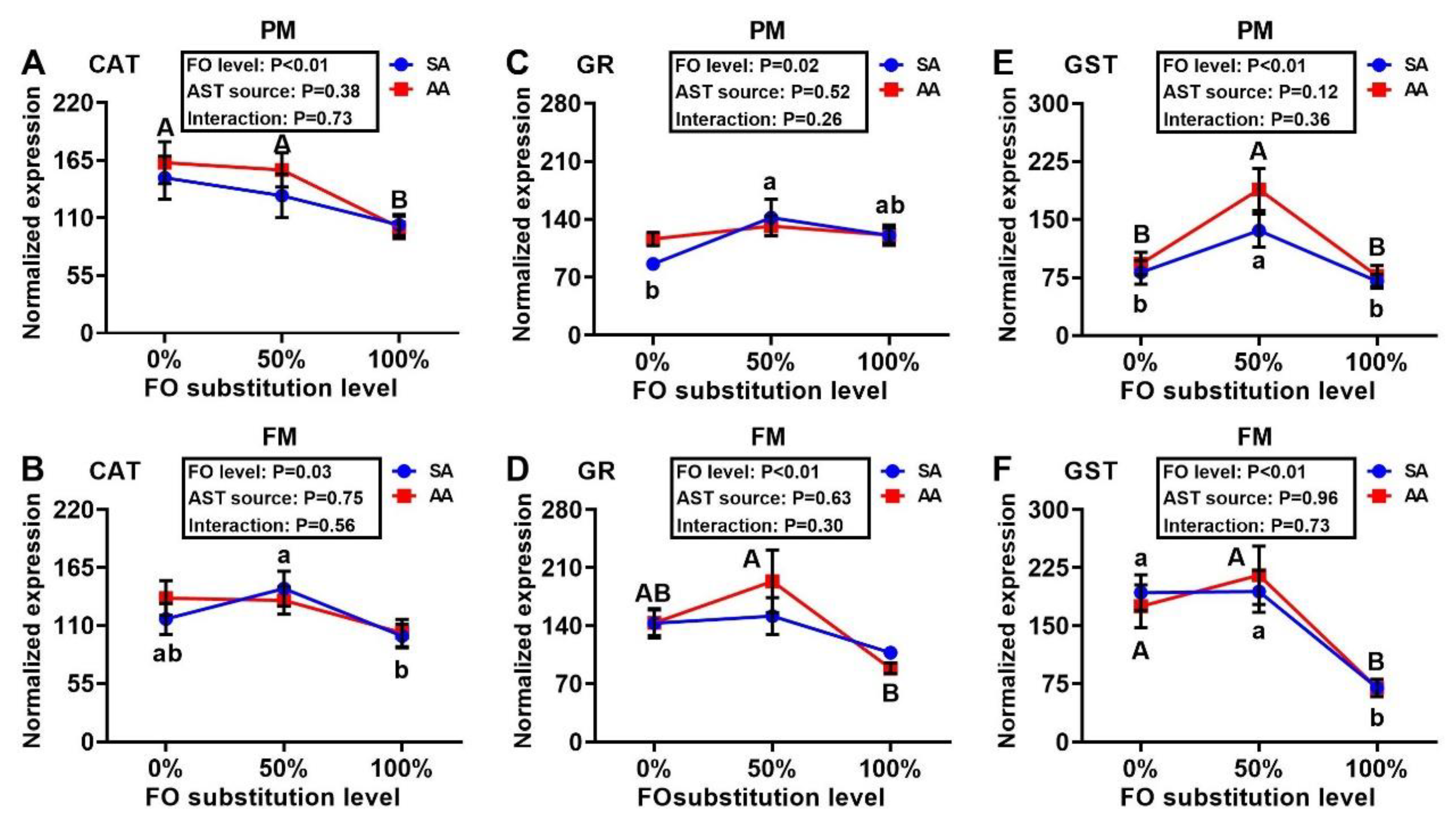
3.4. Antioxidant Enzyme Gene Expressions
3.5. Correlation Analyses
3.6. Stepwise Regression Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Setala, J.; Niukko, J.; Saarni, K. Fish Market Review 2016; Natural Resources Institute Finland (Luke): Helsinki, Finland, 2017. [Google Scholar]
- Adeli, A.; Baghaei, F. Production and supply of rainbow trout in Iran and the world. World J. Fish Mar. Sci. 2013, 5, 335–341. [Google Scholar]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Perez-Velazquez, M.; Gatlin III, D.M.; González-Felix, M.L.; Garcia-Ortega, A.; de Cruz, C.R.; Juarez-Gomez, M.L.; Chen, K. Effect of fishmeal and fish oil replacement by algal meals on biological performance and fatty acid profile of hybrid striped bass (Morone crhysops♀× M. saxatilis♂). Aquaculture 2019, 507, 83–90. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Pentieva, K. Safety of astaxanthin for its use as a novel food in food supplements. EFSA J. 2020, 18, e05993. [Google Scholar]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Capelli, B.; Bagchi, D.; Cysewski, G.R. Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods 2013, 12, 145–152. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on the safety and efficacy of synthetic astaxanthin as feed additive for salmon and trout, other fish, ornamental fish, crustaceans and ornamental birds. EFSA J. 2014, 12, 3724. [Google Scholar] [CrossRef]
- Davinelli, S.; Nielsen, M.E.; Scapagnini, G. Astaxanthin in skin health, repair, and disease: A comprehensive review. Nutrients 2018, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Delgado, A.A.; Khandual, S.; Villanueva-Rodríguez, S.J. Chemical stability of astaxanthin integrated into a food matrix: Effects of food processing and methods for preservation. Food Chem. 2017, 225, 23–30. [Google Scholar] [CrossRef]
- Dominguez-Bocanegra, A.R.; Ponce-Noyola, T.; TorresMunoz, J.A. Astaxanthin production by Phaffia rhodozyma and Haematococcus pluvialis: A comparative study. Appl. Microbiol. Biotechnol. 2007, 75, 783–791. [Google Scholar] [CrossRef]
- Sprague, M.; Walton, J.; Campbell, P.J.; Strachan, F.; Dick, J.R.; Bell, J.G. Replacement of fish oil with a DHA-rich algal meal derived from Schizochytrium sp. on the fatty acid and persistent organic pollutant levels in diets and flesh of Atlantic salmon (Salmo salar, L.) post-smolts. Food Chem. 2015, 185, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Betiku, O.C.; Barrows, F.T.; Ross, C.; Sealey, W.M. The effect of total replacement of fish oil with DHA-Gold® and plant oils on growth and fillet quality of rainbow trout (Oncorhynchus mykiss) fed a plant-based diet. Aquacult. Nutr. 2016, 22, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Belanger-Lamonde, A.; Sarker, P.K.; Ayotte, P.; Bailey, J.L.; Bureau, D.P.; Chouinard, P.Y.; Dewailly, E.; Leblanc, A.; Weber, J.P.; Vandenberg, G.W. Algal and vegetable oils as sustainable fish oil substitutes in rainbow trout diets: An approach to reduce contaminant exposure. J. Food Qual. 2018, 7949782. [Google Scholar] [CrossRef]
- Zhu, S.L.; Portman, M.; Cleveland, B.M.; Magnuson, A.D.; Wu, K.; Sealey, W.; Lei, X.G. Replacing fish oil and astaxanthin by microalgal sources produced different metabolic responses in juvenile rainbow trout fed two types of practical diets. J. Anim. Sci. 2020, in press. [Google Scholar] [CrossRef]
- Sommer, T.R.; D’Souza, F.M.L.; Morrissy, N.M. Pigmentation of adult rainbow trout, Oncorhynchus mykiss, using the green alga Haematococcus pluvialis. Aquaculture 1992, 106, 63–74. [Google Scholar] [CrossRef]
- Lorenz, R.T.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends. Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef]
- Tejera, N.; Cejas, J.R.; Rodriguez, C.; Bjerkeng, B.; Jerez, S.; Bolanos, A.; Lorenzo, A. Pigmentation, carotenoids, lipid peroxides and lipid composition of skin of red porgy (Pagrus pagrus) fed diets supplemented with different astaxanthin sources. Aquaculture 2007, 270, 218–230. [Google Scholar] [CrossRef]
- Sun, T.; Yin, R.; Magnuson, A.D.; Tolba, S.A.; Liu, G.; Lei, X.G. Dose-dependent enrichments and improved redox status in tissues of broiler chicks under heat stress by dietary supplemental microalgal astaxanthin. J. Agric. Food Chem. 2018, 66, 5521–5530. [Google Scholar] [CrossRef]
- Naguib, Y.M. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Ismail, A.; Bannenberg, G.; Rice, H.B.; Schutt, E.; MacKay, D. Oxidation in EPA-and DHA-rich oils: An overview. Lipid Tech. 2016, 28, 55–59. [Google Scholar] [CrossRef]
- Lopez, A.B.; Van Eck, J.; Conlin, B.J.; Paolillo, D.J.; O’Neill, J.; Li, L. Effect of the cauliflower or transgene on carotenoid accumulation and chromoplast formation in transgenic potato tubers. J. Exp. Bot. 2008, 59, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Sowell, A.L.; Huff, D.L.; Yeager, P.R.; Caudill, S.P.; Gunter, E.W. Retinol, alpha-tocopherol, lutein/zeaxanthin, beta-cryptoxanthin, lycopene, alpha-carotene, trans-beta-carotene, and four retinyl esters in serum determined simultaneously by reversed-phase HPLC with multiwavelength detection. Clin. Chem. 1994, 40, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Breithaupt, D.E. Identification and quantification of astaxanthin esters in shrimp (Pandalus borealis) and in a microalga (Haematococcus pluvialis) by liquid chromatography-mass spectrometry using negative ion atmospheric pressure chemical ionization. J. Agric. Food Chem. 2004, 52, 3870–3875. [Google Scholar] [CrossRef] [PubMed]
- Rohrle, F.T.; Moloney, A.P.; Osorio, M.T.; Luciano, G.; Priolo, A.; Caplan, P.; Monahan, F.J. Carotenoid, colour and reflectance measurements in bovine adipose tissue to discriminate between beef from different feeding systems. Meat Sci. 2011, 88, 347–353. [Google Scholar] [CrossRef]
- Mcdonald, R.E.; Hultin, H.O. Some characteristics of the enzymatic lipid-peroxidation system in the microsomal fraction of flounder skeletal-muscle. J. Food Sci. 1987, 52, 15–21. [Google Scholar] [CrossRef]
- Massey, V.; Williams, C.H., Jr. On the reaction mechanism of yeast glutathione reductase. J. Biol. Chem. 1965, 240, 4470–4480. [Google Scholar]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar]
- Mannervik, B.; Guthenberg, C. Glutathione transferase (human placenta). Meth. Enzymol. 1981, 77, 231–235. [Google Scholar]
- Flohe, L.; Gunzler, W.A. Assays of glutathione peroxidase. Meth. Enzymol. 1984, 105, 114–121. [Google Scholar]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovas. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Duval, C.; Auge, N.; Frisach, M.F.; Casteilla, L.; Salvayre, R.; Negre-Salvayre, A. Mitochondrial oxidative stress is modulated by oleic acid via an epidermal growth factor receptor-dependent activation of glutathione peroxidase. Biochem. J. 2002, 367, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanas-Sanchez, V.; Perez, J.A.; Fabelo, N.; Herrera-Herrera, A.V.; Fernandez, C.; Marin, R.; Gonzalez-Montelongo, M.C.; Diaz, M. Addition of docosahexaenoic acid, but not arachidonic acid, activates glutathione and thioredoxin antioxidant systems in murine hippocampal HT22 cells: Potential implications in neuroprotection. J. Neurochem. 2014, 131, 470–483. [Google Scholar] [CrossRef] [PubMed]
- Iuchi, K.; Ema, M.; Suzuki, M.; Yokoyama, C.; Hisatomi, H. Oxidized unsaturated fatty acids induce apoptotic cell death in cultured cells. Mol. Med. Rep. 2019, 19, 2767–2773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, G.; Zarkovic, N. Revealing mechanisms of selective, concentration-dependent potentials of 4-hydroxy-2-nonenal to induce apoptosis in cancer cells through inactivation of membrane-associated catalase. Free Radic. Biol. Med. 2015, 81, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.J.; Tian, L.X.; Yang, H.J.; Liang, G.Y.; Yue, Y.R. Effects of dietary astaxanthin on growth, antioxidant capacity and gene expression in Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2013, 19, 917–927. [Google Scholar] [CrossRef]
- Wang, H.; Dai, A.; Liu, F.; Guan, Y. Effects of dietary astaxanthin on the immune response, resistance to white spot syndrome virus and transcription of antioxidant enzyme genes in Pacific white shrimp Litopenaeus vannamei. Iran. J. Fish. Sci. 2015, 14, 699–718. [Google Scholar]
- Liu, F.; Shi, H.; Guo, Q.; Yu, Y.; Wang, A.; Lv, F.; Shen, W.B. Effects of astaxanthin and emodin on the growth, stress resistance and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immun. 2016, 51, 125–135. [Google Scholar] [CrossRef]
- Rao, A.R.; Reddy, A.H.; Aradhya, S.M. Antibacterial properties of Spirulina platensis, Haematococcus pluvialis, Botryococcus braunii micro algal extracts. Curr. Trends Biotechnol. Pharm. 2010, 4, 809–819. [Google Scholar]
- Rao, A.R.; Sindhuja, H.N.; Dharmesh, S.M.; Sankar, K.U.; Sarada, R.; Ravishankar, G.A. Effective inhibition of skin cancer, tyrosinase, and antioxidative properties by astaxanthin and astaxanthin esters from the green alga Haematococcus pluvialis. J. Agric. Food Chem. 2013, 61, 3842–3851. [Google Scholar] [CrossRef]
- Ghlissi, Z.; Hakim, A.; Sila, A.; Mnif, H.; Zeghal, K.; Rebai, T.; Bougatef, A.; Sahnoun, Z. Evaluation of efficacy of natural astaxanthin and vitamin E in prevention of colistin-induced nephrotoxicity in the rat model. Environ. Toxicol. Phar. 2014, 37, 960–966. [Google Scholar] [CrossRef]
- Chien, Y.H.; Pan, C.H.; Hunter, B. The resistance to physical stresses by Penaeusmonodon juveniles fed diets supplemented with astaxanthin. Aquaculture 2003, 216, 177–191. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chien, Y.H.; Pan, C.H. Effects of dietary supplementation of carotenoids on survival, growth, pigmentation, and antioxidant capacity of characins, Hyphessobry concallistus. Aquaculture 2006, 261, 641–648. [Google Scholar] [CrossRef]
- Pan, C.H.; Chien, Y.H.; Wang, Y.J. Antioxidant defense to ammonia stress of characins (Hyphessobrycon eques Steindachner) fed diets supplemented with carotenoids. Aquac. Nutr. 2011, 17, 258–266. [Google Scholar] [CrossRef]
- Yang, Y.; Pham, T.X.; Wegner, C.J.; Kim, B.; Ku, C.S.; Park, Y.K.; Lee, J.Y. Astaxanthin lowers plasma TAG concentrations and increases hepatic antioxidant gene expression in diet-induced obesity mice. Brit. J. Nutr. 2014, 112, 1797–1804. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Seo, J.M.; Nguyen, A.; Pham, T.X.; Park, H.J.; Park, Y.; Kim, B.; Bruno, R.S.; Lee, J. Astaxanthin-rich extract from the green alga Haematococcus pluvialis lowers plasma lipid concentrations and enhances antioxidant defense in apolipoprotein E knockout mice. J. Nutr. 2011, 141, 1611–1617. [Google Scholar] [CrossRef] [Green Version]
- Harvey, C.J.; Thimmulappa, R.K.; Singh, A.; Blake, D.J.; Ling, G.; Wakabayashi, N.; Fujii, J.; Myers, A.; Biswal, S. Nrf2-regulated glutathione recycling independent of biosynthesis is critical for cell survival during oxidative stress. Free Radic. Biol. Med. 2009, 46, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Itoh, K.; Yamamoto, M.; Zweier, J.L.; Li, Y. Role of Nrf2 signaling in regulation of antioxidants and phase 2 enzymes in cardiac fibroblasts: Protection against reactive oxygen and nitrogen species-induced cell injury. FEBS Lett. 2005, 579, 3029–3036. [Google Scholar] [CrossRef] [Green Version]
- Wischhusen, P.; Larroquet, L.; Durand, T.; Oger, C.; Galano, J.M.; Rocher, A.; Vigor, C.; Prabhu, P.A.J.; Veron, V.; Briens, M.; et al. Oxidative stress and antioxidant response in rainbow trout fry exposed to acute hypoxia is affected by selenium nutrition of parents and during first exogenous feeding. Free Radic. Bio. Med. 2020, 155, 99–113. [Google Scholar] [CrossRef]
- Pacitti, D.; Wang, T.; Page, M.M.; Martin, S.A.M.; Sweetman, J.; Feldmann, J.; Secombes, C.J. Characterization of cytosolic glutathione peroxidase and phospholipid-hydroperoxide glutathione peroxidase genes in rainbow trout (Oncorhynchus mykiss) and their modulation by in vitro selenium exposure. Aquat. Toxicol. 2013, 130, 97–111. [Google Scholar] [CrossRef]
- Miyashita, K. Paradox of omega-3 PUFA oxidation. Eur. J. Lipid Sci. Tech. 2014, 116, 1268–1279. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids in marine animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Showalter, L.A.; Weinman, S.A.; Osterlie, M.; Lockwood, S.F. Plasma appearance and tissue accumulation of non-esterified, free astaxanthin in C57BL/6 mice after oral dosing of a disodium disuccinate diester of astaxanthin (Heptax). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 137, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.H.; Shiau, W.C. The effects of dietary supplementation of algae and synthetic astaxanthin on body astaxanthin, survival, growth, and low dissolved oxygen stress resistance of kuruma prawn, Marsupenaeus japonicus Bate. J. Exp. Mar. Biol. Ecol. 2005, 318, 201–211. [Google Scholar] [CrossRef]
- Barbosa, M.J.; Morais, R.; Choubert, G. Effect of carotenoid source and dietary lipid content on blood astaxanthin concentration in rainbow trout (Oncorhynchus mykiss). Aquaculture 1999, 176, 331–341. [Google Scholar] [CrossRef]
- Okada, Y.; Ishikura, M.; Maoka, T. Bioavailability of astaxanthin in Haematococcus algal extract: The effects of timing of diet and smoking habits. Biosci. Biotechnol. Biochem. 2009, 73, 1928–1932. [Google Scholar] [CrossRef] [Green Version]
- Chimsung, N.; Tantikitti, C.; Milley, J.E.; Verlhac-Trichet, V.; Lall, S.P. Effects of various dietary factors on astaxanthin absorption in Atlantic salmon (Salmo salar). Aquac. Res. 2014, 45, 1611–1620. [Google Scholar] [CrossRef]
- Conchillo, A.; Valencia, I.; Puente, A.; Ansorena, D.; Astiasaran, I. Functional components in fish and algae oils. Nutr. Hosp. 2006, 21, 369–373. [Google Scholar]
- Bjerkeng, B.; Hatlen, B.; Wathne, E. Deposition of astaxanthin in fillets of Atlantic salmon (Salmo salar) fed diets with herring, capelin, sandeel, or Peruvian high PUFA oils. Aquaculture 1999, 180, 307–319. [Google Scholar] [CrossRef]
- Clark, R.M.; Yao, L.; She, L.; Furr, H.C. A comparison of lycopene and astaxanthin absorption from corn oil and olive oil emulsions. Lipids 2000, 35, 803–806. [Google Scholar] [CrossRef]
- Choubert, G.; Heinrich, O. Carotenoid pigments of the green alga Haematococcus pluvialis: Assay on rainbow trout, Oncorhynchus mykiss, pigmentation in comparison with synthetic astaxanthin and canthaxanthin. Aquaculture 1993, 112, 217–226. [Google Scholar] [CrossRef]
- Pan, C.H.; Chien, Y.H. Effects of dietary supplementation of alga Haematococcus pluvialis (Flotow), synthetic astaxanthin and β-carotene on survival, growth, and pigment distribution of red devil, Cichlasoma citrinellum (Gunther). Aquac. Res. 2009, 40, 871–879. [Google Scholar] [CrossRef]
- Storebakken, T.; Foss, P.; Schiedt, K.; Austreng, E.; Liaaen-Jensen, S.; Manz, U. Carotenoid diets in salmonids: IV. Pigmentation of Atlantic salmon with astaxanthin, astaxanthin dipalmitate and canthaxanthin. Aquaculture 1987, 43, 185–193. [Google Scholar]
- Johnson, E.A.; An, G.H. Astaxanthin from microbial sources. Crit. Rev. Biotechnol. 1991, 11, 297–326. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | |||||||
|---|---|---|---|---|---|---|---|
| Fish Oil Replacement Level 2 | 0% | 50% | 100% | SEM | |||
| Astaxanthin Source 3 | SA | AA | SA | AA | SA | AA | |
| Plant meal-based diet | |||||||
| Astaxanthin (μg/g diet) | 42 a | 6.7 b | 33 a | 7.1 b | 32 a | 4.1 b | 0.39 |
| Fishmeal-based diet | |||||||
| Astaxanthin (μg/g diet) | 41 a | 5.8 b | 37 a | 5.1 b | 28 a | 4.0 b | 0.42 |
| Diets | p Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fish Oil Replacement Level 1 | 0% | 50% | 100% | FO Level | AST Source | Interaction | ||||
| Astaxanthin Source 2 | SA | AA | SA | AA | SA | AA | SEM | |||
| Plant Meal-Based Diet | ||||||||||
| Muscle | ||||||||||
| GST 3 (mU/mg protein) | 0.81 a,b | 1.1 a | 0.90 a | 0.85 a,b | 0.72 a,b | 0.54 b | 0.03 | 0.90 | 0.15 | 0.05 |
| GPX (mU/mg protein) | 2.9 a,b | 3.4 a | 2.1 b | 2.9 a,b | 2.3 a,b | 2.5 a,b | 0.15 | 0.11 | 0.71 | 0.16 |
| GR (mU/mg protein) | 0.26 a,b | 0.32 a | 0.19 b | 0.24 a,b | 0.18 b | 0.20 b | 0.02 | 0.13 | 0.71 | 0.02 |
| SOD (mU/mg protein) | 23 a | 24 a | 19 a,b | 24 a | 16 a,b | 12 b | 0.01 | 0.92 | 0.29 | 1.4 |
| Liver | ||||||||||
| GST (mU/mg protein) | 4.6 a,b,c | 5.7 a,b | 4.4 b,c | 6.1 a | 3.9 c | 3.9 c | 0.03 | 0.04 | 0.24 | 0.26 |
| GPX (mU/mg protein) | 13 a,b | 16 a | 16 a | 14 a,b | 11 b | 12 a,b | 0.04 | 0.50 | 0.22 | 0.62 |
| GR (mU/mg protein) | 2.5 | 2.0 | 2.7 | 3.2 | 2.5 | 2.6 | 0.19 | 0.85 | 0.38 | 0.15 |
| SOD (mU/mg protein) | 139 a,b | 143 a | 99.0 b,c | 120 a,b,c | 88.0 c | 105 a,b,c | 0.01 | 0.22 | 0.80 | 6.64 |
| Fishmeal-Based Diet | ||||||||||
| Muscle | ||||||||||
| GST (mU/mg protein) | 1.0 | 1.1 | 0.85 | 0.84 | 0.84 | 0.86 | 0.03 | 0.89 | 0.95 | 0.04 |
| GPX (mU/mg protein) | 2.4 | 2.4 | 2.5 | 1.7 | 2.8 | 2.3 | 0.15 | 0.11 | 0.71 | 0.14 |
| GR (mU/mg protein) | 0.26 | 0.21 | 0.23 | 0.18 | 0.16 | 0.21 | 0.49 | 0.77 | 0.15 | 0.01 |
| SOD (mU/mg protein) | 26 | 14 | 16 | 21 | 22 | 19 | 0.78 | 0.25 | 0.10 | 1.54 |
| Liver | ||||||||||
| GST (mU/mg protein) | 5.2 a | 4.0 a,b | 2.2 b | 2.2 b | 3.7 a,b | 3.1 b | 0.01 | 0.24 | 0.61 | 0.31 |
| GPX (mU/mg protein) | 14 a | 15 a | 15 a | 9.3 b | 9.8 b | 5.3 c | <0.001 | 0.01 | 0.04 | 0.98 |
| GR (mU/mg protein) | 2.8 b,c | 4.0 a | 3.6 a,b | 2.3 c | 2.8 b,c | 2.3 c | 0.03 | 0.24 | 0.01 | 0.18 |
| SOD (mU/mg protein) | 61 b,c | 96 a | 92 a | 86 a,b | 56 c | 36 c | 0.01 | 0.66 | 0.02 | 5.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, K.; Cleveland, B.M.; Portman, M.; Sealey, W.M.; Lei, X.G. Supplemental Microalgal DHA and Astaxanthin Affect Astaxanthin Metabolism and Redox Status of Juvenile Rainbow Trout. Antioxidants 2021, 10, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010016
Wu K, Cleveland BM, Portman M, Sealey WM, Lei XG. Supplemental Microalgal DHA and Astaxanthin Affect Astaxanthin Metabolism and Redox Status of Juvenile Rainbow Trout. Antioxidants. 2021; 10(1):16. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010016
Chicago/Turabian StyleWu, Kun, Beth M. Cleveland, Mark Portman, Wendy M. Sealey, and Xin Gen Lei. 2021. "Supplemental Microalgal DHA and Astaxanthin Affect Astaxanthin Metabolism and Redox Status of Juvenile Rainbow Trout" Antioxidants 10, no. 1: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010016