
Feed Restriction Alleviates Chronic Thermal Stress-Induced Liver Oxidation and Damages via Reducing Lipid Accumulation in Channel Catfish (Ictalurus punctatus)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Channel Catfish and Feeding Trial
2.3. Sample Collection
2.4. Determination of Growth Parameters
2.5. Biochemical and Antioxidative Parameters
2.6. Detection of Lipid Metabolism-Related Enzyme Activity in Liver
2.7. Apoptosis Detection
2.8. Tissue Total RNA Extraction and RT-qPCR Analysis
2.9. Protein Extraction and Western Blot Analysis
2.10. Hematoxylin and Eosin (H&E) Staining
2.11. Statistical Analysis
3. Results
3.1. Feed Restriction (2.5% BW) Suppressed the Thermal Stress Response of Liver without Causing Growth Retardation
3.2. Feed Restriction Reduced Liver Lipid Accumulation and β-Oxidation
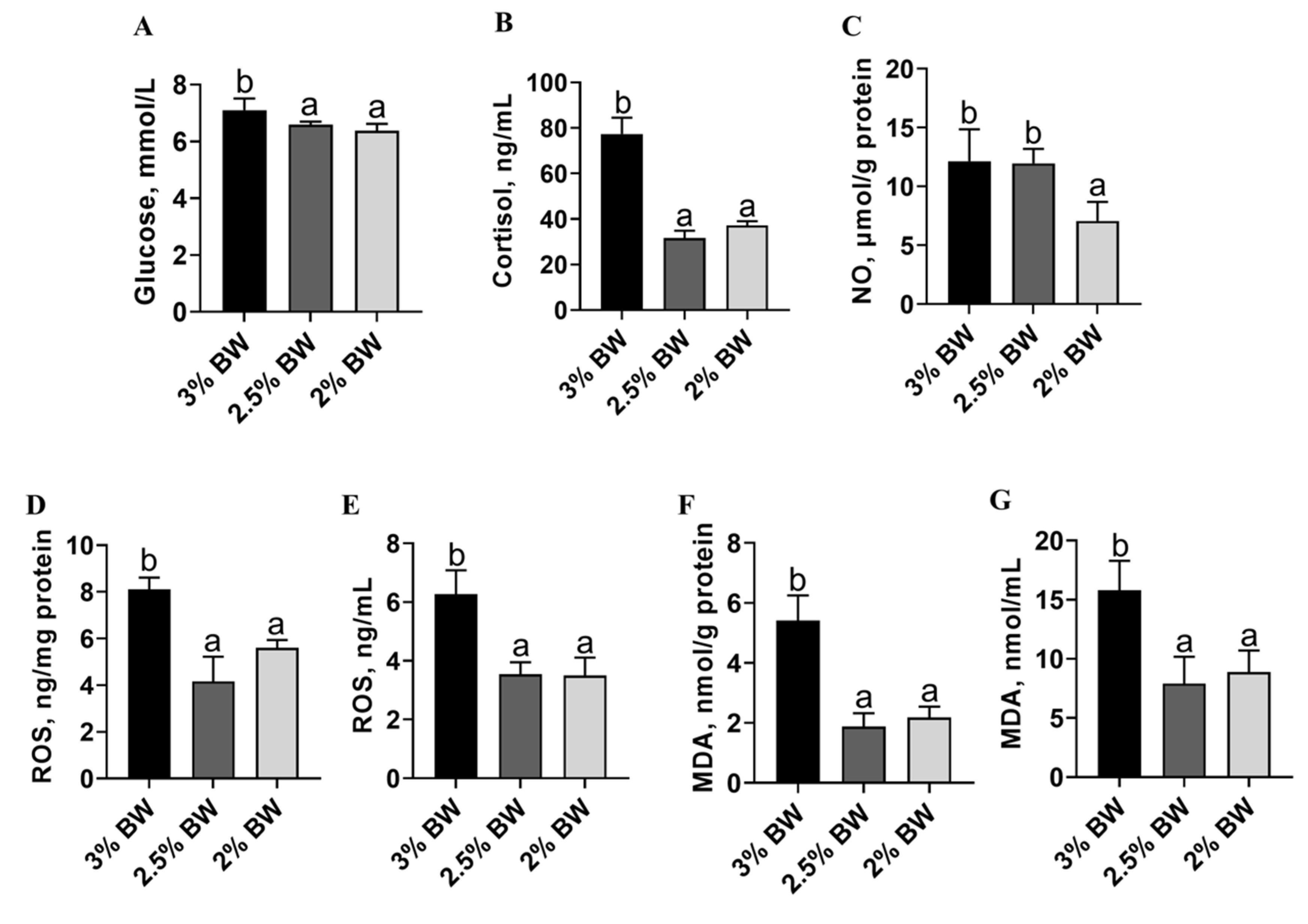
3.3. Feed Restriction Enhanced Liver Antioxidant Capacity and Improved Thermal Stress State
3.4. Feed Restriction Mitigated the Heat-Induced ER Stress and Apoptosis in Liver
3.5. Feed Restriction Alleviated Heat-Induced Liver Inflammation and Damages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahanty, A.; Mohanty, S.; Mohanty, B.P. Dietary supplementation of curcumin augments heat stress tolerance through upregulation of nrf-2-mediated antioxidative enzymes and hsps in Puntius sophore. Fish Physiol. Biochem. 2017, 43, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.; Pan, C.; Liu, E.; Zhao, X.; Ling, Q. Alterations to transcriptomic profile, histopathology, and oxidative stress in liver of pikeperch (Sander lucioperca) under heat stress. Fish Shellfish Immunol. 2019, 95, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Narum, S.R.; Campbell, N.R.; Meyer, K.A.; Miller, M.R.; Hardy, R.W. Thermal adaptation and acclimation of ectotherms from differing aquatic climates. Mol. Ecol. 2013, 22, 3090–3097. [Google Scholar] [CrossRef] [PubMed]
- Gradil, A.M.; Wright, G.M.; Speare, D.J.; Wadowska, D.W.; Purcell, S.; Fast, M.D. The effects of temperature and body size on immunological development and responsiveness in juvenile shortnose sturgeon (Acipenser brevirostrum). Fish Shellfish Immunol. 2014, 40, 545–555. [Google Scholar] [CrossRef]
- Roychowdhury, P.; Aftabuddin, M.; Pati, M.K. Thermal stress-induced oxidative damages in the liver and associated death in fish, Labeo rohita. Fish Physiol. Biochem. 2021, 47, 21–32. [Google Scholar] [CrossRef]
- Forgati, M.; Kandalski, P.K.; Herrerias, T.; Zaleski, T.; Machado, C.; Souza, M.; Donatti, L. Effects of heat stress on the renal and branchial carbohydrate metabolism and antioxidant system of Antarctic fish. J. Comp. Physiol. B 2017, 187, 1137–1154. [Google Scholar] [CrossRef]
- Dahlke, F.T.; Wohlrab, S.; Butzin, M.; Pörtner, H.O. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science 2020, 369, 65–70. [Google Scholar] [CrossRef]
- Spadaro, O.; Youm, Y.; Shchukina, I.; Ryu, S.; Sidorov, S.; Ravussin, A.; Nguyen, K.; Aladyeva, E.; Predeus, A.N.; Smith, S.R.; et al. Caloric restriction in humans reveals immunometabolic regulators of health span. Science 2022, 375, 671–677. [Google Scholar] [CrossRef]
- Anderson, R.M.; Le Couteur, D.G.; de Cabo, R. Caloric restriction research: New perspectives on the biology of aging. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 73, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Xia, E.; Rao, G.; Van Remmen, H.; Heydari, A.R.; Richardson, A. Activities of antioxidant enzymes in various tissues of male Fischer 344 rats are altered by food restriction. J. Nutr. 1995, 125, 195–201. [Google Scholar]
- Gültekin, F.; Nazıroğlu, M.; Savaş, H.B.; Çiğ, B. Calorie restriction protects against apoptosis, mitochondrial oxidative stress and increased calcium signaling through inhibition of TRPV1 channel in the hippocampus and dorsal root ganglion of rats. Metab. Brain Dis. 2018, 33, 1761–1774. [Google Scholar] [CrossRef]
- Venturini, P.R.; Thomazini, B.F.; Oliveira, C.A.; Alves, A.A.; Camargo, T.F.; Domingues, C.; Barbosa-Sampaio, H.; do Amaral, M. Vitamin E supplementation and caloric restriction promotes regulation of insulin secretion and glycemic homeostasis by different mechanisms in rats. Biochem. Cell Biol. 2018, 96, 777–785. [Google Scholar] [CrossRef]
- Li, J.; Xue, B.; Cheng, X.; Hu, J.; Hu, J.; Tian, J.; Li, F.; Yu, X.; Li, B. TiO2 NPs alleviates high-temperature induced oxidative stress in silkworms. J. Econ. Entomol. 2018, 111, 879–884. [Google Scholar] [CrossRef]
- Ma, J.; Hampl, J.S.; Betts, N.M. Antioxidant intakes and smoking status: Data from the continuing survey of food intakes by individuals 1994–1996. Am. J. Clin. Nutr. 2000, 71, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Wang, W.; Li, J.; Yang, Z. Growth, oxidative stress responses, and gene transcription of juvenile bighead carp (Hypophthalmichthys nobilis) under chronic-term exposure of ammonia. Environ. Toxicol. Chem. 2014, 33, 1726–1731. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar]
- Long, S.; Dong, X.; Tan, B.; Zhang, S.; Xie, S.; Yang, Q.; Chi, S.; Liu, H.; Deng, J.; Yang, Y.; et al. Growth performance, antioxidant ability, biochemical index in serum, liver histology and hepatic metabolomics analysis of juvenile hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus) fed with oxidized fish oil. Aquaculture 2021, 545, 737261. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Xie, J.; Ge, X.; Zhou, Q.; Sun, C.; Zhang, H.; Shan, F.; Yang, Z. Oxidized fish oil injury stress in Megalobrama amblycephala: Evaluated by growth, intestinal physiology, and transcriptome-based PI3K-Akt/NF-κB/TCR inflammatory signaling. Fish Shellfish Immunol. 2018, 81, 446–455. [Google Scholar] [CrossRef]
- Biller, J.D.; Takahashi, L.S. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. An. Acad. Bras. Cienc. 2018, 90, 3403–3414. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhao, C.Y.; Guan, J.F.; Liu, X.C.; Li, X.F.; Xie, D.Z.; Xu, C. High-carbohydrate diet alleviates the oxidative stress, inflammation and apoptosis of Megalobrama amblycephala following dietary exposure to silver nanoparticles. Antioxidants 2021, 10, 1343. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, L.; Jiang, W.; Liu, Y.; Jiang, J.; Kuang, S.; Li, S.; Tang, L.; Tang, W.; Zhou, X.; et al. Dietary vitamin A improved the flesh quality of grass carp (Ctenopharyngodon idella) in relation to the enhanced antioxidant capacity through Nrf2/Keap 1a signaling pathway. Antioxidants 2022, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Hu, Y.; Wang, Z.; Zhou, J.; Zhang, J.; Zhong, H.; Fu, G.; Zhong, L. The protective effect of taurine on oxidized fish-oil-induced liver oxidative stress and intestinal barrier-function impairment in juvenile Ictalurus punctatus. Antioxidants 2021, 10, 1690. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Q.; Wang, X.; Wang, X.Y.; Li, H.X.; Xu, L.; Huang, B.; Meng, X.S.; Zhang, T.; Chen, H.B.; Xing, R.; et al. Effects of different feeding frequencies on the growth, plasma biochemical parameters, stress status, and gastric evacuation of juvenile tiger puffer fish (Takifugu rubripes). Aquaculture 2022, 548, 737718. [Google Scholar] [CrossRef]
- Paula, T.G.; Zanella, B.; Fantinatti, B.; Moraes, L.N.; Duran, B.; Oliveira, C.B.; Salomão, R.; Silva, R.; Padovani, C.R.; Santos, V.; et al. Food restriction increase the expression of mTORC1 complex genes in the skeletal muscle of juvenile pacu (Piaractus mesopotamicus). PLoS ONE 2017, 12, e0177679. [Google Scholar] [CrossRef]
- Gong, Y.; Chen, W.; Han, D.; Zhu, X.; Yang, Y.; Jin, J.; Liu, H.; Xie, S. Effects of food restriction on growth, body composition and gene expression related in regulation of lipid metabolism and food intake in grass carp. Aquaculture 2017, 469, 28–35. [Google Scholar] [CrossRef]
- Xu, C.; Liu, W.B.; Remø, S.C.; Wang, B.K.; Shi, H.J.; Zhang, L.; Liu, J.D.; Li, X.F. Feeding restriction alleviates high carbohydrate diet-induced oxidative stress and inflammation of Megalobrama amblycephala by activating the AMPK-SIRT1 pathway. Fish Shellfish Immunol. 2019, 92, 637–648. [Google Scholar] [CrossRef]
- Fishery and Aquaculture Statistics 2018/FAO Annuaire; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020.
- RuthEllen, C.; Vicki, S.B.; Carlos, E. Effects of dietary lipid on the hematology of channel catfish, Ictalurus punctatus. Aquaculture 1996, 147, 225–233. [Google Scholar]
- Yang, B.; Wang, C.; Hu, H.; Tu, Y.; Han, D.; Zhu, X.; Jin, J.; Yang, Y.; Xie, S. Repeated handling compromises the immune suppression and improves the disease resistance in overwintering channel catfish (Ictalurus punctatus). Fish Shellfish Immunol. 2015, 47, 418–428. [Google Scholar] [CrossRef]
- Sun, C.; Shan, F.; Liu, M.; Liu, B.; Zhou, Q.; Zheng, X.; Xu, X. High-fat-diet-induced oxidative stress in giant freshwater prawn (Macrobrachium rosenbergii) via NF-κB/NO signal pathway and the amelioration of vitamin E. Antioxidants 2022, 11, 228. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ning, L.J.; He, A.Y.; Li, J.M.; Lu, D.L.; Jiao, J.G.; Li, L.Y.; Li, D.L.; Zhang, M.L.; Chen, L.Q.; Du, Z.Y. Mechanisms and metabolic regulation of PPARα activation in nile tilapia (Oreochromis niloticus). Biochim. Biophys. Acta 2016, 1861, 1036–1048. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Q.; Xi, L.; Gong, Y.; Su, J.; Han, D.; Zhang, Z.; Liu, H.; Jin, J.; Yang, Y.; et al. Effects of replacement of dietary fishmeal by cottonseed protein concentrate on growth performance, liver health, and intestinal histology of largemouth bass (Micropterus salmoides). Front. Physiol. 2021, 12, 764987. [Google Scholar] [CrossRef]
- Ensminger, D.C.; Salvador-Pascual, A.; Arango, B.G.; Allen, K.N.; Vázquez-Medina, J.P. Fasting ameliorates oxidative stress: A review of physiological strategies across life history events in wild vertebrates. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2021, 256, 110929. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef]
- Pockley, A.G. Heat shock proteins as regulators of the immune response. Lancet 2003, 362, 469–476. [Google Scholar] [CrossRef]
- Basu, N.; Todgham, A.E.; Ackerman, P.A.; Bibeau, M.R.; Nakano, K.; Schulte, P.M.; Iwama, G.K. Heat shock protein genes and their functional significance in fish. Gene 2002, 295, 173–183. [Google Scholar] [CrossRef]
- Poltronieri, C.; Negrato, E.; Bertotto, D.; Majolini, D.; Simontacchi, C.; Radaelli, G. Immunohistochemical localization of constitutive and inducible heat shock protein 70 in carp (Cyprinus carpio) and trout (Oncorhynchus mykiss) exposed to transport stress. Eur. J. Histochem. 2008, 52, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Hung, S.S.; Fangue, N.A.; Haller, L.; Verhille, C.E.; Zhao, J.; Todgham, A.E. Effects of feed restriction on the upper temperature tolerance and heat shock response in juvenile green and white sturgeon. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 198, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Vincent, H.K.; Innes, K.E.; Vincent, K.R. Oxidative stress and potential interventions to reduce oxidative stress in overweight and obesity. Diabetes Obes. Metab. 2007, 9, 813–839. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Dietary taurine supplementation decreases fat synthesis by suppressing the liver X receptor α pathway and alleviates lipid accumulation in the liver of chronic heat-stressed broilers. J. Sci. Food Agric. 2019, 99, 5631–5637. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xie, H.; Pan, P.; Qu, Q.; Xia, Q.; Gao, X.; Zhang, S.; Jiang, Q. Heat stress promotes lipid accumulation by inhibiting the AMPK-PGC-1α signaling pathway in 3T3-L1 preadipocytes. Cell Stress Chaperones 2021, 26, 563–574. [Google Scholar] [CrossRef]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Farzanegi, P.; Dana, A.; Ebrahimpoor, Z.; Asadi, M.; Azarbayjani, M.A. Mechanisms of beneficial effects of exercise training on non-alcoholic fatty liver disease (NAFLD): Roles of oxidative stress and inflammation. Eur. J. Sport Sci. 2019, 19, 994–1003. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Wu, P.; Chu, Y.; Gui, S.; Zheng, Y.; Chen, X. Dietary selenium alleviated mouse liver oxidative stress and NAFLD induced by obesity by regulating the KEAP1/NRF2 pathway. Antioxidants 2022, 11, 349. [Google Scholar] [CrossRef]
- Yin, P.; Xie, S.; Zhuang, Z.; He, X.; Tang, X.; Tian, L.; Liu, Y.; Niu, J. Dietary supplementation of bile acid attenuate adverse effects of high-fat diet on growth performance, antioxidant ability, lipid accumulation and intestinal health in juvenile largemouth bass (Micropterus salmoides). Aquaculture 2021, 531, 735864. [Google Scholar] [CrossRef]
- Jin, M.; Pan, T.; Cheng, X.; Zhu, T.T.; Sun, P.; Zhou, F.; Ding, X.; Zhou, Q. Effects of supplemental dietary l-carnitine and bile acids on growth performance, antioxidant and immune ability, histopathological changes and inflammatory response in juvenile black seabream (Acanthopagrus schlegelii) fed high-fat diet. Aquaculture 2019, 504, 199–209. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, J.; Zhang, G.; Bu, Y.; Zhang, G.; Zhao, X. Resveratrol and caloric restriction prevent hepatic steatosis by regulating SIRT1-autophagy pathway and alleviating endoplasmic reticulum stress in high-fat diet-fed rats. PLoS ONE 2017, 12, e0183541. [Google Scholar] [CrossRef] [Green Version]
- Escalante-Rojas, M.; Martínez-Brown, J.M.; Ibarra-Castro, L.; Llera-Herrera, R.; García-Gasca, A. Effects of feed restriction on growth performance, lipid mobilization, and gene expression in rose spotted snapper (Lutjanus guttatus). J. Comp. Physiol. B 2020, 190, 275–286. [Google Scholar] [CrossRef]
- Hasanpour, S.; Oujifard, A.; Torfi Mozanzadeh, M.; Safari, O. Compensatory growth, antioxidant capacity and digestive enzyme activities of Sobaity (Sparidentex hasta) and yellowfin seabreams (Acanthopagrus latus) subjected to ration restriction. Aquacult. Nutr. 2021, 27, 2448–2458. [Google Scholar] [CrossRef]
- Tang, Q.; Zheng, G.; Feng, Z.; Chen, Y.; Lou, Y.; Wang, C.; Zhang, X.; Zhang, Y.; Xu, H.; Shang, P.; et al. Trehalose ameliorates oxidative stress-mediated mitochondrial dysfunction and ER stress via selective autophagy stimulation and autophagic flux restoration in osteoarthritis development. Cell Death Dis. 2017, 8, e3081. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.F.; Wang, Z.Q.; Ding, Y.; Piao, M.H.; Feng, C.S.; Chi, G.F.; Luo, Y.N.; Ge, P.F. Endoplasmic reticulum stress regulates oxygen-glucose deprivation-induced parthanatos in human SH-SY5Y cells via improvement of intracellular ROS. CNS Neurosci. Ther. 2018, 24, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.Y.; Zhou, X.D.; Yang, J.; Chen, L.X.; Ran, D.H. Inhibition of autophagy enhances heat-induced apoptosis in human non-small cell lung cancer cells through ER stress pathways. Arch. Biochem. Biophys. 2016, 607, 55–66. [Google Scholar] [CrossRef]
- Oikonomou, C.; Hendershot, L.M. Disposing of misfolded ER proteins: A troubled substrate’s way out of the ER. Mol. Cell. Endocrinol. 2020, 500, 110630. [Google Scholar] [CrossRef]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Selman, C.; Kendaiah, S.; Gredilla, R.; Leeuwenburgh, C. Increased hepatic apoptosis during short-term caloric restriction is not associated with an enhancement in caspase levels. Exp. Gerontol. 2003, 38, 897–903. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperthermia 2014, 30, 513–523. [Google Scholar] [CrossRef]
- Dandekar, A.; Mendez, R.; Zhang, K. Cross talk between ER stress, oxidative stress, and inflammation in health and disease. Methods Mol. Biol. 2015, 1292, 205–214. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Forward (5′-3′) | Reverse (5′-3′) | Accession Numbers |
|---|---|---|---|
| aco | AGACCTGAACTTTCTGTCCCG | GCTGGACACCATAGGGATGAA | XM_017482134.1 |
| acad9 | CTGAAAAGAGTGGCCGATACA | CGTCATTGTTCTCTGGGGAAT | XM_017450491.1 |
| acadvl | CTGTGCGATTGACTTGTATGC | CTCTACCACAGCTGCAGAAAT | XM_017472293.1 |
| acads | TCACTCAGCTTGCAGGATTAC | ATGACTGCTTAGCTGGGAAAG | XM_017489904.1 |
| atf4 | GATTCTGATGGCTGACACCTT | GGTCAAAATCACTGAGGTCGA | XM_017482287.1 |
| atf6 | AGGAGTTTGAGGTGATGATGC | TCTACTCCAATTGCTGACACG | XM_017455892.1 |
| bip | GAGACAGGCCAAGATTGAGAG | TTCTGAGTTGGAGCCTGATTG | XM_017460312.1 |
| cpt1a | ACCACATCCCAATCTGCCTG | CCTGAAGTGAGCAAGCTGGA | XM_017485733.1 |
| caspase3 | GTGTGTGTGATCCTAAGCCAT | CAAATCTGTTCCACGACAAGC | NM_001201081.1 |
| caspase8 | TATGAGGAAGAGGACACGGAA | CGCAATGAGGAAATCTGCATC | XM_017480625.1 |
| caspase9 | TGCTTCCTCTGAAACCAACAA | GATCTGAATCGTCTCTCCAGC | XM_017487604.1 |
| caspase10 | ACGTGGAGTTCTTCTGTGATG | AGCTGTTCGAGGTTCTTACAC | XM_017470757.1 |
| dgat | AAGAAATTCCCCGGGATCAAG | ACAGGAGGTAATCGATGGAGT | NM_001201076.1 |
| eif2 | CATTCTTGGAGGTTCAGAGGG | CATTCAAACGTGCCATCCAAA | XM_017468034.1 |
| fasn | CTGGTCAGAGCAACTACGGTT | CTACATCACCGATAGCACCCC | XM_017483746.1 |
| hsc70 | CAAGATCAGTGACGAGGACAAG | GGTTACAGACTTTCTCCAGTTCC | XM_017489684.1 |
| hsp70 | CTTGATGTTACCCCTCTGTCTCT | TCAGAGTAGGTGGTGAAAGTCTG | NM_001200273.1 |
| hsp90 | ATCTGAAGGAGGATCAGACAGAG | CGCTCCTTCTCTACAAAGAGTGT | NM_001329313.1 |
| lpl | CTGGACGGTTACTGGCATGT | GGCAGCTGAGGTTGGGTAAT | XM_017478439.1 |
| mt | CTGCAAATGCTCAAACTGCCA | AGCACTTGGAATCGCAGGTAT | NM_001200077.1 |
| perilipin | GGACATTGACCATTGCTCGC | ACCTCGCGTGTTTATGAGCA | XM_017472797.1 |
| pparα | TCCGCAAGCCTTTCAGTGAT | AGCAAGGTGTCATCAGGGTG | XM_017494485.1 |
| perk | GCAGCAAAACATTCGTGTGTA | TGGTACGATTCGGTCTCTGTA | XM_017455330.1 |
| pdi8 | GAATGAACCGAGGCGATATGA | CAAACCACATTTCCCAGCATC | XM_017482284.1 |
| scd | GTTTGGACGTGTGGAAATGAC | TGTGTATGTTGAAGCTGAGGG | XM_017464581.1 |
| srebp1c | ACTGGCTGCAAGGAGATGAC | TGTCTCTGAACAGGCTGCTG | XM_017452596.1 |
| xbp1 | TCTGGTCAGGAAAGAGAAGGT | AGCTCTGGTTCAATGATGTCC | XM_017452343.1 |
| β-actin | GGATCTGTATGCCAACACTGT | CAGGTGGGGCAATGATCTTAA | XM_017454668.1 |
| 3% BW | 2.5% BW | 2% BW | p-Value | |
|---|---|---|---|---|
| IBW, g | 35.50 ± 0.06 | 35.50 ± 0.06 | 35.50 ± 0.05 | p > 0.05 |
| FBW, g | 99.86 ± 1.04 b | 97.56 ± 1.55 b | 83.84 ± 0.67 a | p < 0.05 |
| WG, % | 181.28 ± 2.83 b | 174.81 ± 4.24 b | 136.33 ± 1.82 a | p < 0.05 |
| SGR, %/d | 1.15 ± 0.01 b | 1.11 ± 0.01 b | 0.99 ± 0.03 a | p < 0.05 |
| FE, % | 73.82 ± 1.29 a | 84.91 ± 1.59 b | 87.93 ± 0.78 b | p < 0.05 |
| FR, %BW/d | 3.02 ± 0.05 c | 2.48 ± 0.03 b | 2.04 ± 0.02 a | p < 0.05 |
| SR, % | 98.96 ± 0.50 | 99.25 ± 0.50 | 98.75 ± 0.68 | p > 0.05 |
| 3% BW | 2.5% BW | 2% BW | p-Value | |
|---|---|---|---|---|
| HSI, % | 1.47 ± 0.05 b | 1.14 ± 0.10 a | 1.17 ± 0.05 a | p < 0.05 |
| VSI, % | 8.43 ± 0.41 b | 8.01 ± 0.17 ab | 7.45 ± 0.20 a | p < 0.05 |
| CF, g/cm3 | 1.40 ± 0.05 b | 1.26 ± 0.04 a | 1.32 ± 0.04 ab | p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Q.; Gong, Y.; Xi, L.; Liu, Y.; Xu, W.; Liu, H.; Jin, J.; Zhang, Z.; Yang, Y.; Zhu, X.; et al. Feed Restriction Alleviates Chronic Thermal Stress-Induced Liver Oxidation and Damages via Reducing Lipid Accumulation in Channel Catfish (Ictalurus punctatus). Antioxidants 2022, 11, 980. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050980
Lu Q, Gong Y, Xi L, Liu Y, Xu W, Liu H, Jin J, Zhang Z, Yang Y, Zhu X, et al. Feed Restriction Alleviates Chronic Thermal Stress-Induced Liver Oxidation and Damages via Reducing Lipid Accumulation in Channel Catfish (Ictalurus punctatus). Antioxidants. 2022; 11(5):980. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050980
Chicago/Turabian StyleLu, Qisheng, Yulong Gong, Longwei Xi, Yulong Liu, Wenjie Xu, Haokun Liu, Junyan Jin, Zhimin Zhang, Yunxia Yang, Xiaoming Zhu, and et al. 2022. "Feed Restriction Alleviates Chronic Thermal Stress-Induced Liver Oxidation and Damages via Reducing Lipid Accumulation in Channel Catfish (Ictalurus punctatus)" Antioxidants 11, no. 5: 980. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050980