
Thiol Reductases in Deinococcus Bacteria and Roles in Stress Tolerance


1
Aix Marseille Univ, Molecular and Environmental Microbiology (MEM) Team, CEA, CNRS, BIAM, F-13108 Saint Paul-Lez-Durance, France
2
University of Lorraine, Inrae, IAM, F-54000 Nancy, France
3
Aix Marseille Univ, Plant Protective Proteins (PPV) Team, CEA, CNRS, BIAM, F-13108 Saint Paul-Lez-Durance, France
*
Authors to whom correspondence should be addressed.
Antioxidants 2022, 11(3), 561; https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030561
Submission received: 15 February 2022
/
Revised: 11 March 2022
/
Accepted: 13 March 2022
/
Published: 16 March 2022
(This article belongs to the Special Issue Thioredoxin and Glutaredoxin Systems II)
Abstract
:Deinococcus species possess remarkable tolerance to extreme environmental conditions that generate oxidative damage to macromolecules. Among enzymes fulfilling key functions in metabolism regulation and stress responses, thiol reductases (TRs) harbour catalytic cysteines modulating the redox status of Cys and Met in partner proteins. We present here a detailed description of Deinococcus TRs regarding gene occurrence, sequence features, and physiological functions that remain poorly characterised in this genus. Two NADPH-dependent thiol-based systems are present in Deinococcus. One involves thioredoxins, disulfide reductases providing electrons to protein partners involved notably in peroxide scavenging or in preserving protein redox status. The other is based on bacillithiol, a low-molecular-weight redox molecule, and bacilliredoxin, which together protect Cys residues against overoxidation. Deinococcus species possess various types of thiol peroxidases whose electron supply depends either on NADPH via thioredoxins or on NADH via lipoylated proteins. Recent data gained on deletion mutants confirmed the importance of TRs in Deinococcus tolerance to oxidative treatments, but additional investigations are needed to delineate the redox network in which they operate, and their precise physiological roles. The large palette of Deinococcus TR representatives very likely constitutes an asset for the maintenance of redox homeostasis in harsh stress conditions.
Keywords:
Deinococcus; thiol; reductase; peroxidase; cysteine; oxidative stress; thioredoxin; bacillithiol; protein redox status1. Introduction
Bacteria belonging to the genus Deinococcus are extremely tolerant to radiation, desiccation, and other conditions that generate oxidative damage to biomolecules, including DNA and proteins. The first described member of this genus, Deinococcus radiodurans, isolated as a contaminant from canned meat that was supposedly sterilised by a high dose of ionising radiation, has been studied most extensively [1,2]. In the last decades, more than 90 other Deinococcus species have been isolated worldwide from different places and environments (e.g., air, water, desert soils). Many genes and several mechanisms contribute to this tolerance [3]. Deinococcus bacteria are able to repair massive DNA damage generated by radiation or desiccation [4,5], indicating efficient limitation or repair of damage to other biomolecules, such as DNA repair proteins and other enzymes, to preserve their function under these conditions. Indeed, proteins in Deinococcus were found to be much better protected against oxidative damage than those in radiation-sensitive species, such as Escherichia coli [6,7,8,9]. Both nonenzymatic (e.g., carotenoids, a high intracellular Mn2+/Fe2+ ratio, and antioxidant complexes containing Mn2+, phosphate, and peptides) and enzymatic (e.g., superoxide dismutases, catalases, peroxidases, thioredoxins) antioxidant systems are present in Deinococcus [3].
Thiol-dependent antioxidant enzymes, namely, thiol reductases (TRs), are ubiquitous enzymes harbouring catalytic cysteines that modulate the redox status of Cys and Met in partner proteins. Living organisms possess various thiol-dependent antioxidant systems displaying specific features and electron sources [10]. Among them, the thioredoxin (Trx) and glutathione (GSH) systems are the most represented and well characterised [11,12,13]. The Trx pathway is composed of NADPH, NADPH-dependent Trx reductase (TrxR), and Trx, a disulfide reductase supplying electrons to many types of partners and targets, including peroxiredoxins (Prxs) and methionine sulfoxide reductases (Msrs), which are involved in the scavenging of peroxides and preservation of protein redox status, respectively. In the GSH system, electrons are provided by NADPH and transferred via NADPH-dependent GSH reductase to GSH, a low-molecular-weight (LMW) compound having direct antioxidant activity. GSH is also able to transfer reducing power to glutaredoxins (Grxs), proteins related to Trxs, and fulfilling a similar function in the control of Cys redox status in partner proteins. Trx and GSH systems play critical roles in the maintenance of protein redox homeostasis, particularly in responses to environmental constraints, upon diseases or during aging [10,14,15,16]. Indeed, these reducing systems participate in the prevention and repair of oxidative damage and in the control of redox post-translational modifications occurring in their partner proteins. This type of modification is associated with changes in enzyme activity, protein conformation or structure, and consequently with redox-based signal transduction pathways. While the Trx system is ubiquitous to all domains of life, the GSH system is absent in many bacteria, including Bacillus species and pathogenic bacteria, such as Mycobacterium tuberculosis or Staphylococcus aureus, in which thiol-based responses are crucial against host oxidative defences [17]. In these species, systems based on other LMW thiol compounds, such as mycothiol (MSH) or bacillithiol (BSH), have been identified and fulfil roles similar to those of GSH [18,19,20].
The extreme environmental conditions that Deinococcus is able to tolerate strongly affect among others cell redox homeostasis and thiol status. Thus, TRs are very likely part of the mechanisms underlying this outstanding feature [3]. Consistently, it was reported that a Deinococcus geothermalis mutant lacking the cystine importer system, which is a source of thiols, was more sensitive to an H2O2 treatment [21]. Some studies based on biochemical, pharmacological, or genetic approaches characterised the Trx system in Deinococcus [22,23,24] and indicated that it likely plays an essential role in responses to oxidative stress [25,26]. In a previous review describing Deinococcus antioxidant systems [3], the main TR families were listed, revealing that this bacterial genus possesses a complete Trx system, including a relatively high number of TRs related to Trxs and Prxs. Of note, Deinococcus does not possess GSH, but BSH and the relevant reducing system, including NADPH–BSH reductase (Bdr) and bacilliredoxin (Brx) [27].
In this review, we aim to carry out a more detailed and exhaustive description of TRs and related proteins among a representative set of various Deinococcus species, the genomes of which have been integrated into the MicroScope platform for microbial comparative genome analysis and functional annotation [28]. These species have been isolated from locations in North America, South America, Europe, Africa, and Asia (Table 1). The TRs and related proteins (locus tags) of these bacteria are listed in Table 2. The presence of most of these proteins has been reported in D. radiodurans and Deinococcus deserti, for which several shotgun proteomics studies have been performed [5,29,30,31,32,33]. We analyse the proteins notably with regard to their sequences, to the Cys distribution, and to the tridimensional structures and compare them with TRs from main model species (generally E. coli and/or Bacillus subtilis). We highlight the specificities of Deinococcus TRs and discuss based on available expression and phenotype data their physiological functions in line with the remarkable tolerance of this bacterial genus.
2. The TrxR/Trx System in Deinococcus
The two main components of the Trx system, TrxR, an NADPH-dependent flavoenzyme, and Trx, a small disulfide reductase, harbour conserved motifs, including two catalytic cysteines, allowing the transfer of reducing power via mixed-disulfide intermediates (Figure 1). This thiol-based system was initially discovered in E. coli through its role in DNA synthesis as an electron donor for ribonucleotide reductase [47]. Since then, this ubiquitous redox system, which regulates protein dithiol/disulfide balance using electrons provided by NADPH, was described as participating in a wide array of signalling and regulation pathways, and also in protective and repair mechanisms [10,15]. E. coli strains deleted for trx genes exhibit increased resistance to H2O2, probably because the more oxidising redox potential causes increased expression of catalase and alkyl hydroperoxidase in these mutants, but is more sensitive to diamide, a compound inducing disulfide bridging [48]. The key roles of the Trx system in the preservation of redox homeostasis and in responses to oxidative stress (Figure 1c) have been extensively documented in many prokaryotes and eukaryotes [10,11,15]. In Deinococcus, the Trx system has been more recently characterised, and in the next sections, we will describe the characteristics of TrxR and Trx families in this genus and what is currently known about their functions.
2.1. Deinococcus Thioredoxin Reductases
TrxR is a homodimeric flavoprotein belonging to the pyridine nucleotide–disulfide oxidoreductase family, which also includes enzymes such as glutathione reductase, alkyl hydroperoxide reductase, and lipoamide dehydrogenase [49]. Bacterial TrxRs, like those of archaea, fungi and plants, are composed of two subunits of 35 kDa, each possessing FAD-(flavin adenine dinucleotide) and NADPH-binding domains. These low-molecular-weight TrxRs are characterised by a specific fold found in several types of enzymes that catalyse diverse chemical reactions [50]. The electron transfer from NADPH via FAD to the active site disulfide, located in the NADPH-binding domain, leads to a large conformational change [51]. Bacterial TrxR-active sites contain two redox-active Cys within CATC or CAVC motifs in E. coli and M. tuberculosis, or Bacillus anthracis and S. aureus, respectively [10]. A search in the genomes of representative Deinococcus species revealed the presence of highly conserved TrxR enzymes, with more than 70% identity between them, and harbouring a CATC motif (Figure S1). Only one trxR gene is present in most Deinococcus species, as is the case in E. coli and B. subtilis. Remarkably, D. geothermalis and D. peraridilitoris have two trxR genes encoding very close proteins (98% and 91% identity, respectively). While Deinococcus TrxRs possess an active site sequence motif similar to that of E. coli, they share more overall homology with TrxR of B. subtilis. Thus, D. radiodurans TrxR (locus tag DR_1982) has 43% and 47% identity with E. coli and B. subtilis counterparts, respectively. Deinococcus and B. subtilis TrxRs do not possess extra Cys residues, such as the E. coli enzyme. Of note, Deinococcus TrxRs are characterised by the presence of a threonine just before the active site, this residue being an alanine or a tyrosine in TrxRs from E. coli or B. subtilis, respectively. There is some variability regarding the identity of this residue among bacterial TrxRs since a threonine is present in the Helicobacter pylori representative and a serine in the M. tuberculosis representative. Whether these residues modulate the activity of redox-active cysteines remains to be investigated.
In a pioneer work regarding the characterisation of the Trx system in Deinococcus, Obiero et al. [22] investigated the biochemical and structural features of TrxR of D. radiodurans. Using recombinant proteins, they showed that the reductase provides electrons to D. radiodurans Trx2 (DR_A0164) at the expense of NADPH, and also to E. coli Trx1, but with a lower affinity. The overall structure of D. radiodurans TrxR is very similar to that of TrxRs from other bacteria, with both NADPH- and FAD-binding domains containing variants of the canonical Rossmann nucleotide-binding fold. However, when compared with E. coli TrxR, some differences in the shape and charge distribution in the Trx-binding pockets were noticed and presumed to underlie the species-specific affinity previously mentioned [22]. This group further analysed the residues involved in TrxR–Trx interaction using computational alanine mutagenesis and by accurate analysis of the interface [23]. They concluded that four residues (M84, K137, F148, and F149) in D. radiodurans TrxR mainly account for the stability of the interface and the preferential binding affinity towards D. radiodurans Trx2. There is currently no genetic study unveiling the physiological function of TrxR in Deinococcus. In D. radiodurans, increased levels of the trxR transcript and TrxR protein have been detected after exposure to cadmium and UV/vacuum, respectively [32,52]. Using inhibitors of TrxR, Maqbool et al. [26] reported that D. radiodurans cells treated with auranofin or ebselen showed increased H2O2 sensitivity and increased levels of intracellular reactive oxygen species (ROS) and protein carbonyls compared with cells treated with H2O2 alone, indicating altered redox homeostasis when TrxR is inhibited.
2.2. Deinococcus Thioredoxins
TrxR supplies electrons to Trx, which is a small heat-stable disulfide reductase of ~10–12 kDa. Canonical Trxs carry a WCGPC-active site motif and share a conserved fold, the basic Trx-fold being composed of three α-helices surrounding four β-strands. The N-terminal catalytic Cys is present mainly as a thiolate, and thus can initiate a nucleophilic attack on oxidised Cys residues present in substrate proteins. This leads to the formation of mixed-disulfide intermediates and triggers deprotonation of the second Cys of the Trx-active site, named resolving Cys, which will catalyse the reduction of the heterodimer and the release of reduced substrate and oxidised Trx (Figure 1b). The E. coli genome, like many other bacterial species, contains two trx genes, trxA and trxC, which encode Trx1 and Trx2, respectively. In E. coli, a substantially higher level of expression of trxA has been reported compared with that of trxC. E. coli strains lacking both trx genes are still viable [48], very likely due to partial overlapping functions with the GSH-dependent system [13]. However, in other bacterial species, such as B. subtilis, Rhodobacter sphaeroides, and Synechocystis sp. PCC 6803, deletion of the trxA gene leads to loss of viability [53].
The characterisation of the E. coli Trx1 interactome in vivo has led to the identification of more than 250 proteins likely to be redox-regulated at the level of their Cys residues and participating in a wide diversity of cellular processes [54]. Similar conclusions, about the broad spectrum of Trx partners, have been drawn from the comprehensive characterisation of Trx interactome in other organisms, such as mammalian or photosynthetic cells [55,56]. Altogether, these findings clearly validate the central role of Trx in the control of most, if not all, cellular processes in living organisms and highlight its importance in redox biology [15].
The Trx1 isoform represents the most typical Trx form containing around 110 residues without any sequence extension or additional domain. Six of the analysed Deinococcus species possess a single trxA gene (Figure S2). Surprisingly, three trxA genes are present in D. peraridilitoris, two being located on a plasmid and encoding proteins more similar to each other (68% identity) than to the chromosome-encoded Trx1 protein. One of these plasmid genes, Deipe_3901, is located directly next to the second trxR gene (Deipe_3902) (Figure S3). D. radiodurans Trx1 (DR_0944) shares between 76% and 92% identity with Trx1 from other Deinococcus species (55% and 60% with the extra Trx1 from D. peraridilitoris) and 44% and 49% with Trx1 from E. coli and B. subtilis, respectively, revealing a closer relationship with the latter, as found for TrxR (Figure S1). Some specificity in Deinococcus Trx1 sequences can be noticed in the proximity of the active site motifs. Indeed, whereas the residues preceding the WCGPC-active site are well conserved in bacterial Trx1 proteins, the two subsequent ones differ in chromosome-encoded Deinococcus Trx1s (generally RI) compared with those (KM) observed in E. coli and B. subtilis Trx1 (Figure S2). This specificity might confer modified catalytic efficiency or altered access of the active site in Deinococcus proteins, but this needs to be further investigated by comparing the biochemical features of mutated recombinant proteins.
The trxC gene, encoding Trx2, is present only in three Deinococcus species (D. radiodurans, D. deserti, and D. geothermalis) analysed here. Trx2 is characterised by the presence of an additional N-terminal domain of 30 to 35 residues, including 2 other CXXC motifs [57] involved in zinc coordination [58]. This extension has been proposed to modulate Trx activity. Furthermore, if conditions of oxidative stress would affect zinc binding, the released zinc might contribute to a redox signalling pathway [59]. The three Trx2 isoforms from Deinococcus share between 51% and 66% identity and around 42% with E. coli Trx2 (Figure S2). Some differences are observed regarding the residue just preceding the WCGPC active site motif (T in D. geothermalis instead of P in other species) and in the one following this motif (V in Deinococcus Trx2 proteins instead of N in E. coli Trx2). Again, these specific features might be associated with changes in catalytic properties.
Similar to E. coli, a substantially higher expression of Trx1 compared with Trx2 has been observed in D. deserti under standard conditions, at both the mRNA and protein levels [31]. Trx1 is also more abundant than Trx2 in D. radiodurans [30,33], and both trxA and trxC showed enhanced expression following treatment with H2O2 [60]. Currently, there are only little data available regarding the biochemical properties and physiological functions of Trxs in Deinococcus. In their pioneer work on the Trx system in D. radiodurans, Obiero et al. [22] cloned the trx gene encoding the Trx2 protein (but improperly named Trx1 since it includes the sequence encoding the N-terminal extension). The recombinant protein catalyses reduction of insulin disulfide bridges, a typical feature of most Trx enzymes, and is reduced by D. radiodurans TrxR. Very recently, Kim et al. [60] provided data regarding the two Trx isoforms from D. radiodurans. First, they characterised their biochemical properties and showed that both display insulin and DTNB (5,5′-dithiobis(2-nitrobenzoic acid), a small-molecule disulfide) reduction activities in the presence of NADPH and TrxR. These activities were completely lost when active site Cys residues in Trx were replaced by Ser. Consistent with the regulatory role of the additional zinc-binding CXXC motifs in Trx2, substitution of Cys by Ser in these motifs impaired its reduction capacity. Of particular interest regarding Trx2, Kim et al. [60] determined its crystal structure and noticed that the orientation of the N-terminal Zn finger domain within the overall structure and with respect to the Trx-fold was clearly different in D. radiodurans from that observed in other bacterial Trx2 isoforms. They proposed that this unique feature could modulate protein–protein interactions, and be involved in the recognition of specific targets in this species. Finally, these authors also constructed D. radiodurans strains deleted for trxA or trxC, and showed that both mutants exhibit strongly reduced survival rates when treated with 60 mM H2O2. Interestingly, the mutant lacking Trx2 appeared to be more sensitive to this reactive oxygen species compared with that impaired for Trx1. Therefore, even though Trx2 is not common to radiation-tolerant Deinococcus species, it seems to contribute to stress tolerance in the species possessing this protein. These data, combined with those of Maqbool et al. [26] on TrxR, clearly show the key role of the TrxR/Trx system in the tolerance of Deinococcus to oxidative stress. No data about mutants lacking either trxR or both trxA and trxC have been reported, perhaps because such mutants are not viable.
2.3. Trx-Like Proteins in Deinococcus
The analysis of the D. radiodurans genome revealed the presence of several other genes (DR_2085, DR_A0072, DR_0057, DR_B0110, and DR_0948) encoding small proteins harbouring CXXC motifs, such a CPDC, CHLC, and CPGC (Table 3), and predicted to display a Trx-fold (Figure S4). These proteins are present in most Deinococcus species analysed here and divided in several subclasses (Table 2; Figure S5e) [3,5]. Based on sequence homology, some of these proteins were first classified as glutaredoxin-like proteins. However, since Grxs generally display a CPYC active site motif and, most importantly, get their reducing power from glutathione, which is absent in Deinococcus, these reductases should rather be considered as Trx-like proteins. Consistently, DR_2085-type proteins contain a specific structural motif (Figure 2a), SGFRP, which is found in E. coli NrdH, a disulfide reductase possessing a canonical C(M/V)QC active site, participating in the reduction of NrdEF, a ribonucleotide reductase, and supplied with electrons by TrxR [61]. A deep analysis of the genomes of various Deinococcus species revealed variability in the number of genes encoding such Trx-like proteins: for instance, three in D. deserti and six in D. gobiensis. The DR_2085 and DR_0057 types, which have CPDC and CHLC active sites, respectively, are present and well conserved in all species selected (Figure 2a; Figure S5a). For D. radiodurans and D. deserti, proteomics indicated low levels of expression of these proteins under standard growth conditions, but their detection might have been limited because of their small size. In the next parts, we will mainly focus on these two Trx-like types, notably in relation to the knowledge gained on related reductases in other organisms.
2.3.1. Trx-Like Proteins Carrying a CPDC Active Site Motif
Thioredoxin-Related Proteins of 14 kDa
Among Trx-like proteins present in Deinococcus, the DR_2085 type is present in all species selected for this study and includes well-conserved members sharing between 74% and 92% identity with DR_2085 (Figure 2a). This Trx type is characterised by the presence of a WCPDC active site. Interestingly, this motif is typical of a specific family of Trx-like proteins named TRP14 for Trx-related protein of 14 kDa or Clot in plants [62]. TRP14, first characterised in mammalian cells [63], is present and well conserved throughout the animal kingdom as well as in plants [64,65,66]. TRP14-related proteins are also present in yeast, but are less represented in bacteria. One ortholog, encoded by locus HP_1458, was identified in H. pylori [67] and shown to be important for maintaining DNA and protein integrity, and for survival under oxidative stress conditions [68]. The WCPDC motif is also present in the thioredoxin-like proteins YdpP and YtpP from B. subtilis, and in YtpP (BA_4945) from Bacillus anthracis [69]. Compared with TRP14s from other organisms, Deinococcus TRP14s are shorter, since they do not display an N-terminal extension (Figure 2b). Furthermore, except for the WCPDC motif, sequence similarity is low between these proteins. Alignment of the sequences of the two TRP14-related proteins from D. radiodurans and H. pylori revealed 15% identity and 30% similarity, while 13% identity and 25% similarity exist between DR_2085 and human TRP14 (Figure 2b). Notably, in addition to the conserved WCPDC active site motif, these proteins share a (V/I)PTL(V/I/L) motif in their C-terminal part.
Mammalian TRP14 has been shown to get electrons from TrxR and displays very poor ability to reduce insulin and typical Trx substrates, such as ribonucleotide reductases, peroxiredoxins, or methionine sulfoxide reductases [63]. Consistently, B. anthracis TRP14 could reduce DTNB, but not insulin in the presence of NADPH and B. anthracis TrxR [69]. Note that H. pylori TRP14 exhibits disulfide reductase activity in both the DTNB and insulin assays [67]. Most interestingly, human TRP14 can efficiently reduce L-cystine, a source of cysteine [70], and also nitrosylated or persulfidated cysteines in proteins [70,71]. Based on these biochemical properties, TRP14 reductases are considered as main modulators in redox signalling pathways and could be involved in protection against metal stress or pathogenic viruses [66]. Whether the DR_2085-type proteins fulfil similar activities and physiological functions in stress responses of Deinococcus remains to be investigated. Induced expression of the DR_2085 gene and its Deide_06390 homolog has been observed after irradiation [31,72,73] and upregulation of DR_2085 upon cadmium exposure [52].
DR_A0072-Related Proteins
Another CPDC-containing Trx-like protein (DR_A0072) has been identified in D. radiodurans, with an Asp preceding this motif, that is present neither in Trx-like such as TRP14 nor in canonical Trxs. Homologs of DR_A0072 sharing between 61% and 70% identity are present only in three of the analysed Deinococcus species (D. geothermalis, D. gobiensis, and D. proteolyticus). Note that these homologs do not harbour a CPDC motif. Such a motif is present in another plasmid-encoded D. proteolyticus homolog (Deipr_2732), displaying less identity (41%) with DR_A0072 (Figure S5b). Interestingly, the DR_A0072 gene is located between a gene encoding a heavy metal translocating P-type ATPase and a gene encoding an ArsR family regulator. This is similar in the three other Deinococcus species, suggesting a possible role of this Trx-like protein in the maintenance of metal homeostasis. Consistently, the DR_A0072 gene is upregulated after exposure of D. radiodurans to cadmium [52].
DR_B0110-Related Proteins
A CPDC motif is also present in DGo_PA0204 from D. gobiensis. However, this motif is not entirely conserved. Instead, a CPXC motif is rather present in homologs found only in three other Deinococcus species, including DR_B0110. The latter has 48% to 53% identity with Dgeo_1776, DGo_PA0204, and Deipr_2190 (Figure S5c). Interestingly, in the four species, the DR_B0110 gene homologs are preceded by and probably in operon with nrdIEF gene homologs, suggesting that these Trx-like proteins could participate in the modulation of ribonucleotide reductase activity, thus being functional equivalents of NrdH proteins. Expression of the DR_B0110 gene was induced after exposure to gamma radiation [72,74].
2.3.2. Trx-Like Proteins Carrying Other Active Site Motifs
DR_0057-Related Proteins
Another type of Trx-like proteins represented by DR_0057 in D. radiodurans is present in the seven Deinococcus species selected. DR_0057 shares between 41% and 65% identity with the six homologs. These proteins are 81 to 106 amino acids long (Figure S5a) and possess a low similarity both with NrdH-redoxin family proteins and proteins possessing a glutaredoxin-like domain (DUF836). In the absence of the C-terminal SGFRP motif typical of NrdH-redoxins, they cannot be classified as such. Instead, they share a dicysteinic CxLC motif that resembles the one present in DUF836 members. With the notable exception of the D. proteolyticus representative (SCKLCQ motif), other members of the Deinococcus genus analysed exhibit a more conserved GCHLCE motif.
Little information is known among the few members of this class. The tridimensional structure of a related protein from Mus musculus has been deposited in the protein databank (pdb code 1WJK), but never published. The yeast protein was referred to as MGP12 (mitochondrial glutaredoxin-like protein of 12 kDa) because it was identified in a study focusing on the mitochondrial proteome of this organism [75]. However, this name obviously does not fit with the widespread presence in prokaryote organisms. Indeed, in addition to their presence in eukaryotes including fungi, plants, and metazoans, homologs are present in some bacterial and archaeal genera. The sequences are highly divergent, and for example, only 17% identity and 17% similarity exist between DR_0057 and Arabidopsis MGP12 (Figure 2c). In fact, only the cis-proline that is present in all Trx superfamily members and the CxLC motif are strictly conserved.
DR_0948-Related Proteins
Finally, another type of Trx-like protein is present in six out of the seven Deinococcus species analysed (no homolog in D. peraridilitoris), the corresponding protein in D. radiodurans being DR_0948. The sequence is well conserved, with DR_0948 sharing 64–86% identity with the proteins from the five other species (Figure S5d). They also exhibit longer sequence (ca 150 residues) than that of canonical Trxs, but differ in the second residue of the active site motif (CSGC, CPGC, or CAGC). In D. deserti, the expression of the Deide_06780 gene is induced by radiation [31].
2.4. FrnE Oxidoreductase
FrnE subfamily proteins are predicted cytoplasmic thiol oxidoreductases belonging to the DsbA family (domain cd03024 in the Conserved Domain Database) [76]. They contain a CXXC motif in a DsbA-like thioredoxin domain. FrnE homologs are present in many bacteria, and many possess an additional conserved CXXXXC motif at the C-terminus. The name frnE was given for the first time to a gene present in a gene cluster (frnA to frnU) associated with the biosynthesis of the polyketide antibiotic frenolicin B in Streptomyces roseofulvus (GenBank accession AF058302.1). An frnE gene homolog is present in Deinococcus species but not located in an frn gene cluster as in S. roseofulvus. FrnE (DR_0659, drFrnE) from D. radiodurans has been studied. The expression of DR_0659 is upregulated after exposure of D. radiodurans to gamma radiation and desiccation [74], cadmium [52], and UV/vacuum [32]. Compared with the wild-type strain, a DR_0659 disruption mutant showed increased sensitivity in particular to cadmium and diamide, and less markedly to gamma rays and H2O2 [77]. Recombinant drFrnE showed disulfide isomerase and insulin reduction activities in vitro. In addition to the 22-CXXC-25 active site motif located near the N-terminus, drFrnE has a C-terminal tail harbouring a 239-CXXXXC-244 motif. Four different drFrnE crystal structures have been solved, with the crystals grown in the absence or presence of different reducing agents [78]. A disulfide bond between Cys22 and Cys25 was observed in the absence of a reducing agent. In all structures, the C-terminal tail of one molecule interacts with the active site of another drFrnE molecule, with the C-terminal CXXXXC located close to the active site CXXC. In crystals grown in the presence of DTT, an intermolecular disulfide bond was observed between Cys244 and Cys22. The C-terminal cysteines are not required for insulin reduction activity in the presence of DTT. The drFrnE protein could also reduce DTNB in the presence of NADPH and E. coli TrxR, but this activity was decreased when the C-terminal cysteines were mutated or deleted. Based on structural and biochemical data, the authors proposed that drFrnE is a cytoplasmic disulfide reductase and that the C-terminal CXXXXC is functionally important with a role in the regeneration of the active site [78]. DR_0659 shares 50–69% identity with FrnE from five species analysed here, but only 37% with FrnE from D. maricopensis (Figure S6a). They all have the active site motif CP(W/F)C. Remarkably, FrnE from D. maricopensis, like S. roseofulvus FrnE, lacks the C-terminal CXXXXC motif (Figure S6a), suggesting that the regeneration mechanism of the oxidised form is different for these homologs.
In addition to FrnE, two other predicted cytoplasmic proteins with DsbA-like domains are present in several Deinococcus species. One of these, belonging to the FrnE-like subfamily of the DsbA family (cd03025), is encoded by the genomes of D. deserti and D. proteolyticus. These two proteins (Deide_3p01230 and Deipr_1559) share 37% identity and have a CGWC or CIWC motif but no other Cys residues (Figure S6b). Another protein with a DsbA-like thioredoxin domain, but with only a single conserved Cys residue, is found in five of the analysed species (e.g., DR_2335 in D. radiodurans) (Figure S6c). To our knowledge, no experimental data concerning the function or expression have been reported for these proteins.
2.5. Periplasmic Disulfide Oxidoreductases
Dsb (disulfide bond) family proteins such as DsbA to DsbE and DsbG from E. coli are involved in the formation or reduction of disulfide bonds, and in the repair of oxidised Cys residues in extracytoplasmic proteins [14]. They have redox-active cysteines, including CXXC motifs. DsbA, DsbC, and DsbG are periplasmic proteins synthesised as precursors with an N-terminal signal peptide. DsbA catalyses the formation of disulfide bonds in proteins exported to the periplasm. DsbA is reoxidised by the cytoplasmic membrane protein DsbB. The DsbA and DsbB homologs in B. subtilis are called BdbD and BdbC, respectively, with BdbD attached to the membrane by an N-terminal transmembrane segment [79]. DsbC and DsbG are part of a periplasmic reducing system that repairs oxidised Cys of single-Cys-containing extracytoplasmic proteins [80]. DsbC also catalyses the isomerisation of incorrect disulfide bonds that are often introduced by DsbA, resulting in proper disulfide bonds and protein folding. DsbE, also called CcmG, reduces the disulfide bond that is formed in apocytochrome c in the periplasm to allow heme ligation. DsbE belongs to the TlpA-like family (domain cd02966 in the Conserved Domain Database) [76], also including TlpA (e.g., from Bradyrhizobium japonicum), ResA (B. subtilis), and similar proteins with a CXXC motif-containing Trx domain in the periplasm. DsbC, DsbG, and DsbE are maintained in the reduced state by the cytoplasmic membrane protein DsbD that catalyses the transfer of electrons from the cytoplasmic NADPH–TrxR–Trx system across the membrane. Many bacteria, such as B. subtilis and H. pylori, possess a shorter DsbD analog called CcdA [14,81].
Deinococcus species have several predicted Dsb proteins, but there is variation in the presence/absence and number of these proteins across the different species (Table 2). To our knowledge, these proteins have not been studied experimentally. The B. subtilis bdbD and bdbC genes are present in an operon (bdbDC). Similarly, two adjacent genes encoding DsbA and DsbB family proteins showing sequence similarity to B. subtilis BdbD and BdbC are present in six of the analysed Deinococcus genomes (Figure S7). Like B. subtilis BdbD, these DsbA family proteins (e.g., DR_0753) have a predicted N-terminal transmembrane segment and a KCPXCK active site motif (Figure S8a). Unlike B. subtilis BdbD, but similar to E. coli DsbC, two additional conserved Cys residues are present, which might form a disulfide bond as in DsbC. D. proteolyticus has an additional more distant homolog (Deipr_2421) encoded by a gene on a plasmid and not adjacent to a bdbC-like gene. The BdbC (DsbB family) homologs (e.g., DR_0754) have a PCXLCW motif and two other conserved Cys residues (Figure S8b). The same six Deinococcus species possess another (two in D. radiodurans: DR_2019 and DR_0560) DsbA-related protein having a predicted signal peptide, a QCPYC motif, and two other conserved Cys residues (Table 2; Figure S8c). Experimental work is needed to establish whether these deinococcal DsbA family proteins have a role, like E. coli DsbA, in generating disulfide bonds in extracytoplasmic proteins or are functionally similar to DsbC or DsbG. Remarkably, DsbA and DsbB family proteins are absent in D. peraridilitoris (Table 2).
As reported previously [82], bdbDC homologs in several Deinococcus species are directly preceded, and possibly in operon [31], by a gene encoding a Deinococcus-specific membrane protein designated dCSP-1 and the gene encoding the DNA repair protein UvrA (Figure S7). It has been proposed that BdbD (DsbA) and, on the cytoplasmic side, UvrA may interact with dCSP-1, and that UvrA may receive electrons from BdbC (DsbB) to reduce oxidatively damaged Cys residues in CXXC motifs of UvrA, restoring its nonoxidised functional state [82]. However, as mentioned above, D. peraridilitoris does not have DsbA and DsbB proteins, but possesses UvrA and dCSP-1 (Figure S7).
Homologs of E. coli DsbD were not found in Deinococcus. However, at least one homolog of the DsbD analog CcdA is present in this bacterial genus. Except for the ccdA gene (DR_1300) in D. radiodurans, ccdA homologs are located in gene clusters encoding cytochrome c-type biogenesis proteins CcmA to CcmH in the other species, albeit with some variation in the genetic organisation (Figure S9). A ccmG (dsbE) gene is also present in these gene clusters. D. deserti and D. peraridilitoris have an additional two-gene cluster encoding homologs of CcdA and CcmG (DsbE) (Figure S9). The CcdA proteins have two conserved Cys surrounded by other conserved residues (Figure S8d). The CcmG (DsbE) proteins contain a WXXWCXXC motif as found in TlpA-like family proteins from other bacteria, such as E. coli DsbE and B. subtilis ResA (Figure S8e). The CXXC motif of the DsbE homolog present in each Deinococcus contains a Pro (CXPC) as in ResA and in the uncharacterised YneN of B. subtilis. The additional DsbE homologs from D. deserti (Deide_2p00420) and D. peraridilitoris (Deipe_0660) have a divergent CLVC motif. Deinococcal CcdA and DsbE (CcmG) proteins may be required only for cytochrome c maturation (reduction of disulfide bond in apocytochrome), but could also participate in the reduction of other substrates in the periplasm, and have a role in defence against oxidative stress, as reported for such proteins in other bacteria [83,84,85,86]. Alternatively, and in particular for additional proteins in D. deserti and D. peraridilitoris, DsbE might function as an oxidant instead of a reductant, as observed for DsbE from M. tuberculosis [87], and be involved in the formation of disulfide bonds since, as described above, no DsbA family proteins were found in D. peraridilitoris.
3. Main Trx Partners in Deinococcus
In addition to their roles in the modification of the structure or activity of numerous partners via redox post-translational modifications, Trxs supply electrons to several types of thiol reductases and peroxidases that have critical roles in the maintenance of redox homeostasis. Among them, two main types of enzymes, methionine sulfoxide reductases (Msrs) and peroxiredoxins (Prxs), control the redox status of proteins and scavenge peroxides, respectively (Figure 1c). Both enzyme types fulfil their biochemical function thanks to catalytic redox-active cysteines.
3.1. Methionine Sulfoxide Reductases
Oxidation of Met results in the formation of Met sulfoxide (MetO) and Met sulfone (MetO2), the latter modification being irreversible. MetO proportion can reach up to 40% of total Met during acute oxidative stress conditions [88]. MetO is mainly reduced back to Met by Msrs [89]. Oxidation of Met generates S- and R-MetO diastereoisomers, and two Msr types, A and B, specifically reduce these isomers, respectively. Although sharing a very similar thiol- and sulfenic acid-based redox chemistry, they do not exhibit any sequence similarity [90,91]. Of note, MsrA is able to reduce both peptide-bound and free MetO forms, in contrast to MsrB, which reduces the peptide-bound form in a more efficient way [92]. Most Msrs achieve their activity thanks to two redox-active Cys and a three-step catalytic mechanism following MetO reduction: (i) formation of a Cys sulfenic acid intermediate, (ii) formation of an intramolecular disulfide bond, and (iii) disulfide reduction generally by Trx and regeneration of activity [91,93]. Note that the regeneration of some Msrs having only one catalytic Cys is ensured by other thiol-containing compounds, such as Grxs [93,94].
Thiol-utilising Msrs are present in most living organisms, except in a few eubacteria and in one group of archaea [95]. They fulfil essential functions in stress tolerance and during ageing through modulation of the Met redox status in proteins [96,97]. In pathogenic bacteria, Msrs are considered essential determinants underlying their virulence and capacity to counteract host defences [98]. Thus, modification of the expression of msr genes leads to impaired responses to oxidative stress notably due to defective protein repair [99]. Consistently, Henry et al. [100] reported that the ubiquitous recombinase RecA, which promotes DNA recombination and induction of the SOS repair system, is a substrate of both Msr types. Indeed, oxidation of two Met residues in E. coli RecA is associated with dramatic changes in DNA recombination activity and SOS induction level, and the Msr system allows preserving the pool of functional RecA. Besides a repair function, these enzymes have also been proposed to play a direct antioxidant function by eliminating ROS via cyclic oxidation of Met in proteins [101]. Most importantly, a lot of evidence highlighted that the control of Met redox status is involved in the transduction of redox signals [102,103,104]. For instance, exposure to HOCl of E. coli cells results in activation via Met oxidation of the HypT transcription factor and enhanced expression of genes involved in protective mechanisms against this harmful agent [105].
In Deinococcus, the knowledge regarding the maintenance of Met redox status has only recently been explored. When submitting E. coli and D. radiodurans cells to ionising radiation (1000 Gy), Bruckbauer et al. [106] observed substantially greater oxidative modifications in E. coli proteome. Of note, methionine sulfoxide formation was the most frequent modification in E. coli, for which 137 instances were counted, while only one was detected in D. radiodurans. In another proteomic-based analysis, Chang et al. [107] exposed these two bacteria to 6700 Gy, a radiation level lethal for E. coli and causing damage in D. radiodurans. A much lower level of carbonylation was measured in the latter. These authors investigated whether protein-intrinsic properties could underlie this differential tolerance and noticed that in permissive conditions Met was less prevalent in D. radiodurans than in E. coli. In contrast, following irradiation, Met prevalence decreased in the latter, but strongly increased in the former, suggesting the occurrence of a more efficient Msr system under oxidative stress in D. radiodurans [107]. Consistently, using structure- and sequence-based models, E. coli MsrB was predicted to be intrinsically more sensitive to carbonylation than its Deinococcus counterpart. These data corroborate the likely importance of Msr proteins in the tolerance of Deinococcus species to severe stress conditions.
3.1.1. MsrA Proteins in Deinococcus
Similar to most non-photosynthetic species, Deinococcus has only one msrA gene. MsrA proteins from the seven species analysed here share 58% to 77% identity and about 45% identity with those of E. coli and B. subtilis (Figure S10a). They all display four conserved Cys, which are at positions 25, 28, 60, and 170 in DR_1849, the Cys25 in the conserved motif A(I/L/F)GGCFWCT being the catalytic one. This motif is close to the consensus sequence present in most MsrAs [91,108]. Of note, this motif displays some specificities in Deinococcus, which are not observed in E. coli and many other organisms. For instance, it includes a second Cys at position 28, whereas Gly or Ser is generally present at this position. Interestingly, this second Cys is also present in B. subtilis MsrA. In E. coli MsrA, Cys198 has been identified as the resolving Cys allowing activity regeneration of the enzyme [109]. Based on sequence alignment, we presume that the resolving Cys in Deinococcus reductases is the one corresponding to Cys170 in DR_1849. Upregulation of DR_1849 gene expression or protein abundance has been observed after exposure of D. radiodurans to radiation [32,74] or cadmium [52].
3.1.2. MsrB Proteins in Deinococcus
Like in most prokaryote genomes [110], one msrB gene is present in Deinococcus. Among the seven species selected for this study, six MsrB proteins display high homology between them (72% to 82% identity) (Figure S10b). MsrB (Deipe_4299) from D. peraridilitoris is more divergent since it exhibits only 51% to 55% identity with MsrBs from other species. Deinococcus MsrBs (except Deipe_4299) are closer to the one of B. subtilis (ca. 62% identity, against ca. 48% when compared with E. coli MsrB). Interestingly, D. peraridilitoris MsrB possesses two CXXC motifs like homologs from E. coli and eukaryotes [108,111]. These motifs are presumed to be involved in Zn fixation and structural stabilisation of the protein, and are absent in the reductases of the six other Deinococcus species analysed here and of B. subtilis. In other respects, all Deinococcus MsrBs share two well-conserved Cys, at positions 71 and 126 in DR_1378, corresponding to Cys63 and Cys117, respectively, in E. coli MsrB. Analysis of the MsrB catalytic mechanism [112] revealed that the latter, which is included in the well-conserved active site motif RYC(I/V)N, corresponds to the catalytic Cys, while the former named resolving Cys is present in the motif GCGWP. Based on these sequence features, we conclude that Deinococcus MsrBs belong to the 2-Cys type, which is regenerated by Trx following the reduction of an intramolecular disulfide bridge [113]. Induced expression of the DR_1378 gene was observed after exposure of D. radiodurans to cadmium [52], while its gene homolog Deide_04050 in D. deserti was induced after gamma radiation [31].
3.1.3. Non-Thiol-Based Periplasmic and Possible Cytoplasmic Msr Systems
As described in Section 2.5, the maintenance of Cys redox status in periplasmic proteins in bacteria is ensured by the thiol-dependent Dsb system thanks to reducing power provided by cytoplasmic Trx. In contrast, non-thiol-dependent systems control the redox status of Met in periplasm [114], except in some bacteria, such as Neisseria or Helicobacter, in which Msrs A and B are present in this compartment as fusion proteins [115,116]. In most other bacteria, various types of molybdenum-containing enzymes exist to reduce MetO [114]. The MsrPQ (YedY/YedZ) system is present in a large subset of bacteria including E. coli and Pseudomonas aeruginosa [117]. In this system, electrons are provided by the respiratory chain via MsrQ, an inner membrane heme b-containing protein, to MsrP, which reduces MetO diastereoisomers thanks to a molybdopterin cofactor. Both MsrP and MsrQ are present and well conserved in all Deinococcus species analysed here (Table 2), the D. radiodurans representatives being DR_2536 and DR_2537, respectively. For DR_2536, upregulated gene expression has been observed following exposure to cadmium [52].
Other molybdenum-containing enzymes exhibiting MetO reductase activity are the cytoplasmic BisC and periplasmic TorZ and DmsA proteins present in E. coli and other bacteria. These are oxidoreductases containing the COG0243 or BisC domain [114]. Periplasmic homologs of this enzyme type are not present in Deinococcus. However, a predicted cytoplasmic protein (DR_0397 in D. radiodurans; Table 2) with only low levels of similarity (less than 30% identity) with BisC is conserved in Deinococcus. Furthermore, another predicted cytoplasmic molybdenum-containing enzyme belonging to the sulfite oxidase family and containing the COG2041 (YedY) domain also present in MsrP is highly conserved in Deinococcus (DR_0716 in D. radiodurans; Table 2). Whether these uncharacterised proteins have MetO reductase activity or another function remains to be established.
3.2. Thioredoxin-Dependent Peroxidases/Peroxiredoxins
Peroxiredoxins are ubiquitous enzymes reducing peroxides using a conserved cysteine, named peroxidatic cysteine [118,119]. These enzymes, which are generally highly abundant in cells, function without any cofactor and exhibit a remarkable efficiency in scavenging peroxides and peroxynitrites [118]. They are recognised as key actors in the maintenance of cell redox homeostasis via several functions, including elimination of peroxides, signalling due to hyperoxidation or via redox modulation of protein partners, and also a chaperone role in relation to their redox status and oligomerisation level [119,120,121]. Since their discovery in yeast [122], Prxs have been the subject of extensive work in all organisms. They are critical actors in responses to environmental constraints, upon diseases such as cancer, or during the course of ageing [118,119]. They also constitute essential virulence factors in pathogenic microorganisms [123].
Prxs are classified in six evolutionary distinct subfamilies (Prx1, Prx6, AhpE (alkyl hydroperoxide reductase E), PrxQ/BCP (bacterioferritin comigratory protein), Tpx (thiol peroxidase), and Prx5), which differ by the presence and position of resolving cysteine, the oligomeric state, and the type of electron donor [118]. All are characterised by the presence of a conserved motif, Pxxx(T/S)xxC, containing the peroxidatic Cys, the thiolate form of which attacks peroxides. The resulting Cys sulfenic acid is generally resolved by the formation of an inter- or intramolecular disulfide with a resolving Cys, which will be further reduced by Trx in most cases, except for AhpE and AhpC, a member of the Prx1 subclass. AhpC-type Prxs receive electrons from AhpF, a flavoprotein pyridine nucleotide–disulfide oxidoreductase related to, but different from, TrxR [124].
The survey of Deinococcus genomes revealed the presence of two types of Prxs, namely, PrxQ/BCP and AhpE (Figure 3), which are present in all analysed species of the genus [3] (Table 2). In this section, we will focus on the Trx-dependent PrxQ/BCP type, first identified and characterised in E. coli, and named bacterioferritin comigratory protein in bacteria [125], but PrxQ in plants [126]. In E. coli, the bcp gene is induced by oxidative stress conditions, and strains lacking it exhibit hypersensitivity to hydrogen and organic peroxides [125]. The first characterised bacterial or plant representatives of this class harboured two Cys, forming an intramolecular disulfide bond following the catalytic act, which was subsequently reduced by Trx [126,127]. However, sequence alignment of BCP proteins from prokaryotes revealed the absence of the resolving Cys in a majority of them and raised the question of the regeneration of their activity [127]. When investigating the catalytic mechanism of 1-Cys BCP from Burkholderia cenocepacia, Clarke et al. [128] observed that the formed Cys sulfenic acid could be reduced directly by Trx, or in a more efficient way by the GSH/Grx pathway.
With regard to Deinococcus, D. deserti and D. proteolyticus have only two bcp genes, whereas the other species possess at least three genes (Table 2). Among the three genes in D. radiodurans, DR_1208 possesses a single catalytic Cys, whereas the two other Prxs (DR_0846 and DR_1209) carry two redox-active Cys. DR_0846 is the closest homolog to E. coli and B. subtilis BCP proteins (40% and 43% identity, respectively) with redox-active cysteines separated by four residues. The percentage identity among Deinococcus DR_0846-related BCPs (Figure S11a) ranges from 64% to 77%. The remaining Deinococcus bcp genes also code for 2-Cys peroxidases, but about 38 residues (Figure S11b) separate the two conserved Cys. DR_1209 shares 45% to 74% identity with the DR_1209-related BCPs of the other species, and 48% with DR_1208. Of note, the Pxxx(T/S)xxC motif present in most BCPs is replaced by a Pxxx(G/A)xxC motif in 2 (out of 11) of these atypical BCPs. Furthermore, the presence of extra Cys was noticed in five of these BCPs (Figure S11b). These sequence features might alter the peroxidase catalytic properties and underlie specific interactions or regeneration pathways with electron donors. An additional, longer and more distant, predicted BCP-type Prx is present in D. gobiensis (Figure S11c). This DGo_CA0314 protein shares 64% identity with D. deserti Deide_23291, but the latter seems truncated. Both DGo_CA0314 and Deide_23291 may represent inactive BCPs since they lack the PxxxTxxC motif, and a 15-residue insertion is present in the corresponding region.
Cho et al. [25] investigated the expression levels of the three bcp genes in D. radiodurans exposed to oxidative stress generated by H2O2 or subjected to gamma irradiation. DR_0846 was strongly induced by both treatments, while the expression of DR_1209 was moderately increased by the peroxide, and that of DR_1208 was substantially triggered by irradiation. In proteomics studies, DR_0846 (and its D. deserti homolog Deide_10900) was well detected compared with DR_1209 and DR_1208 (and Deide_09051), suggesting higher expression levels of the DR_0846-type BCP under standard conditions [5,29,30,31,32,33].
The D. radiodurans DR_0846 BCP protein has been further characterised with regard to its activity and physiological function [25]. The recombinant protein exhibits peroxidase activity towards H2O2 and holdase chaperone activity when measuring thermal-induced aggregation of a heat-sensitive substrate. Interestingly, a D. radiodurans strain lacking DR_0846 displays similar sensitivity to H2O2 and heat stresses compared with WT. Most importantly, a strain lacking both DR_0846 and catalase exhibited increased sensitivity to these stresses compared with the strain lacking only catalase, indicating that the BCP peroxidase activity in Deinococcus may represent a second line of defence towards oxidative stress. In other respects, BCP proteins from various bacterial species have been reported to bind DNA in vitro and protect it against thermal and oxidative damage [129,130]. The physiological relevance of such a function remains to be established, but in the case of Deinococcus, where DNA damage is repaired in an efficient and rapid manner following extreme stress conditions, we can propose that BCPs, which are over-represented in this species, participate in such protective mechanisms.
4. Thioredoxin-Independent Thiol Peroxidases
Besides Trx-dependent Prxs, other less ubiquitous types of thiol peroxidases have been identified in the last decades. Among them, three are present in Deinococcus, namely, AhpE, AhpD, and OsmC/Ohr.
4.1. The AhpF/D-AhpC/E Thiol-Dependent Peroxidase Systems
The alkyl hydroperoxide reductase AhpF-AhpC system has been first identified in Salmonella typhimurium, and later in many other bacteria, yeast, and mammals [131,132,133]. This system, catalysing the reduction of peroxides at the expense of NADH, is composed of AhpF, a FAD-containing protein, which mediates electron transfer to the AhpC thiol peroxidase [124]. As already mentioned, AhpF, like TrxR, belongs to the family of pyridine nucleotide–disulfide oxidoreductases. Both share a domain harbouring two conserved catalytic Cys [131,134]. However, AhpF has a higher molecular mass (57 kDa) due to the presence of an N-terminal extension of 200 residues, including two other Cys ensuring electron transfer from the TrxR-like domain to AhpC [135]. AhpC is a 21-kDa thiol peroxidase belonging to the Prx1 subfamily initially named typical 2-Cys Prx [118]. These highly expressed Prxs function as dimers and form during the catalytic cycle an inter-subunit disulfide bond. Further, as a function of their redox status, they are able to oligomerise and fulfil then a molecular chaperone function [136,137]. The AhpF–AhpC system is widespread in microorganisms, and strains modified for its expression exhibit altered susceptibility to oxidative stress generated by hydrogen or organic peroxides [133,138].
AhpE is an atypical type of AhpC having only one Cys and possessing also both peroxidase and chaperone activities [139]. It has been first characterised in M. tuberculosis, where it shares 34% sequence identity with AhpC [140]. Most interestingly, AhpE has been found to display specificity towards long-chain fatty acid hydroperoxides [141]. Interestingly, another type of reductase named AhpD is able to provide electrons to AhpC [142]. AhpD shares little sequence homology but common structural features with AhpF [10]. This reductase is present in some bacteria (M. tuberculosis, Corynebacterium glutamicum) and cyanobacteria (Anabaena PCC7120) [143]. AhpD is reduced by a lipoamide-containing protein, dihydrolipoamide succinyltransferase or lipoamide reductase, using NADH as a source of electrons [142]. AhpD plays essential roles in response to oxidative stress [143,144] that could be carried out by direct peroxidase activity towards organic peroxides [144] or reduction of AhpC/AhpE [10,142]. However, the experimental evidence for the latter biochemical function remains to be firmly established regarding AhpE.
In Deinococcus genomes, there are no close homologs of ahpF and ahpC. However, all species possess one ahpE gene (DR_2242 in D. radiodurans). DR_2242 shares 53% to 82% identity with the homologs from the other six species and 35% identity with AhpE from M. tuberculosis (Figure S12a). In addition to the AhpE homolog Deipe_1016, the D. peraridilitoris genome encodes Prx-like protein Deipe_4202, which has less than 30% identity with the deinococcal AhpE homologs. Unlike other AhpE proteins, Deipe_4202 has a CxxC motif and lacks the PxxxT/SxxC motif. In M. tuberculosis, which does not possess GSH but another LMW thiol compound referred to as mycothiol (MSH), the reduction of AhpE is ensured by a MSH- and/or mycoredoxin-dependent mechanism [145]. This raises the question of the reducing system for AhpE in Deinococcus, in which the bacillithiol system is present. In other respects, all Deinococcus species harbour one ahpD-like gene (DR_1765 in D. radiodurans), two and five being present in D. deserti and D. peraridilitoris, respectively. Four of the ahpD genes of D. peraridilitoris are located on a plasmid, three being adjacent or close to extra trxR and trx genes in this species (Figure S3). However, these proteins show substantial variation in their sequences. DR_1765 shares 72–76% identity with Deide_13030, Dgeo_1446, and DGo_CA1027; 54% with Deima_0298; 36% with Deipe_3296; but only 17–25% with the other deinococcal AhpD-like proteins (including the single AhpD-like from D. proteolyticus) and 12% with AhpD from M. tuberculosis (Figure S12b).
Biochemical and structural investigations of the D. radiodurans AhpD-like protein were performed by Zhao et al. [146], who reported that a mutant strain lacking the reductase exhibits increased sensitivity to H2O2. Of note, complementation assays using wild-type AhpD revealed partial restoration of this phenotype, and this level of restoration was much lower when using AhpD variants mutated for the two Cys of the conserved CxxC motif [146]. In other respects, determination of a crystal structure revealed that it adopts a fold representative of the AhpD superfamily [146]. Taken collectively, these data strongly support a role of AhpD and AhpE enzymes in Deinococcus responses to oxidative stress via peroxidase activity for both or possibly through electron supply from AhpD to AhpE.
4.2. Ohr/OsmC/YhfA Thiol-Dependent Peroxidases
Another class of bacterial thiol peroxidases named Ohr (organic hydroperoxide resistance) has been identified by complementation with Xanthomonas campestris genes of E. coli strains deficient in the AhpF–AhpC system and susceptible to organic hydroperoxides [147]. This 14-kDa protein shares some sequence homology (around 20% identity) with other stress-induced proteins termed OsmC (Osmotically induced protein C) first identified in E. coli [148]. Ohr and OsmC are structurally related enzymes [149]. Atichartpongkul et al. [150] reported that these two subfamilies are widely distributed in Gram-negative and Gram-positive bacterial species. Later, members of the Ohr/OsmC family have been identified in several eukaryotic clades, especially in fungi but also in mosses, such as Physcomitrella patens [151].
Ohr/OsmC proteins display specific expression patterns depending on stress nature and on peroxide type. In P. aeruginosa, ohr expression is induced only by organic peroxides, and not by other oxidative or stress treatments, while osmC expression is specifically triggered by salt treatment [150]. The phenotype of strains lacking these genes and the presence of two conserved Cys (one positioned in a typical VCPY motif in Ohr) in both protein types indicated that they could fulfil a thiol peroxidase function [150]. This role in metabolising hydrogen and organic peroxides was demonstrated in vitro using recombinant Ohr or OsmC forms and in vivo by generating deletion strains in several bacterial species [152,153,154,155]. Moreover, Ohr was shown to participate in response to peroxynitrite in P. aeruginosa [152]. Site-directed mutagenesis revealed that the two Cys are critical for the catalytic activity [154], and provided clear evidence that Ohr functions thanks to reactive thiol groups. Of note, these enzymes do not share structural relationship with Prxs, such as BCP, AhpC, or AhpE, as shown by the determination of crystal structures of P. aeruginosa Ohr and E. coli OsmC [149,154] (Figure S13). Following peroxide reduction, these enzymes form an intramolecular disulfide bridge that Trx- and GSH-based systems are unable to reduce, but require lipoylated proteins as an electron source (Figure 3) [156]. Indeed, Xylella fastidiosa Ohr was found to interact with lipoylated proteins (dihydrolipoyl dehydrogenase LpdA, pyruvate dehydrogenase beta PDHB, and dihydrolipoamide succinyltransferase SucB), and its peroxidase activity was supported by PDHB and SucB in the presence of lipoamide or by LpdA alone [156]. Similarly, OsmC is reduced by lipoyl-dependent systems [156]. In C. glutamicum, a bacterial species highly resistant to organic hydroperoxides, OsmC activity was preferentially regenerated through a pathway involving the AhpD reductase, another lipoyl-dependent system [157].
Another family of proteins structurally related to Ohr/OsmC has been described by Shin et al. [149]. It is present in E. coli (YhfA) but also in other bacterial species, such as Vibrio cholerae. YhfA proteins possess the two cysteines that are well conserved in Ohr and OsmC peroxidases and likely to participate in the catalytic mechanism, but they lack two residues, one Arg and one Glu, neighbouring the peroxidatic cysteine [149]. The biochemical properties and physiological roles of this class of putative thiol peroxidases remain to be investigated.
Among the Deinococcus species analysed in this review, most possess genes encoding these three types of atypical thiol peroxidases (Ohr, OsmC, and YhfA corresponding to DR_1857, DR_1538, and DR_1177 in D. radiodurans), except D. deserti and D. proteolyticus, which do not have ohr and osmC genes, respectively (Table 2). Note also that the genomes of several species contain two ohr or yhfA genes. Deinococcus OsmC proteins share 54–83% identity, and around 42% with the E. coli homolog (Figure S14a). Concerning Ohr, D. radiodurans shares 55–78% identity with the closest homologs in the five other Deinococcus species, around 40% with the extra Ohr present in three of these species, and around 47% with the two Ohr proteins from B. subtilis (Figure S14b). YfhA homologs are present in the seven Deinococcus species (Figure S14c). They are divided into two subgroups, one containing highly conserved proteins in the seven species sharing 70–92% identity and around 35% with E. coli YhfA, and the second containing the two extra proteins from D. deserti and D. peraridilitoris having 71% identity between them but low-sequence identity with the other YhfA proteins (e.g., 21% between the two from D. deserti).
The knowledge about these atypical thiol peroxidases in Deinococcus remains limited. Atichartpongkul et al. [150] reported that in D. radiodurans, osmC (DR_1538) is induced by ethanol treatment and not by salt, while ohr (DR_1857) expression is enhanced by organic hydroperoxides. Cadmium exposure enhanced the expression of osmC, ohr, and yhfA in D. radiodurans [52]. Radiation-induced expression of D. deserti osmC [31] and D. radiodurans Ohr [158] has also been observed. The crystal structure of D. radiodurans Ohr has been solved [159] (Figure S13). Comparison with the structure of P. aeruginosa Ohr showed that Ohr proteins adopt two different conformations depending on the enzyme redox state. It was postulated that one configuration is consistent with efficient catalysis of the reduction of organic hydroperoxides, whereas the other form is required for enzyme recycling. Compared with wild-type D. radiodurans, a mutant strain deleted for osmC is more sensitive to gamma irradiation, particularly at high doses (10 kGy) [160], indicating the participation of OsmC peroxidase in the radiotolerance of Deinococcus.
5. The Bacillithiol System
Low-molecular-weight thiols are crucial for maintaining an intracellular reducing environment and for protection against ROS and other reactive species. Glutathione is the best-studied LMW thiol present, for example, in E. coli and eukaryotes. Deinococcus bacteria do not produce GSH but BSH, which is the most widespread LMW thiol in bacteria [161]. BSH is also involved in metal homeostasis [18]. The structure of BSH, the α-anomeric glycoside of L-cysteinyl-D-glucosamine with L-malic acid (Figure 4a), was determined after purification from D. radiodurans [162]. However, the BSH system has been studied most extensively in B. subtilis and S. aureus [18]. This system includes NADPH–BSH disulfide reductase (Bdr, previously called YpdA) and three bacilliredoxin (Brx) proteins (Figure 4b) [163,164,165]. Under oxidising conditions, BSH can form a disulfide (BSSB) and also mixed disulfides with protein thiols, a modification referred to as S-bacillithiolation. Like for S-glutathionylation and S-mycothiolation with GSH and MSH, S-bacillithiolation protects protein thiols from overoxidation, and may also function in redox regulation [19]. Brx proteins catalyse protein debacillithiolation, resulting in the formation of Brx-SSB. Oxidised Brx is likely preferentially reactivated by BSH or possibly by another Brx (see below) [18,165]. The regeneration of BSH from BSSB is catalysed by the NADPH-dependent Bdr enzyme [163,166] (Figure 4b). Recently, B. subtilis Bdr was shown to additionally have Brx reductase activity [165].
More than 50 proteins have been identified as S-bacillithiolated following hypochlorite or cumene hydroperoxide stresses in different Bacillus and Staphylococcus species. This includes glyceraldehyde-3-phosphate dehydrogenase (Gap), methionine synthase (MetE) and other amino acid biosynthesis enzymes, organic hydroperoxide resistance transcriptional repressor (OhrR), a predicted 2-Cys Prx (YkuU), translation factors, and chaperones [18,167,168]. The three Brx proteins identified in B. subtilis and S. aureus are named BrxA (previously YphP), BrxB (YqiW), and BrxC (YtxJ). BrxA and BrxB are paralogs and possess a redox-active CGC motif, whereas BrxC has only one Cys in a TCPIS motif. Debacillithiolation of Gap, MetE, and OhrR by BrxA and/or BrxB has been demonstrated [168,169]. Recently, BrxC was shown to debacillithiolate Gap, BrxB and Bdr, while BrxB was not debacillithiolated by BSH excess [165]. Similar to TrxR, Bdr is a FAD-containing NADPH-dependent oxidoreductase. While Bacillus cereus Bdr has a single-Cys residue, the Bdr proteins from B. subtilis and S. aureus have three-Cys residues, including one at position 14 shared with B. cereus Bdr (Figure S15). The characterisation of a S. aureus Bdr variant, in which Cys14 was replaced by Ala, suggested that Cys14 is the redox-active Cys as it seems required for BSSB reductase activity [163]. BSSB reductase activity was also demonstrated for B. cereus Bdr [164]. For B. subtilis Bdr, which could debacillithiolate BrxB-SSB and less efficiently BrxC–SSB, mutation of any of the three Cys residues led to decreased capacity to reduce both mixed disulfides [165]. A variant mutated for all Cys was essentially inactive. Of note, analysis of the crystal structures of Bdr from S. aureus and B. cereus indicated that Cys14 does not directly participate in the BSSB reductase reaction mechanism [164], and the same may be true for the Brx reductase activity [165]. In summary, the precise roles of these Cys in the BSSB or Brx reductase activities of Bdr remain to be delineated.
BSH is expected to be common to all members of the genus Deinococcus because the BSH biosynthesis genes bshA, bshB, and bshC are present in all species [3,27]. However, the other components of the BSH system, in particular Brx, appear to differ from their counterparts in Bacillus and Staphylococcus. Only a BrxC-related protein is present in Deinococcus species. This protein, containing a single Cys, is highly conserved (e.g., D. radiodurans DR_1832 has 71% to 82% identity with BrxC from the other analysed species), but displays low levels of similarity with BrxC from Bacillus and Staphylococcus (Figure S16). Instead of the TCPIS motif found in the latter, the corresponding sequence in deinococcal proteins is TCHKT. Moreover, Deinococcus BrxC proteins are substantially longer (204 to 219 residues instead of 108 residues for B. subtilis BrxC). The Trx-like domain in Deinococcus BrxC is located in the N-terminal part of the protein. The C-terminal region has some similarity with the bacterial self-protective colicin-like immunity domain (pfam09204). The function of this domain in Deinococcus BrxC is unknown. Because of the differences with BrxC from Bacillus and Staphylococcus, DR_1832 has been named AbxC (atypical BrxC) [27]. In D. radiodurans, expression of the abxC gene (DR_1832) is induced following exposure to cadmium [52]. A Bdr homolog is also present in all Deinococcus. D. radiodurans Bdr (DR_2623) has 74% to 89% identity with the homologs of the other six species and 40% with B. subtilis Bdr. DR_2623 contains three Cys residues (Cys22, Cys36, Cys275) that are conserved in deinococcal Bdr proteins, except for Deipe_2475, which lacks the first Cys corresponding to Cys22 of DR_2623 (Figure S15). Whether and how the Cys residues, located at different positions compared with those in Bdrs from Bacillus and Staphylococcus, support the catalytic mechanism of deinococcal Bdr is currently unknown. Upregulated expression of DR_2623 (Bdr) has been observed after exposure of D. radiodurans to cadmium [52], gamma radiation [158,170], and desiccation [171].
Experimental data obtained with D. radiodurans indicated that AbxC (BrxC) contributes to oxidative stress resistance [27]. In vitro, incubation of AbxC with H2O2 resulted in the formation of AbxC dimers through an intermolecular disulfide bond. NADPH consumption was observed when oxidised AbxC was incubated with BSH, NADPH, and Bdr (DR_2623), indicating reduction of the AbxC dimer by BSH and reduction of the formed BSSB by Bdr [27]. S-bacillithiolated proteins in Deinococcus have not been identified yet, and it is currently unknown whether the atypical AbxC can debacillithiolate proteins.
6. Conclusions
The remarkable tolerance of Deinococcus bacteria to extreme environmental conditions relies on a wide array of mechanisms ensuring efficient repair of DNA and limiting damage in proteins, many being present in all species. However, some genes, such as those coding for specific catalase types that confer resistance to oxidative stress, are not present in all species of the genus, revealing diversity in these mechanisms [3]. Regarding thiol reductases, most representatives are found in all Deinococcus species analysed here (Table 2), except Trx2, indicating that they are likely essential actors in the stress responses of this genus, as it is now well established in many other organisms. Consistently, deletion mutants for trx, frnE, bcp, osmC, ahpD-like, and abxC genes show increased susceptibility to oxidative treatments [25,27,60,77,146,160]. Nonetheless, the knowledge about the functions of Deinococcus thiol reductases remains limited. Thorough investigations need to be performed to delineate the precise physiological roles of these enzymes, notably by characterising mutants impaired in their expression in conditions of extreme stress, and by identifying their partners and substrates.
In addition to the NADPH-dependent Trx-and BSH-systems, Deinococcus possesses various types of thiol peroxidases using NADH as an electron source. Of note, Yamashiro et al. [172] reported a much greater concentration of NADH compared with that of NADPH in this genus, and a much higher NADH/NADPH ratio compared with E. coli, indicating that NADH is likely an essential actor in metabolism and stress responses in Deinococcus. Most interestingly, this genus displays a large diversity in thiol peroxidase types. On one hand, Trx-dependent BCPs are well represented in all Deinococcus species, since each possesses two or three genes coding for these Prxs, instead of one in E. coli. In addition, all Deinococcus species have two distinct types of NADH-dependent thiol peroxidases (i.e., AhpE/AhpD and Ohr/OsmC). The gene copy number and diversity of thiol peroxidases in bacteria, notably regarding the specialisation of their active sites, are likely associated with their level of stress tolerance [118,157]. Thus, AhpC, Ohr, and OsmC are widely distributed among bacterial species, but only some species simultaneously possess all of them, the others having generally one or two genes. For instance, Ohr is absent in M. tuberculosis and E. coli. The mycobacterium C. glutamicum, which is highly resistant to oxidative stress, possesses AhpD, Ohr, and OsmC homologs [143,157]. Regarding BCPs, Limauro et al. [173] identified three genes in the hyperthermophilic archaeon Sulfolobus solfataricus and proposed that this copy number was associated with the tolerance of this archaea to extreme temperature. The Deinococcus genus appears thus to be remarkably well equipped with a diversity of thiol peroxidases, since most species possess one or several bcp, ahpD, ahpE, osmC, ohr, and yhfA genes. This palette is very likely an asset in the extreme resistance of Deinococcus via efficient scavenging of all types of peroxides and peroxynitrites. Indeed, these thiol peroxidases display in vitro different specificity towards substrates [144,154,155,174]. For instance, OsmC is more efficient in metabolising organic peroxides than H2O2 [153]. In vivo investigations in P. aeruginosa showed increased susceptibility to artificial and fatty acid hydroperoxides of strains lacking Ohr, this phenotype being not restored by the expression of other types of thiol peroxidases, such as AhpC [152]. Taken collectively, these data strongly suggest that the diversity of thiol peroxidases in Deinococcus is associated with specialisation and efficiency regarding the scavenging of the various peroxide types produced upon stress conditions, and thus contributes to the outstanding tolerance of this genus.
The various electron transfer pathways involving thiol reductases present in Deinococcus are shown in Figure 5. Some pathways need to be precisely delineated, notably regarding the substrates of Trx and Brx proteins, which remain to be identified using biochemical and proteomics approaches. Based on the knowledge gained in other organisms, connections and interplays between these pathways very likely occur in Deinococcus. Indeed, in E. coli, there is functional redundancy between Trx and GSH systems [175], and an interplay between the two systems has been unveiled in plants during development [176]. Consistently, by generating B. subtilis strains impaired in BSH biosynthesis, Gaballa et al. [177] showed that this LMW thiol was not required to maintain the reduced status of protein thiols in the absence of stress, but noticed altered stress sensitivity of these strains. Regarding Deinococcus, it was recently reported that D. deserti and D. radiodurans mutant strains deficient in BSH biosynthesis are viable [27,59], indicating that in the absence of stress conditions the BSH system is not essential and likely compensated by the Trx system. Increased sensitivity to H2O2 has been reported for the D. radiodurans BSH-deficient mutant [27]. The occurrence of such compensation mechanisms and interplays is corroborated by the presence of Trx substrates or of other thiol reductases in bacillithiolated or Brx-interacting proteins. Among the bacillithiolated proteins identified in Bacillus and Staphylococcus, glyceraldehyde-3-phosphate dehydrogenase, a well-known Trx substrate [18,54,165,168], and a predicted 2-Cys Prx (YkuU) [167] have been found. The BSH system in Deinococcus is likely to allow the maintenance of the redox status and/or the regeneration of the activity of Trx-dependent enzymes, as shown for GSH in other bacteria [13]. In Deinococcus, Bihani et al. [78] proposed that in conditions of cadmium stress, which inhibits TrxR, Trx substrates could be supplied with electrons thanks to BSH. In D. radiodurans, 25 proteins potentially interacting with AbxC were isolated after incubation of cell extracts with AbxC-immobilised beads [27]. Among them, Ohr was identified, revealing interplays between the BSH/Brx system and NADH-dependent thiol peroxidases. BSH might also participate in the reduction of AhpE by analogy with the role of mycothiol. Indeed, in M. tuberculosis, reduction of AhpE can be performed via mycoredoxin- or mycothiol-dependent mechanisms [145]. Concerning lipoyl-dependent thiol peroxidases, a link has been unveiled in C. glutamicum between the AhpD system and Ohr/OsmC peroxidases, which in this species can be reduced by AhpD [157]. Taking into consideration all these data, we can thus hypothesise that the various pathways for electron transfer via thiol reductases and peroxidases in Deinococcus give rise to multiple connections and possible redundancy among them. This very likely allows the establishment of protective and repair mechanisms that can substitute or be switched on differentially depending on stress conditions, leading to the implementation of a complex and robust network (Figure 5) maintaining protein redox homeostasis in the extreme conditions that this bacterial genus can tolerate.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/antiox11030561/s1: Figure S1: Multiple sequence alignment of Deinococcus, E. coli, and B. subtilis NADPH thioredoxin reductases. Abbreviations of Deinococcus species names (i.e., the first part of the locus tags (gene numbers)) are as specified in Table 1. TRXB_ECOLI and TRXB_BACSU, thioredoxin reductases from E. coli and B. subtilis, respectively. The alignment was made with ClustalW at NPS@ [178] and ESPript [179]. Figure S2: Multiple sequence alignment of Deinococcus, E. coli, and B. subtilis thioredoxins. (a) Alignment of Trx1 sequences. (b) Alignment of Trx2 sequences. Abbreviations of Deinococcus species names are as specified in Table 1. THIO_ECOLI and THIO_BACSU, thioredoxin 1 from E. coli and B. subtilis, respectively. THIO2_ECOLI, thioredoxin 2 from E. coli. Alignment was made as in Figure S1. Figure S3: Gene clusters with additional trxR, trxA, and ahpD genes in D. peraridilitoris. The two clusters shown are both located on plasmid pDEIPE01. Locus tags (Deipe_3903 etc.) are indicated. Gene sizes not drawn to scale. Figure S4: Three-dimensional models of Trx and Trx-like proteins of D. radiodurans. Models in panels (a) (Trx1) and (c) to (g) (Trx-like) were obtained with AlphaFold2_Advanced [180]. (b) Crystal structure of Trx2 (PDB 7DL6) showing the extra N-terminal zinc binding domain in dark blue [60]. Catalytic and resolving cysteine residues are shown in green and blue, respectively. Three-dimensional structure or model images were generated using PyMOL (PyMOL Molecular Graphics System, Version 2.0, Schrödinger, LLC). Figure S5: Sequence alignments of Trx-like proteins from Deinococcus. (a) Alignment of Deinococcus DR_0057-type sequences. (b) Alignment of Deinococcus DR_A0072-type sequences. (c) Alignment of Deinococcus DR_B0010-type sequences. (d) Alignment of Deinococcus DR_0948-type sequences. (e) Tree showing sequence relationship between Trx-like proteins shown in Figure 2 and in panels (a–d). The guide tree is calculated based on the distance matrix that is generated from the pairwise scores. Trx active site sequences are highlighted in yellow; residues highlighted in cyan in Deipr sequences differ in the proximity of the active site. Abbreviations of Deinococcus species names are as specified in Table 1. Alignments and tree were obtained with UniProt ClustalO (https://www.uniprot.org/align/) [181]. Figure S6: Multiple sequence alignment of Deinococcus FrnE reductases. (a) FrnE proteins (DsbA family, FrnE subfamily). The N-terminal CxxC corresponds to residues 22–25 in D. radiodurans FrnE (DR_0659), and the C-terminal CxxxxC to residues 239–244. (b) FrnE-like proteins found in two Deinococcus species (DsbA family, FrnE-like subfamily). The CxxC motif is highlighted in yellow. (c) Proteins containing a DSBA-like Trx domain. The single Cys is highlighted in yellow. Alignments were made as in Figures S1 and S5. Figure S7: Putative operons encoding DsbA, DsbB, and UvrA. Similar to the B. subtilis bdbDC operon encoding DsbA and DsbB homologs, putative operons encoding DsbA and DsbB family proteins are present in most Deinococcus species. In five of the seven analysed species, these genes are likely in operon with genes encoding DNA repair protein UvrA and a membrane protein designated dCSP-1 (Deinococcus-specific conserved signature protein-1). In D. radiodurans (DR), the uvrA-dCSP-1 genes are located separate from the bdbDC genes. D. peraridilitoris (Deipe) lacks the bdbDC homologs. Gene numbers (locus tags) for D. radiodurans, D. deserti, and D. peraridilitoris are indicated. Figure S8: Multiple sequence alignments of Deinococcus Dsb oxidoreductases. (a) DsbA family proteins with an N-terminal transmembrane helix from Deinococcus species aligned with B. subtilis BdbD. Except for the second homolog of D. proteolyticus (Deipr_2421), the genes are adjacent to a gene encoding the DsbB/BdbC homologs shown in panel (b) (see also Figure S7). Residues highlighted in cyan in Deipr_2421 differ in the proximity of the active site. (c) DsbA family proteins with N-terminal signal peptide. (d) Homologs of the DsbD family protein CcdA from Deinococcus aligned with B. subtilis CcdA. The deinococcal ccdA homologs are in gene clusters also encoding DsbE/CcmG homologs (see Figure S9). (e) DsbE/CcmG homologs from Deinococcus encoded by gene clusters also encoding CcdA and by isolated genes aligned with the TlpA-like family proteins ResA and YneN from B. subtilis and DsbE/CcmG from E. coli. Cys residues and CxxC motifs are highlighted in yellow. Guide trees showing sequence relationship between various proteins are included in case more than one homolog is present in the same Deinococcus species. Alignments and trees were obtained with UniProt ClustalO. Figure S9: Gene clusters encoding CcdA and DsbE/CcmG homologs. (a) The seven analysed Deinococcus species possess ccdA (DsbD family) and dsbE/ccmG homologs in gene clusters containing other cytochrome c biogenesis genes. Different genetic organisations are present in the different species, as shown. The ccm cluster from E. coli is also shown for comparison. Gene numbers (locus tags) for the indicated Deinococcus species are indicated. (b) Additional two-gene cluster encoding CcdA and DsbE/CcmG found in two Deinococcus species. Figure S10: Multiple sequence alignment of Deinococcus Msr proteins. (a) Alignment of Deinococcus, E. coli, and B. subtilis MsrA sequences. MSRA_ECOLI and MSRA_BACSU, MsrAs from E. coli and B. subtilis, respectively. (b) Alignment of Deinococcus, E. coli, and B. subtilis MsrB sequences. MSRB_ECOLI and MSRB_BACSU, MsrBs from E. coli and B. subtilis, respectively. Catalytic and resolving cysteines are highlighted in green and cyan, respectively. Extra cysteines present in some sequences are highlighted in yellow. The black lines indicate the consensus sequences proximal to catalytic cysteines and prevalent in MsrA and MsrB. Figure S11. Multiple sequence alignments of BCP peroxiredoxins from Deinococcus. (a) Alignment of Deinococcus DR_0846-type, B. subtilis, and E. coli BCP sequences. (b) Alignment of other Deinococcus BCP sequences. (c) Tree showing sequence relationship between BCP proteins shown in panels (a,b) and two more distant proteins from D. gobiensis and D. deserti. BCP, bacterioferritin comigratory protein. BCP_ECOLI and BCP_BACSU, BCP peroxiredoxins from E. coli and B. subtilis, respectively. Peroxidatic and resolving cysteines are highlighted in green and cyan, respectively. The black line indicates the sequence proximal to peroxidatic cysteine prevalent in BCPs, PKxxTPGCTxEAC [118]. The two other residues of the catalytic triad are highlighted in grey. Residues highlighted in yellow are highly conserved in bacterial BCPs [128]. Extra cysteines present in some sequences are highlighted in pink. Alignments and tree were obtained with UniProt ClustalO as in Figure S5. Figure S12: Multiple sequence alignment of Deinococcus AhpE and AhpD proteins. (a) Alignment of Deinococcus and M. tuberculosis AhpE sequences. AHPE_MYCTU, AhpE from M. tuberculosis. Peroxidatic cysteine (green) and other catalytic triad residues (grey) are highlighted. (b) Alignment of Deinococcus and M. tuberculosis AhpD sequences. AHPD_MYCTU, AhpD from M. tuberculosis. Catalytic and resolving cysteines are highlighted in green and cyan, respectively. The residues highlighted in grey in AhpD sequences are involved in the catalytic mechanism. The guide tree showing sequence relationship between the various AhpD-like proteins is included. Alignments and tree were obtained with UniProt ClustalO. Figure S13: Three-dimensional models of BCPs and OsmC of D. radiodurans. (a) The three BCPs having a Trx-fold. (b) Model of OsmC and crystal structure of Ohr (PDB 1USP). Peroxidatic and resolving cysteine residues are shown in green and blue, respectively. The OsmC/Ohr monomer is composed of two distinct subdomains. The N-terminal domain (orange) consists of three β-strands folded into a β-sheet. In the C-terminal domain, helices α1 and α2 (the latter severely kinked) are stacked on a β-sheet formed by strands β4 –β6. The active OsmC and Ohr protein is formed by a homodimer [159]. Models and images were obtained as described in Figure S4. Figure S14: Multiple sequence alignment of Deinococcus OsmC, Ohr, and YhfA proteins. (a) Alignment of Deinococcus and E. coli OsmC sequences. OSMC_ECOLI, OsmC from E. coli. (b) Alignment of Deinococcus and B. subtilis Ohr. OHRA_BACSU and OHRB_BACSU, OhrA and OhrB proteins, respectively, from B. subtilis. (c) Alignment of Deinococcus and E. coli YhfA sequences. YHFA_ECOLI, YhfA from E. coli. Peroxidatic and resolving cysteines are highlighted in green and cyan, respectively. The two residues highlighted in grey in Ohr and OsmC sequences are involved in the catalytic mechanism [154]. Figure S15: Multiple sequence alignment of Deinococcus bacillithiol reductases. Bdr homologs from Deinococcus are aligned with Bdr proteins YPDA_BACSU from B. subtilis, BC_1495 from B. cereus, and SACOL1520 from S. aureus. Cysteine residues are highlighted in yellow. Figure S16: Multiple sequence alignment of Deinococcus bacilliredoxins. The BrxC-type bacilliredoxins (also called AbxC for atypical BrxC) from Deinococcus are aligned with BrxC proteins YtxJ_BACCE from B. cereus, YTXJ_BACSU from B. subtilis, and SACOL0804 from S. aureus. Active site cysteine is highlighted in yellow.
Author Contributions
Database search and sequence analysis: A.d.G. Review of literature and writing—original draft preparation: A.d.G. and P.R. Writing—review and editing: A.d.G., L.B., N.R. and P.R. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by Transverse Division No. 4 (Radiobiology) of the French Alternative Energies and Atomic Energy Commission (Programme No. 4 Radiobiologie), a grant from the Agence Nationale de la Recherche (ANR-19-CE12-0010), and by our institute (BIAM) for a transverse project.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in review.
Acknowledgments
We are grateful to Christophe Carles and David Pignol for their constant support and interest.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Battista, J.R. Against All Odds: The Survival Strategies of Deinococcus radiodurans. Annu. Rev. MicroBiol. 1997, 51, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Slade, D.; Radman, M. Oxidative Stress Resistance in Deinococcus radiodurans. MicroBiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.; Jung, J.-H.; Blanchard, L.; de Groot, A. Conservation and Diversity of Radiation and Oxidative Stress Resistance Mechanisms in Deinococcus Species. FEMS MicroBiol. Rev. 2019, 43, 19–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattimore, V.; Battista, J.R. Radioresistance of Deinococcus radiodurans: Functions Necessary to Survive Ionizing Radiation Are Also Necessary to Survive Prolonged Desiccation. J. Bacteriol. 1996, 178, 633–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Groot, A.; Dulermo, R.; Ortet, P.; Blanchard, L.; Guerin, P.; Fernandez, B.; Vacherie, B.; Dossat, C.; Jolivet, E.; Siguier, P.; et al. Alliance of Proteomics and Genomics to Unravel the Specificities of Sahara Bacterium Deinococcus deserti. PLoS Genet. 2009, 5, e1000434. [Google Scholar] [CrossRef]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Leapman, R.D.; Lai, B.; Ravel, B.; Li, S.M.; Kemner, K.M.; et al. Protein Oxidation Implicated as the Primary Determinant of Bacterial Radioresistance. PLoS Biol. 2007, 5, e92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredrickson, J.K.; Li, S.M.; Gaidamakova, E.K.; Matrosova, V.Y.; Zhai, M.; Sulloway, H.M.; Scholten, J.C.; Brown, M.G.; Balkwill, D.L.; Daly, M.J. Protein Oxidation: Key to Bacterial Desiccation Resistance? ISME J. 2008, 2, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krisko, A.; Radman, M. Protein Damage and Death by Radiation in Escherichia coli and Deinococcus radiodurans. Proc. Natl. Acad. Sci. USA 2010, 107, 14373–14377. [Google Scholar] [CrossRef] [Green Version]
- Daly, M.J. Death by Protein Damage in Irradiated Cells. DNA Repair 2012, 11, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Holmgren, A. The Thioredoxin Antioxidant System. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Arnér, E.S.; Holmgren, A. Physiological Functions of Thioredoxin and Thioredoxin Reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Carmel-Harel, O.; Storz, G. Roles of the Glutathione- and Thioredoxin-Dependent Reduction Systems in the Escherichia coli and Saccharomyces cerevisiae Responses to Oxidative Stress. Annu. Rev. MicroBiol. 2000, 54, 439–461. [Google Scholar] [CrossRef] [PubMed]
- Toledano, M.B.; Kumar, C.; Le Moan, N.; Spector, D.; Tacnet, F. The System Biology of Thiol Redox. System in Escherichia coli and Yeast: Differential Functions in Oxidative Stress, Iron Metabolism and DNA Synthesis. FEBS Lett. 2007, 581, 3598–3607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative Stress, Protein Damage and Repair in Bacteria. Nat. Rev. MicroBiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.P. Thioredoxins and Glutaredoxins: Unifying Elements in Redox. Biology. Annu. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef] [PubMed]
- Vieira Dos Santos, C.; Rey, P. Plant Thioredoxins Are Key Actors in the Oxidative Stress Response. Trends Plant. Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.-P.; Fleury, M.J.J. Microbial Antioxidant Defense Enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef]
- Chandrangsu, P.; Loi, V.V.; Antelmann, H.; Helmann, J.D. The Role of Bacillithiol in Gram-Positive Firmicutes. Antioxid. Redox. Signal. 2018, 28, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Imber, M.; Pietrzyk-Brzezinska, A.J.; Antelmann, H. Redox. Regulation by Reversible Protein S-Thiolation in Gram-Positive Bacteria. Redox. Biol. 2019, 20, 130–145. [Google Scholar] [CrossRef]
- Reyes, A.M.; Pedre, B.; De Armas, M.I.; Tossounian, M.-A.; Radi, R.; Messens, J.; Trujillo, M. Chemistry and Redox. Biology of Mycothiol. Antioxid. Redox. Signal. 2018, 28, 487–504. [Google Scholar] [CrossRef]
- Choo, K.; Kim, M.; Nansa, S.A.; Bae, M.K.; Lee, C.; Lee, S.-J. Redox. Potential Change by the Cystine Importer Affected on Enzymatic Antioxidant Protection in Deinococcus geothermalis. Antonie Van Leeuwenhoek 2020, 113, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Obiero, J.; Pittet, V.; Bonderoff, S.A.; Sanders, D.A. Thioredoxin System from Deinococcus radiodurans. J. Bacteriol. 2010, 192, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obiero, J.; Sanders, D.A.R. Design of Deinococcus radiodurans Thioredoxin Reductase with Altered Thioredoxin Specificity Using Computational Alanine Mutagenesis. Protein Sci. 2011, 20, 1021–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Tang, Y.; Hua, Y.; Zhao, Y. Structural Features and Functional Implications of Proteins Enabling the Robustness of Deinococcus radiodurans. Comput. Struct. Biotechnol. J. 2020, 18, 2810–2817. [Google Scholar] [CrossRef]
- Cho, C.; Lee, G.W.; Hong, S.H.; Kaur, S.; Jung, K.-W.; Jung, J.-H.; Lim, S.; Chung, B.Y.; Lee, S.S. Novel Functions of Peroxiredoxin Q from Deinococcus radiodurans R1 as a Peroxidase and a Molecular Chaperone. FEBS Lett. 2019, 593, 219–229. [Google Scholar] [CrossRef]
- Maqbool, I.; Ponniresan, V.K.; Govindasamy, K.; Prasad, N.R. Understanding the Survival Mechanisms of Deinococcus radiodurans against Oxidative Stress by Targeting Thioredoxin Reductase Redox. System. Arch. MicroBiol. 2020, 202, 2355–2366. [Google Scholar] [CrossRef]
- Jeong, S.; Jung, J.-H.; Kim, M.-K.; de Groot, A.; Blanchard, L.; Ryu, S.; Bahn, Y.-S.; Lim, S. Atypical Bacilliredoxin AbxC Plays a Role in Responding to Oxidative Stress in Radiation-Resistant Bacterium Deinococcus radiodurans. Antioxidants 2021, 10, 1148. [Google Scholar] [CrossRef]
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: An Integrated Platform for the Annotation and Exploration of Microbial Gene Functions through Genomic, Pangenomic and Metabolic Comparative Analysis. Nucleic Acids Res. 2020, 48, D579–D589. [Google Scholar] [CrossRef] [Green Version]
- Lipton, M.S.; Pasa-Tolic, L.; Anderson, G.A.; Anderson, D.J.; Auberry, D.L.; Battista, J.R.; Daly, M.J.; Fredrickson, J.; Hixson, K.K.; Kostandarithes, H.; et al. Global Analysis of the Deinococcus radiodurans Proteome by Using Accurate Mass Tags. Proc. Natl. Acad. Sci. USA 2002, 99, 11049–11054. [Google Scholar] [CrossRef] [Green Version]
- Schmid, A.K.; Lipton, M.S.; Mottaz, H.; Monroe, M.E.; Smith, R.D.; Lidstrom, M.E. Global Whole-Cell FTICR Mass Spectrometric Proteomics Analysis of the Heat Shock Response in the Radioresistant Bacterium Deinococcus radiodurans. J. Proteome Res. 2005, 4, 709–718. [Google Scholar] [CrossRef]
- de Groot, A.; Roche, D.; Fernandez, B.; Ludanyi, M.; Cruveiller, S.; Pignol, D.; Vallenet, D.; Armengaud, J.; Blanchard, L. RNA Sequencing and Proteogenomics Reveal the Importance of Leaderless MRNAs in the Radiation-Tolerant Bacterium Deinococcus deserti. Genome Biol. Evol. 2014, 6, 932–948. [Google Scholar] [CrossRef] [PubMed]
- Ott, E.; Kawaguchi, Y.; Kölbl, D.; Chaturvedi, P.; Nakagawa, K.; Yamagishi, A.; Weckwerth, W.; Milojevic, T. Proteometabolomic Response of Deinococcus radiodurans Exposed to UVC and Vacuum Conditions: Initial Studies Prior to the Tanpopo Space Mission. PLoS ONE 2017, 12, e0189381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, E.; Fuchs, F.M.; Moeller, R.; Hemmersbach, R.; Kawaguchi, Y.; Yamagishi, A.; Weckwerth, W.; Milojevic, T. Molecular Response of Deinococcus radiodurans to Simulated Microgravity Explored by Proteometabolomic Approach. Sci. Rep. 2019, 9, 18462. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.W.; Nordan, H.C.; Cain, R.F.; Parrish, G.; Duggan, D. Studies on a Radio-Resistant Micrococcus. I. Isolation, Morphology, Cultural Characteristics, and Resistance to Gamma Radiation. Food Technol. 1956, 10, 575–578. [Google Scholar]
- Brooks, B.W.; Murray, R.G.E. Nomenclature for “Micrococcus radiodurans” and Other Radiation-Resistant Cocci: Deinococcaceae Fam. Nov. and Deinococcus Gen. Nov., Including Five Species. Int. J. Syst. Bacteriol. 1981, 31, 353–360. [Google Scholar] [CrossRef] [Green Version]
- White, O.; Eisen, J.A.; Heidelberg, J.F.; Hickey, E.K.; Peterson, J.D.; Dodson, R.J.; Haft, D.H.; Gwinn, M.L.; Nelson, W.C.; Richardson, D.L.; et al. Genome Sequence of the Radioresistant Bacterium Deinococcus radiodurans R1. Science 1999, 286, 1571–1577. [Google Scholar] [CrossRef] [Green Version]
- de Groot, A.; Chapon, V.; Servant, P.; Christen, R.; Fischer-Le Saux, M.; Sommer, S.; Heulin, T. Deinococcus deserti Sp. Nov., a Gamma-Radiation-Tolerant Bacterium Isolated from the Sahara Desert. Int. J. Syst. Evol. MicroBiol. 2005, 55, 2441–2446. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.C.; Nobre, M.F.; Rainey, F.A.; Silva, M.T.; Wait, R.; Burghardt, J.; Chung, A.P.; da Costa, M.S. Deinococcus geothermalis Sp. Nov. and Deinococcus murrayi Sp. Nov., Two Extremely Radiation-Resistant and Slightly Thermophilic Species from Hot Springs. Int. J. Syst. Bacteriol. 1997, 47, 939–947. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Lapidus, A.; Copeland, A.; Kim, E.; Land, M.; et al. Deinococcus geothermalis: The Pool of Extreme Radiation Resistance Genes Shrinks. PLoS ONE 2007, 2, e955. [Google Scholar] [CrossRef]
- Yuan, M.; Zhang, W.; Dai, S.; Wu, J.; Wang, Y.; Tao, T.; Chen, M.; Lin, M. Deinococcus gobiensis Sp. Nov., an Extremely Radiation-Resistant Bacterium. Int. J. Syst. Evol. MicroBiol. 2009, 59, 1513–1517. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Chen, M.; Zhang, W.; Lu, W.; Wang, J.; Yang, M.; Zhao, P.; Tang, R.; Li, X.; Hao, Y.; et al. Genome Sequence and Transcriptome Analysis of the Radioresistant Bacterium Deinococcus gobiensis: Insights into the Extreme Environmental Adaptations. PLoS ONE 2012, 7, e34458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainey, F.A.; Ray, K.; Ferreira, M.; Gatz, B.Z.; Nobre, M.F.; Bagaley, D.; Rash, B.A.; Park, M.J.; Earl, A.M.; Shank, N.C.; et al. Extensive Diversity of Ionizing-Radiation-Resistant Bacteria Recovered from Sonoran Desert Soil and Description of Nine New Species of the Genus Deinococcus Obtained from a Single Soil Sample. Appl. Environ. MicroBiol. 2005, 71, 5225–5235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pukall, R.; Zeytun, A.; Lucas, S.; Lapidus, A.; Hammon, N.; Deshpande, S.; Nolan, M.; Cheng, J.F.; Pitluck, S.; Liolios, K.; et al. Complete Genome Sequence of Deinococcus maricopensis Type Strain (LB-34). Stand. Genomic Sci. 2011, 4, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Rainey, F.A.; Ferreira, M.; Nobre, M.F.; Ray, K.; Bagaley, D.; Earl, A.M.; Battista, J.R.; Gomez-Silva, B.; McKay, C.P.; da Costa, M.S. Deinococcus peraridilitoris Sp. Nov., Isolated from a Coastal Desert. Int. J. Syst. Evol. MicroBiol. 2007, 57, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Kobatake, M.; Tanabe, S.; Hasegawa, S. New Micrococcus Radioresistant Red Pigment, Isolated from Lama glama Feces, and Its Use as Microbiological Indicator of Radiosterilization. C R Seances Soc. Biol. Fil. 1973, 167, 1506–1510. [Google Scholar] [PubMed]
- Copeland, A.; Zeytun, A.; Yassawong, M.; Nolan, M.; Lucas, S.; Hammon, N.; Deshpande, S.; Cheng, J.F.; Han, C.; Tapia, R.; et al. Complete Genome Sequence of the Orange-Red Pigmented, Radioresistant Deinococcus proteolyticus Type Strain (MRP(T)). Stand. Genomic Sci. 2012, 6, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Laurent, T.C.; Moore, E.C.; Reichard, P. Enzymatic Synthesis Of Deoxyribonucleotides. Iv. Isolation And Characterization of Thioredoxin, The Hydrogen Donor From Escherichia coli B. J. Biol. Chem. 1964, 239, 3436–3444. [Google Scholar] [CrossRef]
- Ritz, D.; Patel, H.; Doan, B.; Zheng, M.; Aslund, F.; Storz, G.; Beckwith, J. Thioredoxin 2 Is Involved in the Oxidative Stress Response in Escherichia coli. J. Biol. Chem. 2000, 275, 2505–2512. [Google Scholar] [CrossRef] [Green Version]
- Argyrou, A.; Blanchard, J.S. Flavoprotein Disulfide Reductases: Advances in Chemistry and Function. Prog. Nucleic Acid Res. Mol. Biol. 2004, 78, 89–142. [Google Scholar] [CrossRef]
- Hammerstad, M.; Hersleth, H.-P. Overview of Structurally Homologous Flavoprotein Oxidoreductases Containing the Low Mr Thioredoxin Reductase-like Fold—A Functionally Diverse Group. Arch. BioChem. Biophys. 2021, 702, 108826. [Google Scholar] [CrossRef] [PubMed]
- Lennon, B.W.; Williams, C.H.; Ludwig, M.L. Twists in Catalysis: Alternating Conformations of Escherichia coli Thioredoxin Reductase. Science 2000, 289, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Joe, M.-H.; Jung, S.-W.; Im, S.-H.; Lim, S.-Y.; Song, H.-P.; Kwon, O.; Kim, D.-H. Genome-Wide Response of Deinococcus radiodurans on Cadmium Toxicity. J. MicroBiol. Biotechnol. 2011, 21, 438–447. [Google Scholar] [CrossRef]
- Zeller, T.; Klug, G. Thioredoxins in Bacteria: Functions in Oxidative Stress Response and Regulation of Thioredoxin Genes. Naturwissenschaften 2006, 93, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.S.; Vertommen, D.; Baldin, F.; Laloux, G.; Collet, J.-F. Comprehensively Characterizing the Thioredoxin Interactome in Vivo Highlights the Central Role Played by This Ubiquitous Oxidoreductase in Redox. Control. Mol. Cell Proteomics 2016, 15, 2125–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chasapis, C.T.; Makridakis, M.; Damdimopoulos, A.E.; Zoidakis, J.; Lygirou, V.; Mavroidis, M.; Vlahou, A.; Miranda-Vizuete, A.; Spyrou, G.; Vlamis-Gardikas, A. Implications of the Mitochondrial Interactome of Mammalian Thioredoxin 2 for Normal Cellular Function and Disease. Free Radic. Biol. Med. 2019, 137, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, M.E.; Mauriès, A.; Maes, A.; Tourasse, N.J.; Hamon, M.; Lemaire, S.D.; Marchand, C.H. The Deep Thioredoxome in Chlamydomonas reinhardtii: New Insights into Redox. Regulation. Mol. Plant. 2017, 10, 1107–1125. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Vizuete, A.; Damdimopoulos, A.E.; Gustafsson, J.; Spyrou, G. Cloning, Expression, and Characterization of a Novel Escherichia coli Thioredoxin. J. Biol. Chem. 1997, 272, 30841–30847. [Google Scholar] [CrossRef] [Green Version]
- Collet, J.F.; D’Souza, J.C.; Jakob, U.; Bardwell, J.C. Thioredoxin 2, an Oxidative Stress-Induced Protein, Contains a High Affinity Zinc Binding Site. J. Biol. Chem. 2003, 278, 45325–45332. [Google Scholar] [CrossRef] [Green Version]
- Magerand, R.; Rey, P.; Blanchard, L.; de Groot, A. Redox. Signaling through Zinc Activates the Radiation Response in Deinococcus Bacteria. Sci. Rep. 2021, 11, 4528. [Google Scholar] [CrossRef]
- Kim, M.-K.; Zhao, L.; Jeong, S.; Zhang, J.; Jung, J.-H.; Seo, H.S.; Choi, J.-I.; Lim, S. Structural and Biochemical Characterization of Thioredoxin-2 from Deinococcus radiodurans. Antioxidants 2021, 10, 1843. [Google Scholar] [CrossRef]
- Rabinovitch, I.; Yanku, M.; Yeheskel, A.; Cohen, G.; Borovok, I.; Aharonowitz, Y. Staphylococcus aureus NrdH Redoxin Is a Reductant of the Class Ib Ribonucleotide Reductase. J. Bacteriol. 2010, 192, 4963–4972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibani, K.; Wingsle, G.; Jacquot, J.-P.; Gelhaye, E.; Rouhier, N. Comparative Genomic Study of the Thioredoxin Family in Photosynthetic Organisms with Emphasis on Populus trichocarpa. Mol. Plant. 2009, 2, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Yoon, H.W.; Lee, S.-R.; Rhee, S.G. Identification and Characterization of TRP14, a Thioredoxin-Related Protein of 14 KDa. New Insights into the Specificity of Thioredoxin Function. J. Biol. Chem. 2004, 279, 3142–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, E.; Peluso, I.; Rendina, R.; Digilio, A.; Furia, M. The Clot Gene of Drosophila melanogaster Encodes a Conserved Member of the Thioredoxin-like Protein Superfamily. Mol. Genet. Genomics 2003, 268, 692–697. [Google Scholar] [CrossRef]
- Chibani, K.; Tarrago, L.; Gualberto, J.M.; Wingsle, G.; Rey, P.; Jacquot, J.-P.; Rouhier, N. Atypical Thioredoxins in Poplar: The Glutathione-Dependent Thioredoxin-like 2.1 Supports the Activity of Target Enzymes Possessing a Single Redox. Active Cysteine. Plant. Physiol. 2012, 159, 592–605. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, B.; Arnér, E.S.J. Thioredoxin-Related Protein of 14 KDa as a Modulator of Redox. Signalling Pathways. Br. J. Pharmacol. 2019, 176, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Baker, L.M.; Raudonikiene, A.; Hoffman, P.S.; Poole, L.B. Essential Thioredoxin-Dependent Peroxiredoxin System from Helicobacter pylori: Genetic and Kinetic Characterization. J. Bacteriol. 2001, 183, 1961–1973. [Google Scholar] [CrossRef] [Green Version]
- Kuhns, L.G.; Wang, G.; Maier, R.J. Comparative Roles of the Two Helicobacter pylori Thioredoxins in Preventing Macromolecule Damage. Infect. Immun. 2015, 83, 2935–2943. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, T.N.; Sahlin, M.; Lu, J.; Sjöberg, B.-M.; Holmgren, A. Bacillus anthracis Thioredoxin Systems, Characterization and Role as Electron Donors for Ribonucleotide Reductase. J. Biol. Chem. 2012, 287, 39686–39697. [Google Scholar] [CrossRef] [Green Version]
- Pader, I.; Sengupta, R.; Cebula, M.; Xu, J.; Lundberg, J.O.; Holmgren, A.; Johansson, K.; Arnér, E.S.J. Thioredoxin-Related Protein of 14 KDa Is an Efficient L-Cystine Reductase and S-Denitrosylase. Proc. Natl. Acad. Sci. USA 2014, 111, 6964–6969. [Google Scholar] [CrossRef] [Green Version]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A Novel Persulfide Detection Method Reveals Protein Persulfide- and Polysulfide-Reducing Functions of Thioredoxin and Glutathione Systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhou, J.; Omelchenko, M.V.; Beliaev, A.S.; Venkateswaran, A.; Stair, J.; Wu, L.; Thompson, D.K.; Xu, D.; Rogozin, I.B.; et al. Transcriptome Dynamics of Deinococcus radiodurans Recovering from Ionizing Radiation. Proc. Natl. Acad. Sci. USA 2003, 100, 4191–4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xu, G.; Chen, H.; Zhao, Y.; Xu, N.; Tian, B.; Hua, Y. DrRRA: A Novel Response Regulator Essential for the Extreme Radioresistance of Deinococcus radiodurans. Mol. MicroBiol. 2008, 67, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Earl, A.M.; Howell, H.A.; Park, M.J.; Eisen, J.A.; Peterson, S.N.; Battista, J.R. Analysis of Deinococcus radiodurans’s Transcriptional Response to Ionizing Radiation and Desiccation Reveals Novel Proteins That Contribute to Extreme Radioresistance. Genetics 2004, 168, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgenstern, M.; Stiller, S.B.; Lübbert, P.; Peikert, C.D.; Dannenmaier, S.; Drepper, F.; Weill, U.; Höß, P.; Feuerstein, R.; Gebert, M.; et al. Definition of a High-Confidence Mitochondrial Proteome at Quantitative Scale. Cell Rep. 2017, 19, 2836–2852. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Khairnar, N.P.; Joe, M.H.; Misra, H.S.; Lim, S.Y.; Kim, D.H. FrnE, a Cadmium-Inducible Protein in Deinococcus radiodurans, Is Characterized as a Disulfide Isomerase Chaperone In Vitro and for Its Role in Oxidative Stress Tolerance In Vivo. J. Bacteriol. 2013, 195, 2880–2886. [Google Scholar] [CrossRef] [Green Version]
- Bihani, S.C.; Panicker, L.; Rajpurohit, Y.S.; Misra, H.S.; Kumar, V. DrFrnE Represents a Hitherto Unknown Class of Eubacterial Cytoplasmic Disulfide Oxido-Reductases. Antioxid. Redox. Signal. 2018, 28, 296–310. [Google Scholar] [CrossRef]
- Crow, A.; Lewin, A.; Hecht, O.; Carlsson Möller, M.; Moore, G.R.; Hederstedt, L.; Le Brun, N.E. Crystal Structure and Biophysical Properties of Bacillus subtilis BdbD. An Oxidizing Thiol:Disulfide Oxidoreductase Containing a Novel Metal Site. J. Biol. Chem. 2009, 284, 23719–23733. [Google Scholar] [CrossRef] [Green Version]
- Depuydt, M.; Leonard, S.E.; Vertommen, D.; Denoncin, K.; Morsomme, P.; Wahni, K.; Messens, J.; Carroll, K.S.; Collet, J.F. A Periplasmic Reducing System Protects Single Cysteine Residues from Oxidation. Science 2009, 326, 1109–1111. [Google Scholar] [CrossRef]
- Bushweller, J.H. Protein Disulfide Exchange by the Intramembrane Enzymes DsbB, DsbD, and CcdA. J. Mol. Biol. 2020, 432, 5091–5103. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.M.N.; Gupta, R.S. Novel Sequence Features of DNA Repair Genes/Proteins from Deinococcus Species Implicated in Protection from Oxidatively Generated Damage. Genes 2018, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, M.E.S.; Hamilton, A.J.; Dankowski, T.; Heras, B.; Schembri, M.S.; Edwards, J.L.; Jennings, M.P.; McEwan, A.G. A Periplasmic Thioredoxin-like Protein Plays a Role in Defense against Oxidative Stress in Neisseria gonorrhoeae. Infect. Immun. 2009, 77, 4934–4939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanboon, W.; Chuchue, T.; Vattanaviboon, P.; Mongkolsuk, S. Inactivation of Thioredoxin-like Gene Alters Oxidative Stress Resistance and Reduces Cytochrome c Oxidase Activity in Agrobacterium tumefaciens. FEMS MicroBiol. Lett. 2009, 295, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Mohorko, E.; Abicht, H.K.; Bühler, D.; Glockshuber, R.; Hennecke, H.; Fischer, H.-M. Thioredoxin-like Protein TlpA from Bradyrhizobium japonicum Is a Reductant for the Copper Metallochaperone ScoI. FEBS Lett. 2012, 586, 4094–4099. [Google Scholar] [CrossRef] [Green Version]
- Roszczenko, P.; Grzeszczuk, M.; Kobierecka, P.; Wywial, E.; Urbanowicz, P.; Wincek, P.; Nowak, E.; Jagusztyn-Krynicka, E.K. Helicobacter pylori HP0377, a Member of the Dsb Family, Is an Untypical Multifunctional CcmG That Cooperates with Dimeric Thioldisulfide Oxidase HP0231. BMC MicroBiol. 2015, 15, 135. [Google Scholar] [CrossRef] [Green Version]
- Goulding, C.W.; Apostol, M.I.; Gleiter, S.; Parseghian, A.; Bardwell, J.; Gennaro, M.; Eisenberg, D. Gram-Positive DsbE Proteins Function Differently from Gram-Negative DsbE Homologs. A Structure to Function Analysis of DsbE from Mycobacterium tuberculosis. J. Biol. Chem. 2004, 279, 3516–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Levine, R.L. Methionine in Proteins Defends against Oxidative Stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Brot, N.; Weissbach, L.; Werth, J.; Weissbach, H. Enzymatic Reduction of Protein-Bound Methionine Sulfoxide. Proc. Natl. Acad. Sci. USA 1981, 78, 2155–2158. [Google Scholar] [CrossRef] [Green Version]
- Grimaud, R.; Ezraty, B.; Mitchell, J.K.; Lafitte, D.; Briand, C.; Derrick, P.J.; Barras, F. Repair of Oxidized Proteins. Identification of a New Methionine Sulfoxide Reductase. J. Biol. Chem. 2001, 276, 48915–48920. [Google Scholar] [CrossRef] [Green Version]
- Lowther, W.T.; Brot, N.; Weissbach, H.; Honek, J.F.; Matthews, B.W. Thiol-Disulfide Exchange Is Involved in the Catalytic Mechanism of Peptide Methionine Sulfoxide Reductase. Proc. Natl. Acad. Sci. USA 2000, 97, 6463–6468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrago, L.; Kaya, A.; Weerapana, E.; Marino, S.M.; Gladyshev, V.N. Methionine Sulfoxide Reductases Preferentially Reduce Unfolded Oxidized Proteins and Protect Cells from Oxidative Protein Unfolding. J. Biol. Chem. 2012, 287, 24448–24459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boschi-Muller, S.; Branlant, G. Methionine Sulfoxide Reductase: Chemistry, Substrate Binding, Recycling Process and Oxidase Activity. Bioorg. Chem. 2014, 57, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Tarrago, L.; Laugier, E.; Zaffagnini, M.; Marchand, C.; Le Maréchal, P.; Rouhier, N.; Lemaire, S.D.; Rey, P. Regeneration Mechanisms of Arabidopsis thaliana Methionine Sulfoxide Reductases B by Glutaredoxins and Thioredoxins. J. Biol. Chem. 2009, 284, 18963–18971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-H.; Weissbach, H. Origin and Evolution of the Protein-Repairing Enzymes Methionine Sulphoxide Reductases. Biol. Rev. Camb. Philos. Soc. 2008, 83, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Koc, A.; Gasch, A.P.; Rutherford, J.C.; Kim, H.-Y.; Gladyshev, V.N. Methionine Sulfoxide Reductase Regulation of Yeast Lifespan Reveals Reactive Oxygen Species-Dependent and -Independent Components of Aging. Proc. Natl. Acad. Sci. USA 2004, 101, 7999–8004. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, I.; Mary, J.; Perichon, M.; Friguet, B. Rat Peptide Methionine Sulphoxide Reductase: Cloning of the CDNA, and down-Regulation of Gene Expression and Enzyme Activity during Aging. BioChem. J. 2001, 355, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine Sulfoxide Reductases in Prokaryotes. Biochim. Biophys. Acta 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Ruan, H.; Tang, X.D.; Chen, M.-L.; Joiner, M.-L.A.; Sun, G.; Brot, N.; Weissbach, H.; Heinemann, S.H.; Iverson, L.; Wu, C.-F.; et al. High-Quality Life Extension by the Enzyme Peptide Methionine Sulfoxide Reductase. Proc. Natl. Acad. Sci. USA 2002, 99, 2748–2753. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.; Loiseau, L.; Vergnes, A.; Vertommen, D.; Mérida-Floriano, A.; Chitteni-Pattu, S.; Wood, E.A.; Casadesús, J.; Cox, M.M.; Barras, F.; et al. Redox. Controls RecA Protein Activity via Reversible Oxidation of Its Methionine Residues. Elife 2021, 10, e63747. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Moskovitz, J.; Berlett, B.S.; Levine, R.L. Cyclic Oxidation and Reduction of Protein Methionine Residues Is an Important Antioxidant Mechanism. Mol. Cell BioChem. 2002, 234–235, 3–9. [Google Scholar] [CrossRef]
- Erickson, J.R.; Joiner, M.A.; Guan, X.; Kutschke, W.; Yang, J.; Oddis, C.V.; Bartlett, R.K.; Lowe, J.S.; O’Donnell, S.E.; Aykin-Burns, N.; et al. A Dynamic Pathway for Calcium-Independent Activation of CaMKII by Methionine Oxidation. Cell 2008, 133, 462–474. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Péterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs Regulate Actin Assembly and Macrophage Function via Reversible Stereoselective Methionine Oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Rey, P.; Tarrago, L. Physiological Roles of Plant Methionine Sulfoxide Reductases in Redox Homeostasis and Signaling. Antioxidants 2018, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drazic, A.; Miura, H.; Peschek, J.; Le, Y.; Bach, N.C.; Kriehuber, T.; Winter, J. Methionine Oxidation Activates a Transcription Factor in Response to Oxidative Stress. Proc. Natl. Acad. Sci. USA 2013, 110, 9493–9498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruckbauer, S.T.; Minkoff, B.B.; Yu, D.; Cryns, V.L.; Cox, M.M.; Sussman, M.R. Ionizing Radiation-Induced Proteomic Oxidation in Escherichia coli. Mol. Cell Proteomics 2020, 19, 1375–1395. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.L.; Stanley, J.A.; Robinson, M.C.; Sher, J.W.; Li, Z.; Chan, Y.A.; Omdahl, A.R.; Wattiez, R.; Godzik, A.; Matallana-Surget, S. Protein Structure, Amino Acid Composition and Sequence Determine Proteome Vulnerability to Oxidation-Induced Damage. EMBO J. 2020, 39, e104523. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Vieira Dos Santos, C.; Tarrago, L.; Rey, P. Plant Methionine Sulfoxide Reductase A and B Multigenic Families. Photosynth. Res. 2006, 89, 247–262. [Google Scholar] [CrossRef]
- Coudevylle, N.; Antoine, M.; Bouguet-Bonnet, S.; Mutzenhardt, P.; Boschi-Muller, S.; Branlant, G.; Cung, M.-T. Solution Structure and Backbone Dynamics of the Reduced Form and an Oxidized Form of E. coli Methionine Sulfoxide Reductase A (MsrA): Structural Insight of the MsrA Catalytic Cycle. J. Mol. Biol. 2007, 366, 193–206. [Google Scholar] [CrossRef]
- Delaye, L.; Becerra, A.; Orgel, L.; Lazcano, A. Molecular Evolution of Peptide Methionine Sulfoxide Reductases (MsrA and MsrB): On the Early Development of a Mechanism That Protects against Oxidative Damage. J. Mol. Evol. 2007, 64, 15–32. [Google Scholar] [CrossRef]
- Kumar, R.A.; Koc, A.; Cerny, R.L.; Gladyshev, V.N. Reaction Mechanism, Evolutionary Analysis, and Role of Zinc in Drosophila Methionine-R-Sulfoxide Reductase. J. Biol. Chem. 2002, 277, 37527–37535. [Google Scholar] [CrossRef] [Green Version]
- Olry, A.; Boschi-Muller, S.; Branlant, G. Kinetic Characterization of the Catalytic Mechanism of Methionine Sulfoxide Reductase B from Neisseria meningitidis. Biochemistry 2004, 43, 11616–11622. [Google Scholar] [CrossRef]
- Neiers, F.; Kriznik, A.; Boschi-Muller, S.; Branlant, G. Evidence for a New Sub-Class of Methionine Sulfoxide Reductases B with an Alternative Thioredoxin Recognition Signature. J. Biol. Chem. 2004, 279, 42462–42468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappler, U.; Nasreen, M.; McEwan, A. New Insights into the Molecular Physiology of Sulfoxide Reduction in Bacteria. Adv. Microb. Physiol. 2019, 75, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Alamuri, P.; Maier, R.J. Methionine Sulphoxide Reductase Is an Important Antioxidant Enzyme in the Gastric Pathogen Helicobacter pylori. Mol. MicroBiol. 2004, 53, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S. Molecular Mechanisms of the Methionine Sulfoxide Reductase System from Neisseria meningitidis. Antioxidants 2018, 7, 131. [Google Scholar] [CrossRef] [Green Version]
- Gennaris, A.; Ezraty, B.; Henry, C.; Agrebi, R.; Vergnes, A.; Oheix, E.; Bos, J.; Leverrier, P.; Espinosa, L.; Szewczyk, J.; et al. Repairing Oxidized Proteins in the Bacterial Envelope Using Respiratory Chain Electrons. Nature 2015, 528, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Perkins, A.; Nelson, K.J.; Parsonage, D.; Poole, L.B.; Karplus, P.A. Peroxiredoxins: Guardians against Oxidative Stress and Modulators of Peroxide Signaling. Trends BioChem. Sci. 2015, 40, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G.; Woo, H.A. Multiple Functions of Peroxiredoxins: Peroxidases, Sensors and Regulators of the Intracellular Messenger H₂O₂, and Protein Chaperones. Antioxid. Redox. Signal. 2011, 15, 781–794. [Google Scholar] [CrossRef] [PubMed]
- Day, A.M.; Brown, J.D.; Taylor, S.R.; Rand, J.D.; Morgan, B.A.; Veal, E.A. Inactivation of a Peroxiredoxin by Hydrogen Peroxide Is Critical for Thioredoxin-Mediated Repair of Oxidized Proteins and Cell Survival. Mol. Cell 2012, 45, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Stöcker, S.; Maurer, M.; Ruppert, T.; Dick, T.P. A Role for 2-Cys Peroxiredoxins in Facilitating Cytosolic Protein Thiol Oxidation. Nat. Chem. Biol. 2018, 14, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.Z.; Kim, I.H.; Kim, K.; Rhee, S.G. Cloning, Sequencing, and Mutation of Thiol-Specific Antioxidant Gene of Saccharomyces cerevisiae. J. Biol. Chem. 1993, 268, 16815–16821. [Google Scholar] [CrossRef]
- de Oliveira, M.A.; Tairum, C.A.; Netto, L.E.S.; de Oliveira, A.L.P.; Aleixo-Silva, R.L.; Cabrera, V.I.M.; Breyer, C.A.; Dos Santos, M.C. Relevance of Peroxiredoxins in Pathogenic Microorganisms. Appl. MicroBiol. Biotechnol. 2021, 105, 5701–5717. [Google Scholar] [CrossRef]
- Poole, L.B.; Ellis, H.R. Flavin-Dependent Alkyl Hydroperoxide Reductase from Salmonella typhimurium. 1. Purification and Enzymatic Activities of Overexpressed AhpF and AhpC Proteins. Biochemistry 1996, 35, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Cha, M.K.; Kim, I.H. Thioredoxin-Dependent Hydroperoxide Peroxidase Activity of Bacterioferritin Comigratory Protein (BCP) as a New Member of the Thiol-Specific Antioxidant Protein (TSA)/Alkyl Hydroperoxide Peroxidase C (AhpC) Family. J. Biol. Chem. 2000, 275, 2924–2930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouhier, N.; Gelhaye, E.; Gualberto, J.M.; Jordy, M.-N.; De Fay, E.; Hirasawa, M.; Duplessis, S.; Lemaire, S.D.; Frey, P.; Martin, F.; et al. Poplar Peroxiredoxin Q. A Thioredoxin-Linked Chloroplast Antioxidant Functional in Pathogen Defense. Plant. Physiol. 2004, 134, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.J.; Mackay, C.L.; Campopiano, D.J.; Langridge-Smith, P.; Brown, A.R. Interrogating the Molecular Details of the Peroxiredoxin Activity of the Escherichia coli Bacterioferritin Comigratory Protein Using High-Resolution Mass Spectrometry. Biochemistry 2009, 48, 3904–3914. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.J.; Ortega, X.P.; Mackay, C.L.; Valvano, M.A.; Govan, J.R.W.; Campopiano, D.J.; Langridge-Smith, P.; Brown, A.R. Subdivision of the Bacterioferritin Comigratory Protein Family of Bacterial Peroxiredoxins Based on Catalytic Activity. Biochemistry 2010, 49, 1319–1330. [Google Scholar] [CrossRef]
- Hicks, L.D.; Raghavan, R.; Battisti, J.M.; Minnick, M.F. A DNA-Binding Peroxiredoxin of Coxiella burnetii Is Involved in Countering Oxidative Stress during Exponential-Phase Growth. J. Bacteriol. 2010, 192, 2077–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Chung, J.M.; Yun, H.J.; Won, J.; Jung, H.S. New Insight into Multifunctional Role of Peroxiredoxin Family Protein: Determination of DNA Protection Properties of Bacterioferritin Comigratory Protein under Hyperthermal and Oxidative Stresses. BioChem. Biophys. Res. Commun. 2016, 469, 1028–1033. [Google Scholar] [CrossRef]
- Chae, H.Z.; Robison, K.; Poole, L.B.; Church, G.; Storz, G.; Rhee, S.G. Cloning and Sequencing of Thiol-Specific Antioxidant from Mammalian Brain: Alkyl Hydroperoxide Reductase and Thiol-Specific Antioxidant Define a Large Family of Antioxidant Enzymes. Proc. Natl. Acad. Sci. USA 1994, 91, 7017–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, F.S.; Morgan, R.W.; Christman, M.F.; Ames, B.N. An Alkyl Hydroperoxide Reductase from Salmonella typhimurium Involved in the Defense of DNA against Oxidative Damage. Purification and Properties. J. Biol. Chem. 1989, 264, 1488–1496. [Google Scholar] [CrossRef]
- Storz, G.; Jacobson, F.S.; Tartaglia, L.A.; Morgan, R.W.; Silveira, L.A.; Ames, B.N. An Alkyl Hydroperoxide Reductase Induced by Oxidative Stress in Salmonella typhimurium and Escherichia coli: Genetic Characterization and Cloning of ahp. J. Bacteriol. 1989, 171, 2049–2055. [Google Scholar] [CrossRef] [Green Version]
- Poole, L.B.; Reynolds, C.M.; Wood, Z.A.; Karplus, P.A.; Ellis, H.R.; Li Calzi, M. AhpF and Other NADH:Peroxiredoxin Oxidoreductases, Homologues of Low Mr Thioredoxin Reductase. Eur. J. BioChem. 2000, 267, 6126–6133. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B.; Godzik, A.; Nayeem, A.; Schmitt, J.D. AhpF Can Be Dissected into Two Functional Units: Tandem Repeats of Two Thioredoxin-like Folds in the N-Terminus Mediate Electron Transfer from the Thioredoxin Reductase-like C-Terminus to AhpC. Biochemistry 2000, 39, 6602–6615. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two Enzymes in One; Two Yeast Peroxiredoxins Display Oxidative Stress-Dependent Switching from a Peroxidase to a Molecular Chaperone Function. Cell 2004, 117, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Parsonage, D.; Youngblood, D.S.; Sarma, G.N.; Wood, Z.A.; Karplus, P.A.; Poole, L.B. Analysis of the Link between Enzymatic Activity and Oligomeric State in AhpC, a Bacterial Peroxiredoxin. Biochemistry 2005, 44, 10583–10592. [Google Scholar] [CrossRef] [Green Version]
- Mongkolsuk, S.; Whangsuk, W.; Vattanaviboon, P.; Loprasert, S.; Fuangthong, M. A Xanthomonas Alkyl Hydroperoxide Reductase Subunit C (AhpC) Mutant Showed an Altered Peroxide Stress Response and Complex Regulation of the Compensatory Response of Peroxide Detoxification Enzymes. J. Bacteriol. 2000, 182, 6845–6849. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Jeong, H.; Lee, J.H.; Chung, J.M.; Kim, R.; Yun, H.J.; Won, J.; Jung, H.S. Characterisation of Conformational and Functional Features of Alkyl Hydroperoxide Reductase E-like Protein. BioChem. Biophys Res. Commun. 2017, 489, 217–222. [Google Scholar] [CrossRef]
- Li, S.; Peterson, N.A.; Kim, M.-Y.; Kim, C.-Y.; Hung, L.-W.; Yu, M.; Lekin, T.; Segelke, B.W.; Lott, J.S.; Baker, E.N. Crystal Structure of AhpE from Mycobacterium tuberculosis, a 1-Cys Peroxiredoxin. J. Mol. Biol. 2005, 346, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.M.; Hugo, M.; Trostchansky, A.; Capece, L.; Radi, R.; Trujillo, M. Oxidizing Substrate Specificity of Mycobacterium tuberculosis Alkyl Hydroperoxide Reductase E: Kinetics and Mechanisms of Oxidation and Overoxidation. Free Radic. Biol. Med. 2011, 51, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Bryk, R.; Lima, C.D.; Erdjument-Bromage, H.; Tempst, P.; Nathan, C. Metabolic Enzymes of Mycobacteria Linked to Antioxidant Defense by a Thioredoxin-like Protein. Science 2002, 295, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.-J.; Jeong, H.; Lee, D.-S.; Kim, Y.; Lee, H.-S. The ahpD Gene of Corynebacterium glutamicum Plays an Important Role in Hydrogen Peroxide-Induced Oxidative Stress Response. J. BioChem. 2019, 165, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hillas, P.J.; del Alba, F.S.; Oyarzabal, J.; Wilks, A.; Ortiz De Montellano, P.R. The AhpC and AhpD Antioxidant Defense System of Mycobacterium tuberculosis. J. Biol. Chem. 2000, 275, 18801–18809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Balakrishna, A.M.; Nartey, W.; Manimekalai, M.S.S.; Grüber, G. Redox. Chemistry of Mycobacterium tuberculosis Alkylhydroperoxide Reductase E (AhpE): Structural and Mechanistic Insight into a Mycoredoxin-1 Independent Reductive Pathway of AhpE via Mycothiol. Free Radic. Biol. Med. 2016, 97, 588–601. [Google Scholar] [CrossRef]
- Zhao, L.; Jeong, S.; Zhang, J.; Jung, J.-H.; Choi, J.-I.; Lim, S.; Kim, M.-K. Crystal Structure of the AhpD-like Protein DR1765 from Deinococcus radiodurans R1. BioChem. Biophys Res. Commun. 2020, 529, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsuk, S.; Praituan, W.; Loprasert, S.; Fuangthong, M.; Chamnongpol, S. Identification and Characterization of a New Organic Hydroperoxide Resistance (ohr) Gene with a Novel Pattern of Oxidative Stress Regulation from Xanthomonas campestris Pv. phaseoli. J. Bacteriol. 1998, 180, 2636–2643. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, C.; Devedjian, J.C. Osmotic Induction of Gene osmC Expression in Escherichia coli K12. J. Mol. Biol. 1991, 220, 959–973. [Google Scholar] [CrossRef]
- Shin, D.H.; Choi, I.G.; Busso, D.; Jancarik, J.; Yokota, H.; Kim, R.; Kim, S.H. Structure of OsmC from Escherichia coli: A Salt-Shock-Induced Protein. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 903–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atichartpongkul, S.; Loprasert, S.; Vattanaviboon, P.; Whangsuk, W.; Helmann, J.D.; Mongkolsuk, S. Bacterial Ohr and OsmC Paralogues Define Two Protein Families with Distinct Functions and Patterns of Expression. Microbiology 2001, 147, 1775–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meireles, D.A.; Domingos, R.M.; Gaiarsa, J.W.; Ragnoni, E.G.; Bannitz-Fernandes, R.; da Silva Neto, J.F.; de Souza, R.F.; Netto, L.E.S. Functional and Evolutionary Characterization of Ohr Proteins in Eukaryotes Reveals Many Active Homologs among Pathogenic Fungi. Redox. Biol. 2017, 12, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Alegria, T.G.P.; Meireles, D.A.; Cussiol, J.R.R.; Hugo, M.; Trujillo, M.; de Oliveira, M.A.; Miyamoto, S.; Queiroz, R.F.; Valadares, N.F.; Garratt, R.C.; et al. Ohr Plays a Central Role in Bacterial Responses against Fatty Acid Hydroperoxides and Peroxynitrite. Proc. Natl. Acad. Sci. USA 2017, 114, E132–E141. [Google Scholar] [CrossRef] [Green Version]
- Lesniak, J.; Barton, W.A.; Nikolov, D.B. Structural and Functional Characterization of the Pseudomonas Hydroperoxide Resistance Protein Ohr. EMBO J. 2002, 21, 6649–6659. [Google Scholar] [CrossRef] [Green Version]
- Lesniak, J.; Barton, W.A.; Nikolov, D.B. Structural and Functional Features of the Escherichia coli Hydroperoxide Resistance Protein OsmC. Protein Sci. 2003, 12, 2838–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Baseman, J.B. Functional Characterization of Osmotically Inducible Protein C (MG_427) from Mycoplasma genitalium. J. Bacteriol. 2014, 196, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Cussiol, J.R.R.; Alegria, T.G.P.; Szweda, L.I.; Netto, L.E.S. Ohr (Organic Hydroperoxide Resistance Protein) Possesses a Previously Undescribed Activity, Lipoyl-Dependent Peroxidase. J. Biol. Chem. 2010, 285, 21943–21950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, M.; Su, T.; Chen, C.; Wei, Z.; Gong, Z.; Li, G. OsmC in Corynebacterium glutamicum Was a Thiol-Dependent Organic Hydroperoxide Reductase. Int. J. Biol. Macromol. 2019, 136, 642–652. [Google Scholar] [CrossRef]
- Basu, B.; Apte, S.K. Gamma Radiation-Induced Proteome of Deinococcus radiodurans Primarily Targets DNA Repair and Oxidative Stress Alleviation. Mol. Cell Proteomics 2012, 11, M111.011734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meunier-Jamin, C.; Kapp, U.; Leonard, G.A.; McSweeney, S. The Structure of the Organic Hydroperoxide Resistance Protein from Deinococcus radiodurans. Do Conformational Changes Facilitate Recycling of the Redox. Disulfide? J. Biol. Chem. 2004, 279, 25830–25837. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, J.; Liu, M.; Yang, S.; Zhao, Y.; Cheng, K.; Xu, G.; Li, M.; Tian, B.; Hua, Y. Proteomic Insights into the Functional Basis for the Response Regulator DrRRA of Deinococcus radiodurans. Int. J. Radiat. Biol. 2016, 92, 273–280. [Google Scholar] [CrossRef]
- Hiras, J.; Sharma, S.V.; Raman, V.; Tinson, R.A.J.; Arbach, M.; Rodrigues, D.F.; Norambuena, J.; Hamilton, C.J.; Hanson, T.E. Physiological Studies of Chlorobiaceae Suggest That Bacillithiol Derivatives Are the Most Widespread Thiols in Bacteria. mBio 2018, 9, e01603-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, G.L.; Rawat, M.; La Clair, J.J.; Jothivasan, V.K.; Budiarto, T.; Hamilton, C.J.; Claiborne, A.; Helmann, J.D.; Fahey, R.C. Bacillithiol Is an Antioxidant Thiol Produced in Bacilli. Nat. Chem. Biol. 2009, 5, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Linzner, N.; Loi, V.V.; Fritsch, V.N.; Tung, Q.N.; Stenzel, S.; Wirtz, M.; Hell, R.; Hamilton, C.J.; Tedin, K.; Fulde, M.; et al. Staphylococcus aureus Uses the Bacilliredoxin (BrxAB)/Bacillithiol Disulfide Reductase (YpdA) Redox. Pathway to Defend against Oxidative Stress under Infections. Front. MicroBiol. 2019, 10, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerstad, M.; Gudim, I.; Hersleth, H.-P. The Crystal Structures of Bacillithiol Disulfide Reductase Bdr (YpdA) Provide Structural and Functional Insight into a New Type of FAD-Containing NADPH-Dependent Oxidoreductase. Biochemistry 2020, 59, 4793–4798. [Google Scholar] [CrossRef]
- Gaballa, A.; Su, T.T.; Helmann, J.D. The Bacillus subtilis Monothiol Bacilliredoxin BrxC (YtxJ) and the Bdr (YpdA) Disulfide Reductase Reduce S-Bacillithiolated Proteins. Redox. Biol. 2021, 42, 101935. [Google Scholar] [CrossRef]
- Mikheyeva, I.V.; Thomas, J.M.; Kolar, S.L.; Corvaglia, A.-R.; Gaϊa, N.; Leo, S.; Francois, P.; Liu, G.Y.; Rawat, M.; Cheung, A.L. YpdA, a Putative Bacillithiol Disulfide Reductase, Contributes to Cellular Redox. Homeostasis and Virulence in Staphylococcus aureus. Mol. MicroBiol. 2019, 111, 1039–1056. [Google Scholar] [CrossRef]
- Chi, B.K.; Roberts, A.A.; Huyen, T.T.T.; Bäsell, K.; Becher, D.; Albrecht, D.; Hamilton, C.J.; Antelmann, H. S-Bacillithiolation Protects Conserved and Essential Proteins against Hypochlorite Stress in Firmicutes Bacteria. Antioxid. Redox. Signal. 2013, 18, 1273–1295. [Google Scholar] [CrossRef] [Green Version]
- Imber, M.; Huyen, N.T.T.; Pietrzyk-Brzezinska, A.J.; Loi, V.V.; Hillion, M.; Bernhardt, J.; Thärichen, L.; Kolšek, K.; Saleh, M.; Hamilton, C.J.; et al. Protein S-Bacillithiolation Functions in Thiol Protection and Redox. Regulation of the Glyceraldehyde-3-Phosphate Dehydrogenase Gap in Staphylococcus aureus under Hypochlorite Stress. Antioxid. Redox. Signal. 2018, 28, 410–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaballa, A.; Chi, B.K.; Roberts, A.A.; Becher, D.; Hamilton, C.J.; Antelmann, H.; Helmann, J.D. Redox. Regulation in Bacillus subtilis: The Bacilliredoxins BrxA(YphP) and BrxB(YqiW) Function in de-Bacillithiolation of S-Bacillithiolated OhrR and MetE. Antioxid. Redox. Signal. 2014, 21, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wei, J.; Zheng, Z.; Ying, N.; Sheng, D.; Hua, Y. Proteomic Analysis of Deinococcus radiodurans Recovering from Gamma-Irradiation. Proteomics 2005, 5, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Ujaoney, A.K.; Padwal, M.K.; Basu, B. Proteome Dynamics during Post-Desiccation Recovery Reveal Convergence of Desiccation and Gamma Radiation Stress Response Pathways in Deinococcus radiodurans. Biochim Biophys Acta Proteins Proteom 2017, 1865, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, T.; Murata, K.; Kawai, S. Extremely High Intracellular Concentration of Glucose-6-Phosphate and NAD(H) in Deinococcus radiodurans. Extremophiles 2017, 21, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Limauro, D.; Pedone, E.; Galdi, I.; Bartolucci, S. Peroxiredoxins as Cellular Guardians in Sulfolobus solfataricus: Characterization of Bcp1, Bcp3 and Bcp4. FEBS J. 2008, 275, 2067–2077. [Google Scholar] [CrossRef]
- Parsonage, D.; Karplus, P.A.; Poole, L.B. Substrate Specificity and Redox. Potential of AhpC, a Bacterial Peroxiredoxin. Proc. Natl. Acad. Sci. USA 2008, 105, 8209–8214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinz, W.A.; Aslund, F.; Holmgren, A.; Beckwith, J. The Role of the Thioredoxin and Glutaredoxin Pathways in Reducing Protein Disulfide Bonds in the Escherichia coli Cytoplasm. J. Biol. Chem. 1997, 272, 15661–15667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashandy, T.; Guilleminot, J.; Vernoux, T.; Caparros-Ruiz, D.; Ljung, K.; Meyer, Y.; Reichheld, J.-P. Interplay between the NADP-Linked Thioredoxin and Glutathione Systems in Arabidopsis Auxin Signaling. Plant. Cell 2010, 22, 376–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaballa, A.; Newton, G.L.; Antelmann, H.; Parsonage, D.; Upton, H.; Rawat, M.; Claiborne, A.; Fahey, R.C.; Helmann, J.D. Biosynthesis and Functions of Bacillithiol, a Major Low-Molecular-Weight Thiol in Bacilli. Proc. Natl. Acad. Sci. USA 2010, 107, 6482–6486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@: Network Protein Sequence Analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The Trx system. (a) NADPH-dependent electron transfer via Trx. (b) Catalytic mechanism of substrate reduction by Trx. (c) Trx thiol reductase substrates involved in the maintenance of cell redox homeostasis. TrxR, thioredoxin reductase; Trx, thioredoxin; Prx, peroxiredoxin; Msr, methionine sulfoxide reductase; SCH, catalytic cysteine thiol; SRH, resolving cysteine thiol; MetO, methionine sulfoxide.
Figure 1.
The Trx system. (a) NADPH-dependent electron transfer via Trx. (b) Catalytic mechanism of substrate reduction by Trx. (c) Trx thiol reductase substrates involved in the maintenance of cell redox homeostasis. TrxR, thioredoxin reductase; Trx, thioredoxin; Prx, peroxiredoxin; Msr, methionine sulfoxide reductase; SCH, catalytic cysteine thiol; SRH, resolving cysteine thiol; MetO, methionine sulfoxide.

Figure 2.
Multiple sequence alignments of TRX-like proteins from Deinococcus. (a) Alignment of Deinococcus DR_2085-type sequences. (b) Alignment of Deinococcus DR_2085 sequence with Homo sapiens (Hs) and Helicobacter pylori (Hp) homologous TRP14 Trxs. (c) Alignment of Deinococcus DR_0057 with Arabidopsis thaliana (At), Mus musculus (Mm), Saccharomyces cerevisiae (Sc), Haloplanus aerogenes (He) and Chlorobium ferrooxidans (Cf) homologous proteins. Trx active site sequences are highlighted in yellow. An asterisk (*) indicates positions which have a single, fully conserved residue; colon (:) and period (.) indicate conservation between groups of strongly and weakly similar properties, respectively. Abbreviations of Deinococcus species names are as specified in Table 1. Alignments were generated using UniProt ClustalO (https://www.uniprot.org/align/; accessed on 14 February 2022).
Figure 2.
Multiple sequence alignments of TRX-like proteins from Deinococcus. (a) Alignment of Deinococcus DR_2085-type sequences. (b) Alignment of Deinococcus DR_2085 sequence with Homo sapiens (Hs) and Helicobacter pylori (Hp) homologous TRP14 Trxs. (c) Alignment of Deinococcus DR_0057 with Arabidopsis thaliana (At), Mus musculus (Mm), Saccharomyces cerevisiae (Sc), Haloplanus aerogenes (He) and Chlorobium ferrooxidans (Cf) homologous proteins. Trx active site sequences are highlighted in yellow. An asterisk (*) indicates positions which have a single, fully conserved residue; colon (:) and period (.) indicate conservation between groups of strongly and weakly similar properties, respectively. Abbreviations of Deinococcus species names are as specified in Table 1. Alignments were generated using UniProt ClustalO (https://www.uniprot.org/align/; accessed on 14 February 2022).

Figure 3.
Electron sources for Deinococcus NADPH- and NADH-dependent thiol peroxidases (BCP, AhpE, AhpD, Ohr, OsmC) scavenging hydrogen and organic peroxides and peroxynitrites and participating in redox signalling. TrxR, thioredoxin reductase; Trx, thioredoxin; BCP, bacterioferritin comigratory protein; SucB, dihydrolipoamide succinyltransferase; PDHB, pyruvate dehydrogenase beta; Ahp, alkyl hydroperoxide reductase; Ohr, organic hydroperoxide resistance enzyme; OsmC, osmotically induced bacterial protein C. Dashed arrows indicate possible transfer of electrons.
Figure 3.
Electron sources for Deinococcus NADPH- and NADH-dependent thiol peroxidases (BCP, AhpE, AhpD, Ohr, OsmC) scavenging hydrogen and organic peroxides and peroxynitrites and participating in redox signalling. TrxR, thioredoxin reductase; Trx, thioredoxin; BCP, bacterioferritin comigratory protein; SucB, dihydrolipoamide succinyltransferase; PDHB, pyruvate dehydrogenase beta; Ahp, alkyl hydroperoxide reductase; Ohr, organic hydroperoxide resistance enzyme; OsmC, osmotically induced bacterial protein C. Dashed arrows indicate possible transfer of electrons.

Figure 4.
The bacillithiol system. (a) Structure of bacillithiol (BSH), glycoside formed of L-cysteinyl-D-glucosamine and malic acid. (b) Electron transfer pathways in the BSH system. Bdr, NADPH–bacillithiol disulfide reductase. Brx, bacilliredoxin; S-Ox, thiol oxidised (bacillithiolated) form; S-Red, thiol reduced form.
Figure 4.
The bacillithiol system. (a) Structure of bacillithiol (BSH), glycoside formed of L-cysteinyl-D-glucosamine and malic acid. (b) Electron transfer pathways in the BSH system. Bdr, NADPH–bacillithiol disulfide reductase. Brx, bacilliredoxin; S-Ox, thiol oxidised (bacillithiolated) form; S-Red, thiol reduced form.

Figure 5.
Relationships and possible interplays between Deinococcus thiol reductase systems. Dashed arrows indicate possible transfer of electrons between Trx-, BSH-, and lipoylated-dependent systems. TrxR, thioredoxin reductase; Trx, thioredoxin; Bdr, NADPH–BSH disulfide reductase; BSH, bacillithiol; Brx, bacilliredoxin; Ahp, alkyl hydroperoxide reductase; Ohr, organic hydroperoxide resistance enzyme; OsmC, osmotically induced bacterial protein C.
Figure 5.
Relationships and possible interplays between Deinococcus thiol reductase systems. Dashed arrows indicate possible transfer of electrons between Trx-, BSH-, and lipoylated-dependent systems. TrxR, thioredoxin reductase; Trx, thioredoxin; Bdr, NADPH–BSH disulfide reductase; BSH, bacillithiol; Brx, bacilliredoxin; Ahp, alkyl hydroperoxide reductase; Ohr, organic hydroperoxide resistance enzyme; OsmC, osmotically induced bacterial protein C.


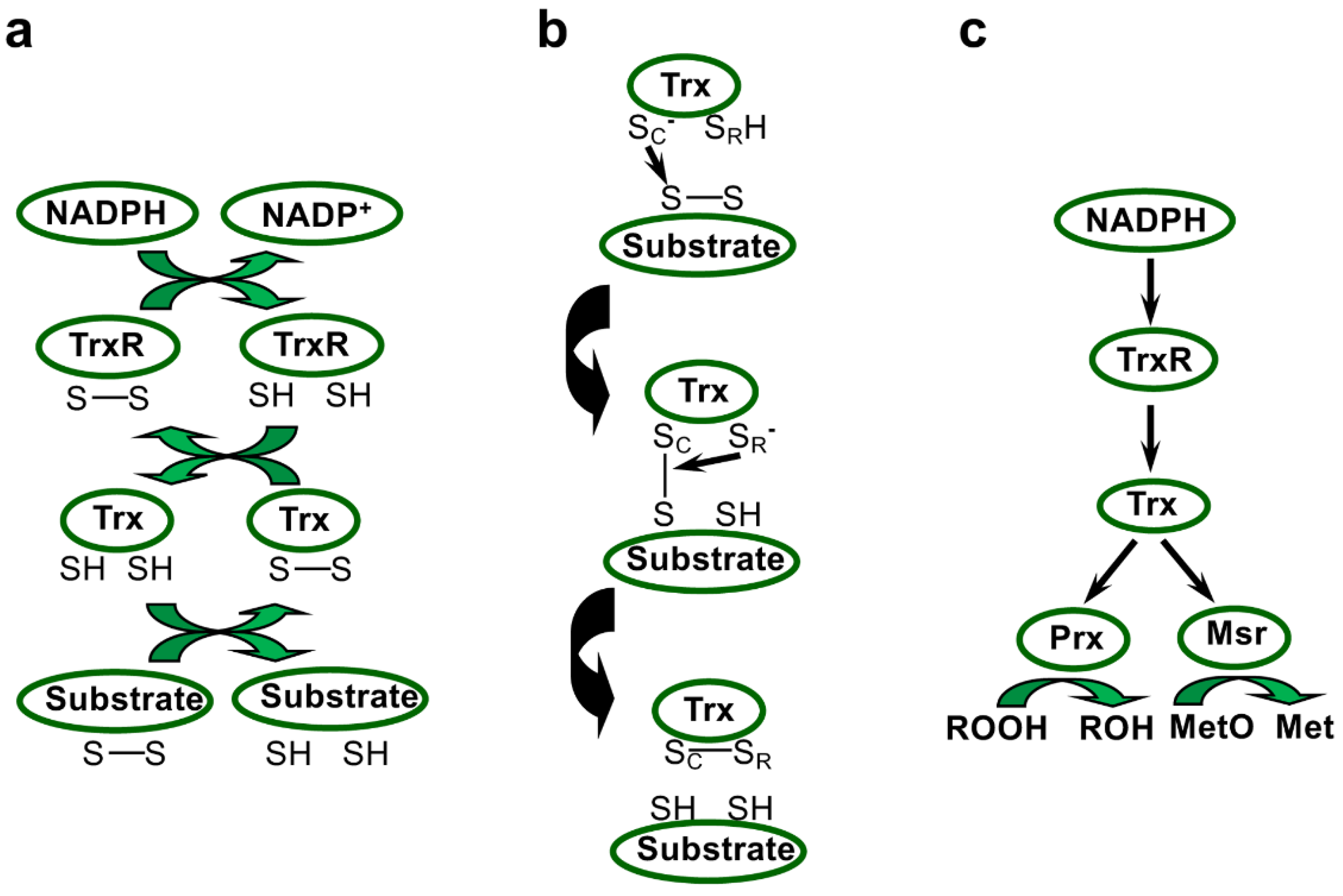{kind=link}
{kind=link}
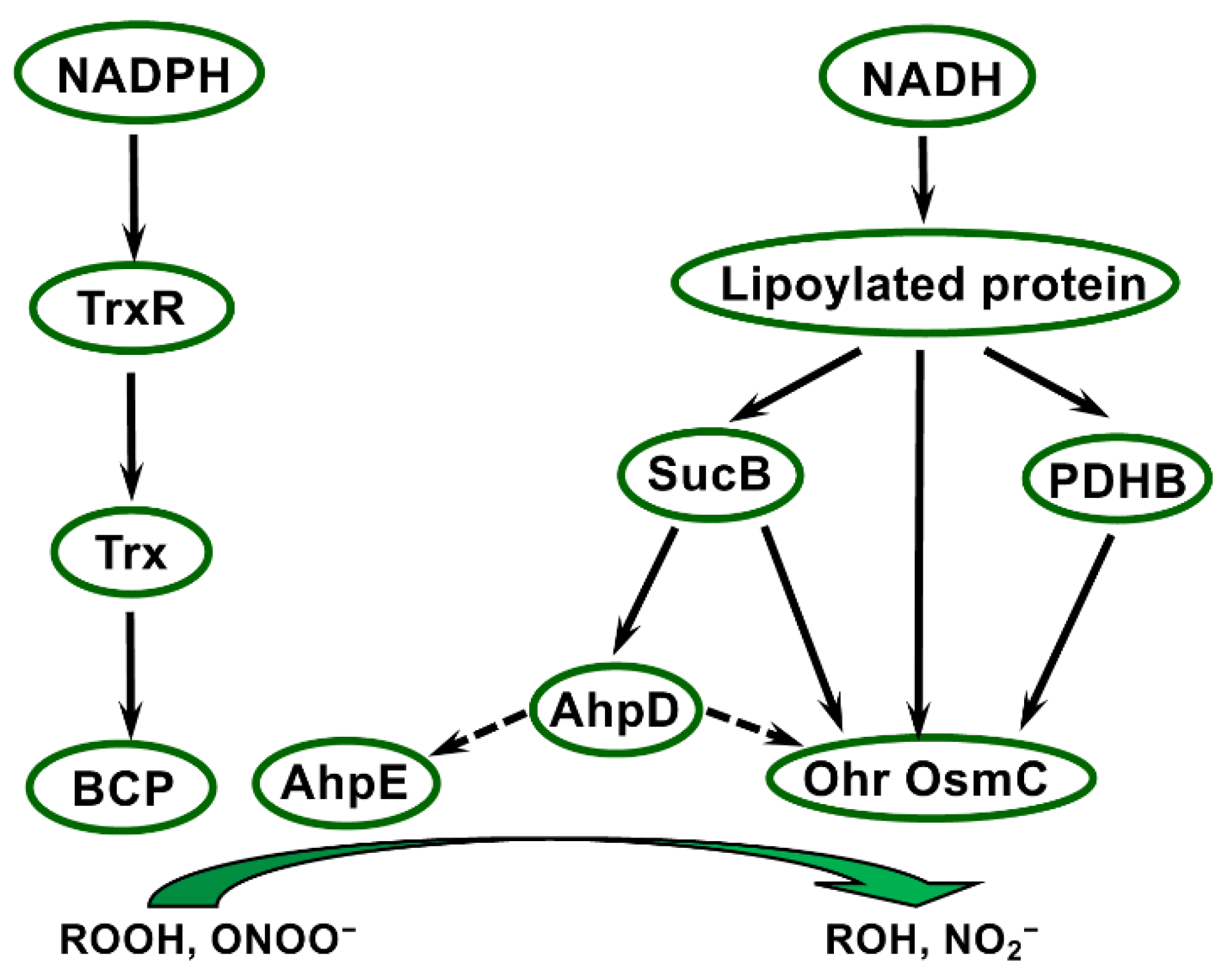{kind=link}
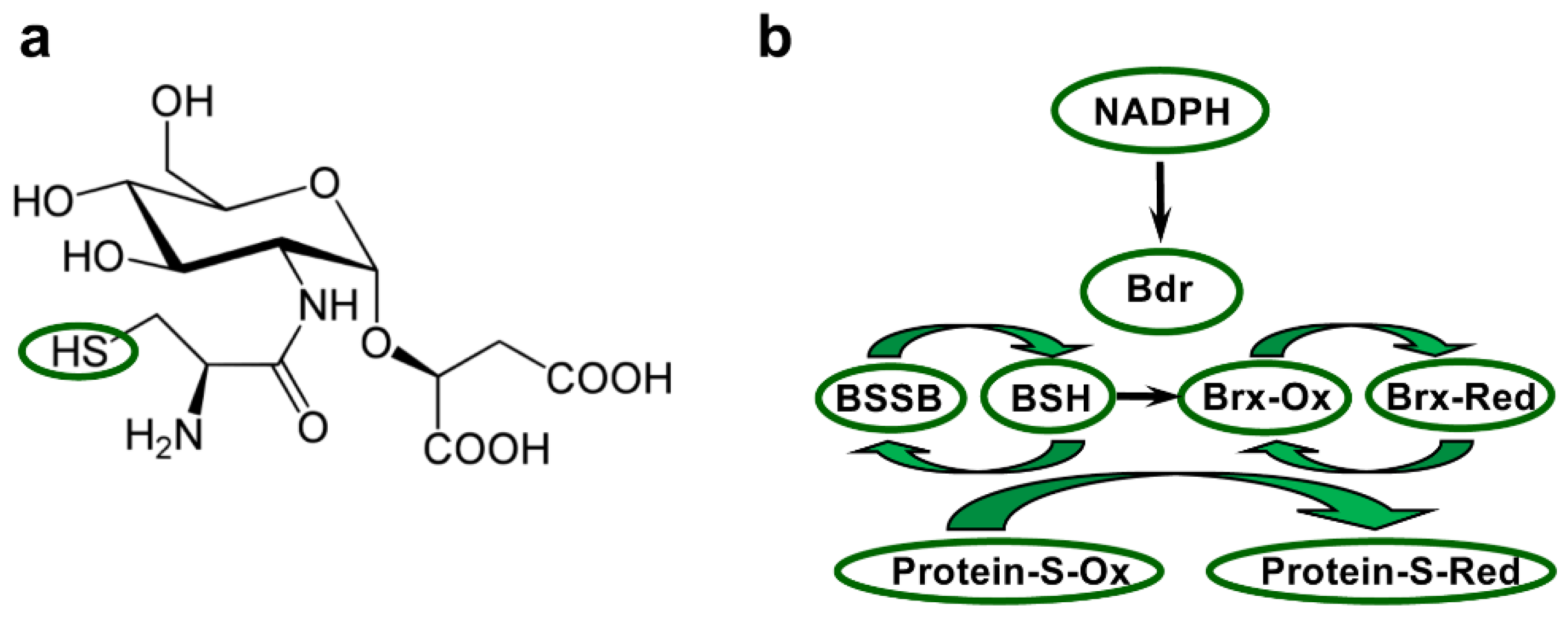{kind=link}
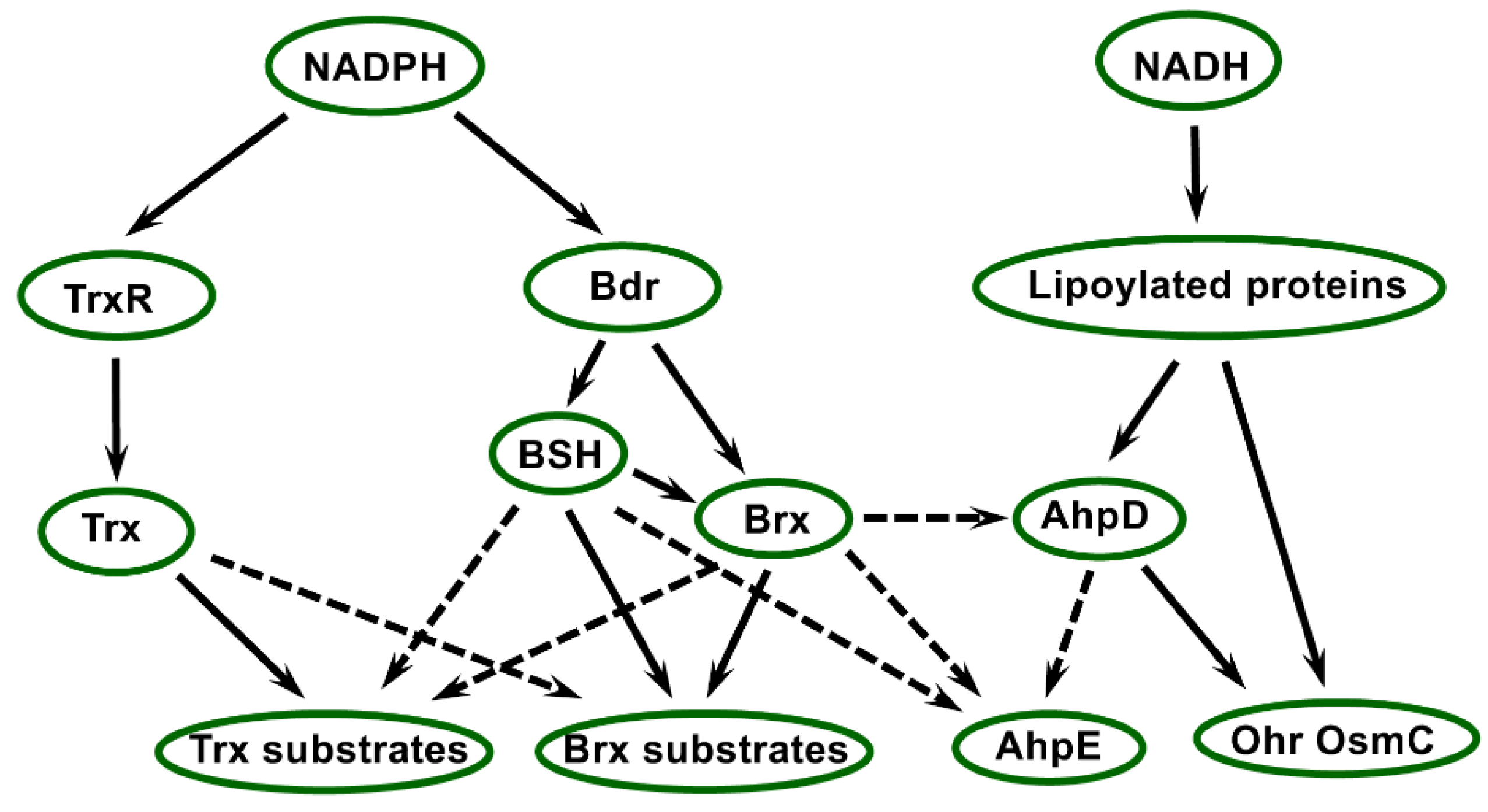{kind=link}
Table 1.
Information of complete genomes of Deinococcus species.
| Species (Abbreviation) | Identified in | Genome Size (Mb) | Replicons (Kb) | Proteins | References |
|---|---|---|---|---|---|
| D. radiodurans (DR) | Canned meat, USA | 3.28 | 4 (2649, 412, 177, 46) | 3167 | [34,35,36] |
| D. deserti (Deide) | Sahara Desert sand, Morocco/Tunisia | 3.86 | 4 (2820, 325, 314, 396) | 3503 | [5,37] |
| D. geothermalis (Dgeo) | Hot spring, Italy | 3.25 | 3 (2467, 574, 206) | 3003 | [38,39] |
| D. gobiensis (DGo) | Gobi Desert sand, China | 4.41 | 7 (3137, 433, 425, 232, 72, 55, 53) | 4140 | [40,41] |
| D. maricopensis (Deima) | Sonoran Desert soil, USA | 3.5 | 1 (3499) | 3242 | [42,43] |
| D. peraridilitoris (Deipe) | Coastal desert soil, Chile | 4.51 | 3 (3882, 557, 75) | 4223 | [44] |
| D. proteolyticus (Deipr) | Lama glama faeces, Japan | 2.89 | 5 (2147, 315, 196, 132, 97) | 2645 | [35,45,46] |
Table 2.
Locus tags of genes encoding thiol reductases and related proteins from seven Deinococcus species.
Table 2.
Locus tags of genes encoding thiol reductases and related proteins from seven Deinococcus species.
| Name/Description | DR | Deide | Dgeo | DGo | Deima | Deipe | Deipr |
|---|---|---|---|---|---|---|---|
| Thioredoxin reductase, thioredoxins, and thioredoxin-like proteins | |||||||
| TrxR | _1982 | _05800 | _1576 _2772 | _CA2339 | _1454 | _0175 _3902 | _0873 |
| TrxA (Trx1) | _0944 | _18600 | _1837 | _CA0861 | _2910 | _3068 _3873 _3901 | _0424 |
| TrxC (Trx2) | _A0164 | _01140 | _2518 | ||||
| Trx-like | _2085 | _06390 | _1508 | _CA2073 | _1013 | _0695 | _0576 |
| Trx-like | _0057 | _13741 | _0729 | _CA0407 | _1186 | _0565 | _1451 |
| Trx-like | _A0072 | _2583 | _PC0201 _PC0211 | _2424 _2732 | |||
| Trx-like | _B0110 | _1776 | _PA0204 | _2190 | |||
| Trx-like | _0948 | _06780 | _1960 | _CA2541 | _2994 | _1792 | |
| Other thiol-based disulfide oxidoreductases, (predicted) cytoplasmic | |||||||
| FrnE | _0659 | _00690 | _2073 | _CA0380 | _0892 | _2202 | _1901 |
| FrnE-like | _3p01230 | _1559 | |||||
| DSBA-like Trx domain-containing protein | _2335 | _22890 | _CA0030 | _0620 | _1019 | ||
| Other thiol-based disulfide oxidoreductases, (predicted) periplasmic or cytoplasmic membrane | |||||||
| DsbA family protein | _2019 _0560 | _06420 | _0747 | _CA1399 | _1134 | _0943 | |
| DsbA family protein | _0753 | _12740 | _0692 | _CA1008 | _1749 | _0493 (and _2421?) | |
| DsbB family | _0754 | _12730 | _0691 | _CA1007 | _1748 | _0492 | |
| DsbD family; CcdA | _1300 | _08350 | _1241 | _CA1639 | _1155 | _0794 | _0982 |
| TlpA-like family; DsbE/CcmG | _0345 _0189 | _08290 | _1248 | _CA2017 _PC0193 | _1148 (and _1627?) | _0801 _4366 | _0892 |
| DsbD family; CcdA | _2p00430 | _0661 | |||||
| TlpA-like family; DsbE/CcmG | _2p00420 | _0660 | |||||
| Thioredoxin-dependent methionine sulfoxide reductases, cytoplasmic | |||||||
| MsrA | _1849 | _10980 | _0843 | _CA1541 | _1788 | _3499 | _1412 |
| MsrB | _1378 | _04050 | _2072 | _CA0919 | _1441 | _4299 | _1900 |
| Potential Mo-dependent methionine sulfoxide reductases, predicted cytoplasmic | |||||||
| Molybdopterin oxidoreductase family protein | _0397 | _18410 | _0402 | _CA0112 | _0538 | _1423 | _1886 |
| Sulfite oxidase family, molybdopterin-binding domain | _0716 | _17540 | _1719 | _CA1115 | _0813 | _2833 | _0695 |
| Mo-dependent methionine sulfoxide reductase system, periplasmic | |||||||
| MsrP | “_2536” (frame-shift) | _20380 | _0877 | _CA2733 | _3114 | _2978 | _1129 |
| MsrQ | _2537 | _20370 | _0878 | _CA2734 | _3115 | _2977 | _1128 |
| Thioredoxin-dependent peroxidases | |||||||
| PRX_BCP | _0846 | _10900 | DgeoAM_1323 a | _CA1364 | _2368 | _0259 | _0703 |
| PRX_BCP | _1209 _1208 | _09051 | _0990 _2729 | _CA1403 | _1714 _0169 | _3580 _3178 | _1557 |
| PRX_BCP | _23291 (partial?) | _CA0314 | |||||
| Alkyl hydroperoxide reductases | |||||||
| AhpE (PRX_AhpE_like) | _2242 | _02430 | _0122 | _CA2657 | _0618 | _1016 | _0175 |
| AhpD-like | _1765 | _13030 _1p00700 | _1446 | _CA1027 | _0298 | _3296 _4199 _3903 _3878 _3900 | _2741 |
| OsmC/Ohr/YhfA family proteins | |||||||
| OsmC | _1538 | _16090 | _0526 | _CA1241 | _0667 | _3743 | |
| Ohr | _1857 | _0446 | _CA0901 _CA1828 | _2331 _0137 | _0225 | _0815 _0816 | |
| YhfA | _1177 | _10790 _21170 | _1268 | _CA1763 | _2343 | _0234 _0648 | _0697 |
| Bacillithiol disulfide reductase, bacilliredoxin | |||||||
| Bdr | _2623 | _23360 | _2331 | _CA0078 | _0670 | _2475 | _1732 |
| Brx (AbxC) | _1832 | _14700 | _1464 | _CA1021 | _1446 | _3166 | _0555 |
a Not initially predicted; corresponds to protein WP_041221145.1.
Table 3.
Active site motifs in Deinococcus Trx and Trx-like proteins.
| Trx1/Trx2 | DR_2085-Type (TRP14-Like) | DR_A0072-Type | DR_B0110-Type | DR_0057-Type (MGP12-Like) | DR_0948-Type | |
|---|---|---|---|---|---|---|
| DR | WCGPC | WCPDC | DCPDC | ACPGC | GCHLC | ECPGC |
| Deide | WCGPC | WCPDC | GCHLC | ECSGC | ||
| Dgeo | WCGPC | WCPDC | NCSSC | NCPAC | GCHLC | ECPGC |
| DGo | WCGPC | WCPDC | QCADC GCASC | ACPDC | GCHLC | ECPGC |
| Deima | WCGPC | WCPDC | GCHLC | ECAGC | ||
| Deipe | WCGPC | WCPDC | GCHLC | |||
| Deipr | WCGPC | WCPDC | DCADC TCPDC | NCPNC | SCKLC | ECAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
de Groot, A.; Blanchard, L.; Rouhier, N.; Rey, P. Thiol Reductases in Deinococcus Bacteria and Roles in Stress Tolerance. Antioxidants 2022, 11, 561. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030561
AMA Style
de Groot A, Blanchard L, Rouhier N, Rey P. Thiol Reductases in Deinococcus Bacteria and Roles in Stress Tolerance. Antioxidants. 2022; 11(3):561. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030561
Chicago/Turabian Stylede Groot, Arjan, Laurence Blanchard, Nicolas Rouhier, and Pascal Rey. 2022. "Thiol Reductases in Deinococcus Bacteria and Roles in Stress Tolerance" Antioxidants 11, no. 3: 561. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030561
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.