
Metabolic Profiling, Chemical Composition, Antioxidant Capacity, and In Vivo Hepato- and Nephroprotective Effects of Sonchus cornutus in Mice Exposed to Cisplatin

 ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Plant Material and Extraction Process
2.2. Metabolic Profiling and Chemical Analysis of S. cornutus
2.2.1. LC-MS/MS Metabolic Profiling
2.2.2. Total Phenolics Content Assay
2.2.3. Total Flavonoids Content Assay
2.3. In Vitro Antioxidant Assays
2.3.1. Determination of Total Antioxidant Capacity
2.3.2. Ferric Reducing Antioxidant Power Assay
2.3.3. DPPH Radical Scavenging Assay
2.4. In Vivo Study of S. cornutus
2.4.1. Drugs and Chemicals
2.4.2. Experimental Animals and Study Protocol
2.5. Biochemical Estimations and Histopathological Examination
2.5.1. Assessment of Liver and Kidney Functions
2.5.2. Assessment of Hepatic and Renal Oxidative Stress and Inflammatory Markers
2.5.3. Quantitative Real-Time—PCR Analysis of Nuclear Factor kappa B (NF-κB), Tumor Necrosis Factor-α (TNF-α), and Nuclear Factor Erythroid 2-related Factor 2 (Nrf2) Expression
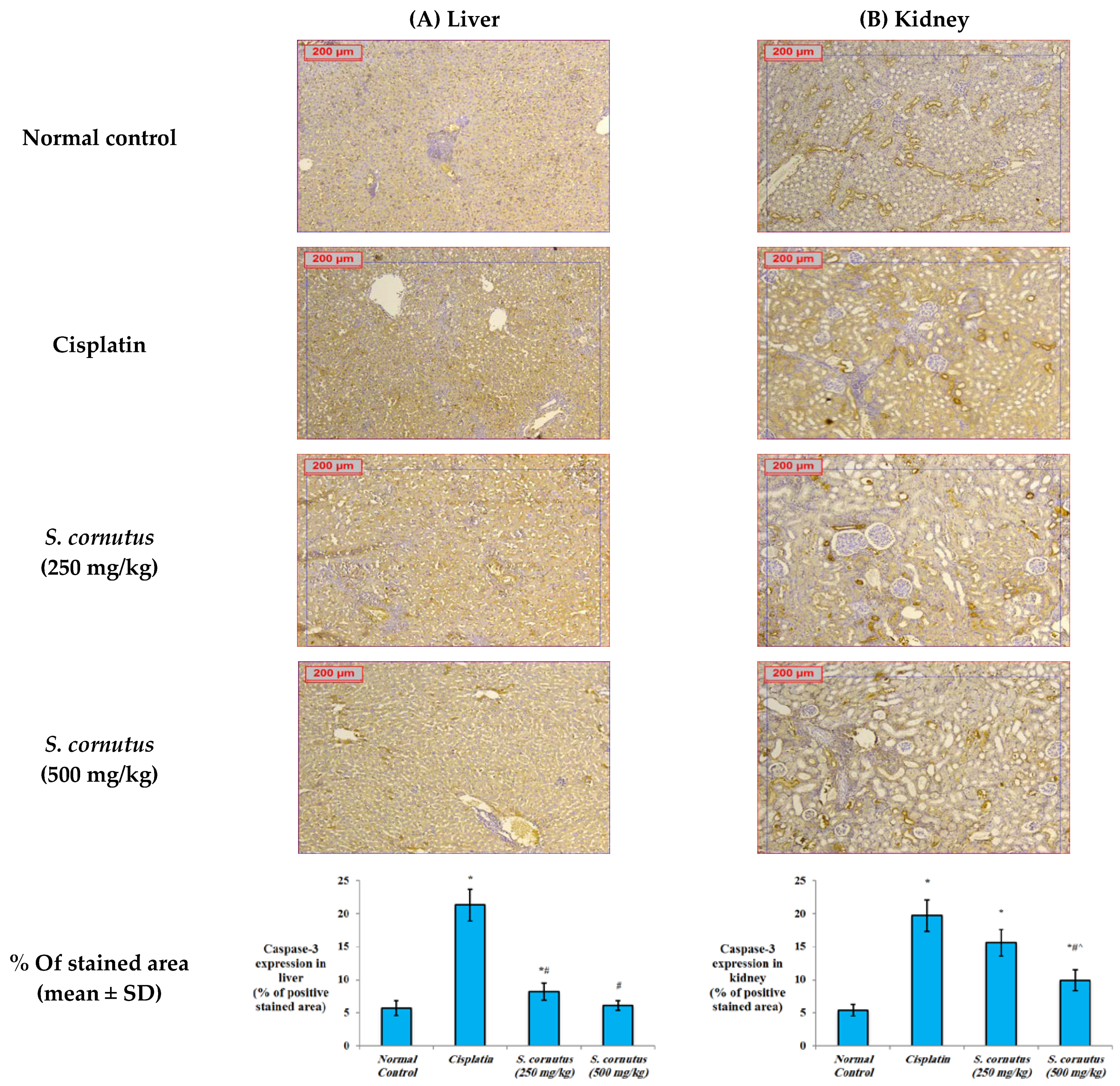
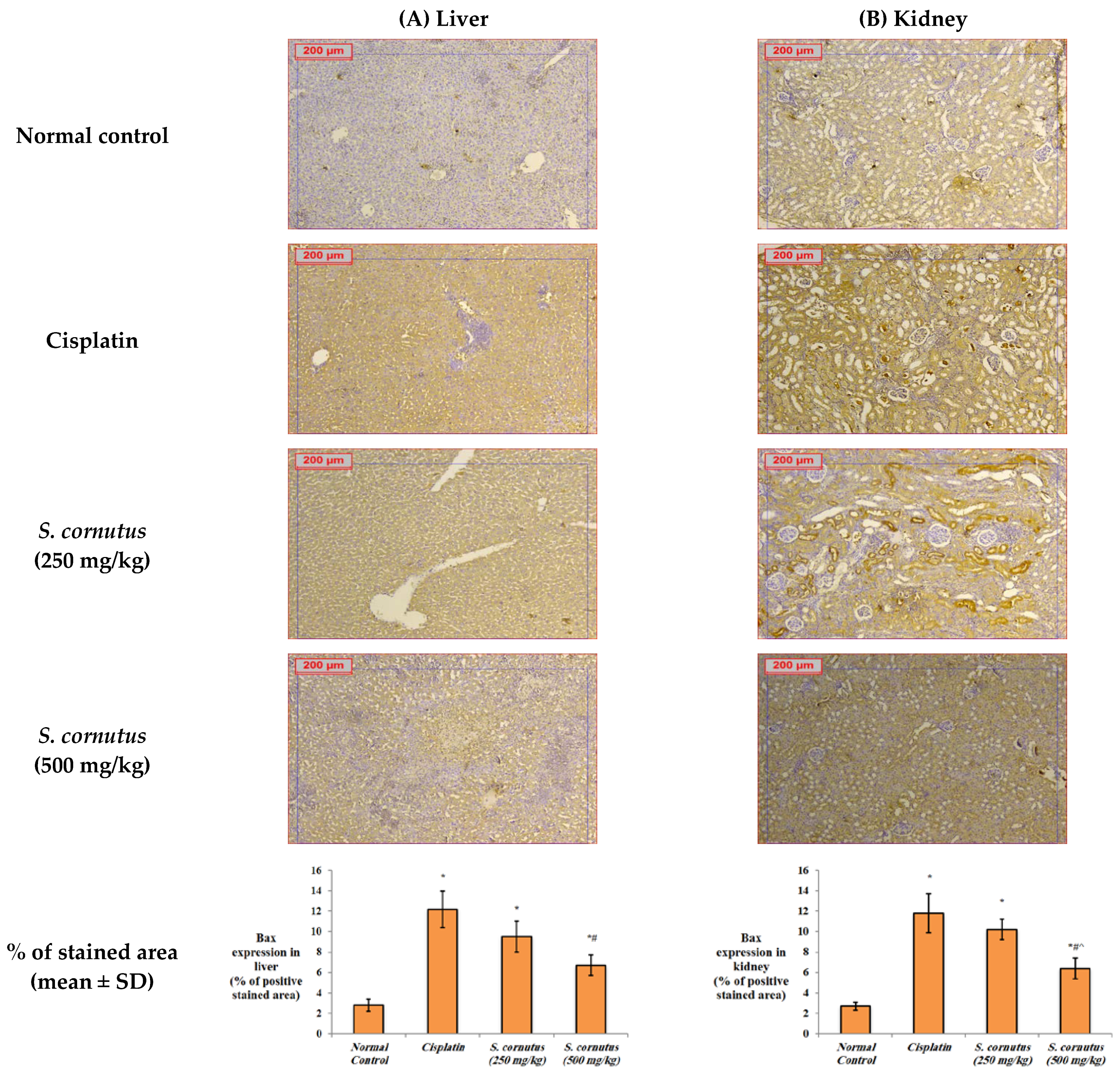
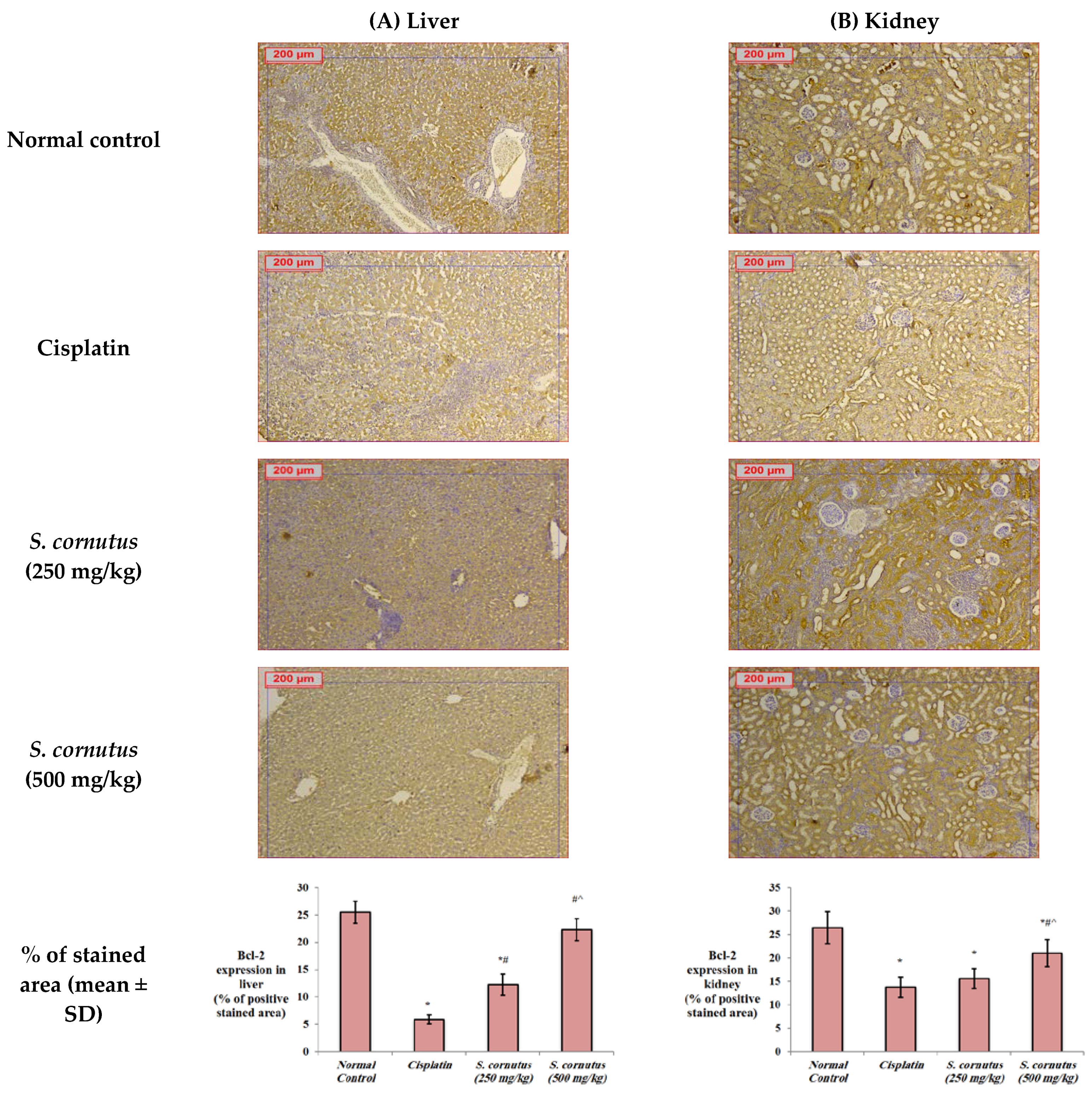
2.5.4. Immunohistochemistry for Detection of Bax, Bcl-2, and Caspase-3
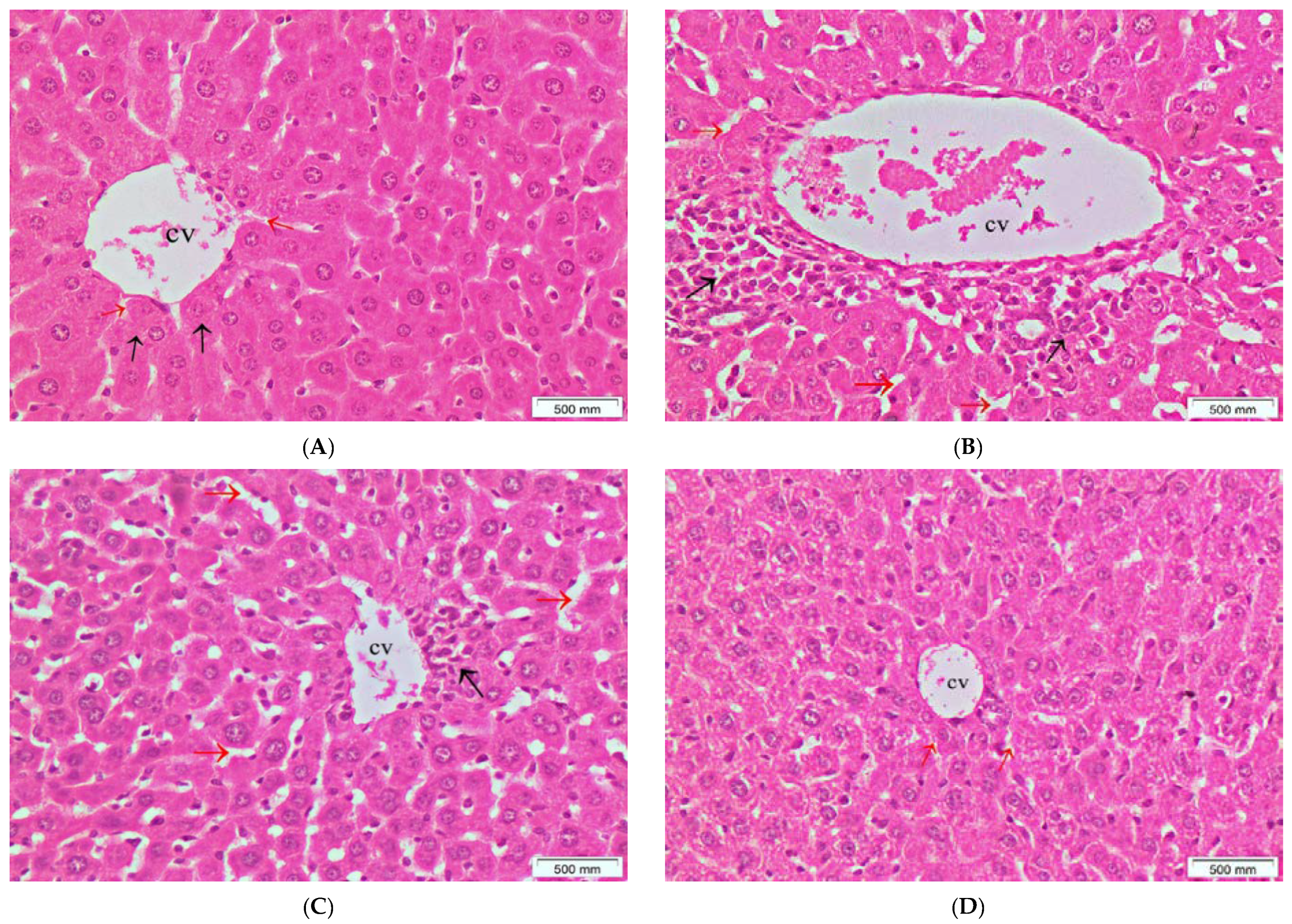
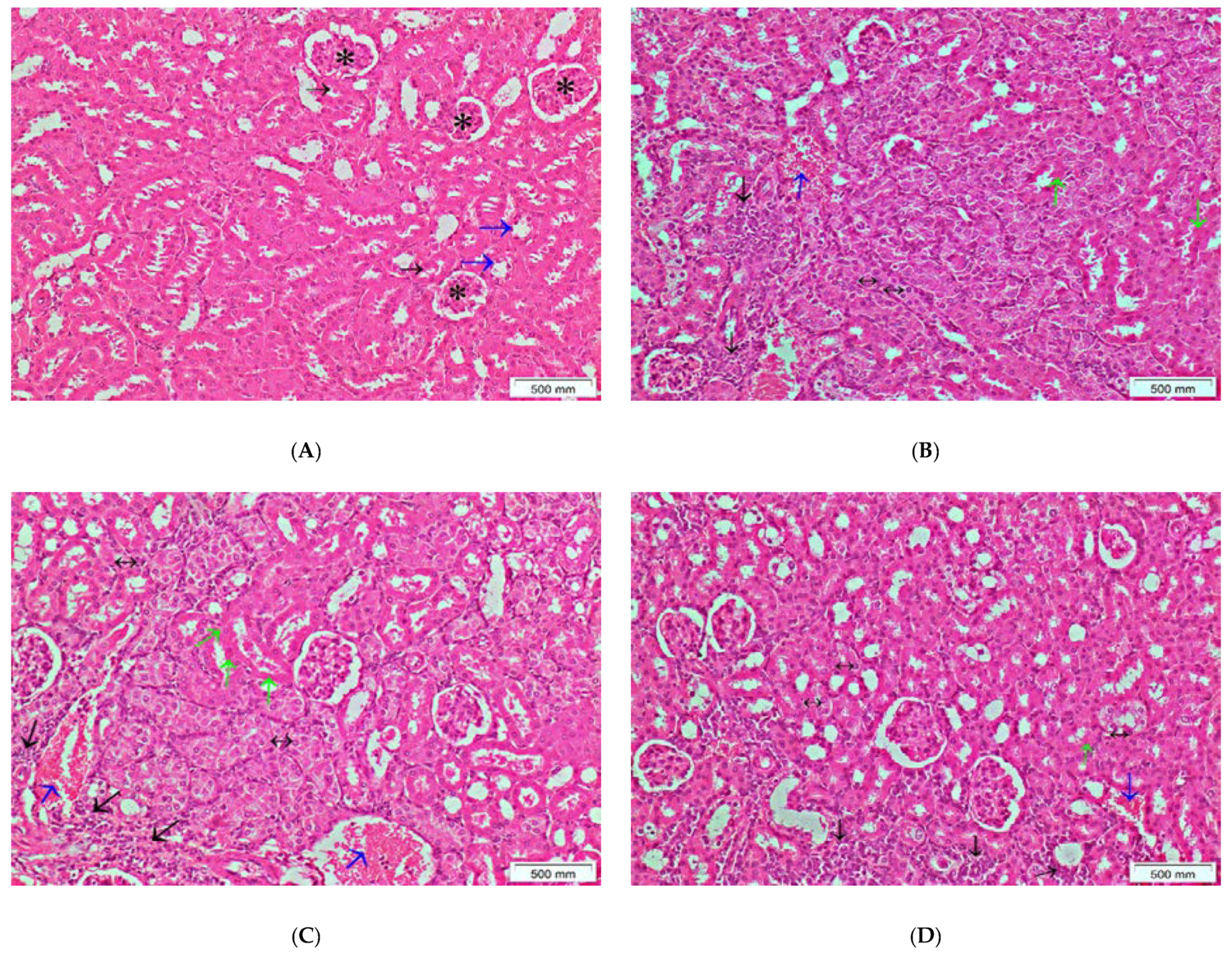
2.5.5. Histopathological Examination
2.5.6. Statistical Analysis
3. Results and Discussion
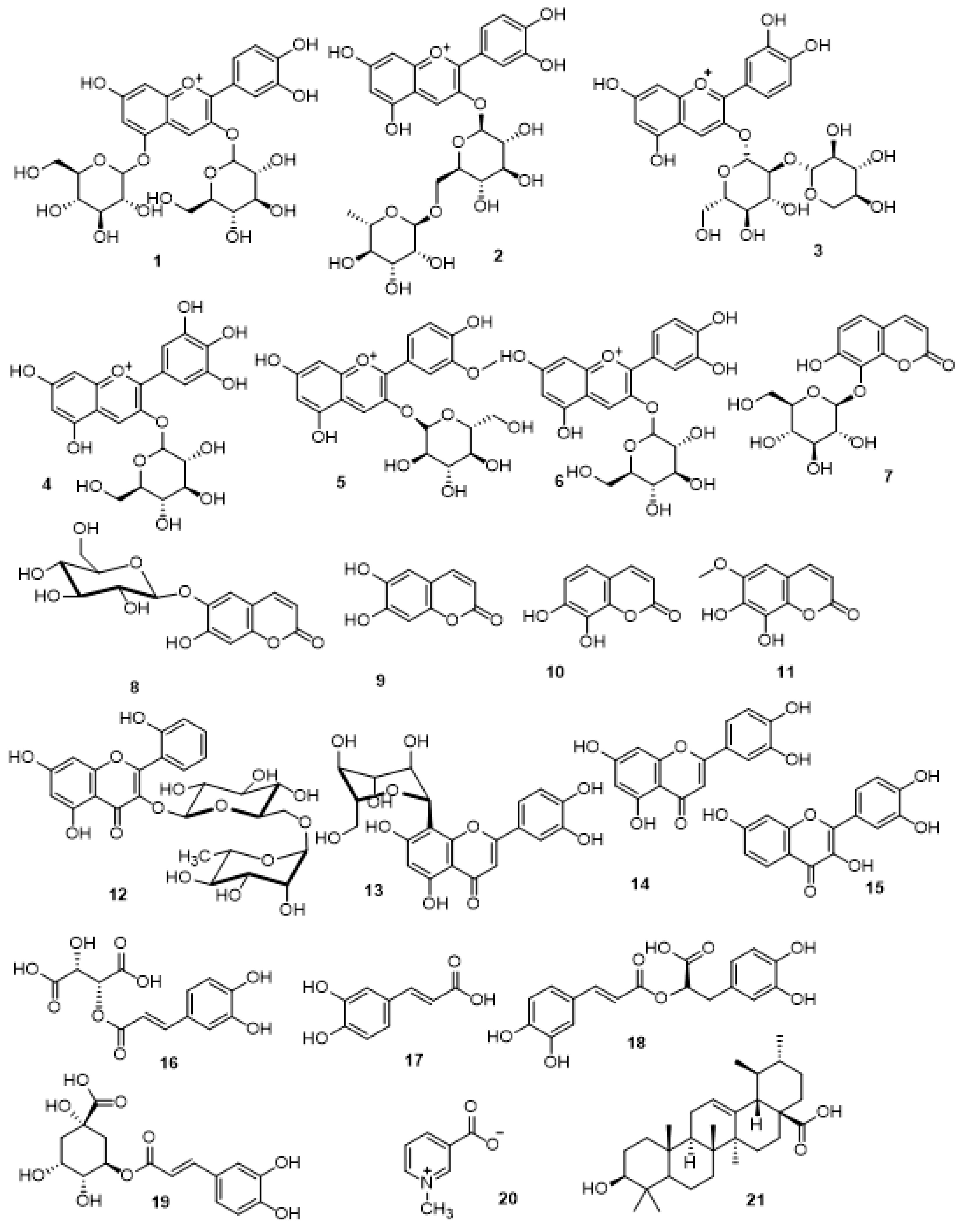
3.1. LC-MS/MS Metabolic Profiling of Crude Extract of S. cornutus
3.2. Quantification of Total Phenolics and Total Flavonoids in S. cornutus
3.3. In Vitro Antioxidant Activity of S. cornutus
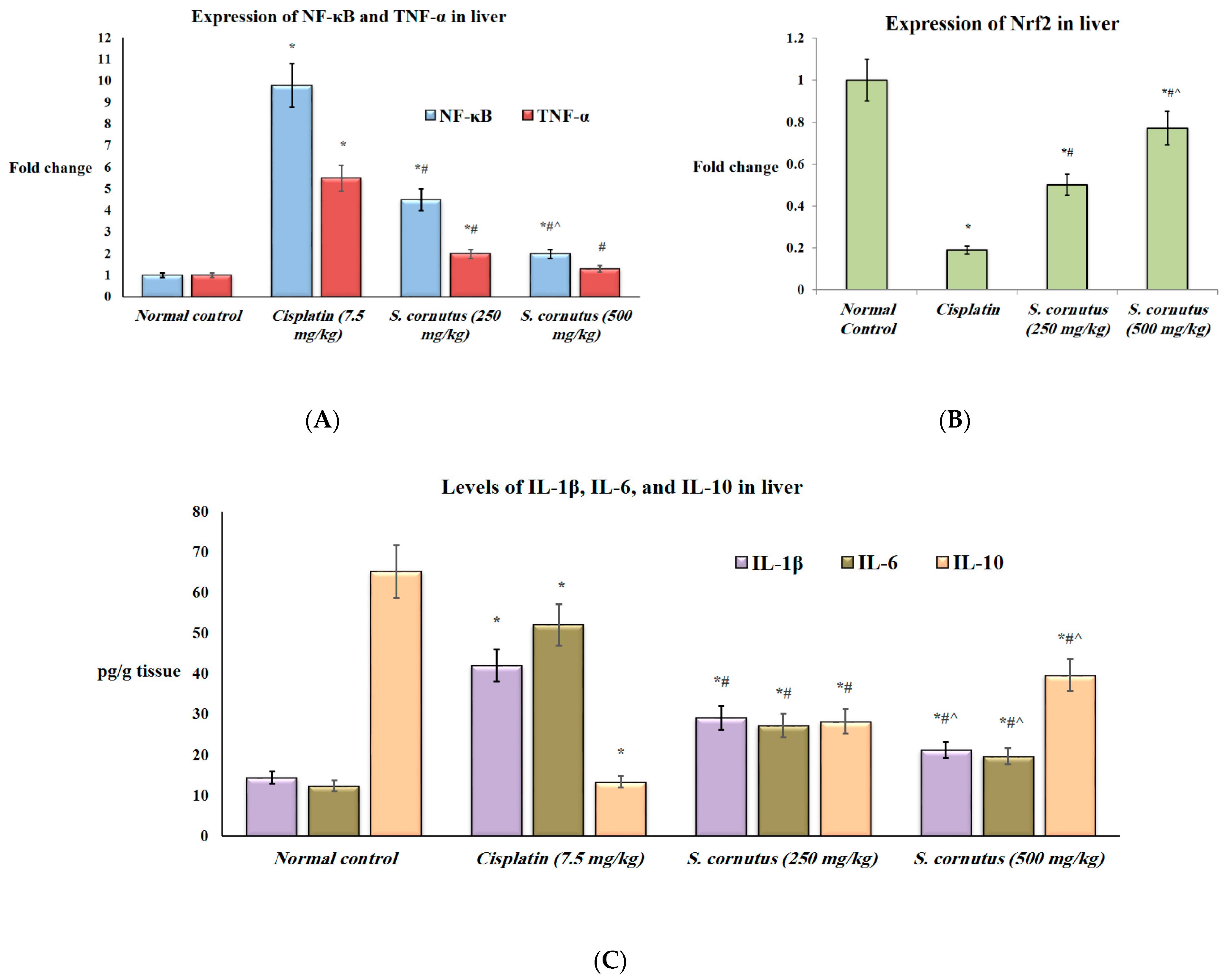
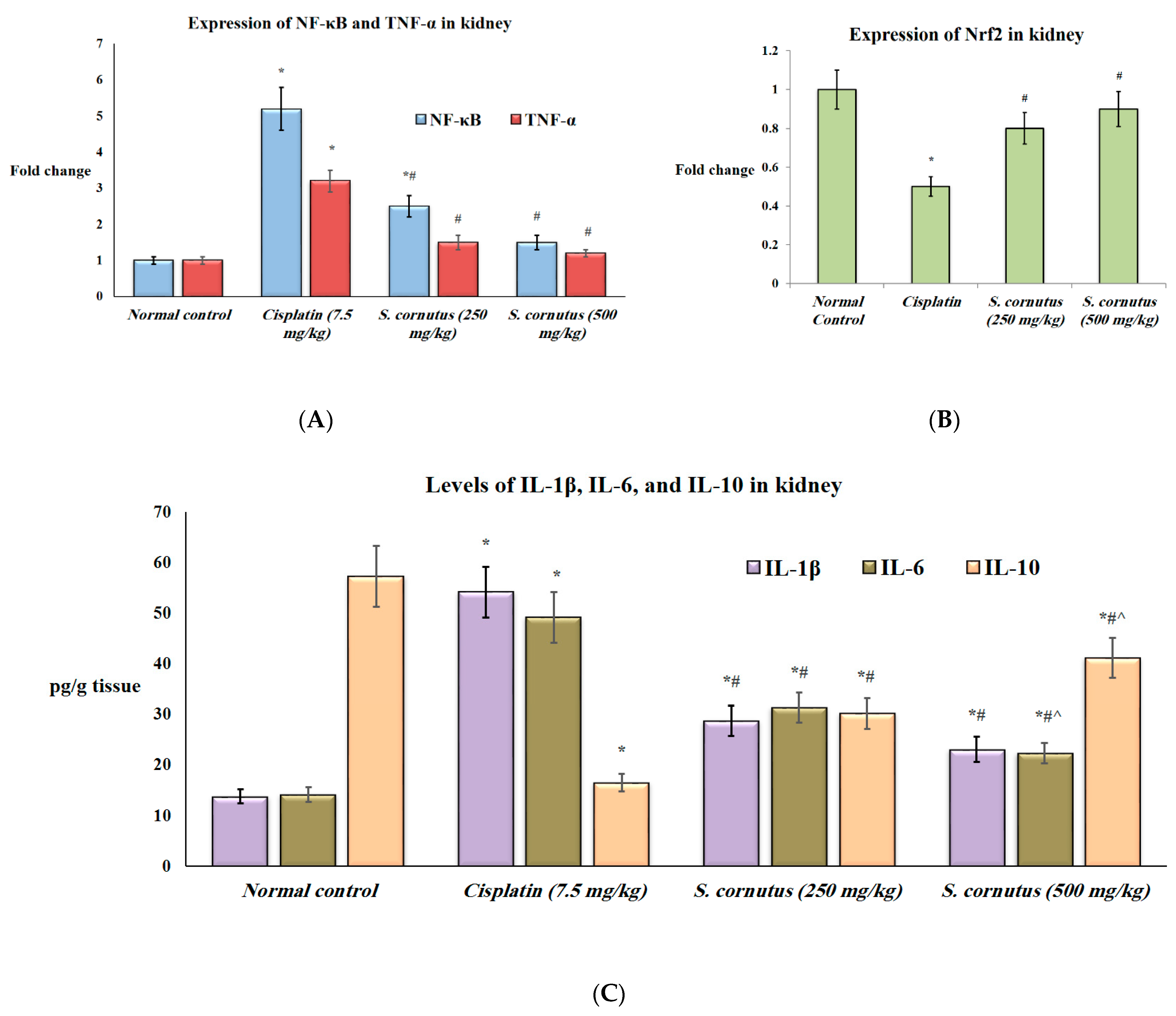
3.4. The Effects of the Crude Extract of S. cornutus against Cisplatin-Induced Hepatic and Renal Toxicity in Experimental Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, S. Cisplatin: The first metal based anticancer drug. Bioorg. Chem. 2019, 88, 102925. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Magotra, R.; Sharma, A.K.; Ahmed, M.; Khajuria, V. Effect of cisplatin on liver of male albino rats. J. Evol. Med. Dent. Sci. 2015, 4, 8993–8998. [Google Scholar] [CrossRef]
- Abdel Moneim, A.E.; Othman, M.S.; Aref, A.M. Azadirachta indica attenuates cisplatin-induced nephrotoxicity and oxidative stress. Biomed. Res. Int. 2014, 2014, 647131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Wang, H.; Wang, C.; Su, J.; Xie, Q.; Xu, L.; Yu, Y.; Liu, S.; Li, S.; Xu, Y. Failure of elevating calcium induces oxidative stress tolerance and imparts cisplatin resistance in ovarian cancer cells. Aging Dis. 2016, 7, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Abouzed, T.; Soliman, M.; Khatab, S.; Elgazzar, A.; Gouda, W.; Dorghamm, D. The protective impacts of Spriulina platensis against cisplatin-induced renal injury through the regulation of oxidative stress, pro-inflammatory cytokines and Bax/Bcl2 expression cascade. Toxicol. Res. 2022, 11, 169–178. [Google Scholar] [CrossRef]
- Ko, J.W.; Lee, I.C.; Park, S.H.; Moon, C.; Kang, S.S.; Kim, S.H.; Kim, J.C. Protective effects of pine bark extract against cisplatin-induced hepatotoxicity and oxidative stress in rats. Lab. Anim. Res. 2014, 30, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Rolnik, A.; Olas, B. The plants of the Asteraceae family as agents in the protection of human health. Int. J. Mol. Sci. 2021, 22, 3009. [Google Scholar] [CrossRef]
- Li, X.-M.; Yang, P.-L. Research progress of Sonchus species. Int. J. Food Prop. 2018, 21, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Farah, H.M.; Hussein, A.R.M.E.; Khalid, H.E.; Osman, H. Assessment of sub chronic toxicity of Sonchus cornutus in rats. AIR 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamed, A.R.; Mehanna, E.T.; Hazem, R.M.; Badr, J.M.; Abo-Elmatty, D.M.; Abdel-Kader, M.S.; Goda, M.S. Plicosepalus acacia extract and its major constituents, methyl gallate and quercetin, potentiate therapeutic angiogenesis in diabetic hind limb ischemia: HPTLC quantification and LC-MS/MS metabolic profiling. Antioxidants 2021, 10, 1701. [Google Scholar] [CrossRef]
- Goda, M.S.; Nafie, M.S.; Awad, B.M.; Abdel-Kader, M.S.; Ibrahim, A.K.; Badr, J.M.; Eltamany, E.E. In vitro and in vivo studies of anti-lung cancer activity of Artemesia judaica L. crude extract combined with LC-MS/MS metabolic profiling, docking simulation and HPLC-DAD quantification. Antioxidants 2022, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Abdelhameed, R.F.A.; Habib, E.S.; Goda, M.S.; Fahim, J.R.; Hassanean, H.A.; Eltamany, E.E.; Ibrahim, A.K.; AboulMagd, A.M.; Fayez, S.; El-kader, A.M.A.; et al. Thalassosterol, a new cytotoxic aromatase inhibitor ergosterol derivative from the Red Sea seagrass Thalassodendron ciliatum. Mar. Drugs 2020, 18, 354. [Google Scholar] [CrossRef] [PubMed]
- Eltamany, E.E.; Elhady, S.S.; Goda, M.S.; Aly, O.M.; Habib, E.S.; Ibrahim, A.K.; Hassanean, H.A.; Abdelmohsen, U.R.; Safo, M.K.; Ahmed, S.A. Chemical composition of the Red Sea green algae Ulva lactuca: Isolation and in silico studies of new anti-COVID-19 ceramides. Metabolites 2021, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Elhady, S.S.; Habib, E.S.; Abdelhameed, R.F.A.; Goda, M.S.; Hazem, R.M.; Mehanna, E.T.; Helal, M.A.; Hosny, K.M.; Diri, R.M.; Hassanean, H.A.; et al. Anticancer effects of new ceramides isolated from the Red Sea red algae Hypnea musciformis in a model of Ehrlich ascites carcinoma: LC-HRMS analysis profile and molecular modeling. Mar. Drugs 2022, 20, 63. [Google Scholar] [CrossRef] [PubMed]
- Hatami, T.; Emami, S.A.; Miraghaee, S.S.; Mojarrab, M. Total Phenolic contents and antioxidant activities of different extracts and fractions from the aerial parts of Artemisia biennis Willd. Iran. J. Pharm. Res. 2014, 13, 551–559. [Google Scholar] [PubMed]
- Kamtekar, S.; Keer, V.; Patil, V. Estimation of phenolic content, flavonoid content, antioxidant and alpha amylase inhibitory activity of marketed polyherbal formulation. J. Appl. Pharm. Sci. 2014, 4, 61–65. [Google Scholar] [CrossRef]
- Kasangana, P.B.; Haddad, P.S.; Stevanovic, T. Study of polyphenol content and antioxidant capacity of Myrianthus arboreus (Cecropiaceae) root bark extracts. Antioxidants 2015, 4, 410–426. [Google Scholar] [CrossRef] [Green Version]
- Batool, R.; Khan, M.R.; Sajid, M.; Ali, S.; Zahra, Z. Estimation of phytochemical constituents and in vitro antioxidant potencies of Brachychiton populneus (Schott & Endl.) R.Br. BMC Chem. 2019, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Eltamany, E.E.; Elhady, S.S.; Ahmed, H.A.; Badr, J.M.; Noor, A.O.; Ahmed, S.A.; Nafie, M.S. Chemical profiling, antioxidant, cytotoxic activities, and molecular docking simulation of Carrichtera annua DC. (Cruciferae). Antioxidants 2020, 9, 1286. [Google Scholar] [CrossRef]
- Leta, B.; Kenenisa, C.; Wondimnew, T.; Sime, T. Evaluation of renoprotective effects of our locally grown green coffee beans against cisplatin-induced nephrotoxicity in Swiss albino mice. Int. J. Nephrol. 2021, 2021, 2805068. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Ren, C.; Mallery, S.R.; Ugalde, C.M.; Pei, P.; Saradhi, U.V.R.V.; Stoner, G.D.; Chan, K.K.; Liu, Z. A rapid and sensitive LC–MS/MS method for quantification of four anthocyanins and its application in a clinical pharmacology study of a bio adhesive black raspberry gel. J. Chromatogr. B 2009, 877, 4027–4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, L.; Coletta, W.; Rapisarda, P.; Donati, M.B.; Rotilio, D. Development and validation of an LC-MS/MS analysis for simultaneous determination of delphinidin-3-glucoside, cyanidin-3-glucoside, and cyanidin-3-(6-malonylglucoside) in human plasma and urine after blood orange juice administration. J. Sep. Sci. 2007, 30, 3127–3136. [Google Scholar] [CrossRef] [PubMed]
- Scorrano, S.; Lazzoi, M.R.; Mergola, L.; Bello, M.P.D.; Sole, R.D.; Vasapollo, G. Anthocyanins profile by Q-TOF LC/MS in Myrtus communis berries from Salento area. Food Anal. Methods 2017, 10, 2404–2411. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; He, B.; Zhang, W.; Liu, L.; Deng, M.; Lü, M.; Qi, X.; Liang, S. Determination of daphnetin and its 8-O-methylated metabolite in rat plasma by UFLC-MS/MS: Application to a pharmacokinetic study. Chromatographia 2022, 85, 333–341. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, W.; Liu, H.; Wu, G.; Song, M.; Yang, B.; Yang, D.; Wang, Q.; Kuang, H. Simultaneous determination of aesculin, aesculetin, fraxetin, fraxin and polydatin in beagle dog plasma by UPLC-ESI-MS/MS and its application in a pharmacokinetic study after oral administration extracts of Ledum palustre L. Molecules 2018, 23, 2285. [Google Scholar] [CrossRef] [Green Version]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. HPLC–DAD–ESI-MS/MS Screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yu, C.; Sun, W.; Liu, G.; Jia, J.; Wang, Y. Simultaneous determination of magnesium lithospermate B, rosmarinic acid, and lithospermic acid in beagle dog serum by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2878–2882. [Google Scholar] [CrossRef]
- Ye, J.-X.; Wei, W.; Quan, L.-H.; Liu, C.-Y.; Chang, Q.; Liao, Y.-H. An LC–MS/MS method for the simultaneous determination of chlorogenic acid, forsythiaside A and baicalin in rat plasma and its application to pharmacokinetic study of Shuang-huang-lian in rats. J. Pharm. Biomed. Anal. 2010, 52, 625–630. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Yibchok-Anun, S.; Charoenlertkul, P.; Wongsasiripat, N. Cyanidin-3-rutinoside alleviates postprandial hyperglycemia and its synergism with acarbose by inhibition of intestinal α-glucosidase. J. Clin. Biochem. Nutr. 2011, 49, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Luo, E.; Liu, X.; Han, B.; Yu, X.; Peng, X. Delphinidin-3-glucoside suppresses breast carcinogenesis by inactivating the Akt/HOTAIR signaling pathway. BMC Cancer 2016, 16, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sari, D.R.T.; Cairns, J.R.K.; Safitri, A.; Fatchiyah, F. Virtual prediction of the delphinidin-3-O-glucoside and peonidin-3-O-glucoside as anti-inflammatory of TNF-α signaling. Acta Inform. Med. 2019, 27, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Y.; Hou, D.-X.; Wu, S. The effects and mechanisms of cyanidin-3-glucoside and its phenolic metabolites in maintaining intestinal integrity. Antioxidants 2019, 8, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmohsen, U.R.; Albohy, A.; Abdulrazik, B.S.; Bayoumi, S.A.L.; Malak, L.G.; Khallaf, I.S.A.; Bringmann, G.; Farag, S.F. Natural coumarins as potential anti-SARS-CoV-2 agents supported by docking analysis. RSC Adv. 2021, 11, 16970–16979. [Google Scholar] [CrossRef]
- Solís-Salas, L.M.; Sierra-Rivera, C.A.; Cobos-Puc, L.E.; Ascacio-Valdés, J.A.; Silva-Belmares, S.Y. Antibacterial potential by rupture membrane and antioxidant capacity of purified phenolic fractions of Persea americana leaf extract. Antibiotics 2021, 10, 508. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Lu, Z.; Zhang, H.; Kang, Y.; Mao, Y.; Wang, H.; Ge, W.; Shi, L. Anti-inflammatory, and protective properties of daphnetin in endotoxin-induced lung injury. J. Agric. Food Chem. 2014, 62, 12315–12325. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Pang, J.; Hu, X.; Nie, T.; Lu, X.; Li, X.; Wang, X.; Lu, Y.; Yang, X.; Jiang, J.; et al. Daphnetin: A novel anti-Helicobacter pylori agent. Int. J. Mol. Sci. 2019, 20, 850. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Wang, Z.; Liu, Y.; Xu, S.; Huang, G.; Xiong, Y.; Zhang, S.; Xu, L.; Deng, Z.; Guan, S. Immunosuppressive activity of daphnetin, one of coumarin derivatives, is mediated through suppression of NF-κB and NFAT signaling pathways in mouse T cells. PLoS ONE 2014, 9, e96502. [Google Scholar] [CrossRef]
- Wang, H.; Zou, D.; Xie, K.; Xie, M. Antibacterial mechanism of fraxetin against Staphylococcus aureus. Mol. Med. Rep. 2014, 10, 2341–2345. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Yang, C.; Park, S.; Song, G.; Lim, W. Fraxetin induces cell death in colon cancer cells via mitochondria dysfunction and enhances therapeutic effects in 5-fluorouracil resistant cells. J. Cell. Biochem. 2022, 123, 469–480. [Google Scholar] [CrossRef]
- Lam, K.Y.; Ling, A.P.K.; Koh, R.Y.; Wong, Y.P.; Say, Y.H. A Review on medicinal properties of orientin. Adv. Pharmacol. Sci. 2016, 2016, 4104595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziyan, L.; Yongmei, Z.; Nan, Z.; Ning, T.; Baolin, L. Evaluation of the anti-inflammatory activity of luteolin in experimental animal models. Planta Med. 2007, 73, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-H.; Lee, S.; Oh, J.-M.; Lee, M.-S.; Yoon, K.-H.; Park, B.H.; Kim, J.W.; Song, H.; Kim, S.-H. Anti-inflammatory activity of fisetin in human mast cells (HMC-1). Pharmacol. Res. 2007, 55, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Silva Rosario, A.C.R.; da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and pharmacological aspects of caffeic acid and its activity in hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Iswandana, R.; Pham, B.T.; van Haaften, W.T.; Luangmonkong, T.; Oosterhuis, D.; Mutsaers, H.A.M.; Olinga, P. Organ- and species-specific biological activity of rosmarinic acid. Toxicol. In Vitro 2016, 32, 261–268. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, Z.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, Y.J.; Chung, S.O.; Park, S.U. Recent studies on ursolic acid and its biological and pharmacological activity. EXCLI J. 2016, 15, 221–228. [Google Scholar] [CrossRef]
- Yin, J.; Kwon, G.J.; Wang, M.H. The antioxidant and cytotoxic activities of Sonchus oleraceus L. extracts. Nutr. Res. Pract. 2007, 1, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A. Evaluation of flavonoids and diverse antioxidant activities of Sonchus arvensis. Chem. Cent. J. 2012, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A. Protective effects of Sonchus asper (L.) Hill, (Asteraceae) against CCl4-induced oxidative stress in the thyroid tissue of rats. BMC Complement. Altern Med. 2012, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Minatel, I.O.; Borges, C.V.; Ferreira, M.I.; Gomez, H.A.; Chen, C.-Y.O.; Lima, G.P.P. Phenolic compounds: Functional properties, impact of processing and bioavailability. In Phenolic Compounds—Biological Activity, 1st ed.; Soto-Hernandez, M., Palma-Tenango, M., del Rosario Gracia Mateos, M., Eds.; InTech: Nappanee, IN, USA, 2017; pp. 1–24. [Google Scholar] [CrossRef] [Green Version]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Ahmed, M. Evaluation of phenolic contents and antioxidant activity of various solvent extracts of Sonchus asper (L.) Hill. Chem. Cent J. 2012, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, N.H.; Hassan, M.O.; Fahmy, G.M.; González, L.; Hammouda, O.; Atteya, A.M. Allelopathic effects of Sonchus oleraceus L. on the germination and seedling growth of crop and weed species. Acta Bot. Bras. 2014, 28, 408–416. [Google Scholar] [CrossRef]
- Fouad, H.; Habib, E.S.; Ahmed, S.A. Phytochemistry and pharmacological effects of plants in genus Sonchus (Asteraceae). Rec. Pharm. Biomed. Sci. 2020, 4, 40–50. [Google Scholar]
- Rahman, M.M.; Islam, M.B.; Biswas, M.; Alam, A.H.M.K. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 621. [Google Scholar] [CrossRef] [Green Version]
- Quintanilha, J.C.F.; de Sousa, V.M.; Visacri, M.B.; Amaral, L.S.; Santos, R.M.M.; Zambrano, T.; Salazar, L.A.; Moriel, P. Involvement of cytochrome P450 in cisplatin treatment: Implications for toxicity. Cancer Chemother. Pharmacol. 2017, 80, 223–233. [Google Scholar] [CrossRef]
- Lu, Y.; Cederbaum, A.I. Cytochrome P450s and alcoholic liver disease. Curr. Pharm. Des. 2018, 24, 1502–1517. [Google Scholar] [CrossRef]
- Lu, Y.; Cederbaum, A.I. Cisplatin-induced hepatotoxicity is enhanced by elevated expression of cytochrome P450 2E1. Toxicol. Sci. 2006, 89, 515–523. [Google Scholar] [CrossRef]
- Marullo, R.; Werner, E.; Degtyareva, N.; Moore, B.; Altavilla, G.; Ramalingam, S.S.; Doetsch, P.W. Cisplatin induces a mitochondrial-ROS response that contributes to cytotoxicity depending on mitochondrial redox status and bioenergetic functions. PLoS ONE 2013, 8, e81162. [Google Scholar] [CrossRef]
- Ahmadipour, A.; Sharififar, F.; Nakhaipour, F.; Samanian, M.; Karami-Mohajeri, S. Hepatoprotective effect of Zataria Multiflora Boisson cisplatin-induced oxidative stress in male rat. J. Med. Life 2015, 8, 275–281. [Google Scholar]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noeman, S.A.; Hamooda, H.E.; Baalash, A.A. Biochemical study of oxidative stress markers in the liver, kidney and heart of high fat diet induced obesity in rats. Diabetol. Metab. Syndr. 2011, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadhukhan, P.; Saha, S.; Dutta, S.; Sil, P.C. Mangiferin ameliorates cisplatin induced acute kidney injury by upregulating Nrf-2 via the activation of PI3K and exhibits synergistic anticancer activity with cisplatin. Front. Pharmacol. 2018, 9, 638. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Gao, W.; Guo, L.; Yang, Y.; Wang, Y.; Gong, H.; Xia, S.; Zhang, B.-K. Dissecting the crosstalk between Nrf2 and NF-κB response pathways in drug-induced toxicity. Front. Cell Dev. Biol. 2021, 9, 809952. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of Nrf2 signaling pathway and its role in inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Cüre, M.C.; Cüre, E.; Kalkan, Y.; Kırbaş, A.; Tümkaya, L.; Yılmaz, A.; Türkyılmaz, A.K.; Şehitoğlu, İ.; Yüce, S. Infliximab modulates cisplatin-induced hepatotoxicity in rats. Balk. Med. J. 2016, 33, 504–511. [Google Scholar] [CrossRef]
- Bilgic, Y.; Akbulut, S.; Aksungur, Z.; Erdemli, M.E.; Ozhan, O.; Parlakpinar, H.; Vardi, N.; Turkoz, Y. Protective effect of dexpanthenol against cisplatin-induced hepatotoxicity. Exp. Ther. Med. 2018, 16, 4049–4057. [Google Scholar] [CrossRef] [Green Version]
- Omar, H.A.; Mohamed, W.R.; Arab, H.H.; Arafa, E.-S.A. Tangeretin alleviates cisplatin-induced acute hepatic injury in rats: Targeting MAPKs and apoptosis. PLoS ONE 2016, 11, e0151649. [Google Scholar] [CrossRef]
- El-Shitany, N.A.; Eid, B. Proanthocyanidin protects against cisplatin-induced oxidative liver damage through inhibition of inflammation and NF-κβ/TLR-4 pathway. Environ. Toxicol. 2017, 32, 1952–1963. [Google Scholar] [CrossRef]
- Aboraya, D.M.; El Baz, A.; Risha, E.F.; Abdelhamid, F.M. Hesperidin ameliorates cisplatin induced hepatotoxicity and attenuates oxidative damage, cell apoptosis, and inflammation in rats. Saudi J. Biol. Sci. 2022, 29, 3157–3166. [Google Scholar] [CrossRef]
- Abd Rashid, N.; Abd Halim, S.A.S.; Teoh, S.L.; Budin, S.B.; Hussan, F.; Adib Ridzuan, N.R.; Abdul Jalil, N.A. The role of natural antioxidants in cisplatin-induced hepatotoxicity. Biomed. Pharmacother. 2021, 144, 112328. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Panichpisal, K.; Kurtzman, N.; Nugent, K. Cisplatin nephrotoxicity: A review. Am. J. Med. Sci. 2007, 334, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, N.A.; Catão, C.S.; Martins, N.M.; Curti, C.; Bianchi, M.L.; Santos, A.C. Cisplatin-induced nephrotoxicity is associated with oxidative stress, redox state unbalance, impairment of energetic metabolism and apoptosis in rat kidney mitochondria. Arch. Toxicol. 2007, 81, 95–504. [Google Scholar] [CrossRef]
- Townsend, D.M.; Deng, M.; Zhang, L.; Lapus, M.G.; Hanigan, M.H. Metabolism of cisplatin to a nephrotoxin in proximal tubule cells. J. Am. Soc. Nephrol. 2003, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres, L.A.; da Cunha, A.D., Jr. Acute nephrotoxicity of cisplatin: Molecular mechanisms. J. Bras. Nefrol. 2013, 35, 332–340. [Google Scholar] [CrossRef]
- Morsy, M.A.; Heeba, G.H. Nebivolol ameliorates cisplatin-induced nephrotoxicity in rats. Basic Clin. Pharmacol. Toxicol. 2016, 118, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.P.; Tadagavadi, R.K.; Ramesh, G.; Reeves, W.B. Mechanisms of cisplatin nephrotoxicity. Toxins 2010, 2, 2490–2518. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Ramesh, G.; Norbury, C.C.; Reeves, W.B. Cisplatin-induced nephrotoxicity is mediated by tumor necrosis factor-alpha produced by renal parenchymal cells. Kidney Int. 2007, 72, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Miao, J.; Liu, W.; Peng, L.; Chen, Y.; Zhong, Q. Formononetin protects against cisplatin-induced acute kidney injury through activation of the PPARα/Nrf2/HO-1/NQO1 pathway. Int. J. Mol. Med. 2021, 47, 511–522. [Google Scholar] [CrossRef]
- McSweeney, K.R.; Gadanec, L.K.; Qaradakhi, T.; Ali, B.A.; Zulli, A.; Apostolopoulos, V. Mechanisms of cisplatin-induced acute kidney injury: Pathological mechanisms, pharmacological interventions, and genetic mitigations. Cancers 2021, 13, 1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xia, W.; Long, G.; Pei, Z.; Li, Y.; Wu, M.; Wang, Q.; Zhang, Y.; Jia, Z.; Chen, H. Isoquercitrin ameliorates cisplatin-induced nephrotoxicity via the inhibition of apoptosis, inflammation, and oxidative stress. Front. Pharmacol. 2020, 11, 599416. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.Y.; Lou, D.Y.; Zhou, L.Q.; Wang, J.C.; Yang, B.; He, Q.J.; Wang, J.J.; Weng, Q.J. Natural products: Potential treatments for cisplatin-induced nephrotoxicity. Acta Pharmacol. Sin. 2021, 42, 1951–1969. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Bokhari, J. Prevention of CCl4-induced nephrotoxicity with Sonchus asper in rat. Food Chem. Toxicol. 2010, 48, 2469–2476. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Shah, N.A. Hepatoprotective activity of Sonchus asper against carbon tetrachloride-induced injuries in male rats: A randomized controlled trial. BMC Complement. Altern. Med. 2012, 12, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-González, L.; Cienfuegos-Pecina, E.; Perales-Quintana, M.M.; Alarcon-Galvan, G.; Muñoz-Espinosa, L.E.; Pérez-Rodríguez, E.; Cordero-Pérez, P. Nephroprotective effect of Sonchus oleraceus extract against kidney injury induced by ischemia-reperfusion in wistar rats. Oxid. Med. Cell Longev. 2018, 2018, 9572803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.; Sharma, A.K.; Kumar, S.; Saxena, A.K.; Pandey, A.K. Bauhinia variegata leaf extracts exhibit considerable antibacterial, antioxidant, and anticancer activities. Biomed. Res. Int. 2013, 2013, 915436. [Google Scholar] [CrossRef] [Green Version]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Santangelo, C.; Varì, R.; Scazzocchio, B.; Di Benedetto, R.; Filesi, C.; Masella, R. Polyphenols, intracellular signaling and inflammation. Ann. Ist. Super. Sanita 2007, 43, 394–405. [Google Scholar]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative stress, and inflammation: What polyphenols can do for us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Soares, E.R.; Monteiro, E.B.; Graziele, F.; Inada, K.O.; Torres, A.G.; Perrone, D.; Soulage, C.O.; Monteiro, M.C.; Resende, A.C.; Moura-Nunes, N.; et al. Up-regulation of Nrf2-antioxidant signaling by Açaí (Euterpe oleracea Mart.) extract prevents oxidative stress in human endothelial cells. J. Funct. Foods 2017, 37, 107–115. [Google Scholar] [CrossRef]
- Domitrović, R.; Potočnjak, I.; Crnčević-Orlić, Z.; Škoda, M. Nephroprotective activities of rosmarinic acid against cisplatin-induced kidney injury in mice. Food Chem. Toxicol. 2014, 66, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Domitrović, R.; Cvijanović, O.; Šušnić, V.; Katalinić, N. Renoprotective mechanisms of chlorogenic acid in cisplatin-induced kidney injury. Toxicology 2014, 324, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Li, J.; Zha, D.; Zhang, L.; Gao, P.; Yao, T.; Wu, X. Chlorogenic acid prevents diabetic nephropathy by inhibiting oxidative stress and inflammation through modulation of the Nrf2/HO-1 and NF-ĸB pathways. Int. Immunopharmacol. 2018, 54, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Alshahrani, S. Mitigation of ILβ-1, ILβ-6, TNF-α, and markers of apoptosis by ursolic acid against cisplatin-induced oxidative stress and nephrotoxicity in rats. Hum. Exp. Toxicol. 2021, 40, S397–S405. [Google Scholar] [CrossRef]
- Owumi, S.E.; Irozuru, C.E.; Arunsi, U.O.; Oyelere, A.K. Caffeic acid protects against DNA damage, oxidative and inflammatory mediated toxicities, and upregulated caspases activation in the hepatorenal system of rats treated with aflatoxin B1. Toxicon 2022, 207, 1–12. [Google Scholar] [CrossRef]
- Pari, L.; Prasath, A. Efficacy of caffeic acid in preventing nickel induced oxidative damage in liver of rats. Chem. Biol. Interact 2008, 173, 77–83. [Google Scholar] [CrossRef]
- Domitrović, R.; Cvijanović, O.; Pugel, E.P.; Zagorac, G.B.; Mahmutefendić, H.; Škoda, M. Luteolin ameliorates cisplatin-induced nephrotoxicity in mice through inhibition of platinum accumulation, inflammation, and apoptosis in the kidney. Toxicology 2013, 310, 115–123. [Google Scholar] [CrossRef]
- Sahu, B.D.; Kalvala, A.K.; Koneru, M.; Mahesh Kumar, J.; Kuncha, M.; Rachamalla, S.S.; Sistla, R. Ameliorative effect of fisetin on cisplatin-induced nephrotoxicity in rats via modulation of NF-κB activation and antioxidant defence. PLoS ONE 2014, 9, e105070. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, Y.; Li, H.; Cao, H.; Liu, B.; Zhang, H.; Shao, F. Daphnetin protects against cisplatin-induced nephrotoxicity by inhibiting inflammatory and oxidative response. Int. Immunopharmacol. 2018, 65, 402–407. [Google Scholar] [CrossRef]
- Ekinci-Akdemi, R.F.N.; Bi Ngöl, Ç.; Yıldırım, S.; Kandemi, R.F.M.; Küçükler, S.; Sağlam, Y.S. The investigation of the effect of fraxin on hepatotoxicity induced by cisplatin in rats. Iran. J. Basic Med. Sci. 2020, 23, 1382–1387. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Polarity Mode | MZmine ID | Ret. Time (min) | Measured m/z | Calculated m/z | Mass Error (ppm) | Adduct | Molecular Formula | MS/MS Spectrum | Deduced Compound | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| I. Anthocyanidin glycosides | |||||||||||
| 1. | Positive | 2385 | 5.43 | 611.1656 | 611.1607 | 8.02 | M + | C27H31O16 + | 611, 449, 287 | Cyanidin-3, 5-di-O-glucopyranoside | [22] |
| 2. | Positive | 2563 | 6.33 | 595.1599 | 595.1657 | −9.75 | M + | C27H31O15 + | 595, 433, 287 | Cyanidin-3-O-rutinoside | [22] |
| 3. | Positive | 2660 | 6.49 | 581.1516 | 581.1501 | 2.58 | M + | C26H29O15 + | 581, 449, 287 | Cyanidin-3-O-(2″-O-β-xylopyranosyl-β-glucopyranoside) | [22] |
| 4. | Positive | 2672 | 6.51 | 465.1022 | 465.1028 | −1.29 | M + | C21H21O12 + | 465, 303 | Delphinidin-3-glucoside | [23] |
| 5. | Positive | 3059 | 7.81 | 463.1282 | 463.1235 | 10.15 | M + | C22H23O11 + | 463, 301 | Peonidin-3-O-glucoside | [24] |
| 6. | Positive | 3103 | 7.99 | 449.1082 | 449.1078 | 0.89 | M + | C21H21O11 + | 449, 287 | Cyanidin-3-O-glucoside | [22] |
| II. Coumarins and their glycosides | |||||||||||
| 7. | Positive | 2122 | 3.73 | 341.0866 | 341.0873 | −2.05 | [M + H] + | C15H16O9 | 341, 179, 133 | Daphnetin-8- glucopyranoside | [25] |
| 8. | Negative | 2357 | 3.79 | 339.0732 | 339.0716 | 4.72 | [M − H] − | C15H16O9 | 339, 177 | Aesculin | [11] |
| 9. | Negative | 2547 | 5.07 | 177.0192 | 177.0188 | 2.26 | [M − H] − | C9H6O4 | 177, 89 | Aesculetin | [26] |
| 10. | Positive | 3035 | 5.75 | 179.0356 | 179. 0344 | 6.70 | [M + H] + | C9H6O4 | 179, 133, 77 | Daphnetin | [25] |
| 11. | Negative | 2757 | 6.01 | 207.0293 | 207.0293 | 0.0 | [M − H] − | C10H8O5 | 207, 192 | Fraxetin | [26] |
| III. Flavonoids and their glycosides | |||||||||||
| 12. | Negative | 2910 | 6.52 | 593.1481 | 593.1506 | −4.21 | [M − H] − | C27H29O15 | 593, 285 | Datiscin | [10] |
| 13. | Negative | 3045 | 6.85 | 447.0922 | 447.0927 | −1.12 | [M − H] − | C21H20O11 | 447, 327 | Orientin | [11] |
| 14. | Negative | 3564 | 9.43 | 285.0404 | 285.0399 | 1.75 | [M − H] − | C15H10O6 | 285, 133 | Luteolin | [11] |
| 15. | Positive | 3505 | 9.72 | 287.0574 | 287.0556 | 6.27 | [M + H] + | C15H10O6 | 287, 269, 241, 213, 149, 137 | Fisetin | [11] |
| IV. Phenolic derivatives | |||||||||||
| 16. | Negative | 383 | 1.14 | 311.0427 | 311.0403 | 7.72 | [M − H] − | C13H12O9 | 311, 133 | Caftaric acid | [27] |
| 17. | Negative | 973 | 1.24 | 179.0355 | 179.0344 | 6.14 | [M − H] − | C9H8O4 | 179, 135, 134 | Caffeic acid | [11] |
| 18. | Negative | 2496 | 4.62 | 359.0758 | 359.0767 | −2.51 | [M − H] − | C18H16O8 | 359, 161 | Rosmarinic acid | [28] |
| 19. | Negative | 2536 | 4.88 | 353.0860 | 353.0873 | −3.68 | [M − H] − | C16H18O9 | 353, 191 | Chlorogenic acid | [29] |
| V. Other chemical classes | |||||||||||
| 20. | Positive | 1680 | 1.39 | 138.0546 | 138.0555 | −6.52 | [M + H] + | C7H7NO2 | 138, 94 | Trigonelline | [10] |
| 21. | Negative | 4434 | 22.81 | 455.3539 | 455.3525 | 3.07 | [M − H] − | C30H48O3 | 455 | Ursolic acid | [11] |
| Crude Extract | Total Phenolics (mg GAE/gm) | Total Flavonoids (mg QE/gm) |
|---|---|---|
| S. cornutus | 206.28 ± 14.64 | 45.56 ± 1.78 |
| Samples | Total Antioxidant Capacity (mg GAE/gm) | Ferric Reducing Power (mM Fe+2/gm) | IC50 of DPPH Scavenging Activity (µg/mL) |
|---|---|---|---|
| S. cornutus crude extract | 49.06 ± 3.62 | 1.92 ± 0.71 | 16.10 ± 2.14 |
| Ascorbic acid | 69.32 ± 4.51 | 3.14 ± 0.82 | 10.64 ± 0.82 |
| Parameter | Normal Control | Cisplatin (7.5 mg/kg) | S. cornutus (250 mg/kg) | S. cornutus (500 mg/kg) |
|---|---|---|---|---|
| ALT (U/L) | 51.55 ± 5.60 | 98.11 ± 10.10 * | 76.13 ± 8.20 *# | 63.12 ± 6.22 *#^ |
| AST (U/L) | 45.93 ± 5.50 | 92.77 ± 9.98 * | 77.37 ± 8.23 *# | 60.21 ± 7.05 *#^ |
| ALP (U/L) | 91.54 ± 8.73 | 280.38 ± 21.39 * | 210.20 ± 18.28 *# | 140.28 ± 15.30 *#^ |
| Creatinine (mg/dL) | 0.16 ± 0.03 | 0.44 ± 0.09 * | 0.35 ± 0.07 *# | 0.22 ± 0.05 #^ |
| BUN (mg/dL) | 45.34 ± 3.18 | 79.90 ± 9.23 * | 63.36 ± 7.22 *# | 51.29 ± 5.82 #^ |
| Parameter | Organ | Normal Control | Cisplatin (7.5 mg/kg) | S. cornutus (250 mg/kg) | S. cornutus (500 mg/kg) |
|---|---|---|---|---|---|
| MDA (nmol/g tissue) | Liver | 40.4 ± 5.1 | 287.7 ± 32.5 * | 182.7 ± 21.3 *# | 136.3 ± 15.4 *#^ |
| Kidney | 122.4 ± 12.3 | 558.3 ± 63.3 * | 304.2 ± 32.3 *# | 162.4 ± 18.3 *#^ | |
| ROS (U/g tissue) | Liver | 17.2 ± 2.0 | 56.1 ± 5.9 * | 29.8 ± 3.0 *# | 23.4 ± 2.5 *#^ |
| Kidney | 15.9 ± 1.8 | 55.3 ± 5.8 * | 33.1 ± 3.4 *# | 25.8 ± 2.8 *#^ | |
| GSH (ng/mg tissue) | Liver | 70.2 ± 8.1 | 21.3 ± 2.1 * | 38.9 ± 4.2 *# | 54.6 ± 6.3 *#^ |
| Kidney | 59.2 ± 6.5 | 17.8 ± 1.9 * | 28.7 ± 3.1 *# | 41.2 ± 4.2 *#^ | |
| GSSG (ng/mg tissue) | Liver | 19.7 ± 2.2 | 67.3 ± 7.5 * | 36.5 ± 3.8 *# | 28.7 ± 3.2 *#^ |
| Kidney | 20.1 ± 2.2 | 63.2 ± 6.8 * | 38.4 ± 4.0 *# | 27.6 ± 2.8 *#^ | |
| GSH/GSSG ratio | Liver | 3.6 ± 0.5 | 0.3 ± 0.1 * | 1.1 ± 0.1 *# | 1.9 ± 0.2 *#^ |
| Kidney | 2.9 ± 0.4 | 0.3 ± 0.1 * | 0.7 ± 0.1 *# | 1.5 ± 0.2 *#^ | |
| SOD (U/g tissue) | Liver | 698.9 ± 73.3 | 270.4 ± 30.2 * | 398.2 ± 42.1 *# | 470.7 ± 45.3 *# |
| Kidney | 187.3 ± 2.2 | 117.2 ± 12.3 * | 125.2 ± 14.2 * | 150.3 ± 16.2 *# | |
| Catalase (U/g tissue) | Liver | 16.8 ± 2.3 | 5.1 ± 0.7 * | 9.2 ± 1.1 *# | 13.2 ± 1.5#^ |
| Kidney | 18.9 ± 2.5 | 8.1 ± 0.9 * | 10.9 ± 1.2 * | 15.3 ± 1.7#^ | |
| NO (µmol/g tissue) | Liver | 11.3 ± 1.6 | 44.5 ± 5.4 * | 23.5 ± 2.8 *# | 18.6 ± 2.0 *# |
| Kidney | 8.6 ± 0.9 | 27.2 ± 3.2 * | 21.3 ± 2.6 * | 14.6 ± 1.5 *#^ | |
| iNOS (ng/g tissue) | Liver | 3.5 ± 0.4 | 16.5 ± 2.0 * | 10.4 ± 1.4 *# | 7.6 ± 0.9 *#^ |
| Kidney | 3.4 ± 0.5 | 11.2 ± 1.3 * | 7.7 ± 0.9 *# | 5.9 ± 0.7 *# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elhady, S.S.; Abdelhameed, R.F.A.; Mehanna, E.T.; Wahba, A.S.; Elfaky, M.A.; Koshak, A.E.; Noor, A.O.; Bogari, H.A.; Malatani, R.T.; Goda, M.S. Metabolic Profiling, Chemical Composition, Antioxidant Capacity, and In Vivo Hepato- and Nephroprotective Effects of Sonchus cornutus in Mice Exposed to Cisplatin. Antioxidants 2022, 11, 819. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050819
Elhady SS, Abdelhameed RFA, Mehanna ET, Wahba AS, Elfaky MA, Koshak AE, Noor AO, Bogari HA, Malatani RT, Goda MS. Metabolic Profiling, Chemical Composition, Antioxidant Capacity, and In Vivo Hepato- and Nephroprotective Effects of Sonchus cornutus in Mice Exposed to Cisplatin. Antioxidants. 2022; 11(5):819. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050819
Chicago/Turabian StyleElhady, Sameh S., Reda F. A. Abdelhameed, Eman T. Mehanna, Alaa Samir Wahba, Mahmoud A. Elfaky, Abdulrahman E. Koshak, Ahmad O. Noor, Hanin A. Bogari, Rania T. Malatani, and Marwa S. Goda. 2022. "Metabolic Profiling, Chemical Composition, Antioxidant Capacity, and In Vivo Hepato- and Nephroprotective Effects of Sonchus cornutus in Mice Exposed to Cisplatin" Antioxidants 11, no. 5: 819. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050819