Total Phenol Content and Antioxidant Activity of Different Celta Pig Carcass Locations as Affected by the Finishing Diet (Chestnuts or Commercial Feed)

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design and Pig Slaughter
2.2. Finishing Diet Material
2.3. Extract Preparations
2.4. Determination of Total Phenol Content (TPC)
2.5. Determination of Total Flavonoids
2.6. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity
2.7. 2-2′-Azino-di-[3-Ethylbenzthiazoline Sulfonate] (ABTS) Radical Scavenging Activity
2.8. Ferric Reducing Antioxidant Power (FRAP) Assay
2.9. Statistical Analysis
3. Results and Discussion
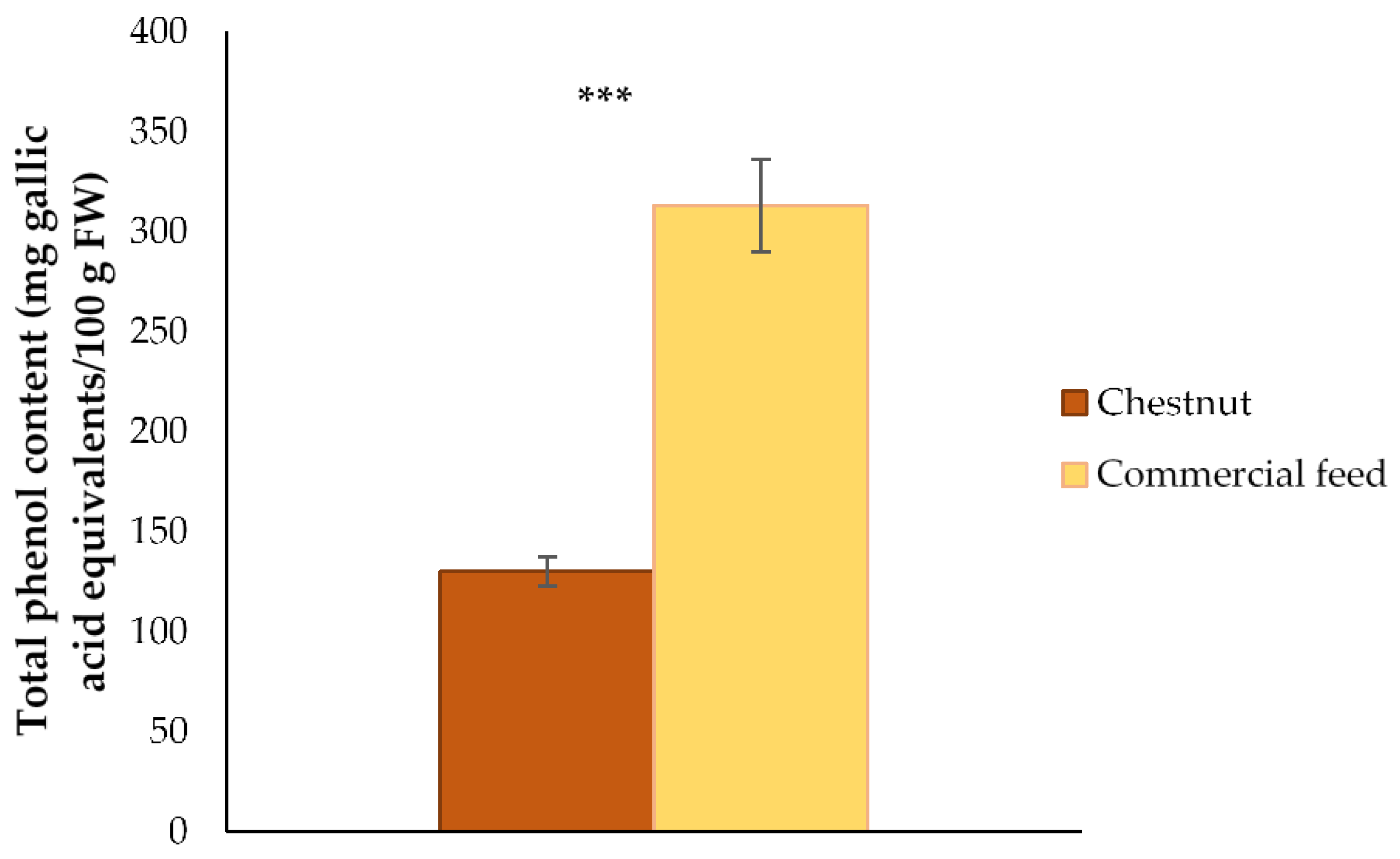
3.1. Total Phenol Content of Feed Used in the Finishing Diet of Celta Pigs
3.2. Total Flavonoids of Feed Used in the Finishing Diet of Celta Pigs
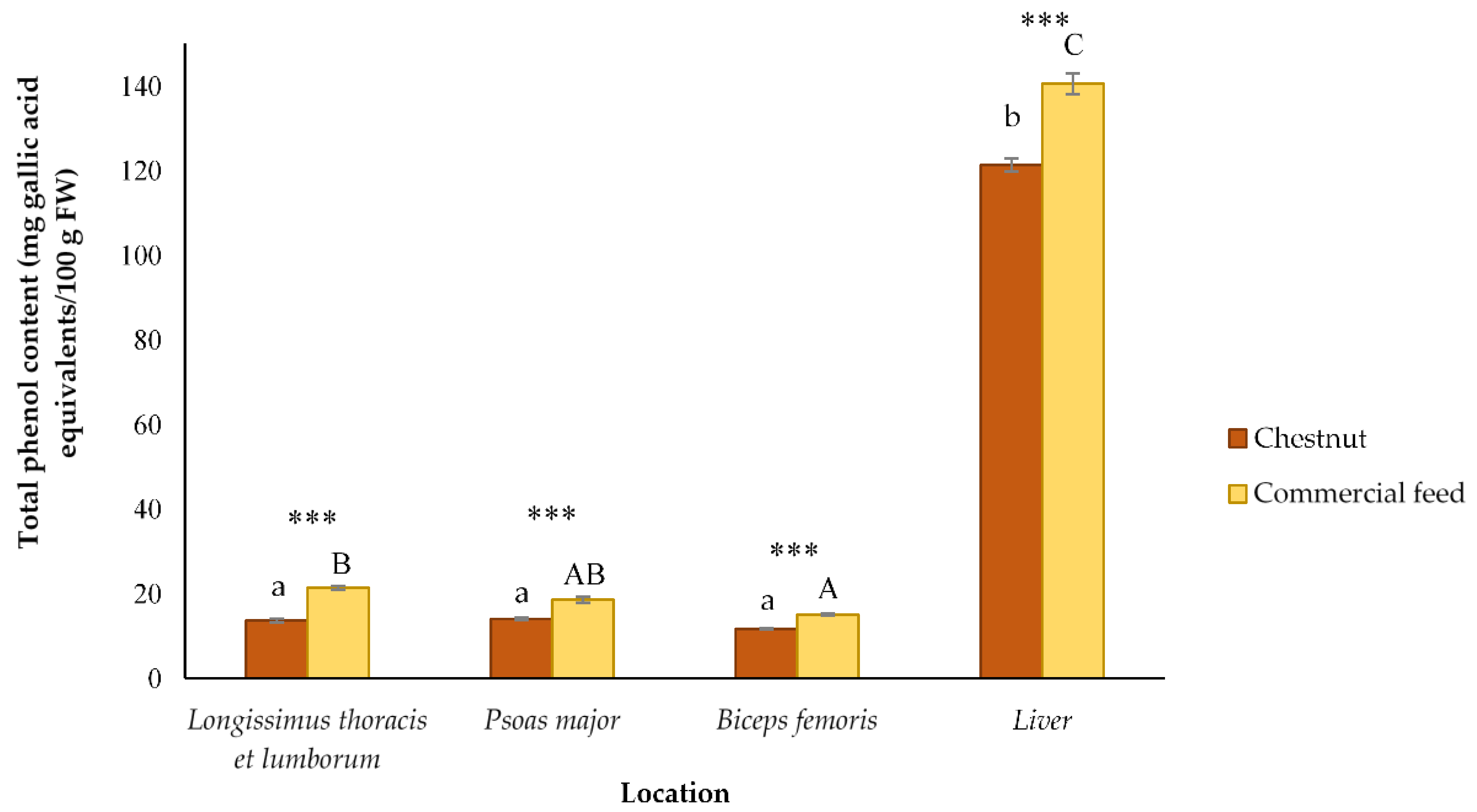
3.3. Total Phenol Content of Different Celta Pig Locations
3.4. Antioxidant Activity of Feed Used in the Finishing Diet of Celta Pigs
3.5. Correlation Analysis in Feed Used in the Finishing Diet of Celta Pigs
3.6. Antioxidant Activity of Different Celta Pig Locations
3.7. Correlation Analysis of Different Celta Pig Locations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moure, A.; Cruz, J.M.; Franco, D.; Domínguez, M.J.; Sineiro, J.; Domínguez, H.; Núñez, M.J.; Parajó, J.C. Natural antioxidants from residual sources. Food Chem. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Rodríguez-Carpena, J.G.; Morcuende, D.; Estévez, M. Avocado by-products as inhibitors of color deterioration and lipid and protein oxidation in raw porcine patties subjected to chilled storage. Meat Sci. 2011, 89, 166–173. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [Green Version]
- Devatkal, S.K.; Thorat, P.; Manjunatha, M. Effect of vacuum packaging and pomegranate peel extract on quality aspects of ground goat meat and nuggets. J. Food Sci. Technol. 2014, 51, 2685–2691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, M.; Lorenzo, J.M. Effect of packaging conditions on shelf-life of fresh foal meat. Meat Sci. 2012, 91, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.P.P.; Trindade, M.A.; Tonin, F.G.; Pugine, S.M.P.; Lima, C.G.; Lorenzo, J.M.; de Melo, M.P. Evaluation of oxidative stability of lamb burger with Origanum vulgare extract. Food Chem. 2017, 233, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Zinoviadou, K.G.; Koutsoumanis, K.P.; Biliaderis, C.G. Physico-chemical properties of whey protein isolate films containing oregano oil and their antimicrobial action against spoilage flora of fresh beef. Meat Sci. 2009, 82, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.P.P.; Trindade, M.A.; Lorenzo, J.M.; de Melo, M.P. Assessment of the stability of sheep sausages with the addition of different concentrations of Origanum vulgare extract during storage. Meat Sci. 2018, 137, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Pateiro, M.; Domínguez, R.; Barba, F.J.; Putnik, P.; Kovačević, D.B.; Shpigelman, A.; Granato, D.; Franco, D. Berries extracts as natural antioxidants in meat products: A review. Food Res. Int. 2018, 106, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Nunes, D.S.; Barba, F.J. An integrated strategy between food chemistry, biology, nutrition, pharmacology, and statistics in the development of functional foods: A proposal. Trends Food Sci. Technol. 2017, 62, 13–22. [Google Scholar] [CrossRef]
- Pateiro, M.; Lorenzo, J.M.M.; Amado, I.R.R.; Franco, D. Effect of addition of green tea, chestnut and grape extract on the shelf-life of pig liver pâté. Food Chem. 2014, 147, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Lanigan, R.S.; Yamarik, T.A.; Andersen, F.A. Final report on the safety assessment of BHT. J. Am. Coll. Toxicol. 2011, 1, 19–94. [Google Scholar]
- Thatoi, H.; Patra, J.K. Biotechnology and pharmacological evaluation of medicinal plants: An overview. J. Herbs Spices Med. Plants 2011, 17, 214–248. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Rocchetti, G.; Pateiro, M.; Lucini, L.; Domínguez, R.; Lorenzo, J.M. Addition of plant extracts to meat and meat products to extend shelf-life and health-promoting attributes: An overview. Curr. Opin. Food Sci. 2020, 31, 81–87. [Google Scholar] [CrossRef]
- Hernández Suárez, M.; Rodríguez Galdón, B.; Ríos Mesa, D.; Díaz Romero, C.; Rodríguez Rodríguez, E. Sugars, organic acids and total phenols in varieties of chestnut fruits from Tenerife (Spain). Food Nutr. Sci. 2012, 3, 705–715. [Google Scholar]
- De Vasconcelos, M.d.C.B.M.; Bennett, R.N.; Quideau, S.; Jacquet, R.; Rosa, E.A.S.; Ferreira-Cardoso, J.V. Evaluating the potential of chestnut (Castanea sativa Mill.) fruit pericarp and integument as a source of tocopherols, pigments and polyphenols. Ind. Crops Prod. 2010, 31, 301–311. [Google Scholar] [CrossRef]
- Vázquez, G.; Fernández-Agulló, A.; Gómez-Castro, C.; Freire, M.S.; Antorrena, G.; González-Álvarez, J. Response surface optimization of antioxidants extraction from chestnut (Castanea sativa) bur. Ind. Crops Prod. 2012, 35, 126–134. [Google Scholar] [CrossRef]
- Nazzaro, M.; Barbarisi, C.; La Cara, F.; Volpe, M.G. Chemical and biochemical characterisation of an IGP ecotype chestnut subjected to different treatments. Food Chem. 2011, 128, 930–936. [Google Scholar] [CrossRef]
- Simonetti, A.; Gambacorta, E.; Perna, A. Antioxidative and antihypertensive activities of pig meat before and after cooking and in vitro gastrointestinal digestion: Comparison between Italian autochthonous pig Suino Nero Lucano and a modern crossbred pig. Food Chem. 2016, 212, 590–595. [Google Scholar] [CrossRef]
- Simonetti, A.; Perna, A.; Gambacorta, E. Comparison of antioxidant compounds in pig meat from Italian autochthonous pig Suino Nero Lucano and a modern crossbred pig before and after cooking. Food Chem. 2019, 292, 108–112. [Google Scholar] [CrossRef]
- Bermúdez, R.; Franco, I.; Franco, D.; Carballo, J.; Lorenzo, J.M. Influence of inclusion of chestnut in the finishing diet on fatty acid profile of dry-cured ham from Celta pig breed. Meat Sci. 2012, 92, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Montes, R.; Purriños, L.; Cobas, N.; Franco, D. Fatty acid composition of Celta pig breed as influenced by sex and location of fat in the carcass. J. Sci. Food Agric. 2012, 92, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Martínez, S.; Gómez, M.; Carballo, J.; Franco, I. Fatty acids, retinol and cholesterol composition in various fatty tissues of Celta pig breed: Effect of the use of chestnuts in the finishing diet. J. Food Compos. Anal. 2015, 37, 104–111. [Google Scholar] [CrossRef]
- Pugliese, C.; Sirtori, F.; Acciaioli, A.; Bozzi, R.; Campodoni, G.; Franci, O. Quality of fresh and seasoned fat of Cinta Senese pigs as affected by fattening with chestnut. Meat Sci. 2013, 93, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Temperan, S.; Lorenzo, J.M.J.M.; Castiñeiras, B.D.; Franco, I.; Carballo, J.; Castsiñeiras, B.; Franco, I.; Carballo, J. Carcass and meat quality traits of Celta heavy pigs. Effect of the inclusion of chestnuts in the finishing diet. Span. J. Agric. Res. 2014, 12, 694–707. [Google Scholar] [CrossRef] [Green Version]
- Díaz, O.; Ros, C.; Veiga, A.; Cobos, Á. Including chestnuts and sugar beet pulp in diets for pigs: The effects on the quality of pork meat and the sensory properties of dry-cured sausage (chorizo gallego). J. Muscle Foods 2009, 20, 449–464. [Google Scholar] [CrossRef]
- Cobos, Á.; Veiga, A.; Díaz, O. Influencia de un pienso con castañas y pulpa de remolacha azucarera en la composición lipídica del lacón gallego. Grasas y Aceites 2008, 59, 121–127. [Google Scholar] [CrossRef]
- Echegaray, N.; Paterio, M.; Domínguez, R.; Purriños, L.; Bermúdez, R.; Carballo, J.; Lorenzo, J.M. Effects of different cooking methods and of the inclusion of chestnut (Castanea sativa Miller) in the finishing diet of Celta pig breed on the physicochemical parameters and volatile profile of Longissimus thoracis et lumborum muscle. Food Res. Int. 2020, 137, 109407. [Google Scholar] [CrossRef]
- Echegaray, N.; Gómez, B.; Barba, F.J.; Franco, D.; Estévez, M.; Carballo, J.; Marszałek, K.; Lorenzo, J.M. Chestnuts and by-products as source of natural antioxidants in meat and meat products: A review. Trends Food Sci. Technol. 2018, 82, 110–121. [Google Scholar] [CrossRef]
- Gómez, M.; Fonseca, S.; Cachaldora, A.; Carballo, J.; Franco, I. Effect of chestnuts intake by Celta pigs on lipolytic, oxidative and fatty acid profile changes during ripening and vacuum-packed storage of Galician “chorizo”. J. Food Compos. Anal. 2017, 56, 73–83. [Google Scholar] [CrossRef]
- Gómez, M.; Cachaldora, A.; Fonseca, S.; Domínguez, R.; Carballo, J.; Franco, I. Biochemical, oxidative, and lipolytic changes during vacuum-packed storage of dry-cured loin: Effect of chestnuts intake by celta pigs. J. Food Qual. 2018, 2018, 7690501. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, D.O. Polyphenols and the human brain: Plant “secondary metabolite” ecologic roles and endogenous signaling functions drive benefits. Adv. Nutr. 2014, 5, 515–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, M.; Morcuende, D.; Cava, R. Extensively reared Iberian pigs versus intensively reared white pigs for the manufacture of frankfurters. Meat Sci. 2006, 72, 356–364. [Google Scholar] [CrossRef] [PubMed]
- European Union Council Directive 2008/120/EC laying down minimum standards for the protection of pigs. Off. J. Eur. Union 2008, 47, 5–13.
- Santos, J.; Mendiola, J.A.; Oliveira, M.B.P.P.; Ibáñez, E.; Herrero, M. Sequential determination of fat- and water-soluble vitamins in green leafy vegetables during storage. J. Chromatogr. 2012, 1261, 179–188. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Rodrigues, F.; Palmeira-de-Oliveira, A.; das Neves, J.; Sarmento, B.; Amaral, M.H.; Oliveira, M.B. Medicago spp. extracts as promising ingredients for skin care products. Ind. Crops Prod. 2013, 49, 634–644. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH-assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.S.; Karathanos, V.T. Nutritional evaluation and health promoting activities of nuts and seeds cultivated in Greece. Int. J. Food Sci. Nutr. 2013, 64, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Neri, L.; Dimitri, G.; Sacchetti, G. Chemical composition and antioxidant activity of cured chestnuts from three sweet chestnut (Castanea sativa Mill.) ecotypes from Italy. J. Food Compos. Anal. 2010, 23, 23–29. [Google Scholar] [CrossRef]
- Özel, H.B. The effects of origin difference on some chemical properties of the fruit of chestnut (Castanea sativa Mill.). Rom. Biotechnol. Lett. 2015, 20, 10969–10975. [Google Scholar]
- Carocho, M.; Antonio, A.L.; Barros, L.; Bento, A.; Luisa Botelho, M.; Kaluska, I.; Ferreira, I.C.F.R. Comparative effects of gamma and electron beam irradiation on the antioxidant potential of Portuguese chestnuts (Castanea sativa Mill.). Food Chem. Toxicol. 2012, 50, 3452–3455. [Google Scholar] [CrossRef]
- Otles, S.; Selek, I. Effect of processing on the phenolic content and antioxidant activity of chestnuts. Qual. Assur. Saf. Crop. Foods 2012, 4, 3–11. [Google Scholar] [CrossRef]
- Tejerina, D.; García-Torres, S.; Cabeza De Vaca, M.; Vázquez, F.M.; Cava, R. Effect of production system on physical-chemical, antioxidant and fatty acids composition of Longissimus dorsi and Serratus ventralis muscles from Iberian pig. Food Chem. 2012, 133, 293–299. [Google Scholar] [CrossRef]
- González, E.; Tejeda, J.F. Effects of dietary incorporation of different antioxidant extracts and free-range rearing on fatty acid composition and lipid oxidation of Iberian pig meat. Animal 2007, 1, 1060–1067. [Google Scholar] [CrossRef] [Green Version]
- Baik, B.K.; Ullrich, S.E. Barley for food: Characteristics, improvement, and renewed interest. J. Cereal Sci. 2008, 48, 233–242. [Google Scholar] [CrossRef]
- Van Hung, P. Phenolic compounds of cereals and their antioxidant capacity. Crit. Rev. Food Sci. Nutr. 2014, 56, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Nordkvist, E.; Salomonssona, A.-C.; Aman, P. Distribution of insoluble bound phenolic acids in barley grain. J. Sci. Food Agric. 1984, 35, 657–661. [Google Scholar] [CrossRef]
- Bendelow, V.M.; LaBerge, D.E. Relationships among barley, malt, and beer phenolics. J. Am. Soc. Brew. Chem. 1979, 37, 89–90. [Google Scholar] [CrossRef]
- Suriano, S.; Iannucci, A.; Codianni, P.; Fares, C.; Russo, M.; Pecchioni, N.; Marciello, U.; Savino, M. Phenolic acids profile, nutritional and phytochemical compounds, antioxidant properties in colored barley grown in southern Italy. Food Res. Int. 2018, 113, 221–233. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, J.; Cai, S.; Chen, X.; Quan, X.; Zhang, G. Association mapping for total polyphenol content, total flavonoid content and antioxidant activity in barley. BMC Genom. 2018, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Martinez, L.X.; Parkin, K.L.; Garcia, H.S. Phase II-Inducing, polyphenols content and antioxidant capacity of corn (Zea mays L.) from phenotypes of white, blue, red and purple colors processed into masa and tortillas. Plant Foods Hum. Nutr. 2011, 66, 41–47. [Google Scholar] [CrossRef]
- Saikaew, K.; Lertrat, K.; Meenune, M.; Tangwongchai, R. Effect of high-pressure processing on colour, phytochemical contents and antioxidant activities of purple waxy corn (Zea mays L. var. ceratina) kernels. Food Chem. 2017, 243, 328–337. [Google Scholar] [CrossRef]
- Antonio, A.L.; Fernandes, Â.; Barreira, J.C.M.; Bento, A.; Botelho, M.L.; Ferreira, I.C.F.R. Influence of gamma irradiation in the antioxidant potential of chestnuts (Castanea sativa Mill.) fruits and skins. Food Chem. Toxicol. 2011, 49, 1918–1923. [Google Scholar] [CrossRef]
- Živković, J.; Zeković, Z.; Mujić, I.; Tumbas, V.; Cvetković, D.; Spasojević, I. Antioxidant properties of phenolics in Castanea sativa Mill. extracts. Food Technol. Biotechnol. 2009, 47, 421–427. [Google Scholar]
- Dinis, L.T.; Oliveira, M.M.; Almeida, J.; Costa, R.; Gomes-Laranjo, J.; Peixoto, F. Antioxidant activities of chestnut nut of Castanea sativa Mill. (cultivar “Judia”) as function of origin ecosystem. Food Chem. 2012, 132, 1–8. [Google Scholar] [CrossRef]
- Tejerina, D.; García-Torres, S.; Cabeza de Vaca, M.; Vázquez, F.M.; Cava, R. Study of variability in antioxidant composition and fatty acids profile of Longissimus dorsi and Serratus ventralis muscles from Iberian pigs reared in two different Montanera seasons. Meat Sci. 2012, 90, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Flis, M.; Sobotka, W.; Antoszkiewicz, Z.; Lipiński, K.; Zduńczyk, Z. The effect of grain polyphenols and the addition of vitamin E to diets enriched with α-linolenic acid on the antioxidant status of pigs. J. Anim. Feed Sci. 2010, 19, 539–553. [Google Scholar] [CrossRef] [Green Version]
- González, E.; Tejeda, J.F.; Motilva, M.J.; Romero, M.P. Phenolic compounds in subcutaneous adipose tissue from Iberian pigs. Options Méditerranéennes Ser. A 2003, 118, 115–118. [Google Scholar]
- Perna, A.; Simonetti, A.; Grassi, G.; Gambacorta, E. Effect of a cauliflower (Brassica oleraceae var. Botrytis) leaf powder-enriched diet on performance, carcass and meat characteristics of growing rabbit. Meat Sci. 2018, 149, 134–140. [Google Scholar]
- Jang, A.; Liu, X.D.; Shin, M.H.; Lee, B.D.; Lee, S.K.; Lee, J.H.; Jo, C. Antioxidative potential of raw breast meat from broiler chicks fed a dietary medicinal herb extract mix. Poult. Sci. 2008, 87, 2382–2389. [Google Scholar] [CrossRef]
- Yu, H.C.; Hsu, J.-L.L.; Chang, C.-I.I.; Tan, F.-J.J. Antioxidant properties of porcine liver proteins hydrolyzed using Monascus purpureus. Food Sci. Biotechnol. 2017, 26, 1217–1225. [Google Scholar] [CrossRef]
- Kakimov, A.; Suychinov, A.; Tsoy, A.; Mustambayev, N.; Ibragimov, N.; Kuderinova, N.; Mirasheva, G.; Yessimbekov, Z. Nutritive and biological value of liver and blood of various slaughtered animals. J. Pharm. Res. Int. 2018, 22, 1–5. [Google Scholar] [CrossRef]
- Rey, A.I.; López-Bote, C.J.; Litta, G. Effects of dietary vitamin E (DL-α-tocopheryl acetate) and vitamin C combination on piglets oxidative status and immune response at weaning. J. Anim. Feed Sci. 2017, 26, 226–235. [Google Scholar] [CrossRef]
- Amazan, D.; Cordero, G.; López-Bote, C.J.; Lauridsen, C.; Rey, A.I. Effects of oral micellized natural vitamin E (d-α-tocopherol) v. synthetic vitamin E (dl-α-tocopherol) in feed on α-tocopherol levels, stereoisomer distribution, oxidative stress and the immune response in piglets. Animal 2014, 8, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Rey, A.I.; Lopez-Bote, C.J.; Kerry, J.P.; Lynch, P.B.; Buckley, D.J.; Morrissey, P.A. Modification of lipid composition and oxidation in porcine muscle and muscle microsomes as affected by dietary supplementation of n-3 with either n-9 or n-6 fatty acids and α-tocopheryl acetate. Anim. Feed Sci. Technol. 2004, 113, 223–238. [Google Scholar] [CrossRef]
- Abe, L.T.; Lajolo, F.M.; Genovese, M.I. Comparison of phenol content and antioxidant capacity of nuts. Ciencia e Tecnol. Alimentar 2010, 30, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Blomhoff, R.; Carlsen, M.H.; Andersen, L.F.; Jacobs, D.R. Health benefits of nuts: Potential role of antioxidants. Br. J. Nutr. 2006, 96, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.I.R.N.A.; Nunes, F.M.; Gonalves, B.; Bennett, R.N.; Silva, A.P. Effect of cooking on total vitamin C contents and antioxidant activity of sweet chestnuts (Castanea sativa Mill.). Food Chem. 2011, 128, 165–172. [Google Scholar] [CrossRef]
- Smet, K.; Raes, K.; de Smet, S. Novel approaches in measuring the antioxidative potential of animal feeds: The FRAP and DPPH methods. J. Sci. Food Agric. 2006, 86, 2412–2416. [Google Scholar] [CrossRef]
- Yang, X.J.; Dang, B.; Fan, M.T. Free and bound phenolic compound content and antioxidant activity of different cultivated blue highland barley varieties from the qinghai-tibet plateau. Molecules 2018, 23, 879. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Sotero, M.Y.; Cruz-Hernández, C.D.; Trujillo-Carretero, C.; Rodríguez-Dorantes, M.; García-Galindo, H.S.; Chávez-Servia, J.L.; Oliart-Ros, R.M.; Guzmán-Gerónimo, R.I. Antioxidant and antiproliferative activity of blue corn and tortilla from native maize. Chem. Cent. J. 2017, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Ventura, J.; Castro, C.; Boone, V.; Rojas, R.; Ascacio-Valdés, J.; Martínez-Ávila, G. UPLC-ESI-QTOF-MS2-based identification and antioxidant activity assessment of phenolic compounds from red corn Cob (Zea mays L.). Molecules 2018, 23, 1425. [Google Scholar] [CrossRef] [Green Version]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Živković, J.; Zeković, Z.; Mujić, I.; Vidović, S.; Cvetković, D.; LepoJević, Ž.; Nikolić, G.; Trutić, N. Scavenging capacity of superoxide radical and screening of antimicrobial activity of Castanea sativa Mill. extracts. Czech J. Food Sci. 2010, 28, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Dudonné, S.; Vitrac, X.; Coutiére, P.; Woillez, M.; Mérillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Todorovic, V.; Milenkovic, M.; Vidovic, B.; Todorovic, Z.; Sobajic, S. Correlation between antimicrobial, antioxidant activity, and polyphenols of alkalized/nonalkalized cocoa powders. J. Food Sci. 2017, 82, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Olivares, A.; Rey, A.I.; Daza, A.; Lopez-Bote, C.J. High dietary vitamin A interferes with tissue -tocopherol concentrations in fattening pigs: A study that examines administration and withdrawal times. Animal 2009, 3, 1264–1270. [Google Scholar] [CrossRef]
- Rey, A.I.; Segura, J.F.; Castejón, D.; Fernández-Valle, E.; Cambero, M.I.; Calvo, L. Vitamin D3 supplementation in drinking water prior to slaughter improves oxidative status, physiological stress, and quality of pork. Antioxidants 2020, 9, 559. [Google Scholar] [CrossRef]
- Echegaray, N.; Pateiro, M.; Zhang, W.; Domínguez, R.; Campagnol, P.C.B.; Carballo, J.; Lorenzo, J.M. Influence of the Inclusion of chestnut (Castanea sativa Miller) in the finishing diet and cooking technique on the physicochemical parameters and volatile profile of biceps femoris muscle. Foods 2020, 9, 754. [Google Scholar] [CrossRef]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, α-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef]
- Prevolnik, M.; Škrlep, M.; Brus, M.; Pugliese, C.; Čandek-Potokar, M.; Škorjanc, D. Supplementing pig diet with 0.2% sweet chestnut (Castanea sativa Mill.) wood extract had no effect on growth, carcass or meat quality. Acta Agric. Slov. 2012, 100, 83–88. [Google Scholar]
- Echegaray, N.; Domínguez, R.; Franco, D.; Lorenzo, J.M.; Carballo, J. Effect of the use of chestnuts (Castanea sativa Miller) in the finishing diet of Celta pig breed on the shelf-life of meat refrigerated and frozen. Food Res. Int. 2018, 114, 114–122. [Google Scholar] [CrossRef]




{kind=link}
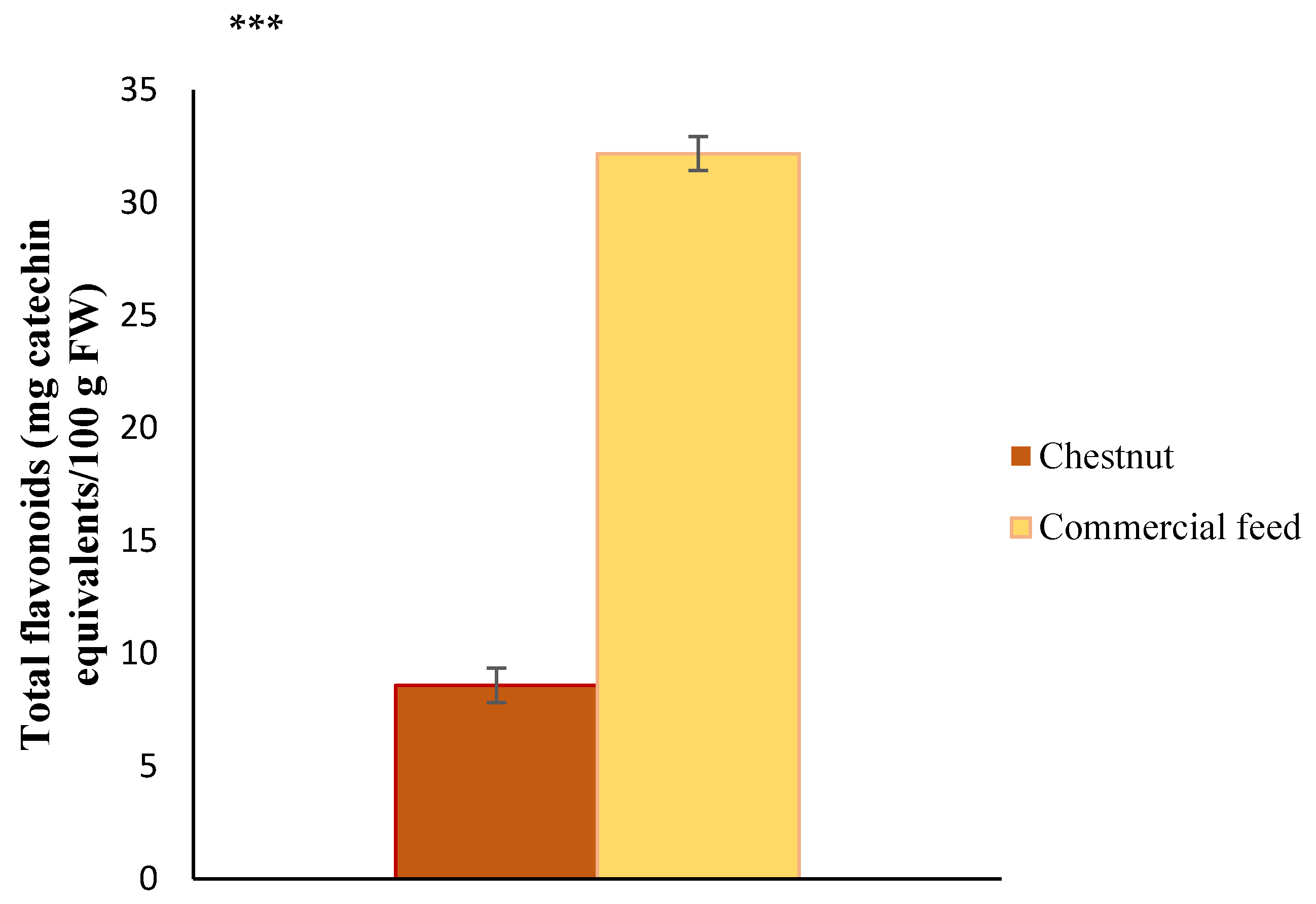{kind=link}
{kind=link}
| Component | (%) | Supplements | UI (Vitamins) or mg (Minerasls)/kg |
|---|---|---|---|
| Crude protein | 16.01 | Vitamin A | 6500 |
| Crude fiber | 5.01 | Vitamin D3 | 1500 |
| Crude fat | 2.98 | Vitamin E | 15 |
| Crude ashes | 5.83 | Manganese sulphate | 40 |
| Lys | 0.72 | Zinc oxide | 60 |
| Met | 0.25 | Iron sulphate | 50 |
| Thr | 0.12 | Copper sulphate | 15 |
| Trp | 0.01 | Potassium iodide | 0.5 |
| Ca | 0.88 | Sodium selenite | 0.1 |
| P | 0.61 | ||
| Na | 0.17 |
| Chestnut | Commercial Feed | SEM | F | |
|---|---|---|---|---|
| Antioxidant Activity | ||||
| DPPH (µg TE/g FW) | 1152.42 ± 89.84 | 957.33 ± 101.50 | 69.878 | ns |
| ABTS (µg TE/g FW | 9379.74 ± 183.61 | 7613.44 ± 139.96 | 241.704 | *** |
| FRAP (µmol Fe2+/100 g FW) | 1777.49 ± 124.72 | 1946.09 ± 24.45 | 64.953 | ns |
| ABTS | FRAP | TPC | TF | |
|---|---|---|---|---|
| Chestnut | ||||
| DPPH | −0.832 ** | 0.847 ** | 0.794 * | 0.895 ** |
| ABTS | −0.861 ** | −0.838 ** | −0.834 ** | |
| FRAP | 0.974 ** | 0.973 ** | ||
| TPC | 0.903 ** | |||
| Commercial Feed | ||||
| DPPH | 0.379 | 0.483 | 0.298 | −0.066 |
| ABTS | 0.432 | 0.960 ** | 0.061 | |
| FRAP | 0.358 | −0.413 | ||
| TPC | 0.092 | |||
| Location | Longissimus thoracis et Lumborum | Psoas major | Biceps femoris | Liver | SEM | L | LxF |
|---|---|---|---|---|---|---|---|
| Antioxidant Activity | |||||||
| DPPH (µg TE/g FW) | |||||||
| Chestnut pigs | 85.77 ± 3.27 a | 97.55 ± 2.38 a | 97.42 ± 2.02 a | 669.08 ± 18.03 b | 29.927 | *** | *** |
| Commercial feed pigs | 124.23 ± 2.78 a | 125.76 ± 2.55 a | 131.78 ± 2.16 a | 811.64 ± 12.91 b | 35.329 | *** | |
| SEM | 3.879 | 2.939 | 3.248 | 16.265 | |||
| F | *** | *** | *** | *** | |||
| ABTS (µg TE/g FW) | |||||||
| Chestnut pigs | 503.94 ± 19.90 b | 625.83 ± 19.35 c | 343.76 ± 10.37 a | 1889.67 ± 58.32 d | 74.579 | *** | *** |
| Commercial feed pigs | 932.10 ± 30.71 b | 831.21 ± 45.42 b | 395.15 ± 14.27 a | 2047.10 ± 67.58 c | 75.456 | *** | |
| SEM | 40.430 | 29.889 | 9.717 | 45.960 | |||
| F | *** | *** | ** | ns | |||
| FRAP (µmol Fe2+/100 g FW) | |||||||
| Chestnut pigs | 41.58 ± 1.40 a | 53.19 ± 0.87 a | 52.64 ± 1.32 a | 659.46 ± 7.66 b | 31.429 | *** | ns |
| Commercial feed pigs | 54.96 ± 1.26 a | 63.06 ± 0.98 a | 59.09 ± 0.78 a | 692.52 ± 13.00 b | 32.714 | *** | |
| SEM | 1.464 | 1.055 | 0.932 | 7.942 | |||
| F | *** | *** | *** | * | |||
| ABTS | FRAP | TPC | |
|---|---|---|---|
| Chestnut Pigs | |||
| Longissimus thoracis et lumborum | |||
| DPPH | 0.488 * | 0.401 | 0.222 |
| ABTS | 0.304 | 0.621 ** | |
| FRAP | 0.549 * | ||
| Psoas major | |||
| DPPH | 0.243 | 0.389 | 0.143 |
| ABTS | −0.020 | 0.637 ** | |
| FRAP | 0.393 | ||
| Biceps femoris | |||
| DPPH | 0.545 * | 0.447 | 0.527 * |
| ABTS | 0.045 | 0.413 | |
| FRAP | 0.303 | ||
| Liver | |||
| DPPH | −0.171 | 0.535 * | −0.336 |
| ABTS | −0.470 * | 0.628 ** | |
| FRAP | −0.351 | ||
| Commercial Feed Pigs | |||
| Longissimus thoracis et lumborum | |||
| DPPH | −0.045 | 0.164 | 0.008 |
| ABTS | 0.526 * | 0.783 ** | |
| FRAP | 0.688 ** | ||
| Psoas major | |||
| DPPH | 0.354 | 0.495 * | 0.444 |
| ABTS | 0.711 ** | 0.918 ** | |
| FRAP | 0.633 ** | ||
| Biceps femoris | |||
| DPPH | 0.224 | 0.386 | −0.426 |
| ABTS | −0.033 | −0.257 | |
| FRAP | −0.232 | ||
| Liver | |||
| DPPH | −0.089 | 0.395 | −0.003 |
| ABTS | −0.387 | 0.433 | |
| FRAP | −0.153 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Echegaray, N.; Munekata, P.E.S.; Centeno, J.A.; Domínguez, R.; Pateiro, M.; Carballo, J.; Lorenzo, J.M. Total Phenol Content and Antioxidant Activity of Different Celta Pig Carcass Locations as Affected by the Finishing Diet (Chestnuts or Commercial Feed). Antioxidants 2021, 10, 5. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010005
Echegaray N, Munekata PES, Centeno JA, Domínguez R, Pateiro M, Carballo J, Lorenzo JM. Total Phenol Content and Antioxidant Activity of Different Celta Pig Carcass Locations as Affected by the Finishing Diet (Chestnuts or Commercial Feed). Antioxidants. 2021; 10(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010005
Chicago/Turabian StyleEchegaray, Noemí, Paulo E. S. Munekata, Juan A. Centeno, Rubén Domínguez, Mirian Pateiro, Javier Carballo, and José M. Lorenzo. 2021. "Total Phenol Content and Antioxidant Activity of Different Celta Pig Carcass Locations as Affected by the Finishing Diet (Chestnuts or Commercial Feed)" Antioxidants 10, no. 1: 5. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010005