
Calcium Dobesilate Reverses Cognitive Deficits and Anxiety-Like Behaviors in the D-Galactose-Induced Aging Mouse Model through Modulation of Oxidative Stress


 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Body Weight and Behavioral Tests
2.3. Oxidative Stress Evaluation
2.4. Statistical Analysis
3. Results
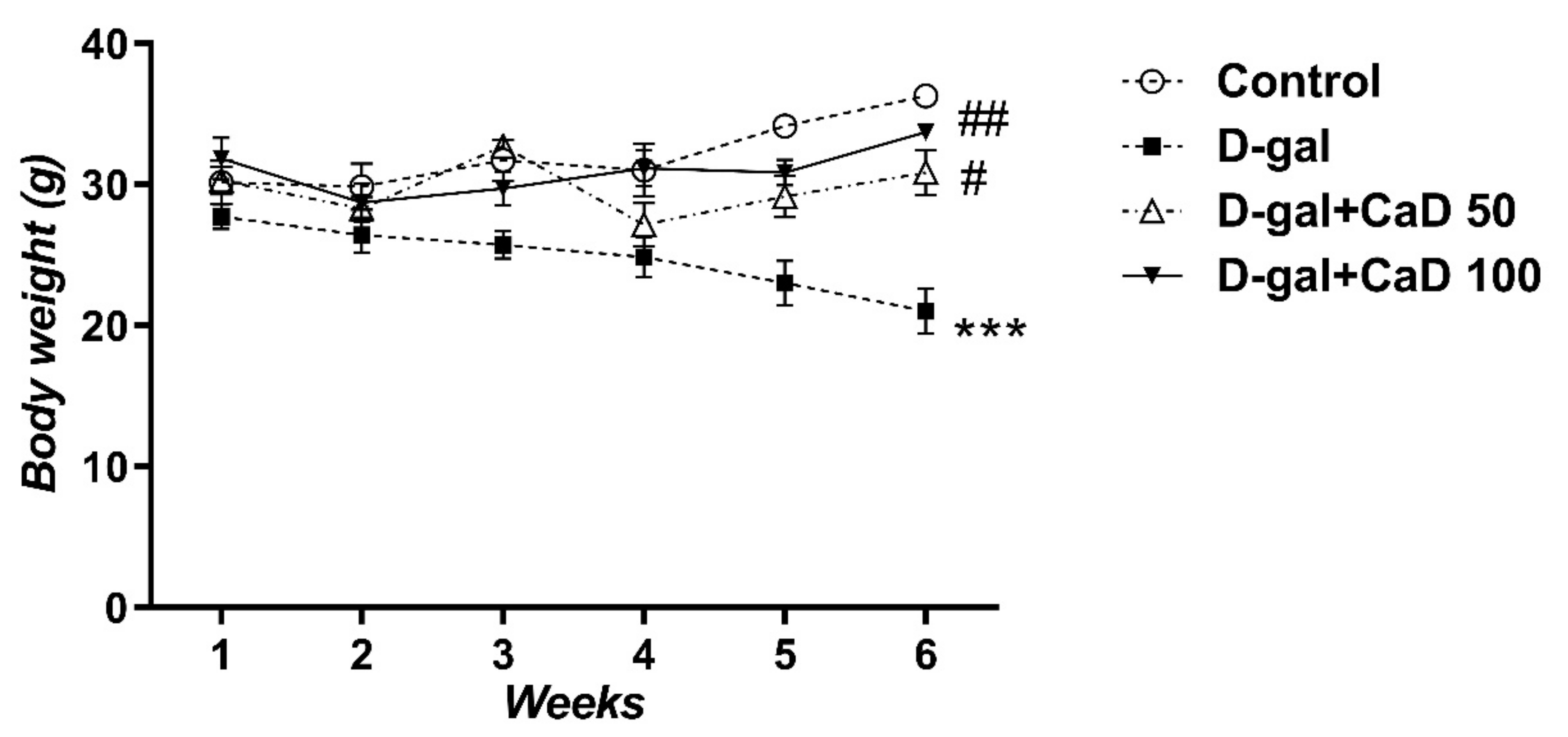
3.1. CaD Inhibited the Body Weight Loss Induced by D-Gal Treatment
3.2. CaD Decreased the Anxiety-Like Behavior in D-Gal Treated Mice
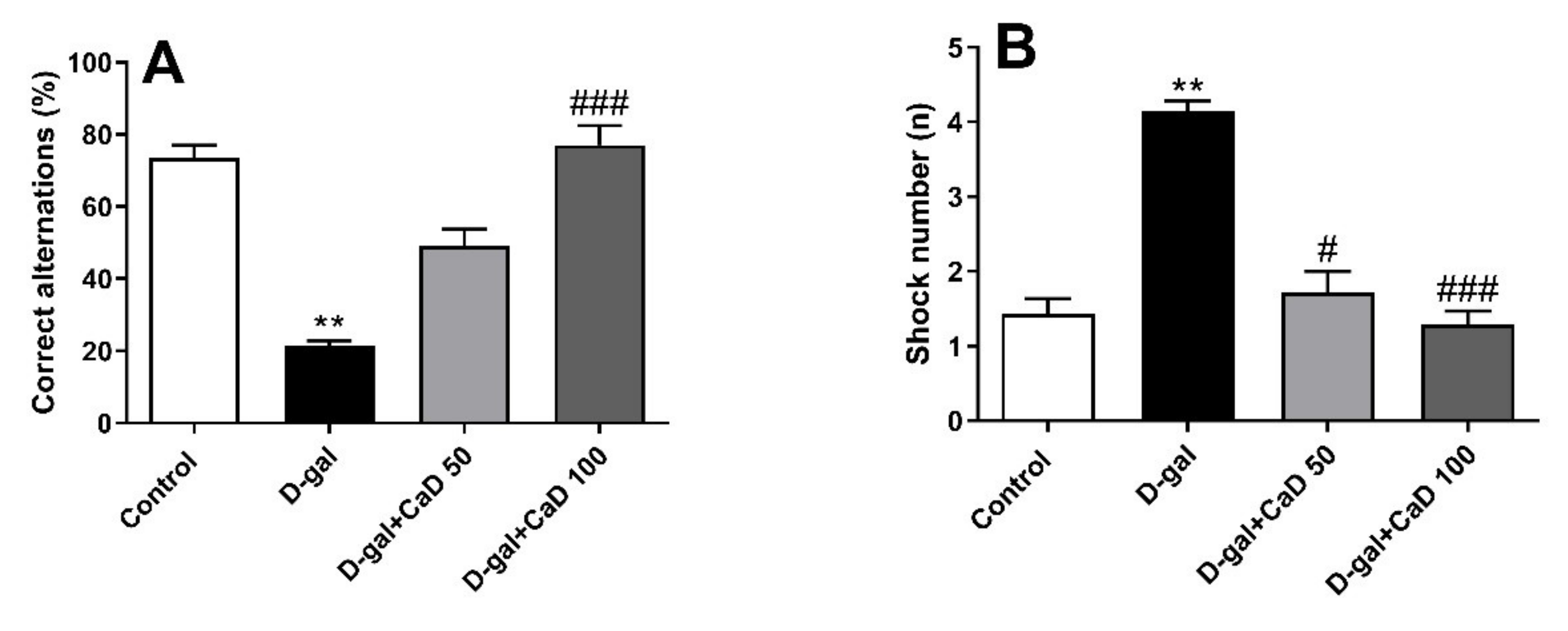
3.3. CaD Attenuated the Cognitive Impairments in D-Gal Treated Mice
3.4. CaD Reduced the Oxidative Stress s in D-Gal Treated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shin, K.R.; Kim, M.Y.; Kim, Y.H. Study on the lived experience of aging. Nurs. Health Sci. 2003, 5, 245–252. [Google Scholar] [CrossRef]
- Kowalska, M.; Piekut, T.; Prendecki, M.; Sodel, A.; Kozubski, W.; Dorszewska, J. Mitochondrial and nuclear DNA oxidative damage in physiological and pathological aging. DNA Cell. Biol. 2020, 39, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Forster, M.J.; Dubey, A.; Dawson, K.M.; Stutts, W.A.; Lal, H.; Sohal, R.S. Age-related losses of cognitive function and motor skills in mice are associated with oxidative protein damage in the brain. Proc. Natl. Acad. Sci. USA 1996, 93, 4765–4769. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Dong, B.; Pang, N.; Hu, J. ROS generation and DNA damage contribute to abamectin-induced cytotoxicity in mouse macrophage cells. Chemosphere 2019, 234, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef] [PubMed]
- Aydın, S.; Yanar, K.; Atukeren, P.; Dalo, E.; Sitar, M.E.; Uslu, E.; Caf, N.; Cakatay, U. Comparison of oxidative stress biomarkers in renal tissues of D-galactose induced, naturally aged and young rats. Biogerontology 2012, 13, 251–260. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, Y.; Xu, X.; Xiang, H.; Shi, Y.; Gao, Y.; Wang, X.; Jiang, X.; Li, N.; Pan, J. Autophagy inhibits the mesenchymal stem cell aging induced by D-galactose through ROS/JNK/p38 signalling. Clin. Exp. Pharmacol. Physiol. 2020, 47, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F.; Zhou, B.H. Leonurine ameliorates D-galactose-induced aging in mice through activation of the Nrf2 signalling pathway. Aging 2019, 11, 7339. [Google Scholar] [CrossRef]
- Baeta-Corral, R.; Castro-Fuentes, R.; Giménez-Llort, L. Sexual dimorphism in the behavioral responses and the immuno-endocrine status in D-galactose induced aging. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1147–1157. [Google Scholar] [CrossRef]
- Ho, S.C.; Liu, J.H.; Wu, R.Y. Establishment of the mimetic aging effect in mice caused by D-galactose. Biogerontology 2003, 4, 15–18. [Google Scholar] [CrossRef]
- Farsa, O. Calcium dobesilate in prevention and treatment of diabetic retinopathy. Diabet. Retin. 2012, 356. [Google Scholar]
- Liu, J.; Li, S.; Sun, D. Calcium dobesilate and micro-vascular diseases. Life Sci. 2019, 22, 348–353. [Google Scholar] [CrossRef]
- Bazmandegan, G.; Fatemi, I.; Kaeidi, A.; Khademalhosseini, M.; Fathinejad, A.; Amirteimoury, M. Calcium dobesilate prevents cisplatin-induced nephrotoxicity by modulating oxidative and histopathological changes in mice. Naunyn. Schmiedebergs. Arch. Pharmacol. 2020, 394, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Unal, Y.; Tuncal, S.; Kosmaz, K.; Kucuk, B.; Kismet, K.; Cavusoglu, T.; Celepli, P.; Senes, M.; Yildiz, S.; Hucumenoglu, S. The effect of calcium dobesilate on liver damage in experimental obstructive jaundice. J. Investig. Surg. 2019, 32, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.Y.; Jiang, Y.; Wang, R.N.; Hao, L.L.; Xu, D. Evaluation of calcium dobesilate for its anti-cataract potential in experimental rat models. Chin. J. Ophthalmol. 2013, 49, 927–933. [Google Scholar]
- Constantin, G.; Della Bianca, V.; Pietronigro, E.C.; Zenaro, E.; Piacentino, G.; Nagarajan, R.; Toffali, L.; Mirenda, M.; Bauer, J. Treatment with calcium dobesilate reduces neuroinflammation and improves cognition in a mouse model of alzheimer’s disease. Alzheimer’s Dement. 2017, 13, P321. [Google Scholar] [CrossRef]
- Kaviani, E.; Rahmani, M.; Kaeidi, A.; Shamsizadeh, A.; Allahtavakoli, M.; Mozafari, N.; Fatemi, I. Protective effect of atorvastatin on d-galactose-induced aging model in mice. Behav. Brain Res. 2017, 334, 55–60. [Google Scholar] [CrossRef]
- Fatemi, I.; Delrobaee, F.; Bahmani, M.; Shamsizadeh, A.; Allahtavakoli, M. The effect of the anti-diabetic drug metformin on behavioral manifestations associated with ovariectomy in mice. Neurosci. Lett. 2019, 690, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 2004, 28, 497–505. [Google Scholar] [CrossRef]
- Goudarzi, M.; Mombeini, M.A.; Fatemi, I.; Aminzadeh, A.; Kalantari, H.; Nesari, A.; Najafzadehvarzi, H.; Mehrzadi, S. Neuroprotective effects of Ellagic acid against acrylamide-induced neurotoxicity in rats. Neurol. Res. 2019, 41, 419–428. [Google Scholar] [CrossRef]
- Sadigh-Eteghad, S.; Majdi, A.; McCann, S.K.; Mahmoudi, J.; Vafaee, M.S.; Macleod, M.R. Correction: D-galactose-induced brain ageing model: A systematic review and meta-analysis on cognitive outcomes and oxidative stress indices. PLoS ONE. 2017, 12, e0190328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Therrien, Z.; Hunsley, J. Assessment of anxiety in older adults: A systematic review of commonly used measures. Aging Ment. Health 2012, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Liaquat, L.; Shahzad, S.; Sadir, S.; Madiha, S.; Batool, Z.; Tabassum, S.; Saleem, S.; Naqvi, F.; Perveen, T. A high dose of short term exogenous D-galactose administration in young male rats produces symptoms simulating the natural aging process. Life Sci. 2015, 124, 110–119. [Google Scholar] [CrossRef]
- Haider, S.; Naqvi, F.; Batool, Z.; Tabassum, S.; Sadir, S.; Liaquat, L.; Naqvi, F.; Zuberi, N.A.; Shakeel, H.; Perveen, T. Pretreatment with curcumin attenuates anxiety while strengthens memory performance after one short stress experience in male rats. Brain Res. Bul. 2015, 115, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, G.; Farajdokht, F.; Mohaddes, G.; Babri, S.; Ebrahimi, V.; Ebrahimi, H. Garlic (Allium sativum) improves anxiety-and depressive-related behaviors and brain oxidative stress in diabetic rats. Arch. Physiol. Biochem. 2020, 126, 95–100. [Google Scholar] [CrossRef]
- Samad, N.; Saleem, A. Administration of Allium cepa L. bulb attenuates stress-produced anxiety and depression and improves memory in male mice. Metabolic Brain Dis. 2018, 33, 271–281. [Google Scholar] [CrossRef]
- Cerrahoglu, M.; Kurdal, A.T.; Iskesen, I.; Onur, E.; Sirin, H. Calcium dobesilate reduces oxidative stress in cardiac surgery. J. Cardiovasc. Surg. 2009, 50, 695. [Google Scholar]
- Sola-Adell, C.; Bogdanov, P.; Hernandez, C.; Sampedro, J.; Valeri, M.; Garcia-Ramirez, M.; Pasquali, C.; Simó, R. Calcium dobesilate prevents neurodegeneration and vascular leakage in experimental diabetes. Curr. Eye Res. 2017, 42, 1273–1286. [Google Scholar] [CrossRef]
- Jafarey, M.; Changizi Ashtiyani, S.; Najafi, H. Calcium dobesilate for prevention of gentamicin-induced nephrotoxicity in rats. Iran. J. Kidney Dis. 2014, 8, 46–52. [Google Scholar] [PubMed]
- Zhang, X.; Liu, W.; Wu, S.; Jin, J.; Li, W.; Wang, N. Calcium dobesilate for diabetic retinopathy: A systematic review and meta-analysis. Sci. China Life Sci. 2015, 58, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, I.; Khaluoi, A.; Kaeidi, A.; Shamsizadeh, A.; Heydari, S.; Allahtavakoli, M.A. Protective effect of metformin on D-galactose-induced aging model in mice. Iran. J. Basic Med. Sci. 2018, 21, 19–25. [Google Scholar] [PubMed]
- Fatemi, I.; Heydari, S.; Kaeidi, A.; Shamsizadeh, A.; Hakimizadeh, E.; Khaluoi, A.; Allahtavakoli, M. Metformin ameliorates the age-related changes of d-galactose administration in ovariectomized mice. Fundam. Clin. Pharmacol. 2018, 32, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Rej, S.; Saleem, M.; Herrmann, N.; Stefatos, A.; Rau, A.; Lanctot, K.L. Serum low-density lipoprotein levels; statin use, and cognition in patients with coronary artery disease. Neuropsych. Dis. Treat. 2016, 12, 2913–2920. [Google Scholar] [CrossRef] [Green Version]
- Pak, J.W.; Herbst, A.; Bua, E.; Gokey, N.; McKenzie, D.; Aiken, J.M. Mitochondrial DNA mutations as a fundamental mechanism in physiological declines associated with aging. Aging Cell. 2003, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Van Velzen, L.S.; Wijdeveld, M.; Black, C.N.; van Tol, M.J.; van der Wee, N.J.; Veltman, D.J.; Penninx, B.W.J.H.; Schmaal, L. Oxidative stress and brain morphology in individuals with depression, anxiety and healthy controls. Progr. Neuropsychopharmacol. Biol. Psych. 2017, 76, 140–144. [Google Scholar] [CrossRef]
- Seker, A.; Bardakci, O.; Eryilmaz, S.; Kocarslan, S.; Incebiyik, A.; Yucel, Y.; Taskin, A.; Soyalp, M.; Gokalp, O.; Uzunkoy, A. Does calcium dobesilate protect against intestinal ischemia-reperfusion injury induced in rats? Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2168–2173. [Google Scholar]
- Szabo, M.E.; Haines, D.; Garay, E.; Chiavaroli, C.; Farine, J.C.; Hannaert, P.; Berta, A.; Garay, R.P. Antioxidant properties of calcium dobesilate in ischemic/reperfused diabetic rat retina. Eur. J. Pharmacol. 2001, 428, 277–286. [Google Scholar] [CrossRef]
- Sun, J.; Wang, B.; Hao, Y.; Yang, X. Effects of calcium dobesilate on Nrf2, Keap1 and HO-1 in the lenses of D-galactose-induced cataracts in rats. Exp. Ther. Med. 2018, 15, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Malinski, T. Nitric oxide and nitroxidative stress in Alzheimer’s disease. J. Alzheimers Dis. 2007, 11, 207–218. [Google Scholar] [CrossRef]
- Sobocanec, S.; Balog, T.; Sverko, V.; Marotti, T. Sex-dependent antioxidant enzyme activities and lipid peroxidation in ageing mouse brain. Free Radic. Res. 2003, 37, 743–748. [Google Scholar] [CrossRef]
- Picón-Pagès, P.; Garcia-Buendia, J.; Muñoz, F.J. Functions and dysfunctions of nitric oxide in brain. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1949–1967. [Google Scholar] [CrossRef]
- Snyder, S.H. Janus faces of nitric oxide. Nature 1993, 364, 577. [Google Scholar] [CrossRef] [PubMed]
- Ledo, A.; Lourenço, C.F.; Cadenas, E.; Barbosa, R.M.; Laranjinha, J. The bioactivity of neuronal-derived nitric oxide in aging and neurodegeneration: Switching signaling to degeneration. Free Red. Biol. Med. 2021, 162, 500–513. [Google Scholar] [CrossRef]
- Banji, D.; Banji, O.J.; Dasaroju, S.; Kranthi, K.C. Curcumin and piperine abrogate lipid and protein oxidation induced by D-galactose in rat brain. Brain Res. 2013, 1515, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Suschek, C.; Kolb, H.; Kolb-Bachofen, V. Dobesilate enhances endothelial nitric oxide synthase-activity in macro- and microvascular endothelial cells. British. J. Pharmacol. 1997, 122, 1502–1508. [Google Scholar] [CrossRef] [PubMed]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, M.; Zhu, Z.; Wen, Z.; Ke, S. Impairments of tight junctions are involved in D-galactose-induced brain aging. Neuroreport 2013, 24, 671–676. [Google Scholar] [CrossRef]
- González-Marrero, I.; Giménez-Llort, L.; Johanson, C.E.; Carmona-Calero, E.M.; Castañeyra-Ruiz, L.; Brito-Armas, J.M.; Castañeyra-Perdomo, A.; Castro-Fuentes, R. Choroid plexus dysfunction impairs beta-amyloid clearance in a triple transgenic mouse model of Alzheimer’s disease. Front. Cell. Neurosci. 2015, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Simó-Servat, O.; Solà-Adell, C.; Bogdanov, P.; Hernández, C.; Simó, R. Mechanisms of retinal neuroprotection of calcium dobesilate: Therapeutic implications. Neural Regen. Res. 2017, 12, 1620. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yuan, J.; Qi, C.; Shao, X.; Mou, S.; Ni, Z. Calcium dobesilate may alleviate diabetes-induced endothelial dysfunction and inflammation. Mol. Med. Rep. 2017, 16, 8635–8642. [Google Scholar] [CrossRef]
- Liu, J.; Chen, D.; Wang, Z.; Chen, C.; Ning, D.; Zhao, S. Protective effect of walnut on d-galactose-induced aging mouse model. Food Sci. Nutr. 2019, 7, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hakimizadeh, E.; Zamanian, M.; Giménez-Llort, L.; Sciorati, C.; Nikbakhtzadeh, M.; Kujawska, M.; Kaeidi, A.; Hassanshahi, J.; Fatemi, I. Calcium Dobesilate Reverses Cognitive Deficits and Anxiety-Like Behaviors in the D-Galactose-Induced Aging Mouse Model through Modulation of Oxidative Stress. Antioxidants 2021, 10, 649. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050649
Hakimizadeh E, Zamanian M, Giménez-Llort L, Sciorati C, Nikbakhtzadeh M, Kujawska M, Kaeidi A, Hassanshahi J, Fatemi I. Calcium Dobesilate Reverses Cognitive Deficits and Anxiety-Like Behaviors in the D-Galactose-Induced Aging Mouse Model through Modulation of Oxidative Stress. Antioxidants. 2021; 10(5):649. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050649
Chicago/Turabian StyleHakimizadeh, Elham, Mohammad Zamanian, Lydia Giménez-Llort, Clara Sciorati, Marjan Nikbakhtzadeh, Małgorzata Kujawska, Ayat Kaeidi, Jalal Hassanshahi, and Iman Fatemi. 2021. "Calcium Dobesilate Reverses Cognitive Deficits and Anxiety-Like Behaviors in the D-Galactose-Induced Aging Mouse Model through Modulation of Oxidative Stress" Antioxidants 10, no. 5: 649. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050649