
Galangin Resolves Cardiometabolic Disorders through Modulation of AdipoR1, COX-2, and NF-κB Expression in Rats Fed a High-Fat Diet

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Research Designs
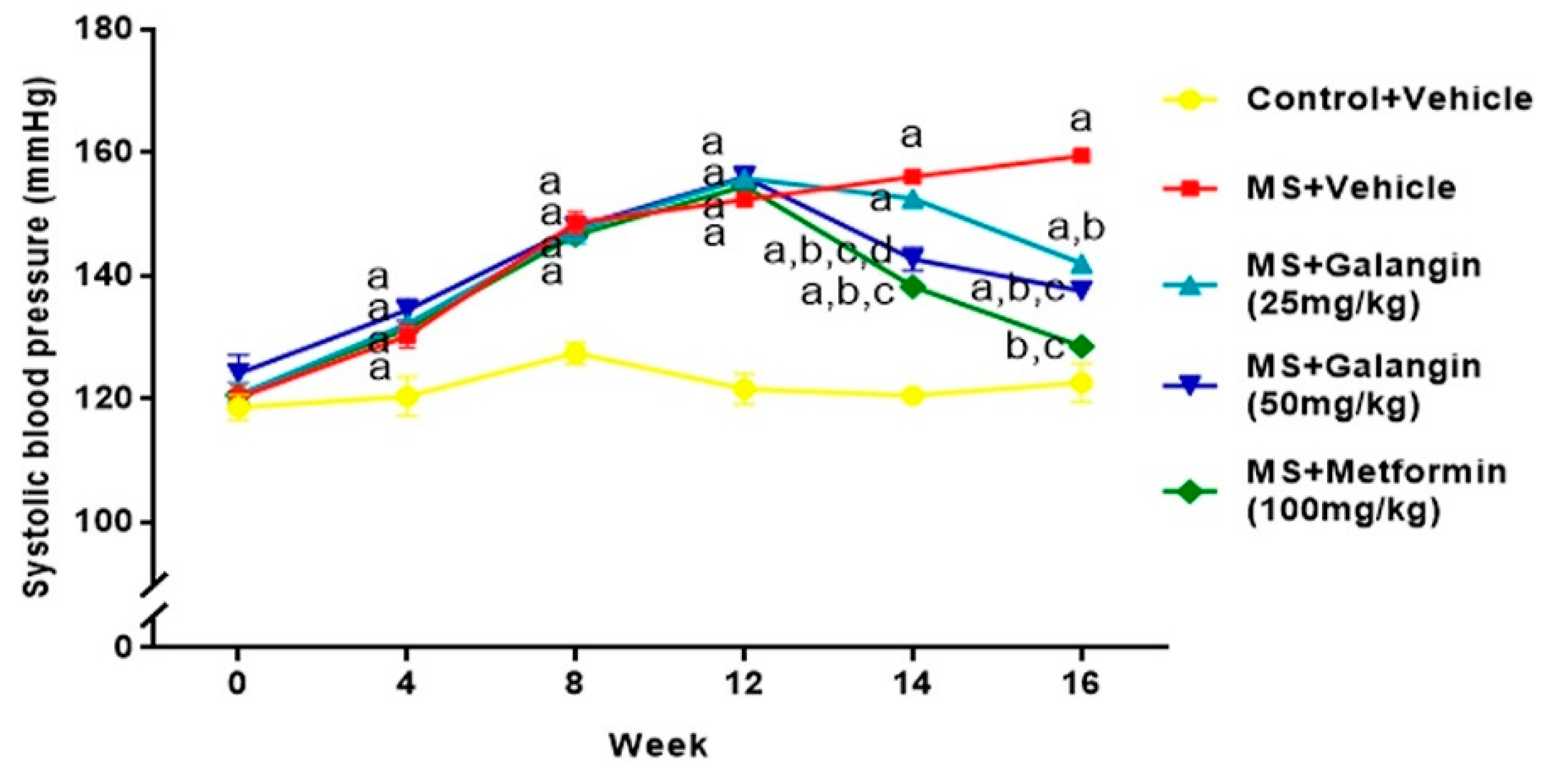
2.3. Measurement of Systolic Blood Pressure
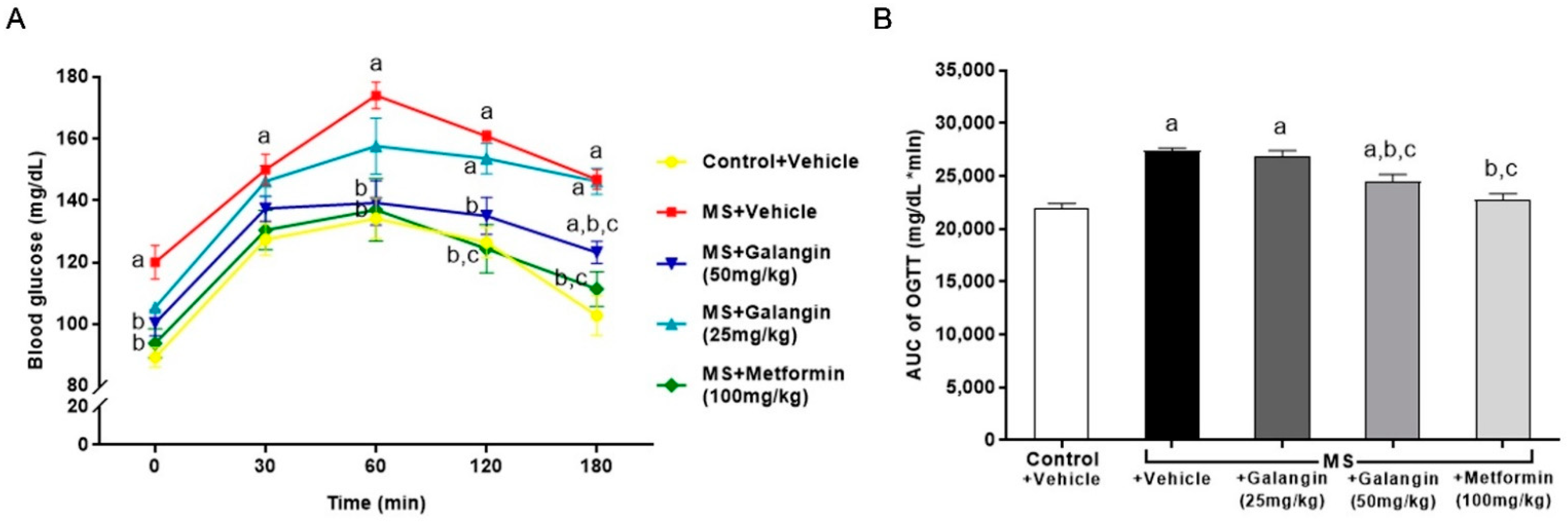
2.4. Measurements of Fasting Blood Glucose, Serum Insulin Level, and the Oral Glucose Tolerance Test
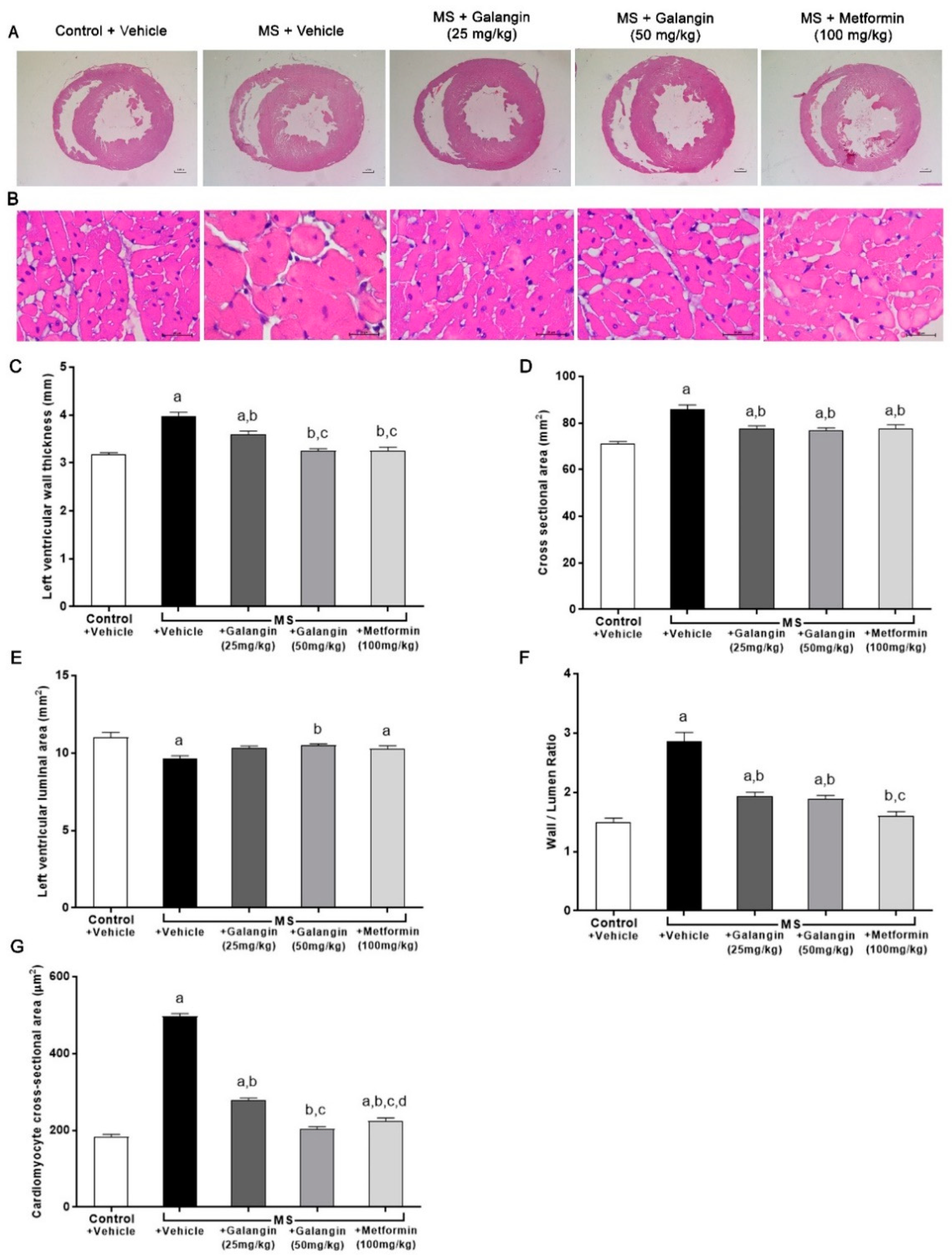
2.5. Echocardiography
2.6. Direct Blood Pressure Measurements
2.7. Assessment of Biochemical Profiles
2.8. Tissue Harvesting
2.9. Hematoxylin and Eosin Staining of Cardiac and Fat Tissue
2.10. Immunohistochemical Staining of Myocardial Sections
2.11. Assays of Cytokine Levels
2.12. Oxidative Stress Marker Assessment
2.13. Antioxidant Endogenous Enzyme Activity Assessment
2.14. Western Blotting Analysis
2.15. Statistical Analysis
3. Results
3.1. Effects of Galangin on Body and Organ Weight in Metabolic Syndrome Rats
3.2. Effects of Galangin on Metabolic Parameters
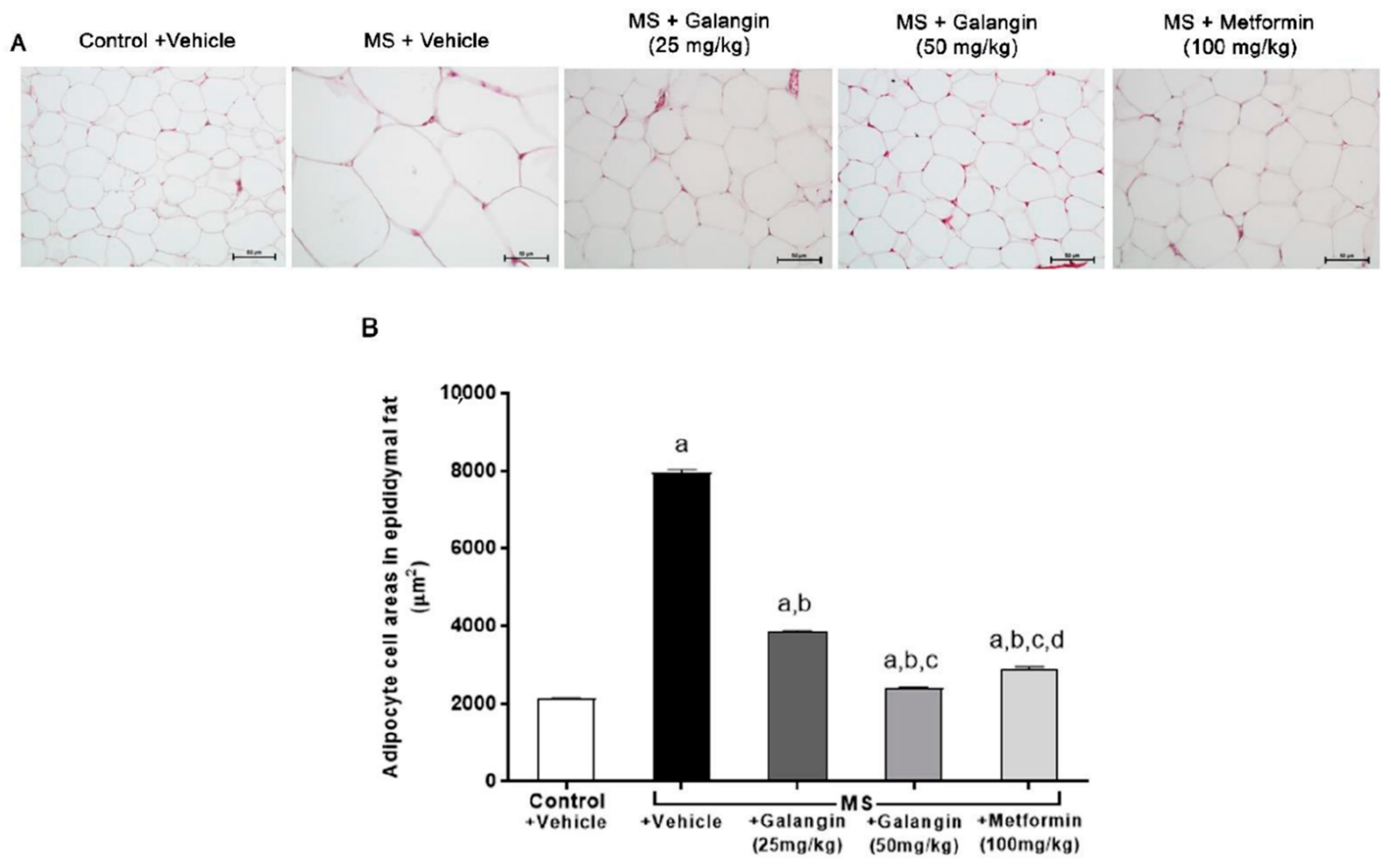
3.3. Effects of Galangin on Epididymal Fat Pads’ Morphology
3.4. Effects of Galangin on Blood Pressure and Heart Rate
3.5. Effects of Galangin on Cardiac Function Parameters
3.6. Effects of Galangin on Cardiac Morphology
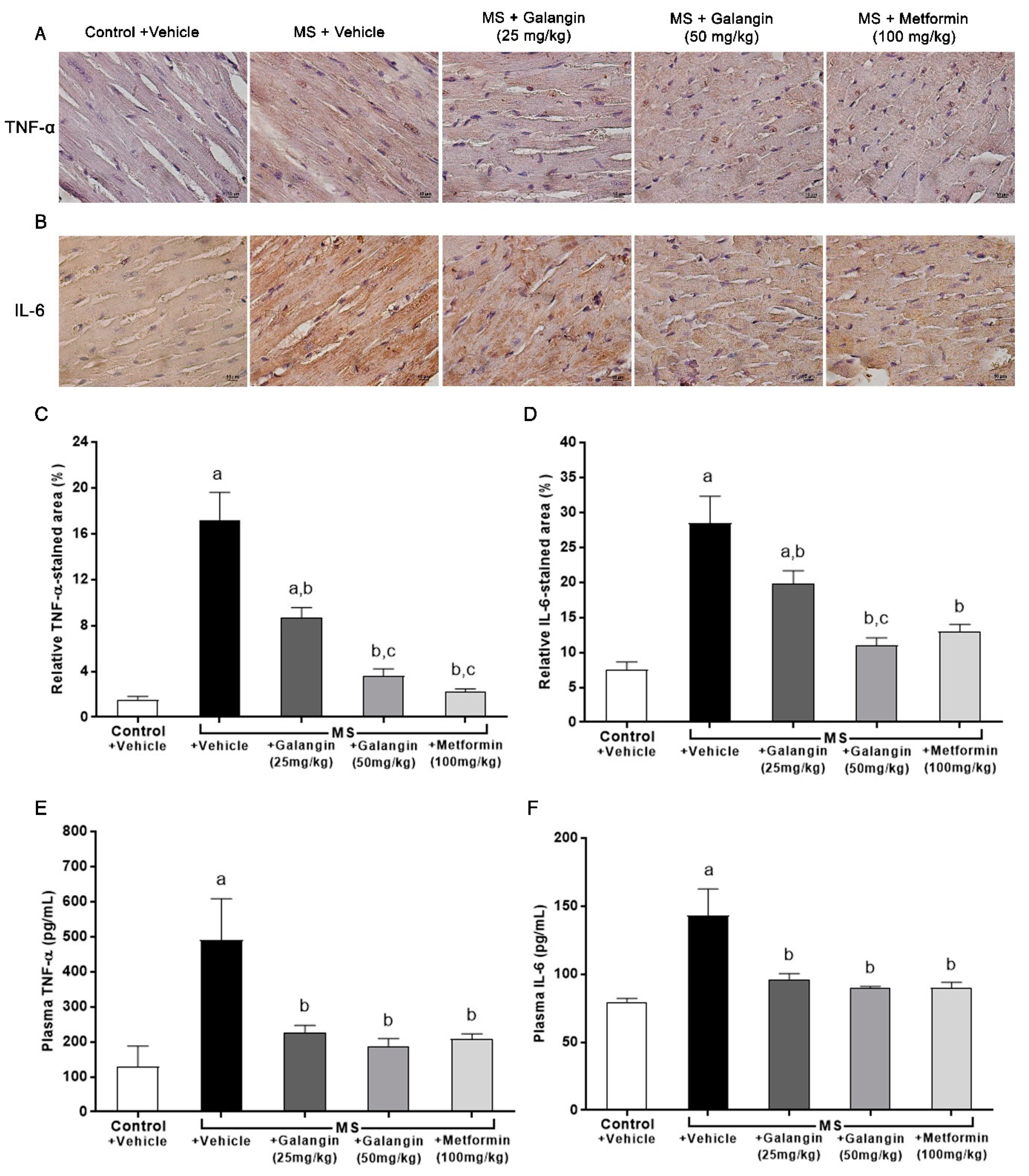
3.7. Effects of Galangin on Myocardial Inflammation
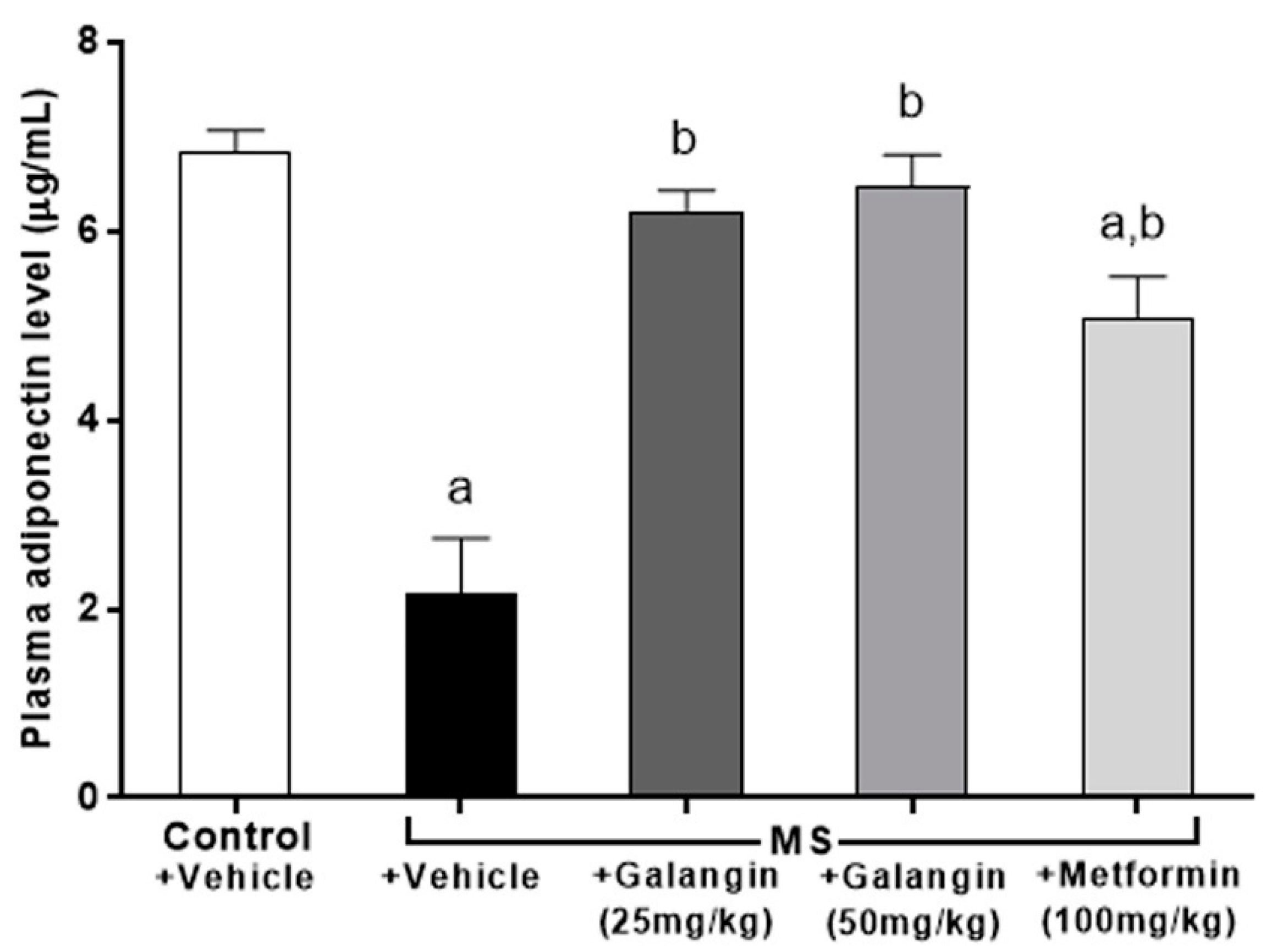
3.8. Effects of Galangin on Plasma Adiponectin Levels
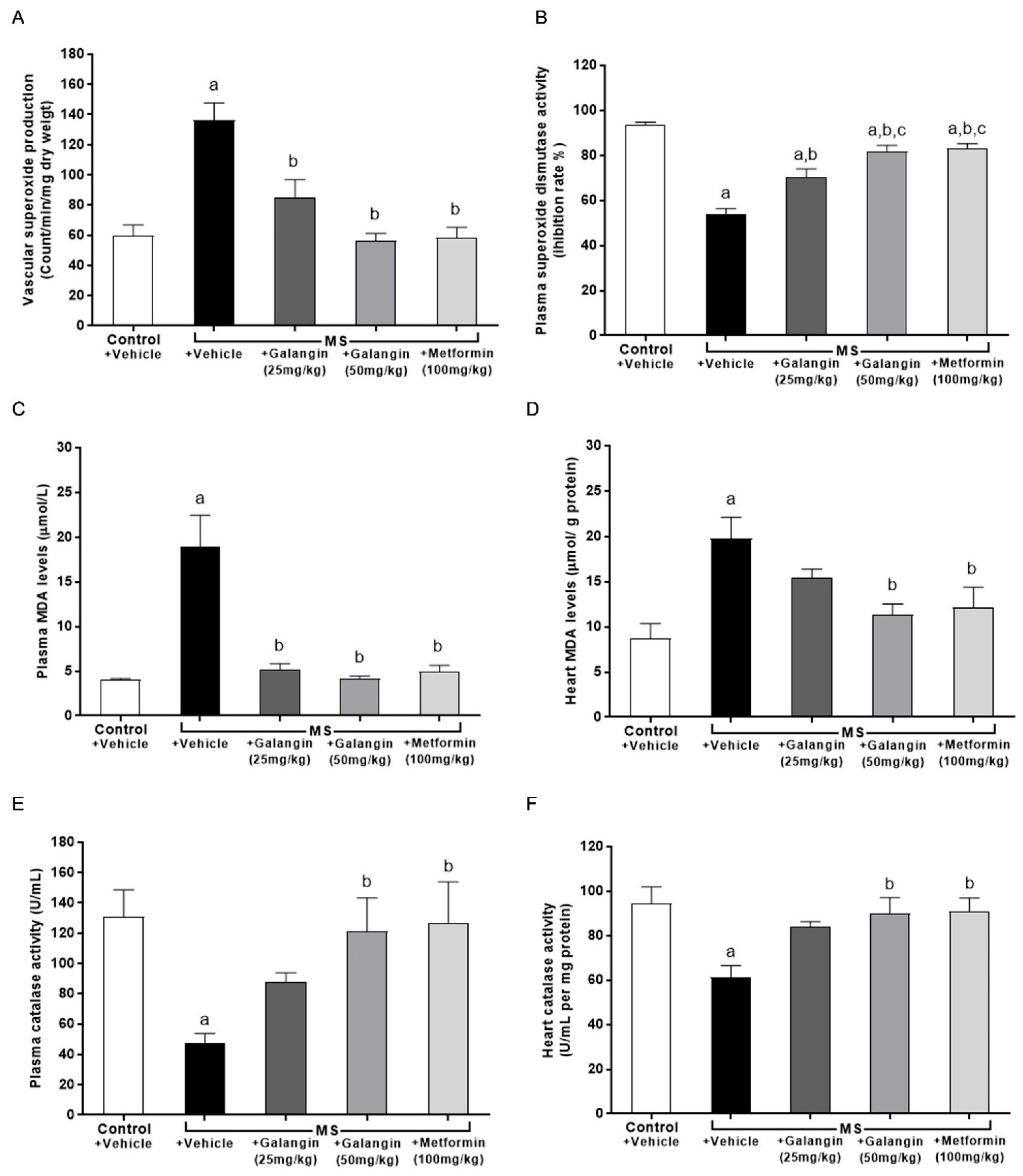
3.9. Effects of Galangin on Oxidative Stress Markers and Endogenous Antioxidant Enzymes
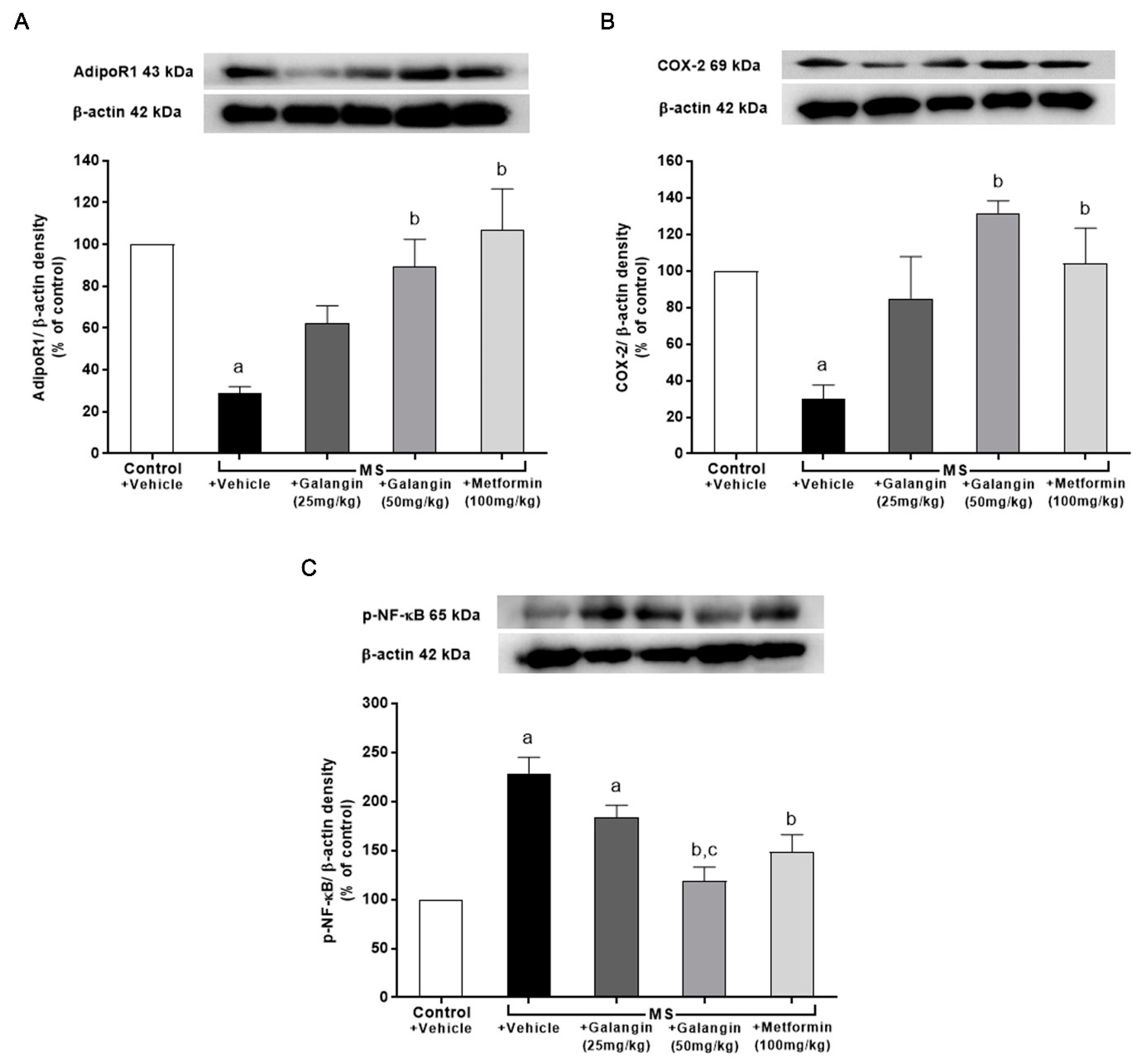
3.10. Effects of Galangin on AdipoR1, COX-2, and p-NF-κB Expression in Cardiac Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014, 943162. [Google Scholar] [CrossRef]
- Lee, S.; Lee, H.J. Association between nutrients and metabolic syndrome in middle-aged Korean women. Arch. Endocrinol. Metab. 2020, 64, 298–305. [Google Scholar] [CrossRef]
- Calle, E.E.; Kaaks, R. Overweight, obesity and cancer: Epidemiological evidence and proposed mechanisms. Nat. Rev. Cancer 2004, 4, 579–591. [Google Scholar] [CrossRef]
- Abo El-Magd, N.F.; El-Mesery, M. Amelioration effect of black seed oil against high-fat diet-induced obesity in rats through Nrf2/HO-1 pathway. J. Food Biochem. 2021, 45, 13693. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, Y.; Suo, M.; Jin, X.; Tian, G. Telmisartan prevents high-fat diet-induced hypertension and decreases perirenal fat in rats. J. Biomed. Res. 2012, 26, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Guo, Y.; Sun, L.; Lai, X.; Li, Q.; Zhang, W.; Xiang, L.; Sun, S.; Cao, F. Six types of tea reduce high-fat-diet-induced fat accumulation in mice by increasing lipid metabolism and suppressing inflammation. Food Funct. 2019, 10, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, A.M.; Al Suleimani, Y.M.; Ashique, M.; Manoj, P.; Ali, B.H. Effect of infliximab and tocilizumab on fructose-induced hyperinsulinemia and hypertension in rats. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 105, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Dupas, J.; Feray, A.; Goanvec, C.; Guernec, A.; Samson, N.; Bougaran, P.; Guerrero, F.; Mansourati, J. Metabolic Syndrome and Hypertension Resulting from Fructose Enriched Diet in Wistar Rats. BioMed Res. Int. 2017, 2017, 2494067. [Google Scholar] [CrossRef]
- Ouwens, D.M.; Diamant, M.; Fodor, M.; Habets, D.D.J.; Pelsers, M.; El Hasnaoui, M.; Dang, Z.C.; van den Brom, C.E.; Vlasblom, R.; Rietdijk, A.; et al. Cardiac contractile dysfunction in insulin-resistant rats fed a high-fat diet is associated with elevated CD36-mediated fatty acid uptake and esterification. Diabetologia 2007, 50, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Prasatthong, P.; Meephat, S.; Rattanakanokchai, S.; Bunbupha, S.; Prachaney, P.; Maneesai, P.; Pakdeechote, P. Hesperidin ameliorates signs of the metabolic syndrome and cardiac dysfunction via IRS/Akt/GLUT4 signaling pathway in a rat model of diet-induced metabolic syndrome. Eur. J. Nutr. 2021, 60, 833–848. [Google Scholar] [CrossRef]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burhans, M.S.; Hagman, D.K.; Kuzma, J.N.; Schmidt, K.A.; Kratz, M. Contribution of Adipose Tissue Inflammation to the Development of Type 2 Diabetes Mellitus. Compr. Physiol. 2018, 9, 1–58. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Zavos, C.; Tsiaousi, E. The role of adiponectin in the pathogenesis and treatment of non-alcoholic fatty liver disease. Diabetes Obes. Metab. 2010, 12, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Shimada, K.; Fukuda, D.; Shimada, Y.; Ehara, S.; Hirose, M.; Kataoka, T.; Kamimori, K.; Shimodozono, S.; Kobayashi, Y.; et al. Implications of plasma concentrations of adiponectin in patients with coronary artery disease. Heart 2004, 90, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.J.; Park, C.G.; Seo, H.S.; Oh, D.J.; Ro, Y.M. Associations among plasma adiponectin, hypertension, left ventricular diastolic function and left ventricular mass index. Blood Press. 2004, 13, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Pistilli, D.; Gucciardo, M.; Leonetti, F.; Miraldi, F.; Brancaccio, G.; Gallo, P.; Di Gioia, C.R. Adiponectin expression in human epicardial adipose tissue in vivo is lower in patients with coronary artery disease. Cytokine 2005, 29, 251–255. [Google Scholar] [CrossRef]
- Essick, E.E.; Ouchi, N.; Wilson, R.M.; Ohashi, K.; Ghobrial, J.; Shibata, R.; Pimentel, D.R.; Sam, F. Adiponectin mediates cardioprotection in oxidative stress-induced cardiac myocyte remodeling. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, 984–993. [Google Scholar] [CrossRef] [Green Version]
- Iwabu, M.; Yamauchi, T.; Okada-Iwabu, M.; Sato, K.; Nakagawa, T.; Funata, M.; Yamaguchi, M.; Namiki, S.; Nakayama, R.; Tabata, M.; et al. Adiponectin and AdipoR1 regulate PGC-1alpha and mitochondria by Ca(2+) and AMPK/SIRT1. Nature 2010, 464, 1313–1319. [Google Scholar] [CrossRef]
- Shibata, R.; Sato, K.; Pimentel, D.R.; Takemura, Y.; Kihara, S.; Ohashi, K.; Funahashi, T.; Ouchi, N.; Walsh, K. Adiponectin protects against myocardial ischemia-reperfusion injury through AMPK- and COX-2-dependent mechanisms. Nat. Med. 2005, 11, 1096–1103. [Google Scholar] [CrossRef]
- Bolli, R.; Shinmura, K.; Tang, X.L.; Kodani, E.; Xuan, Y.T.; Guo, Y.; Dawn, B. Discovery of a new function of cyclooxygenase (COX)-2: COX-2 is a cardioprotective protein that alleviates ischemia/reperfusion injury and mediates the late phase of preconditioning. Cardiovasc. Res. 2002, 55, 506–519. [Google Scholar] [CrossRef]
- Gilroy, D.W.; Colville-Nash, P.R.; Willis, D.; Chivers, J.; Paul-Clark, M.J.; Willoughby, D.A. Inducible cyclooxygenase may have anti-inflammatory properties. Nat. Med. 1999, 5, 698–701. [Google Scholar] [CrossRef]
- Villarreal-Molina, M.T.; Antuna-Puente, B. Adiponectin: Anti-inflammatory and cardioprotective effects. Biochimie 2012, 94, 2143–2149. [Google Scholar] [CrossRef] [PubMed]
- Kawano, S.; Kubota, T.; Monden, Y.; Tsutsumi, T.; Inoue, T.; Kawamura, N.; Tsutsui, H.; Sunagawa, K. Blockade of NF-kappaB improves cardiac function and survival after myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Nagasawa, K.; Matsuura, N.; Takeshita, Y.; Ito, S.; Sano, Y.; Yamada, Y.; Uchinaka, A.; Murohara, T.; Nagata, K. Attenuation of cold stress-induced exacerbation of cardiac and adipose tissue pathology and metabolic disorders in a rat model of metabolic syndrome by the glucocorticoid receptor antagonist RU486. Nutr. Diabetes 2016, 6, 207. [Google Scholar] [CrossRef] [Green Version]
- Balderas-Villalobos, J.; Molina-Muñoz, T.; Mailloux-Salinas, P.; Bravo, G.; Carvajal, K.; Gómez-Viquez, N.L. Oxidative stress in cardiomyocytes contributes to decreased SERCA2a activity in rats with metabolic syndrome. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, 1344–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meephat, S.; Prasatthong, P.; Rattanakanokchai, S.; Bunbupha, S.; Maneesai, P.; Pakdeechote, P. Diosmetin attenuates metabolic syndrome and left ventricular alterations via the suppression of angiotensin II/AT1 receptor/gp(91phox)/p-NF-κB protein expression in high-fat diet fed rats. Food Funct. 2021, 12, 1469–1481. [Google Scholar] [CrossRef]
- Bilginoglu, A. Cardiovascular protective effect of pioglitazone on oxidative stress in rats with metabolic syndrome. J. Chin. Med. Assoc. 2019, 82, 452–456. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Torres-Narváez, J.C.; Guarner-Lans, V.; Díaz-Díaz, E.; Perezpeña-Diazconti, M.; Palacios, A.R.; Manzano-Pech, L. Myocardial Protection from Ischemia-Reperfusion Damage by the Antioxidant Effect of Hibiscus sabdariffa Linnaeus on Metabolic Syndrome Rats. Oxidative Med. Cell. Longev. 2019, 2019, 1724194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grycel, S.; Markowski, A.R.; Hady, H.R.; Zabielski, P.; Kojta, I.; Imierska, M.; Górski, J.; Blachnio-Zabielska, A.U. Metformin treatment affects adipocytokine secretion and lipid composition in adipose tissues of diet-induced insulin-resistant rats. Nutrition 2019, 63–64, 126–133. [Google Scholar] [CrossRef]
- Rai, R.C.; Bagul, P.K.; Banerjee, S.K. NLRP3 inflammasome drives inflammation in high fructose fed diabetic rat liver: Effect of resveratrol and metformin. Life Sci. 2020, 253, 117727. [Google Scholar] [CrossRef]
- Mohan, M.; Al-Talabany, S.; McKinnie, A.; Mordi, I.R.; Singh, J.S.S.; Gandy, S.J.; Baig, F.; Hussain, M.S.; Bhalraam, U.; Khan, F.; et al. A randomized controlled trial of metformin on left ventricular hypertrophy in patients with coronary artery disease without diabetes: The MET-REMODEL trial. Eur. Heart J. 2019, 40, 3409–3417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cushnie, T.P.; Hamilton, V.E.; Chapman, D.G.; Taylor, P.W.; Lamb, A.J. Aggregation of Staphylococcus aureus following treatment with the antibacterial flavonol galangin. J. Appl. Microbiol. 2007, 103, 1562–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, M.Y.; Sohn, S.J.; Au, W.W. Anti-genotoxicity of galangin as a cancer chemopreventive agent candidate. Mutat. Res. 2001, 488, 135–150. [Google Scholar] [CrossRef]
- Huang, Y.C.; Tsai, M.S.; Hsieh, P.C.; Shih, J.H.; Wang, T.S.; Wang, Y.C.; Lin, T.H.; Wang, S.H. Galangin ameliorates cisplatin-induced nephrotoxicity by attenuating oxidative stress, inflammation and cell death in mice through inhibition of ERK and NF-kappaB signaling. Toxicol. Appl. Pharmacol. 2017, 329, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gong, G.; Yang, W.; Li, Y.; Jiang, M.; Li, L. Antifibrotic activity of galangin, a novel function evaluated in animal liver fibrosis model. Environ. Toxicol. Pharmacol. 2013, 36, 288–295. [Google Scholar] [CrossRef]
- Jung, Y.C.; Kim, M.E.; Yoon, J.H.; Park, P.R.; Youn, H.-Y.; Lee, H.-W.; Lee, J.S. Anti-inflammatory effects of galangin on lipopolysaccharide-activated macrophages via ERK and NF-κB pathway regulation. Immunopharmacol. Immunotoxicol. 2014, 36, 426–432. [Google Scholar] [CrossRef]
- Aloud, A.A.; Veeramani, C.; Govindasamy, C.; Alsaif, M.A.; El Newehy, A.S.; Al-Numair, K.S. Galangin, a dietary flavonoid, improves antioxidant status and reduces hyperglycemia-mediated oxidative stress in streptozotocin-induced diabetic rats. Redox Rep. Commun. Free Radic. Res. 2017, 22, 290–300. [Google Scholar] [CrossRef]
- Aloud, A.A.; Chinnadurai, V.; Govindasamy, C.; Alsaif, M.A.; Al-Numair, K.S. Galangin, a dietary flavonoid, ameliorates hyperglycaemia and lipid abnormalities in rats with streptozotocin-induced hyperglycaemia. Pharm. Biol. 2018, 56, 302–308. [Google Scholar] [CrossRef]
- Aladaileh, S.H.; Abukhalil, M.H.; Saghir, S.A.M.; Hanieh, H.; Alfwuaires, M.A.; Almaiman, A.A.; Bin-Jumah, M.; Mahmoud, A.M. Galangin Activates Nrf2 Signaling and Attenuates Oxidative Damage, Inflammation, and Apoptosis in a Rat Model of Cyclophosphamide-Induced Hepatotoxicity. Biomolecules 2019, 9, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.J.; Lin, J.T.; Wang, H.P.; Huang, W.C. A simple, sensitive, non-stimulated photon counting system for detection of superoxide anion in whole blood. Experientia 1996, 52, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Lasker, S.; Rahman, M.M.; Parvez, F.; Zamila, M.; Miah, P.; Nahar, K.; Kabir, F.; Sharmin, S.B.; Subhan, N.; Ahsan, G.U.; et al. High-fat diet-induced metabolic syndrome and oxidative stress in obese rats are ameliorated by yogurt supplementation. Sci. Rep. 2019, 9, 20026. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L.; Schwarz, J.M.; Keim, N.L.; Griffen, S.C.; Bremer, A.A.; Graham, J.L.; Hatcher, B.; Cox, C.L.; Dyachenko, A.; Zhang, W.; et al. Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral adiposity and lipids and decreases insulin sensitivity in overweight/obese humans. J. Clin. Investig. 2009, 119, 1322–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Tian, Y.; Chen, Y.; Shen, X.; Pan, L.; Li, G. Hyperinsulinemia rather than insulin resistance itself induces blood pressure elevation in high fat diet-fed rats. Clin. Exp. Hypertens. 2020, 42, 614–621. [Google Scholar] [CrossRef]
- Kluge, A.; Rangrez, A.Y.; Kilian, L.S.; Pott, J.; Bernt, A.; Frauen, R.; Rohrbeck, A.; Frey, N.; Frank, D. Rho-family GTPase 1 (Rnd1) is a biomechanical stress-sensitive activator of cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol. 2019, 129, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, P.; López de Pablo, A.L.; García-Prieto, C.F.; Somoza, B.; Quintana-Villamandos, B.; Gómez de Diego, J.J.; Gutierrez-Arzapalo, P.Y.; Ramiro-Cortijo, D.; González, M.C.; Arribas, S.M. Long term effects of fetal undernutrition on rat heart. Role of hypertension and oxidative stress. PLoS ONE 2017, 12, e0171544. [Google Scholar] [CrossRef]
- Suthahar, N.; Meijers, W.C.; Silljé, H.H.W.; De Boer, R.A. From Inflammation to Fibrosis-Molecular and Cellular Mechanisms of Myocardial Tissue Remodelling and Perspectives on Differential Treatment Opportunities. Curr. Heart Fail. Rep. 2017, 14, 235–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, E.D.; Litwin, S.E.; Sweeney, G. Cardiac remodeling in obesity. Physiol. Rev. 2008, 88, 389–419. [Google Scholar] [CrossRef] [PubMed]
- Elrashidy, R.A. Dysregulation of nuclear factor erythroid 2-related factor 2 signaling and activation of fibrogenic pathways in hearts of high fat diet-fed rats. Mol. Biol. Rep. 2020, 47, 2821–2834. [Google Scholar] [CrossRef]
- Kondo, K.; Shibata, R.; Unno, K.; Shimano, M.; Ishii, M.; Kito, T.; Shintani, S.; Walsh, K.; Ouchi, N.; Murohara, T. Impact of a single intracoronary administration of adiponectin on myocardial ischemia/reperfusion injury in a pig model. Circ. Cardiovasc. Interv. 2010, 3, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Berendoncks, A.M.V.; Stensvold, D.; Garnier, A.; Fortin, D.; Sente, T.; Vrints, C.J.; Arild, S.S.; Ventura-Clapier, R.; Wisløff, U.; Conraads, V.M. Disturbed adiponectin–AMPK system in skeletal muscle of patients with metabolic syndrome. Eur. J. Prev. Cardiol. 2015, 22, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Schindler, M.; Pendzialek, M.; Grybel, K.J.; Seeling, T.; Gürke, J.; Fischer, B.; Navarrete Santos, A. Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts. Hum. Reprod. 2017, 32, 1382–1392. [Google Scholar] [CrossRef] [Green Version]
- Cook, W.S.; Yeldandi, A.V.; Rao, M.S.; Hashimoto, T.; Reddy, J.K. Less extrahepatic induction of fatty acid beta-oxidation enzymes by PPAR alpha. Biochem. Biophys. Res. Commun. 2000, 278, 250–257. [Google Scholar] [CrossRef]
- Chan, P.C.; Liao, M.T.; Hsieh, P.S. The Dualistic Effect of COX-2-Mediated Signaling in Obesity and Insulin Resistance. Int. J. Mol. Sci. 2019, 20, 3115. [Google Scholar] [CrossRef] [Green Version]
- Vegiopoulos, A.; Müller-Decker, K.; Strzoda, D.; Schmitt, I.; Chichelnitskiy, E.; Ostertag, A.; Berriel Diaz, M.; Rozman, J.; Hrabe de Angelis, M.; Nüsing, R.M.; et al. Cyclooxygenase-2 controls energy homeostasis in mice by de novo recruitment of brown adipocytes. Science 2010, 328, 1158–1161. [Google Scholar] [CrossRef]
- Francés, D.E.; Motiño, O.; Agrá, N.; González-Rodríguez, Á.; Fernández-Álvarez, A.; Cucarella, C.; Mayoral, R.; Castro-Sánchez, L.; García-Casarrubios, E.; Boscá, L.; et al. Hepatic cyclooxygenase-2 expression protects against diet-induced steatosis, obesity, and insulin resistance. Diabetes 2015, 64, 1522–1531. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Alagawadi, K.R. Anti-obesity effects of galangin, a pancreatic lipase inhibitor in cafeteria diet fed female rats. Pharm. Biol. 2013, 51, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, 001651. [Google Scholar] [CrossRef] [Green Version]
- Hausenloy, D.J.; Yellon, D.M. Survival kinases in ischemic preconditioning and postconditioning. Cardiovasc. Res. 2006, 70, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Bolli, R.; Rokosh, G.D.; Bi, Q.; Dai, S.; Shirk, G.; Tang, X.-L. The cardioprotection of the late phase of ischemic preconditioning is enhanced by postconditioning via a COX-2-mediated mechanism in conscious rats. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, 2557–2564. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Po, S.S.; Zhang, B.; Bai, F.; Li, J.; Qin, F.; Liu, N.; Sun, C.; Xiao, Y.; Tu, T.; et al. Metformin regulates adiponectin signalling in epicardial adipose tissue and reduces atrial fibrillation vulnerability. J. Cell. Mol. Med. 2020, 24, 7751–7766. [Google Scholar] [CrossRef]
- Bhamra, G.S.; Hausenloy, D.J.; Davidson, S.M.; Carr, R.D.; Paiva, M.; Wynne, A.M.; Mocanu, M.M.; Yellon, D.M. Metformin protects the ischemic heart by the Akt-mediated inhibition of mitochondrial permeability transition pore opening. Basic Res. Cardiol. 2008, 103, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Adamia, N.; Virsaladze, D.; Charkviani, N.; Skhirtladze, M.; Khutsishvili, M. Effect of metformin therapy on plasma adiponectin and leptin levels in obese and insulin resistant postmenopausal females with type 2 diabetes. Georgian Med. News 2007, 145, 52–55. [Google Scholar]
- Su, J.R.; Lu, Z.H.; Su, Y.; Zhao, N.; Dong, C.L.; Sun, L.; Zhao, S.F.; Li, Y. Relationship of Serum Adiponectin Levels and Metformin Therapy in Patients with Type 2 Diabetes. Horm. Metab. Res. 2016, 48, 92–98. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control + Vehicle | MS + Vehicle | MS + Galangin (25 mg/kg) | MS + Galangin (50 mg/kg) | MS + Metformin (100 mg/kg) | |
|---|---|---|---|---|---|
| BW (g) | 714.17 ± 22.69 | 837.14 ± 35.59 a | 793.29 ± 17.75 | 789.50 ± 16.37 | 739.13 ± 28.12 |
| HW (g) | 1.47 ± 0.05 | 1.76 ± 0.06 a | 1.72 ± 0.04 a | 1.59 ± 0.04 | 1.54 ± 0.06 b |
| HW/BW (mg/g) | 2.06 ± 0.05 | 2.11 ± 0.07 | 2.17 ± 0.02 | 2.01 ± 0.06 | 2.09 ± 0.05 |
| VW (g) | 1.30 ± 0.04 | 1.52 ± 0.05 a | 1.44 ± 0.02 | 1.41 ± 0.03 | 1.32 ± 0.05 b |
| VW/BW (mg/g) | 1.82 ± 0.03 | 1.84 ± 0.04 | 1.82 ± 0.02 | 1.79 ± 0.03 | 1.80 ± 0.04 |
| RP pads weight (g) | 18.96 ± 2.66 | 59.91 ± 5.23 a | 55.62 ± 3.13 a | 52.25 ± 1.24 a | 40.53 ± 6.13 a,b |
| RP pads weight/BW (mg/g) | 26.43 ± 3.35 | 70.86 ± 3.57 a | 69.85 ± 2.84 a | 67.00 ± 1.00 a | 53.54 ± 6.63 a,b |
| EP pads weight (g) | 14.81 ± 0.72 | 29.09 ± 2.85 a | 26.13 ± 0.76 a | 25.52 ± 1.94 a | 23.85 ± 2.68 a |
| EP pads weight/BW (mg/g) | 20.76 ± 0.88 | 34.39 ± 2.09 a | 33.15 ± 1.15 a | 32.62 ± 1.89 a | 31.81 ± 3.09 a |
| LW(g) | 19.01 ± 1.17 | 24.34 ± 1.20 a | 21.74 ± 0.92 | 19.72 ± 0.70 b | 19.79 ± 0.75 b |
| LW/BW (mg/g) | 26.81 ± 1.92 | 29.00 ± 1.03 | 27.41 ± 0.95 | 25.34 ± 0.51 | 26.85 ± 0.86 |
| Control + Vehicle | MS + Vehicle | MS + Galangin (25 mg/kg) | MS + Galangin (50 mg/kg) | MS + Metformin (100 mg/kg) | |
|---|---|---|---|---|---|
| Fasting blood glucose (mg/dL) | 89.20 ± 3.18 | 120.00 ± 5.42 a | 105.40 ± 1.33 | 100.40 ± 4.17 b | 93.80 ± 4.65 b |
| Fasting serum insulin (ng/mL) | 1.99 ± 0.26 | 4.02 ± 0.71 a | 2.76 ± 0.25 | 2.06 ± 0.52 b | 2.13 ± 0.25 b |
| HOMA-IR index | 10.87 ± 1.38 | 30.26 ± 6.95 a | 17.73 ± 1.38 | 12.49 ± 3.12 b | 12.33 ± 1.89 b |
| Plasma total cholesterol (mmol/L) | 0.92 ± 0.03 | 2.57 ± 0.30 a | 1.35 ± 0.08 b | 1.00 ± 0.10 b | 1.17 ± 0.07 b |
| Plasma triglyceride (mmol/L) | 0.41 ± 0.07 | 2.49 ± 0.43 a | 0.71 ± 0.19 b | 0.54 ± 0.09 b | 0.45 ± 0.06 b |
| Plasma HDL-C (mmol/L) | 1.71 ± 0.13 | 0.33 ± 0.03 a | 0.79 ± 0.09 a,b | 1.30 ± 0.15 b,c | 0.99 ± 0.11 a,b |
| Liver total cholesterol content (mg/g tissue) | 11.17 ± 1.87 | 23.87 ± 2.71 a | 17.64 ± 1.46 | 11.53 ± 1.57 b | 11.58 ± 2.96 b |
| Liver triglyceride content (mg/g tissue) | 13.79 ± 1.10 | 33.13 ± 1.58 a | 19.42 ± 1.32 b | 16.29 ± 2.26 b | 15.42 ± 1.71 b |
| AST (U/L) | 49.00 ± 4.63 | 83.55 ± 4.20 a | 76.17 ± 4.56 a | 52.20 ± 1.43 b,c | 65.50 ± 3.59 a,b |
| ALT (U/L) | 19.13 ± 2.88 | 39.60 ± 2.58 a | 29.17 ± 3.20 b | 22.86 ± 0.86 b | 24.75 ± 1.66 b |
| Control + Vehicle | MS + Vehicle | MS + Galangin (25 mg/kg) | MS + Galangin (50 mg/kg) | MS + Metformin (100 mg/kg) | |
|---|---|---|---|---|---|
| Systolic blood pressure (mmHg) | 118.49 ± 2.50 | 156.78 ± 1.96 a | 146.71 ± 2.24 a | 131.41 ± 2.75 a,b,c | 120.86 ± 3.04 b,c,d |
| Diastolic blood pressure (mmHg) | 80.28 ± 3.06 | 107.32 ± 1.11 a | 97.46 ± 3.12 a | 85.72 ± 2.00 b,c | 75.24 ± 3.43 b,c |
| Mean arterial pressure (mmHg) | 93.58 ± 2.82 | 123.90 ± 1.19 a | 113.73 ± 2.55 a | 100.95 ± 2.05 a,b,c | 90.48 ± 2.78 b,c |
| Pulse pressure | 39.89 ± 1.67 | 49.73 ± 2.11 | 48.81 ± 2.99 | 45.68 ± 2.10 | 45.72 ± 3.81 |
| Heart rate (bmp) | 331.86 ± 6.06 | 378.57 ± 9.84 a | 336.15 ± 11.29 b | 336.09 ± 9.99 b | 314.95 ± 4.68 b |
| Control + Vehicle | MS + Vehicle | MS + Galangin (25 mg/kg) | MS + Galangin (50 mg/kg) | MS + Metformin (100 mg/kg) | |
|---|---|---|---|---|---|
| IVSd (cm) | 0.178 ± 0.005 | 0.200 ± 0.008 | 0.177 ± 0.008 | 0.174 ± 0.012 | 0.180 ± 0.008 |
| IVSs (cm) | 0.273 ± 0.010 | 0.272 ± 0.015 | 0.271 ± 0.10 | 0.283 ± 0.010 | 0.281 ± 0.012 |
| LVIDd (cm) | 0.759 ± 0.018 | 0.626 ± 0.030 a | 0.793 ± 0.018 b | 0.776 ± 0.026 b | 0.747 ± 0.008 b |
| LVIDs (cm) | 0.463 ± 0.023 | 0.434 ± 0.024 | 0.501 ± 0.020 | 0.463 ± 0.014 | 0.459 ± 0.008 |
| LVPWd (cm) | 0.197 ± 0.010 | 0.244 ± 0.013 a | 0.191 ± 0.011 b | 0.190 ± 0.010 b | 0.198 ± 0.006 b |
| LVPWs (cm) | 0.278 ± 0.009 | 0.307 ± 0.014 | 0.279 ± 0.008 | 0.277 ± 0.009 | 0.285 ± 0.009 |
| EDV (mL) | 0.969 ± 0.061 | 0.586 ± 0.077 a | 1.096 ± 0.070 b | 1.036 ± 0.092 b | 0.930 ± 0.029 b |
| ESV (mL) | 0.252 ± 0.031 | 0.212 ± 0.030 | 0.314 ± 0.037 | 0.249 ± 0.020 | 0.238 ± 0.013 |
| EF (%) | 74.480 ± 1.880 | 63.591 ± 3.185 a | 71.683 ± 1.417 | 75.839 ± 0.928 b | 74.093 ± 1.416 b |
| SV (mL) | 0.717 ± 0.033 | 0.374 ± 0.058 a | 0.780 ± 0.034 b | 0.790 ± 0.074 b | 0.689 ± 0.029 b |
| FS (%) | 38.840 ± 1.680 | 30.518 ± 2.137 a | 36.854 ± 0.972 b | 39.859 ± 0.851 b | 38.369 ± 1.204 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasatthong, P.; Meephat, S.; Rattanakanokchai, S.; Khamseekaew, J.; Bunbupha, S.; Prachaney, P.; Maneesai, P.; Pakdeechote, P. Galangin Resolves Cardiometabolic Disorders through Modulation of AdipoR1, COX-2, and NF-κB Expression in Rats Fed a High-Fat Diet. Antioxidants 2021, 10, 769. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050769
Prasatthong P, Meephat S, Rattanakanokchai S, Khamseekaew J, Bunbupha S, Prachaney P, Maneesai P, Pakdeechote P. Galangin Resolves Cardiometabolic Disorders through Modulation of AdipoR1, COX-2, and NF-κB Expression in Rats Fed a High-Fat Diet. Antioxidants. 2021; 10(5):769. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050769
Chicago/Turabian StylePrasatthong, Patoomporn, Sariya Meephat, Siwayu Rattanakanokchai, Juthamas Khamseekaew, Sarawoot Bunbupha, Parichat Prachaney, Putcharawipa Maneesai, and Poungrat Pakdeechote. 2021. "Galangin Resolves Cardiometabolic Disorders through Modulation of AdipoR1, COX-2, and NF-κB Expression in Rats Fed a High-Fat Diet" Antioxidants 10, no. 5: 769. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050769