
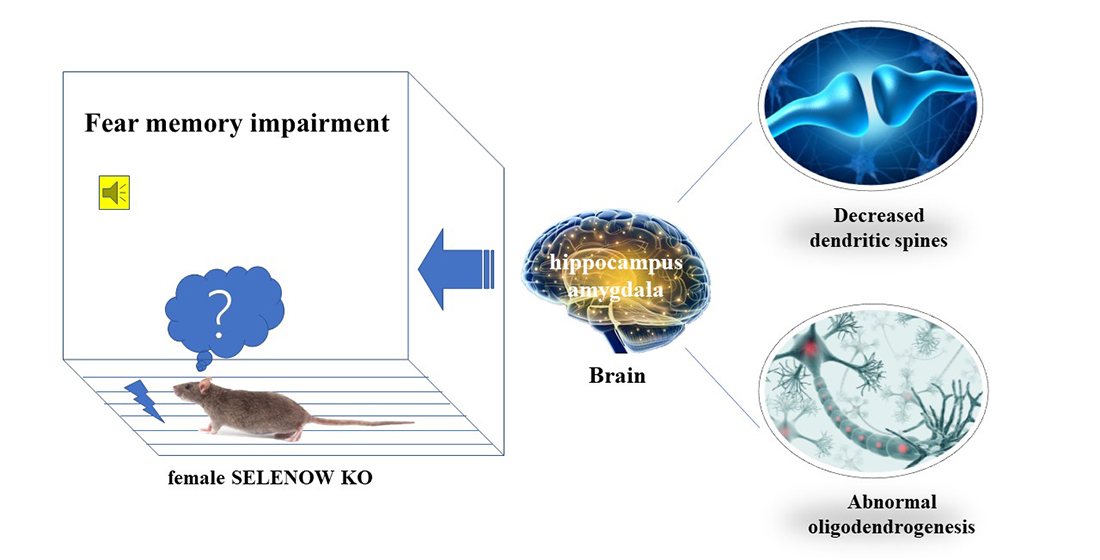
Comparative Proteomic Analysis Reveals the Effect of Selenoprotein W Deficiency on Oligodendrogenesis in Fear Memory


Abstract
:
1. Introduction
2. Materials and Methods
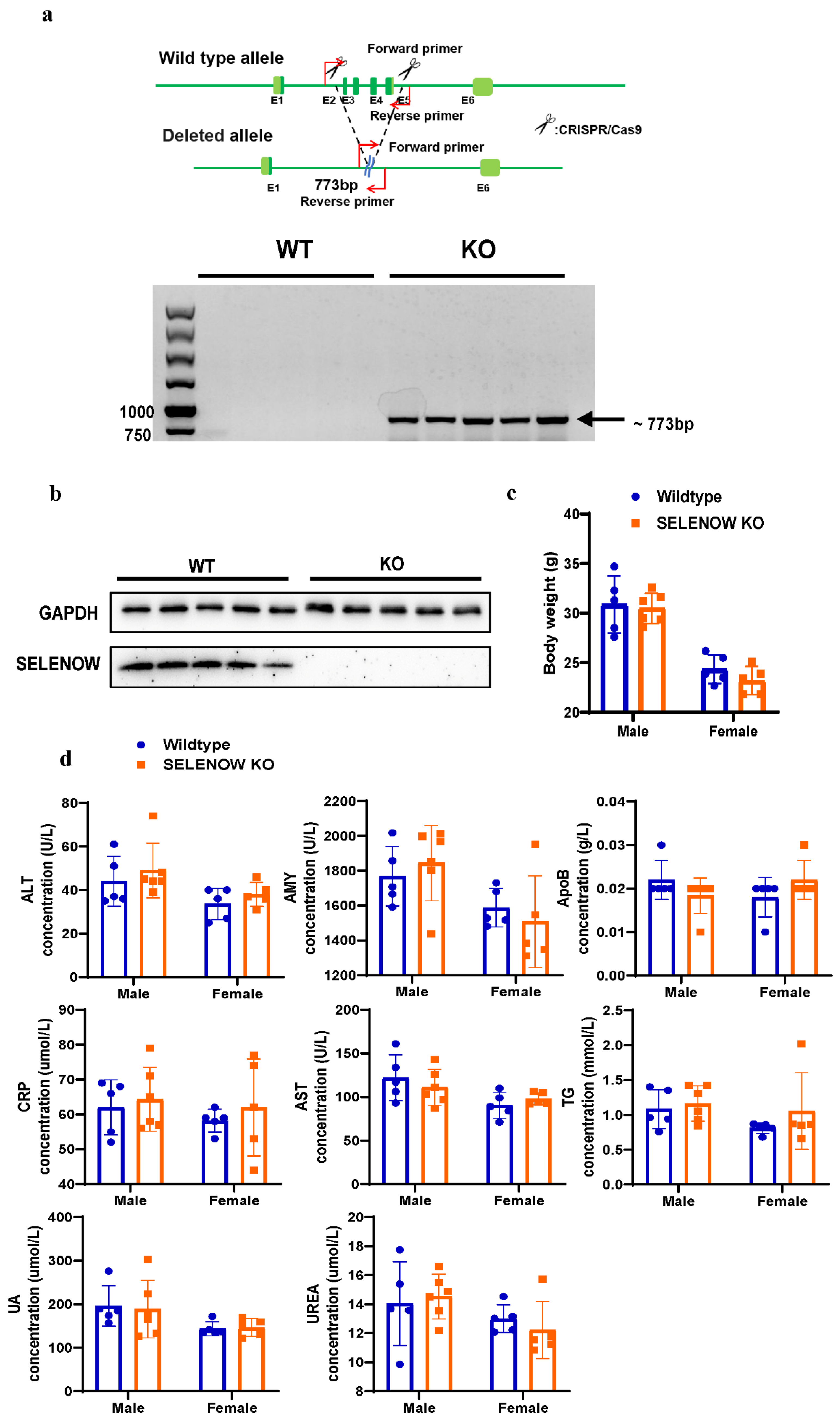
2.1. Animals
2.2. Antibodies
2.3. PCR Amplification
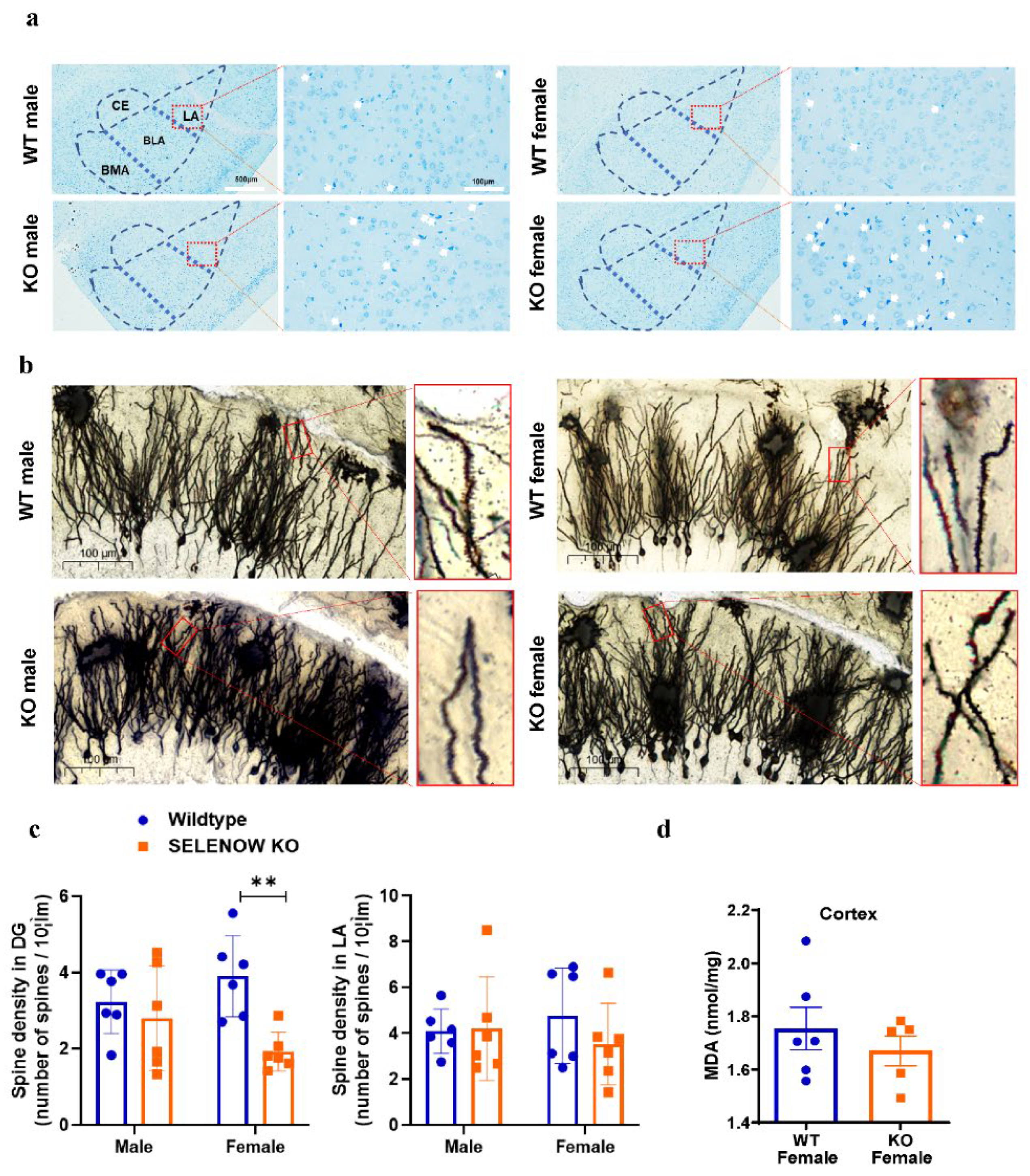
2.4. Brain Tissue Staining and Imaging
2.5. Blood Biochemical and Malondialdehyde (MDA) Analysis
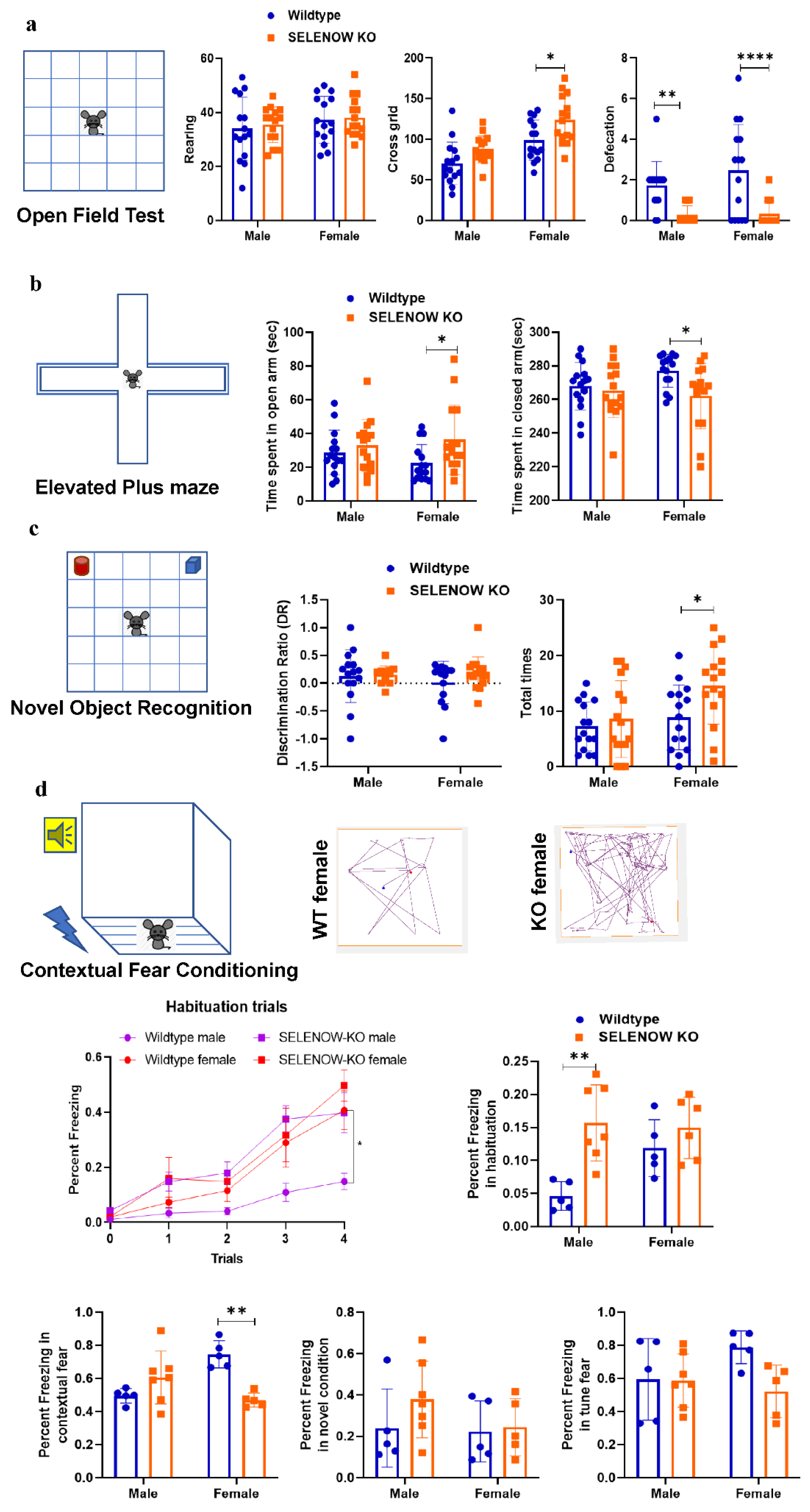
2.6. Behavioral Tests
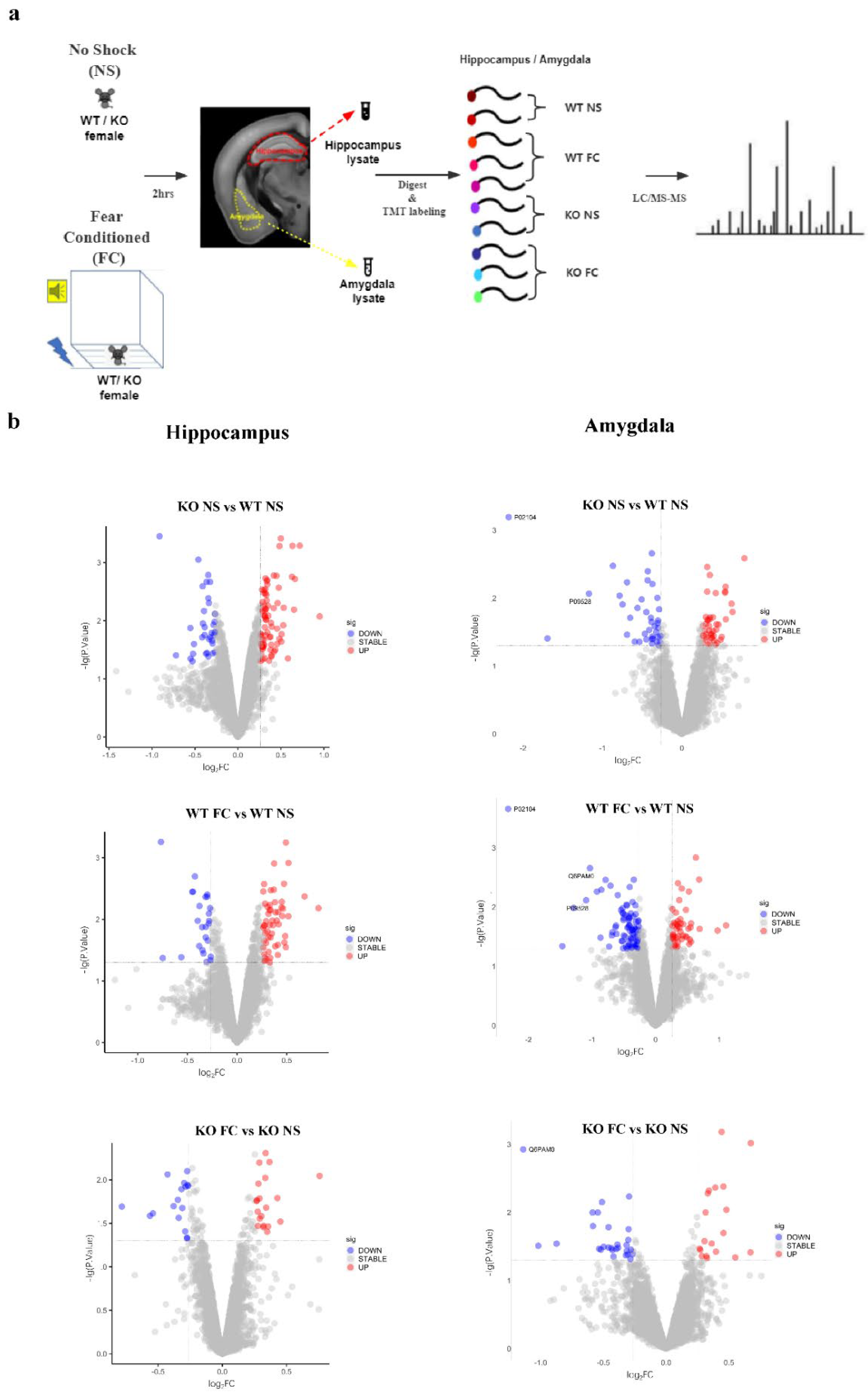
2.7. Tandem Mass Tag (TMT)-Based Quantitative Proteomics
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. Less Anxiety-like Behavior and Impaired Contextual Fear Memory in SELENOW KO Female Mice
3.2. Histomorphologic Alteration of Amygdala and Hippocampus in SELENOW KO Female Mice
3.3. Identification of DEPs by TMT-Based Quantitative Proteomic Analysis
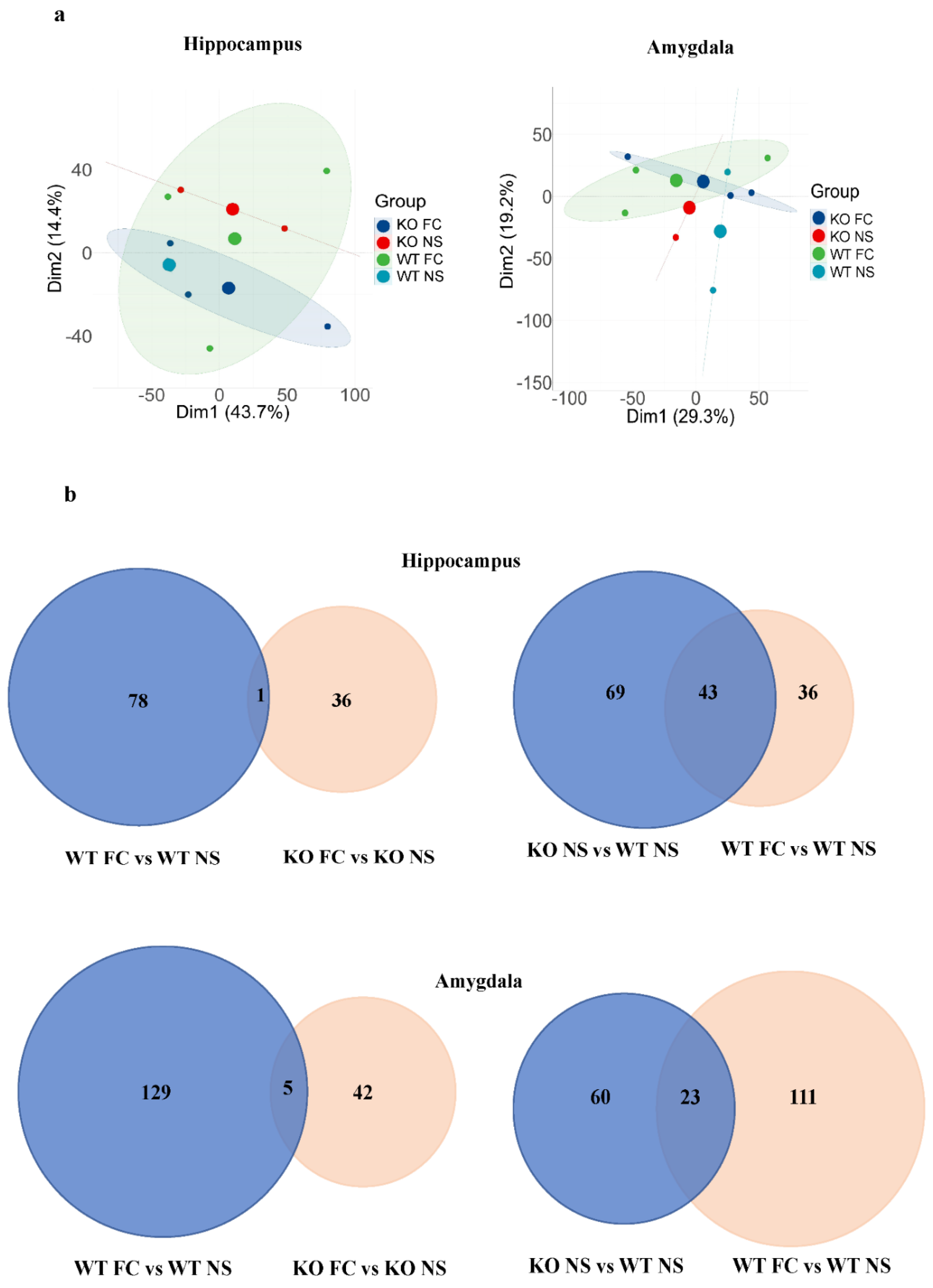
3.4. Analysis and Comparison of Protein Expression Profiles
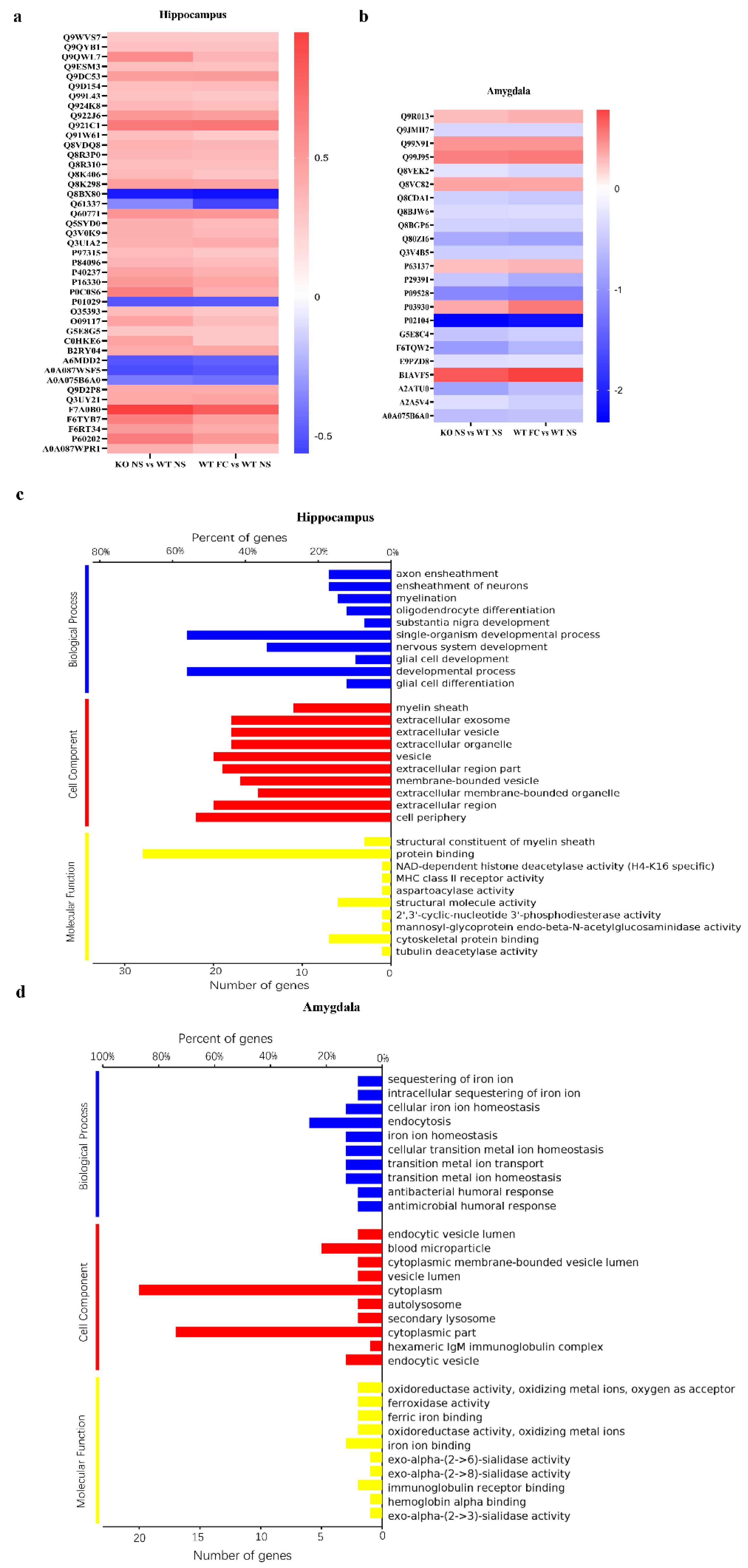
3.5. DEPs Common in Female SELENOW KO and Fear Conditioning WT Mice and Their Gene Ontology Categorization
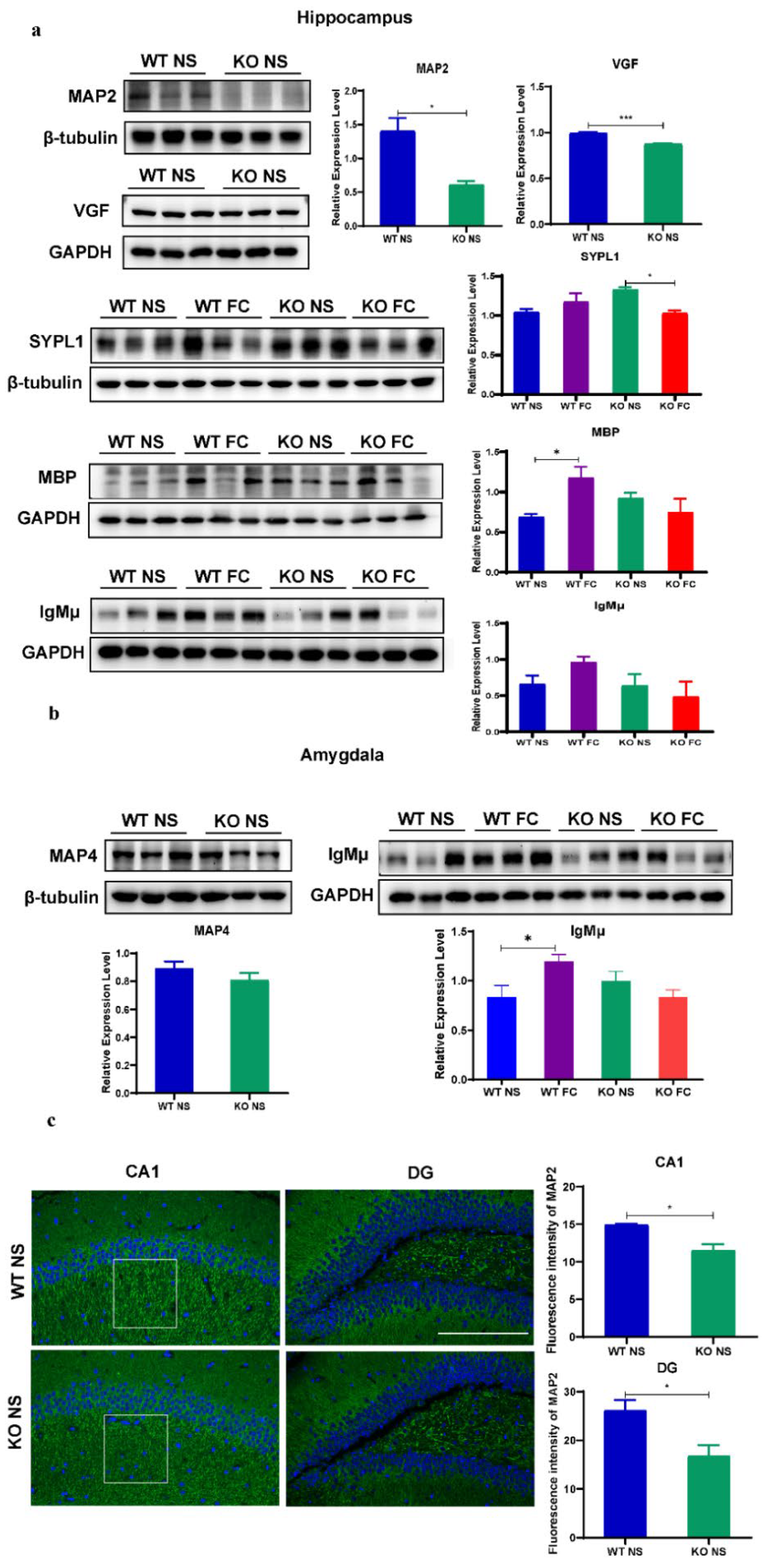
3.6. Validation of Proteomic Results Revealing Abnormal Synaptic Plasticity and Oligodendrogenesis in Female SELENOW KO Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kato, D.; Wake, H. Myelin plasticity modulates neural circuitry required for learning and behavior. Neurosci. Res. 2021, 167, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Chaaya, N.; Battle, A.R.; Johnson, L.R. An update on contextual fear memory mechanisms: Transition between Amygdala and Hippocampus. Neurosci. Biobehav. Rev. 2018, 92, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Terranova, J.I.; Yokose, J.; Osanai, H.; Marks, W.D.; Yamamoto, J.; Ogawa, S.K.; Kitamura, T. Hippocampal-amygdala memory circuits govern experience-dependent observational fear. Neuron 2022, 110, 1416–1431.e13. [Google Scholar] [CrossRef] [PubMed]
- Gresack, J.E.; Schafe, G.E.; Orr, P.T.; Frick, K.M. Sex differences in contextual fear conditioning are associated with differential ventral hippocampal extracellular signal-regulated kinase activation. Neuroscience 2009, 159, 451–467. [Google Scholar] [CrossRef]
- Keiser, A.A.; Turnbull, L.M.; Darian, M.A.; Feldman, D.E.; Song, I.; Tronson, N.C. Sex Differences in Context Fear Generalization and Recruitment of Hippocampus and Amygdala during Retrieval. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2017, 42, 397–407. [Google Scholar] [CrossRef]
- Garcia, R. Neurobiology of fear and specific phobias. Learn. Mem. 2017, 24, 462–471. [Google Scholar] [CrossRef]
- Voĭtsekhovskis, V.V.; Voĭtsekhovska Iu, G.; Shkesters, A.; Antsane, G.; Silova, A.; Ivashchenko, T.; Michans, I.; Vaĭvads, N. Advances of selenium supplementation in posttraumatic stress disorder risk group patients. Biomeditsinskaia Khimiia 2014, 60, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Torres, D.J.; Alfulaij, N.; Berry, M.J. Stress and the Brain: An Emerging Role for Selenium. Front. Neurosci. 2021, 15, 440. [Google Scholar] [CrossRef]
- Low, S.C.; Berry, M.J. Knowing when not to stop: Selenocysteine incorporation in eukaryotes. Trends Biochem. Sci. 1996, 21, 203–208. [Google Scholar] [CrossRef]
- Whanger, P.D. Selenoprotein W: A review. Cell. Mol. Life Sci. CMLS 2000, 57, 1846–1852. [Google Scholar] [CrossRef]
- Gladyshev, V.N.; Arnér, E.S.; Berry, M.J.; Brigelius-Flohé, R.; Bruford, E.A.; Burk, R.F.; Carlson, B.A.; Castellano, S.; Chavatte, L.; Conrad, M.; et al. Selenoprotein Gene Nomenclature. J. Biol. Chem. 2016, 291, 24036–24040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunde, R.A.; Raines, A.M.; Barnes, K.M.; Evenson, J.K. Selenium status highly regulates selenoprotein mRNA levels for only a subset of the selenoproteins in the selenoproteome. Biosci. Rep. 2009, 29, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whanger, P.D. Selenoprotein expression and function—Selenoprotein W. Biochim. Biophys. Acta (BBA) Gen. Subj. 2009, 1790, 1448–1452. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.; Kim, J.M.; Kim, M.Y.; Kim, J.R.; Lee, H.W.; Chung, Y.W.; Shin, H.I.; Kim, T.; Park, E.S.; et al. Selenoprotein W ensures physiological bone remodeling by preventing hyperactivity of osteoclasts. Nat. Commun. 2021, 12, 2258. [Google Scholar] [CrossRef]
- Hawkes, W.C.; Alkan, Z. Regulation of Redox Signaling by Selenoproteins. Biol. Trace Elem. Res. 2010, 134, 235–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Liu, S.; Li, S.; Chen, J.; Ni, J.; Liu, Q. Blocking the Thiol at Cysteine-322 Destabilizes Tau Protein and Prevents Its Oligomer Formation. ACS Chem. Neurosci. 2018, 9, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Raman, A.V.; Pitts, M.W.; Seyedali, A.; Hashimoto, A.C.; Bellinger, F.P.; Berry, M.J. Selenoprotein W expression and regulation in mouse brain and neurons. Brain Behav. 2013, 3, 562–574. [Google Scholar] [CrossRef]
- Kubo, A.; Misonou, H.; Matsuyama, M.; Nomori, A.; Wada-Kakuda, S.; Takashima, A.; Kawata, M.; Murayama, S.; Ihara, Y.; Miyasaka, T. Distribution of endogenous normal tau in the mouse brain. J. Comp. Neurol. 2019, 527, 985–998. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Schweizer, U.; Savaskan, N.E.; Hua, D.; Kipnis, J.; Hatfield, D.L.; Gladyshev, V.N. Comparative analysis of selenocysteine machinery and selenoproteome gene expression in mouse brain identifies neurons as key functional sites of selenium in mammals. J. Biol. Chem. 2008, 283, 2427–2438. [Google Scholar] [CrossRef] [Green Version]
- Yeh, J.Y.; Vendeland, S.C.; Gu, Q.; Butler, J.A.; Ou, B.R.; Whanger, P.D. Dietary selenium increases selenoprotein W levels in rat tissues. J. Nutr. 1997, 127, 2165–2172. [Google Scholar] [CrossRef] [Green Version]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The Open Field Test. In Mood and Anxiety Related Phenotypes in Mice: Characterization Using Behavioral Tests; Gould, T.D., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 1–20. [Google Scholar] [CrossRef]
- Lister, R.G. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology 1987, 92, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, J.D. Chapter 4—Rodent Behavioral Learning and Memory Models. In Mechanisms of Memory, 2nd ed.; Sweatt, J.D., Ed.; Academic Press: London, UK, 2010; pp. 76–103. [Google Scholar] [CrossRef]
- Wahlsten, D. Chapter 3—Tests of Mouse Behavior. In Mouse Behavioral Testing; Wahlsten, D., Ed.; Academic Press: London, UK, 2011; pp. 39–51. [Google Scholar] [CrossRef]
- Steffenach, H.A.; Witter, M.; Moser, M.B.; Moser, E.I. Spatial memory in the rat requires the dorsolateral band of the entorhinal cortex. Neuron 2005, 45, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Kim, W.B.; Cho, J.H. Encoding of contextual fear memory in hippocampal–amygdala circuit. Nat. Commun. 2020, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhang, Z.W.; Yao, H.D.; Li, S.; Xu, S.W. Antioxidative role of selenoprotein W in oxidant-induced chicken splenic lymphocyte death. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2014, 27, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Fan, R.; Zhao, X.; Zhao, W.; Liu, W.; Yang, J.; Sattar, H.; Zhao, J.; Zhang, Z.; Xu, S. Selenoprotein W redox-regulated Ca2+ channels correlate with selenium deficiency-induced muscles Ca2+ leak. Oncotarget 2016, 7, 57618–57632. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.G.; Cha, H.N.; Park, S.; Kim, Y.W.; Kim, J.Y.; Park, S.Y. Selenoprotein W deficiency does not affect oxidative stress and insulin sensitivity in the skeletal muscle of high-fat diet-fed obese mice. Am. J. Physiol. Cell Physiol. 2019, 317, C1172–C1182. [Google Scholar] [CrossRef]
- Yeh, J.Y.; Ou, B.R.; Gu, Q.P.; Whanger, P.D. Influence of gender on selenoprotein W, glutathione peroxidase and selenium in tissues of rats. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1998, 119, 151–155. [Google Scholar] [CrossRef]
- Nebel, R.A.; Aggarwal, N.T.; Barnes, L.L.; Gallagher, A.; Goldstein, J.M.; Kantarci, K.; Mallampalli, M.P.; Mormino, E.C.; Scott, L.; Yu, W.H.; et al. Understanding the impact of sex and gender in Alzheimer’s disease: A call to action. Alzheimer’s Dement. 2018, 14, 1171–1183. [Google Scholar] [CrossRef]
- Xiong, J.; Kang, S.S.; Wang, Z.; Liu, X.; Kuo, T.C.; Korkmaz, F.; Padilla, A.; Miyashita, S.; Chan, P.; Zhang, Z.; et al. FSH blockade improves cognition in mice with Alzheimer’s disease. Nature 2022, 603, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ren, B.; Li, X.; Yan, H.; Xie, Q.; Liu, H.; Zhou, J.; Tian, J.; Huang, K. Selenoprotein F knockout leads to glucose and lipid metabolism disorders in mice. J. Biol. Inorg. Chem. JBIC Publ. Soc. Biol. Inorg. Chem. 2020, 25, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Feng, T.; Liu, L.; Liu, H.; Huang, K.; Zhou, J. Hepatic Proteomic Analysis of Selenoprotein T Knockout Mice by TMT: Implications for the Role of Selenoprotein T in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2021, 22, 8515. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Q.; Zhou, J.C.; Wu, Y.Y.; Ren, F.Z.; Lei, X.G. Role of glutathione peroxidase 1 in glucose and lipid metabolism-related diseases. Free Radic. Biol. Med. 2018, 127, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Leube, R.E. Expression of the synaptophysin gene family is not restricted to neuronal and neuroendocrine differentiation in rat and human. Differ. Res. Biol. Divers. 1994, 56, 163–171. [Google Scholar] [CrossRef]
- Ramkumar, A.; Jong, B.Y.; Ori-McKenney, K.M. ReMAPping the microtubule landscape: How phosphorylation dictates the activities of microtubule-associated proteins. Dev. Dyn. 2018, 247, 138–155. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, R.; Huber, G.; Matus, A. Differences in the developmental patterns of three microtubule-associated proteins in the rat cerebellum. J. Neurosci. Off. J. Soc. Neurosci. 1985, 5, 977–991. [Google Scholar] [CrossRef]
- Matsushima, K.; Tokuraku, K.; Hasan, M.R.; Kotani, S. Microtubule-associated protein 4 binds to actin filaments and modulates their properties. J. Biochem. 2012, 151, 99–108. [Google Scholar] [CrossRef]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Mayoral, S.R.; Choi, H.S.; Chan, J.R.; Kheirbek, M.A. Preservation of a remote fear memory requires new myelin formation. Nat. Neurosci. 2020, 23, 487–499. [Google Scholar] [CrossRef] [Green Version]
- Francis, J.S.; Strande, L.; Markov, V.; Leone, P. Aspartoacylase supports oxidative energy metabolism during myelination. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2012, 32, 1725–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steadman, P.E.; Xia, F.; Ahmed, M.; Mocle, A.J.; Penning, A.R.A.; Geraghty, A.C.; Steenland, H.W.; Monje, M.; Josselyn, S.A.; Frankland, P.W. Disruption of Oligodendrogenesis Impairs Memory Consolidation in Adult Mice. Neuron 2020, 105, 150–164.e156. [Google Scholar] [CrossRef] [PubMed]
- Houyoux, N.; Wattiez, R.; Ris, L. A proteomic analysis of contextual fear conditioned rats reveals dynamic modifications in neuron and oligodendrocyte protein expression in the dentate gyrus. Eur. J. Neurosci. 2017, 46, 2177–2189. [Google Scholar] [CrossRef] [PubMed]
- French, H.M.; Reid, M.; Mamontov, P.; Simmons, R.A.; Grinspan, J.B. Oxidative stress disrupts oligodendrocyte maturation. J. Neurosci. Res. 2009, 87, 3076–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Waly, B.; Macchi, M.; Cayre, M.; Durbec, P. Oligodendrogenesis in the normal and pathological central nervous system. Front. Neurosci. 2014, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.W.; Kim, T.S.; Chung, Y.W.; Lee, B.J.; Kim, I.Y. Selenoprotein W is a glutathione-dependent antioxidant in vivo. FEBS Lett. 2002, 517, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Loflin, J.; Lopez, N.; Whanger, P.D.; Kioussi, C. Selenoprotein W during development and oxidative stress. J. Inorg. Biochem. 2006, 100, 1679–1684. [Google Scholar] [CrossRef]
- Xiao-Long, W.; Chuan-Ping, Y.; Kai, X.; Ou-Jv, Q. Selenoprotein W depletion in vitro might indicate that its main function is not as an antioxidative enzyme. Biochem. Biokhimiia 2010, 75, 201–207. [Google Scholar] [CrossRef]
- Hassan, W.; Gomes Vde, C.; Pinton, S.; Batista Teixeira da Rocha, J.; Landeira-Fernandez, J. Association between oxidative stress and contextual fear conditioning in Carioca high- and low-conditioned freezing rats. Brain Res. 2013, 1512, 60–67. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Region | Uniprot ID | Name | Fold Change | |
|---|---|---|---|---|
| WT FC vs. WT NS | KO FC vs. KO NS | |||
| Hippocampus | O09117 | Synaptophysin-like protein 1 (SYPL1) | 1.26 | 0.83 |
| Amygdala | A0A075B6A0 | Ig mu chain C region (IgMμ) | 0.66 | 0.69 |
| A0A1L1SU40 | Choline transporter-like protein 2 (CTL2) | 0.41 | 0.49 | |
| Q03717 | Potassium voltage-gated channel subfamily B member 1 (KCNB1) | 1.31 | 1.25 | |
| Q6PAM0 | 5’-AMP-activated protein kinase subunit beta-2 (PRKAB2) | 0.49 | 0.45 | |
| Q99J36 | THUMP domain-containing protein 1 (THUMPD1) | 1.55 | 1.60 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Situ, J.; Huang, X.; Zuo, M.; Huang, Y.; Ren, B.; Liu, Q. Comparative Proteomic Analysis Reveals the Effect of Selenoprotein W Deficiency on Oligodendrogenesis in Fear Memory. Antioxidants 2022, 11, 999. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050999
Situ J, Huang X, Zuo M, Huang Y, Ren B, Liu Q. Comparative Proteomic Analysis Reveals the Effect of Selenoprotein W Deficiency on Oligodendrogenesis in Fear Memory. Antioxidants. 2022; 11(5):999. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050999
Chicago/Turabian StyleSitu, Jiaxin, Xuelian Huang, Mingyang Zuo, Yingying Huang, Bingyu Ren, and Qiong Liu. 2022. "Comparative Proteomic Analysis Reveals the Effect of Selenoprotein W Deficiency on Oligodendrogenesis in Fear Memory" Antioxidants 11, no. 5: 999. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11050999