Mechanistic Insight into the Regulation of Lipoxygenase-Driven Lipid Peroxidation Events in Human Spermatozoa and Their Impact on Male Fertility


 , , and
, , and 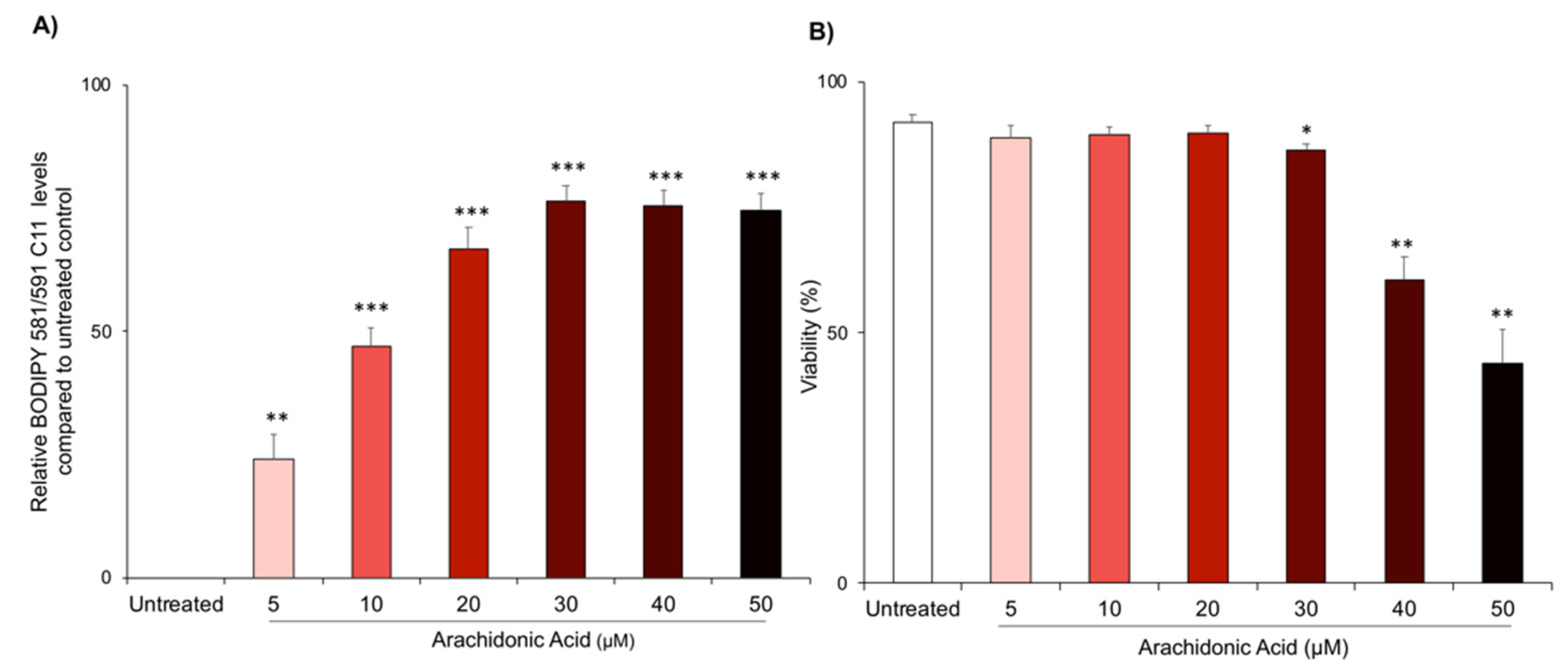{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Reagents
2.3. Preparation of Human Spermatozoa
2.4. Induction of Lipid Peroxidation and Inhibitor Treatment
2.5. Assessment of Lipid Peroxidation
2.6. Comparison of ALOX15 Expression in Infertile and Fertile Spermatozoa
2.7. Statistical Analysis
3. Results
3.1. Arachidonic Acid Treatment Induces Lipid Peroxidation and Cell Death
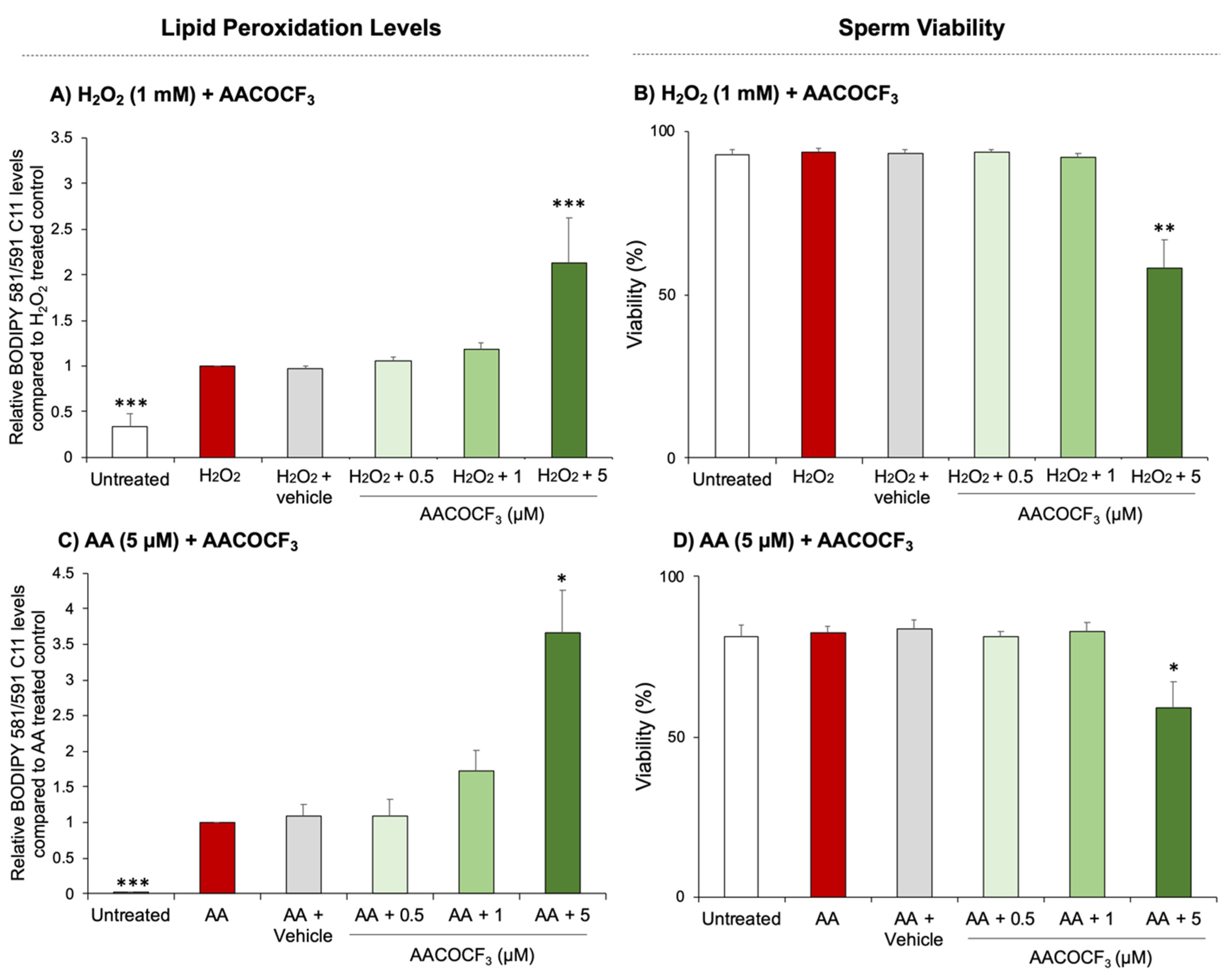
3.2. Phospholipase Inhibition Exacerbates Lipid Peroxidation in Human Spermatozoa
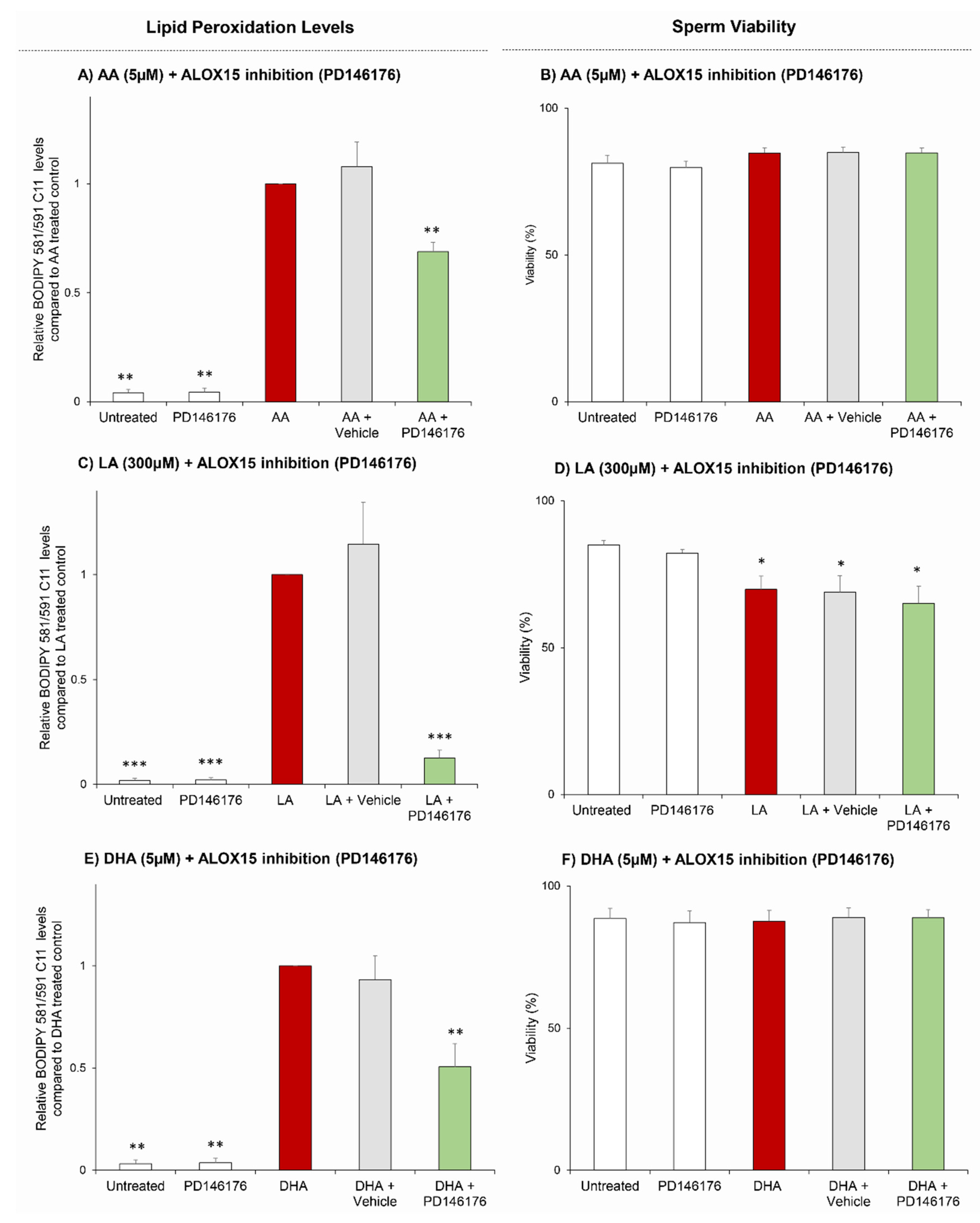
3.3. PUFAs (AA, LA, DHA) Serve as Key Substrates for ALOX15 Driven Lipid Peroxidation in Human Spermatozoa
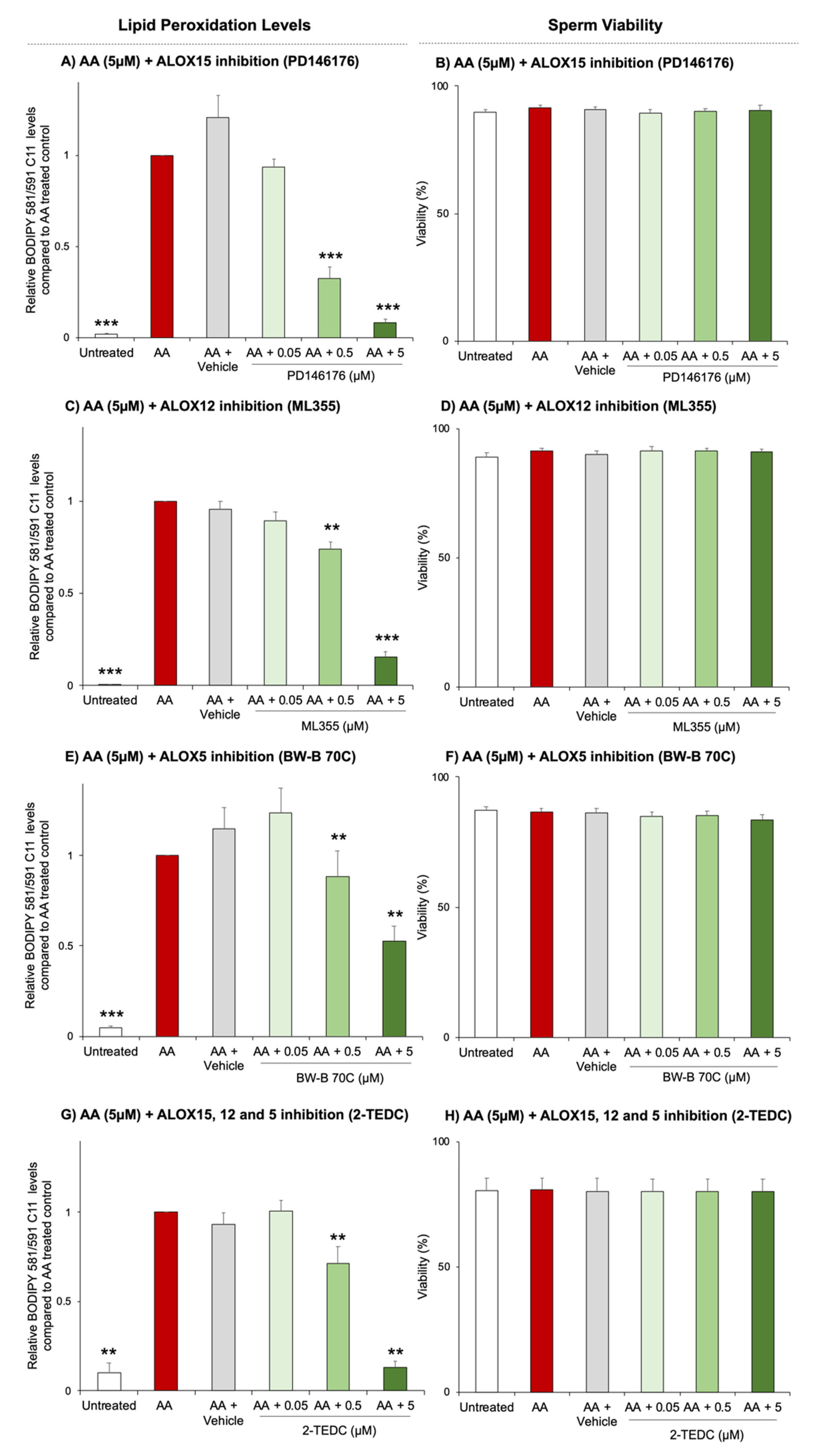
3.4. ALOX15 Plays a Dominant Role in Lipoxygenase-Mediated Lipid Peroxidation Pathways in Human Spermatozoa
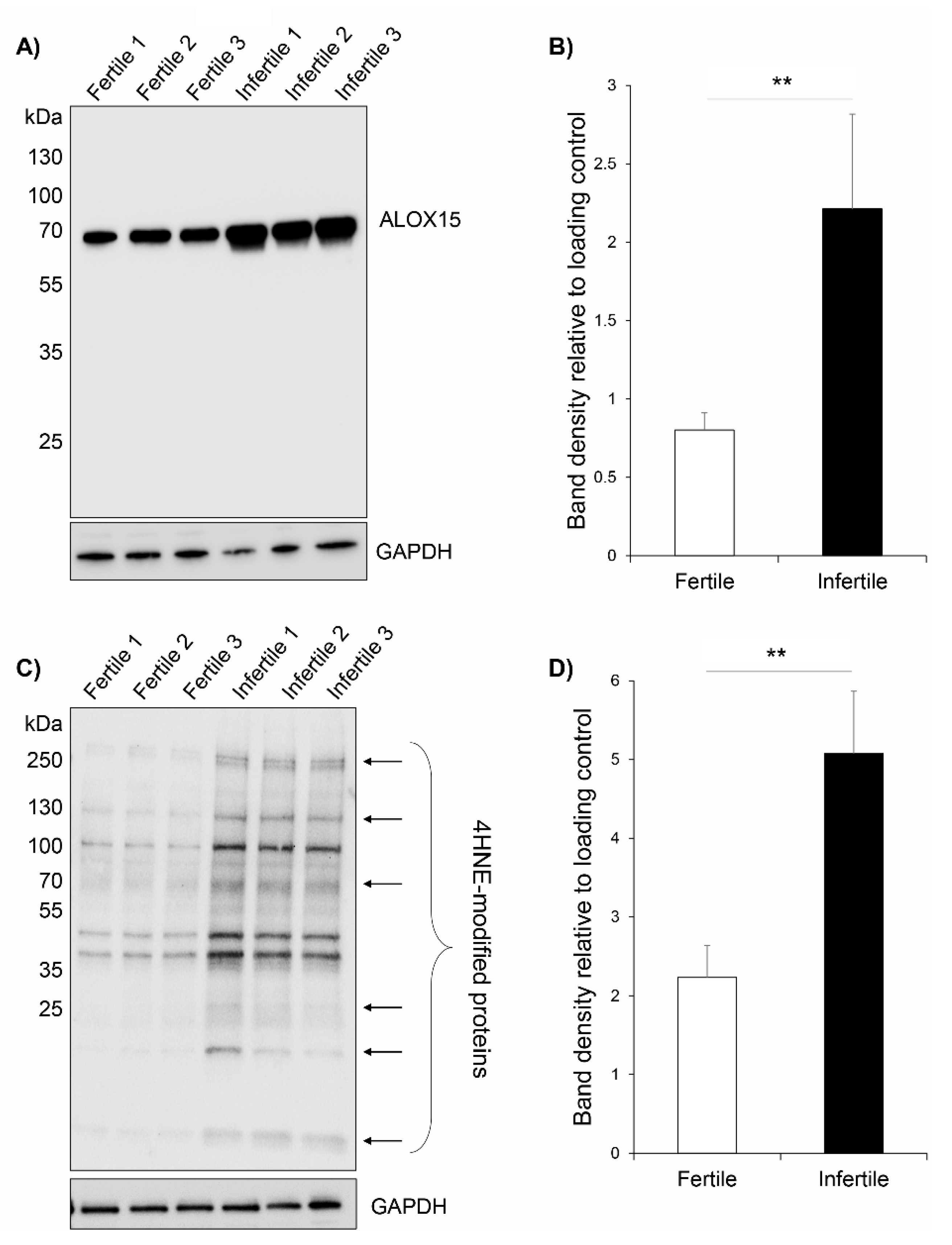
3.5. ALOX15 Protein Expression and 4HNE Protein Modifications Are Elevated in Infertile Patient Sperm Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barratt, C.L.R.; Björndahl, L.; De Jonge, C.J.; Lamb, D.L.; Martini, F.O.; McLachlan, R.; Oates, R.D.; Van der Poel, S.; St John, B.; Sigman, M.; et al. The diagnosis of male infertility: An analysis of the evidence to support the development of global WHO guidance—challenges and future research opportunities. Hum. Reprod. Update 2017, 23, 660–680. [Google Scholar] [CrossRef] [PubMed]
- Oborna, I.; Wojewodka, G.; De Sanctis, J.; Fingerova, H.; Svobodova, M.; Brezinova, J.; Hajduch, M.; Novotny, J.; Radova, L.; Radzioch, D. Increased lipid peroxidation and abnormal fatty acid profiles in seminal and blood plasma of normozoospermic males from infertile couples. Hum. Reprod. 2010, 25, 308–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualotto, F.; Sharma, R.; Kobayashi, H.; Nelson, D.; Agarwal, A. Oxidative stress in normospermic men undergoing infertility evaluation. J. Androl. 2001, 22, 316–322. [Google Scholar] [PubMed]
- Aitken, R.J.; Clarkson, J.S.; Fishel, S. Generation of reactive oxygen species, lipid peroxidation, and human sperm function. Biol. Reprod. 1989, 41, 183–197. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Aitken, R.J.; Wingate, J.K.; De Iuliis, G.N.; McLaughlin, E.A. Analysis of lipid peroxidation in human spermatozoa using BODIPY C11. MHR Basic Sci. Reprod. Med. 2007, 13, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Bromfield, E.G.; Aitken, R.J.; Anderson, A.L.; McLaughlin, E.A.; Nixon, B. The impact of oxidative stress on chaperone-mediated human sperm–egg interaction. Hum. Reprod. 2015, 30, 2597–2613. [Google Scholar] [CrossRef] [Green Version]
- Walters, J.L.; De Iuliis, G.N.; Dun, M.D.; Aitken, R.J.; McLaughlin, E.A.; Nixon, B.; Bromfield, E.G. Pharmacological inhibition of arachidonate 15-lipoxygenase protects human spermatozoa against oxidative stress. Biol. Reprod. 2018, 98, 784–794. [Google Scholar] [CrossRef]
- Walters, J.L.; De Iuliis, G.N.; Nixon, B.; Bromfield, E.G. Oxidative stress in the male germline: A review of novel strategies to reduce 4-hydroxynonenal production. Antioxidants 2018, 7, 132. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence based review. Int. J. Reprod. Biomed. 2016, 14, 729–736. [Google Scholar] [CrossRef]
- Ross, C.; Morriss, A.; Khairy, M.; Khalaf, Y.; Braude, P.; Coomarasamy, A.; El-Toukhy, T. A systematic review of the effect of oral antioxidants on male infertility. Reprod. Biomed. Online 2010, 20, 711–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majzoub, A.; Agarwal, A. Systematic review of antioxidant types and doses in male infertility: Benefits on semen parameters, advanced sperm function, assisted reproduction and live-birth rate. Arab J. Urol. 2018, 16, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromfield, E.G.; Mihalas, B.P.; Dun, M.D.; Aitken, R.J.; McLaughlin, E.A.; Walters, J.L.H.; Nixon, B. Inhibition of arachidonate 15-lipoxygenase prevents 4-hydroxynonenal-induced protein damage in male germ cells. Biol. Reprod. 2017, 96, 598–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suleiman, S.A.; Ali, M.E.; Zaki, Z.; El-Malik, E.; Nasr, M. Lipid peroxidation and human sperm motility: Protective role of vitamin E. J. Androl. 1996, 17, 530–537. [Google Scholar]
- Colagar, A.H.; Pouramir, M.; Marzony, E.T.; Jorsaraei, S.G.A. Relationship between seminal malondialdehyde levels and sperm quality in fertile and infertile men. Braz. Arch. Biol. Technol. 2009, 52, 1387–1392. [Google Scholar] [CrossRef]
- Hermo, L.; Pelletier, R.M.; Cyr, D.G.; Smith, C.E. Surfing the wave, cycle, life history, and genes/proteins expressed by testicular germ cells. Part 1: Background to spermatogenesis, spermatogonia, and spermatocytes. Microsc. Res. Tech. 2010, 73, 241–278. [Google Scholar] [CrossRef]
- Hermo, L.; Pelletier, R.M.; Cyr, D.G.; Smith, C.E. Surfing the wave, cycle, life history, and genes/proteins expressed by testicular germ cells. Part 3: Developmental changes in spermatid flagellum and cytoplasmic droplet and interaction of sperm with the zona pellucida and egg plasma membrane. Microsc. Res. Tech. 2010, 73, 320–363. [Google Scholar] [CrossRef]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.; Heydeck, D.; Hofheinz, K.; Roffeis, J.; O’Donnell, V.B.; Kuhn, H.; Walther, M. Molecular enzymology of lipoxygenases. Arch. Biochem. Biophys. 2010, 503, 161–174. [Google Scholar] [CrossRef]
- Ivanov, I.; Kuhn, H.; Heydeck, D. Structural and functional biology of arachidonic acid 15-lipoxygenase-1 (ALOX15). Gene 2015, 573, 1–32. [Google Scholar] [CrossRef]
- Nixon, B.; Mitchell, L.A.; Anderson, A.L.; Mclaughlin, E.A.; O’bryan, M.K.; Aitken, R.J. Proteomic and functional analysis of human sperm detergent resistant membranes. J. Cell. Physiol. 2011, 226, 2651–2665. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Oshio, S.; Kobanawa, K.; Kobayashi, T.; Mohri, H.; Iizuka, R. Purification of human sperm by a discontinuous percoll density gradient with an innercolumn. Biol. Reprod. 1986, 35, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Geshi, M.; Yamauchi, N.; Nagai, T. Functional changes and motility characteristics of Japanese Black bull spermatozoa separated by percoll. Anim. Reprod. Sci. 2003, 77, 157–172. [Google Scholar] [CrossRef]
- Biggers, J.; Whitten, W.; Whittingham, D. The culture of mouse embryos in vitro. In Methods in Mammalian Embryology; Daniel, J.C., Ed.; W. H. Freeman and Company: San Francisco, CA, USA, 1971; pp. 86–116. [Google Scholar]
- Ackermann, E.J.; Conde-Frieboes, K.; Dennis, E.A. Inhibition of macrophage Ca-independent phospholipase A by bromoenol lactone and trifluoromethyl ketones. J. Biol. Chem. 1995, 270, 445–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, W.; Farooqui, T.; Kokotos, G.; Farooqui, A.A. Synthetic and natural inhibitors of phospholipases A2: Their importance for understanding and treatment of neurological disorders. ACS Chem. Neurosci. 2015, 6, 814–831. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Picardo, M.; Gandini, L.; Dondero, F. Lipids of the sperm plasma membrane: From polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Hum. Reprod. Update 1996, 2, 246–256. [Google Scholar] [CrossRef] [Green Version]
- Roig-Pérez, S.; Guardiola, F.; Moretó, M.; Ferrer, R. Lipid peroxidation induced by DHA enrichment modifies paracellular permeability in Caco-2 cells protective role of taurine. J. Lipid Res. 2004, 45, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Allard, J.P.; Kurian, R.; Aghdassi, E.; Muggli, R.; Royall, D. Lipid peroxidation during n−3 fatty acid and vitamin E supplementation in humans. Lipids 1997, 32, 535–541. [Google Scholar] [CrossRef]
- Vento, R.; D’Alessandro, N.; Giuliano, M.; Lauricella, M.; Carabillo, M.; Tesoriere, G. Induction of apoptosis by arachidonic acid in human retinoblastoma Y79 cells: Involvement of oxidative stress. Exp. Eye Res. 2000, 70, 503–517. [Google Scholar] [CrossRef]
- Das, U. Essential fatty acids enhance free radical generation and lipid peroxidation toinduce apoptosis of tumor cells. Clin. Lipidol. 2011, 6, 463–489. [Google Scholar] [CrossRef]
- Koppers, A.J.; Garg, M.L.; Aitken, R.J. Stimulation of mitochondrial reactive oxygen species production by unesterified, unsaturated fatty acids in defective human spermatozoa. Free Radic. Biol. Med. 2010, 48, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.T.; Lord, T.; Stanger, S.J.; Roman, S.D.; McCluskey, A.; Robinson, P.J.; Aitken, R.J.; Nixon, B. Dynamin regulates specific membrane fusion events necessary for acrosomal exocytosis in mouse spermatozoa. J. Biol. Chem. 2012, 287, 37659–37672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Kühn, H.; Borchert, A. Regulation of enzymatic lipid peroxidation: The interplay of peroxidizing and peroxide reducing enzymes. Free Radic. Biol. Med. 2002, 33, 154–172. [Google Scholar] [CrossRef]
- Aitken, R.; Smith, T.; Lord, T.; Kuczera, L.; Koppers, A.; Naumovski, N.; Connaughton, H.; Baker, M.; De Iuliis, G. On methods for the detection of reactive oxygen species generation by human spermatozoa: Analysis of the cellular responses to catechol oestrogen, lipid aldehyde, menadione and arachidonic acid. Andrology 2013, 1, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Adibhatla, R.M.; Hatcher, J.F. Phospholipase A2, reactive oxygen species, and lipid peroxidation in CNS pathologies. BMB Rep. 2008, 41, 560–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Yu, S.; Wood, W.G.; González, F.A.; Simonyi, A.; Sun, A.Y.; Weisman, G.A. Phospholipase A2 in astrocytes. Mol. Neurobiol. 2005, 31, 27–41. [Google Scholar] [CrossRef]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Walters, J.L.; Gadella, B.M.; Sutherland, J.M.; Nixon, B.; Bromfield, E.G. Male infertility: Shining a light on lipids and lipid-modulating enzymes in the male germline. J. Clin. Med. 2020, 9, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Jiang, L.; Wang, Y.; Yao, B.; Yang, S.; Zhang, B.; Zhang, M. 12/15 Lipoxygenase regulation of colorectal tumorigenesis is determined by the relative tumor levels of its metabolite 12-HETE and 13-HODE in animal models. Oncotarget 2015, 6, 2879–2888. [Google Scholar] [CrossRef] [PubMed]
- Dobrian, A.D.; Lieb, D.C.; Cole, B.K.; Taylor-Fishwick, D.A.; Chakrabarti, S.K.; Nadler, J.L. Functional and pathological roles of the 12-and 15-lipoxygenases. Prog. Lipid Res. 2011, 50, 115–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangaveti, V.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids: Novel regulators of macrophage differentiation and atherogenesis. Adv. Endocrinol. Metab. 2010, 1, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisastra, R.; Dekker, F.J. Inflammation, cancer and oxidative lipoxygenase activity are intimately linked. Cancers 2014, 6, 1500–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Huang, Y.; Li, B.; Xia, X.; Wang, Z. Detection and clinical significance of phospholipase A2 in semen of male infertile patients. Natl. J. Androl. 2003, 9, 90–93. [Google Scholar]
- Sato, H.; Taketomi, Y.; Isogai, Y.; Miki, Y.; Yamamoto, K.; Masuda, S.; Hosono, T.; Arata, S.; Ishikawa, Y.; Ishii, T. Group III secreted phospholipase A2 regulates epididymal sperm maturation and fertility in mice. J. Clin. Investig. 2010, 120, 1400–1414. [Google Scholar] [CrossRef] [Green Version]
- Moawad, A.R.; Fernandez, M.C.; Scarlata, E.; Dodia, C.; Feinstein, S.I.; Fisher, A.B.; O’Flaherty, C. Deficiency of peroxiredoxin 6 or inhibition of its phospholipase A2 activity impair the in vitro sperm fertilizing competence in mice. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Moawad, A.R.; Morielli, T.; Fernandez, M.C.; O’Flaherty, C. Peroxiredoxins prevent oxidative stress during human sperm capacitation. MHR Basic Sci. Reprod. Med. 2017, 23, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Sevenian, A.; Muakkaah-Kelly, S.; Montetruque, S. The influence of phospholipase A2 glutathione peroxidase on the membrane lipid peroxidase. Arch. Biochem. Biophys. 1983, 223, 441–452. [Google Scholar] [CrossRef]
- Kuhn, H.; Banthiya, S.; Van Leyen, K. Mammalian lipoxygenases and their biological relevance. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 308–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Schneider, C.; Boeglin, W.E.; Brash, A.R. Human and mouse eLOX3 have distinct substrate specificities: Implications for their linkage with lipoxygenases in skin. Arch. Biochem. Biophys. 2006, 455, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieg, P.; Fürstenberger, G. The role of lipoxygenases in epidermis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2014, 1841, 390–400. [Google Scholar] [CrossRef] [PubMed]
- García-Herrero, S.; Meseguer, M.; Martínez-Conejero, J.A.; Remohí, J.; Pellicer, A.; Garrido, N. The transcriptome of spermatozoa used in homologous intrauterine insemination varies considerably between samples that achieve pregnancy and those that do not. Fertil. Steril. 2010, 94, 1360–1373. [Google Scholar] [CrossRef]
- Yu, L.; Yang, X.; Ma, B.; Ying, H.; Shang, X.; He, B.; Zhang, Q. Abnormal arachidonic acid metabolic network may reduce sperm motility via P38 MAPK. Open Biol. 2019, 9, 180091. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, S.E.; Tyurina, Y.Y.; Zhao, J.; Croix, C.M.S.; Dar, H.H.; Mao, G.; Tyurin, V.A.; Anthonymuthu, T.S.; Kapralov, A.A.; Amoscato, A.A. PEBP1 wardens ferroptosis by enabling lipoxygenase generation of lipid death signals. Cell 2017, 171, 628–641.e26. [Google Scholar] [CrossRef]
- Bromfield, E.G.; Walters, J.L.; Cafe, S.L.; Bernstein, I.R.; Stanger, S.J.; Anderson, A.L.; Aitken, R.J.; McLaughlin, E.A.; Dun, M.D.; Gadella, B.M. Differential cell death decisions in the testis: Evidence for an exclusive window of ferroptosis in round spermatids. MHR Basic Sci. Reprod. Med. 2019, 25, 241–256. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walters, J.L.H.; Anderson, A.L.; Martins da Silva, S.J.; Aitken, R.J.; De Iuliis, G.N.; Sutherland, J.M.; Nixon, B.; Bromfield, E.G. Mechanistic Insight into the Regulation of Lipoxygenase-Driven Lipid Peroxidation Events in Human Spermatozoa and Their Impact on Male Fertility. Antioxidants 2021, 10, 43. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010043
Walters JLH, Anderson AL, Martins da Silva SJ, Aitken RJ, De Iuliis GN, Sutherland JM, Nixon B, Bromfield EG. Mechanistic Insight into the Regulation of Lipoxygenase-Driven Lipid Peroxidation Events in Human Spermatozoa and Their Impact on Male Fertility. Antioxidants. 2021; 10(1):43. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010043
Chicago/Turabian StyleWalters, Jessica L. H., Amanda L. Anderson, Sarah J. Martins da Silva, R. John Aitken, Geoffry N. De Iuliis, Jessie M. Sutherland, Brett Nixon, and Elizabeth G. Bromfield. 2021. "Mechanistic Insight into the Regulation of Lipoxygenase-Driven Lipid Peroxidation Events in Human Spermatozoa and Their Impact on Male Fertility" Antioxidants 10, no. 1: 43. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010043