
Discovering the Potential Mechanisms of Medicinal Mushrooms Antidepressant Activity: A Review

Abstract
:1. Introduction
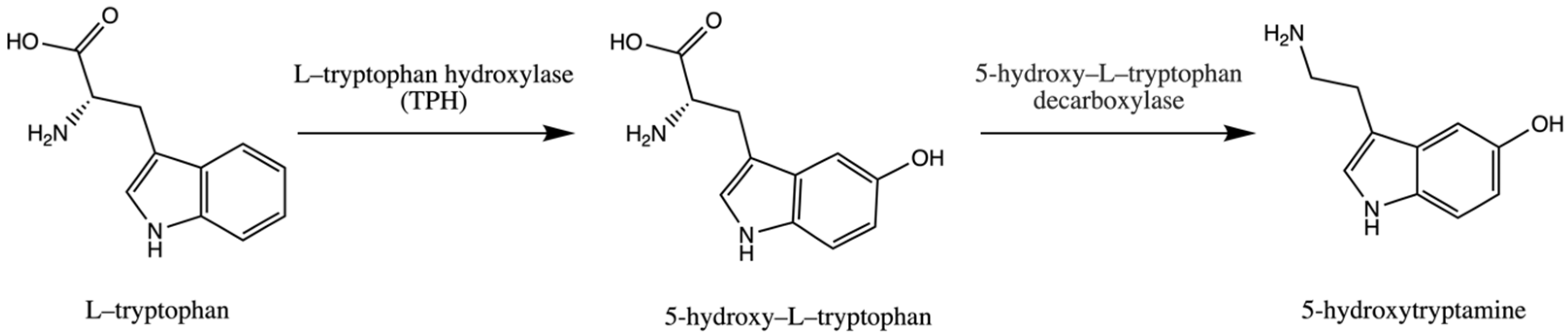
2. L–Tryptophan Derivatives—Essential Compounds for Serotonin Synthesis
3. Tryptamine Derivatives—Psilocybin as a Potential Rapid Acting Antidepressant
4. Anti-Inflammatory Activity of Medicinal Mushrooms in Beating Depression
5. Medicinal Mushrooms and Their Impact on Neurotrophins—Neurotrophin and Neurogenesis-Based Depression
6. Medicinal Mushroom and the Brain–Gut Axis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Moreno-Agostino, D.; Wu, Y.-T.; Daskalopoulou, C.; Hasan, M.T.; Huisman, M.; Prina, M. Global trends in the prevalence and incidence of depression:a systematic review and meta-analysis. J. Affect. Disord. 2021, 281, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Uher, R.; Payne, J.L.; Pavlova, B.; Perlis, R.H. Major Depressive Disorder in DSM-5: Implications for clinical practice and research of changes from DSM-IV. Depress. Anxiety 2014, 31, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.K.; Barton, D.A. Depression and the link with cardiovascular disease. Front. Psychiatry 2016, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galts, C.P.C.; Bettio, L.E.B.; Jewett, D.C.; Yang, C.C.; Brocardo, P.S.; Rodrigues, A.L.S.; Thacker, J.S.; Gil-Mohapel, J. Depression in neurodegenerative diseases: Common mechanisms and current treatment options. Neurosci. Biobehav. Rev. 2019, 102, 56–84. [Google Scholar] [CrossRef]
- Ettman, C.K.; Cohen, G.H.; Abdalla, S.M.; Sampson, L.; Trinquart, L.; Castrucci, B.C.; Bork, R.H.; Clark, M.A.; Wilson, I.; Vivier, P.M.; et al. Persistent depressive symptoms during COVID-19: A national, population-representative, longitudinal study of U.S. adults. Lancet Reg. Health Am. 2022, 5, 100091. [Google Scholar] [CrossRef]
- Calcagno, N.; Colombo, E.; Maranzano, A.; Pasquini, J.; Keller Sarmiento, I.J.; Trogu, F.; Silani, V. Rising evidence for neurological involvement in COVID-19 Pandemic. Neurol. Sci. 2020, 41, 1339. [Google Scholar] [CrossRef]
- Aghagoli, G.; Gallo Marin, B.; Katchur, N.J.; Chaves-Sell, F.; Asaad, W.F.; Murphy, S.A. Neurological involvement in COVID-19 and potential mechanisms: A review. Neurocrit. Care 2021, 34, 1062–1071. [Google Scholar] [CrossRef]
- Ban, T.A. Pharmacotherapy of mental illness—A historical analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2001, 25, 709–727. [Google Scholar] [CrossRef]
- Bloch, R.G.; Dooneief, A.S.; Buchberg, A.S.; Sellman, S. The clinical effect of isoniazid and iproniazid in the treatment of pulmonary tuberculosis. Ann. Intern. Med. 1954, 40, 881–900. [Google Scholar] [CrossRef]
- Ayd, F.J. A preliminary report on marsilid. Am. J. Psychiatry 1957, 114, 459. [Google Scholar] [CrossRef]
- Goldberg, L.I. Monoamine Oxidase Inhibitors. JAMA 1964, 190, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Perez-Caballero, L.; Torres-Sanchez, S.; Romero-López-Alberca, C.; González-Saiz, F.; Mico, J.A.; Berrocoso, E. Monoaminergic system and depression. Cell Tissue Res. 2019, 377, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, M.; Linda, F.K.; Ramesh, S. Role of selective serotonin reuptake inhibitors in psychiatric disorders: A comprehensive review. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.D.; Duman, R.S. The role of neurotrophic factors in adult hippocampal neurogenesis, antidepressant treatments and animal models of depressive-like behavior. Behav. Pharmacol. 2007, 18, 391–418. [Google Scholar] [CrossRef] [PubMed]
- Castrén, E.; Võikar, V.; Rantamäki, T. Role of neurotrophic factors in depression. Curr. Opin. Pharmacol. 2007, 7, 18–21. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Lopes, M.; Fregni, F. A systematic review and meta-analysis of clinical studies on Major Depression and BDNF levels: Implications for the role of neuroplasticity in depression. Int. J. Neuropsychopharmacol. 2008, 11, 1169–1180. [Google Scholar] [CrossRef]
- Sairanen, M.; Lucas, G.; Ernfors, P.; Castrén, M.; Castrén, E. Brain-Derived Neurotrophic Factor and antidepressant drugs have different but coordinated effects on neuronal turnover, proliferation, and survival in the adult dentate gyrus. J. Neurosci. 2005, 25, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and TrkB MRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef]
- Neto, F.L.; Borges, G.; Torres-Sanchez, S.; Mico, J.A.; Berrocoso, E. Neurotrophins role in depression neurobiology: A review of basic and clinical evidence. Curr. Neuropharmacol. 2011, 9, 530–552. [Google Scholar] [CrossRef] [Green Version]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Y.; Ho, K.J.; Hsieh, Y.J.; Wang, L.T.; Mau, J.L. Contents of lovastatin, γ-aminobutyric acid and ergothioneine in mushroom fruiting bodies and mycelia. LWT 2012, 47, 274–278. [Google Scholar] [CrossRef]
- Muszyńska, B.; Kała, K.; Włodarczyk, A.; Krakowska, A.; Ostachowicz, B.; Gdula-Argasińska, J.; Suchocki, P. Lentinula edodes as a source of bioelements released into artificial digestive juices and potential anti-inflammatory material. Biol. Trace Elem. Res. 2020, 194, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Ba, D.M.; Gao, X.; Al-Shaar, L.; Muscat, J.E.; Chinchilli, V.M.; Beelman, R.B.; Richie, J.P. Mushroom intake and depression: A population-based study using data from the US National Health and Nutrition Examination Survey (NHANES), 2005–2016. J. Affect. Disord. 2021, 294, 686–692. [Google Scholar] [CrossRef]
- Vigna, L.; Morelli, F.; Agnelli, G.M.; Napolitano, F.; Ratto, D.; Occhinegro, A.; di Iorio, C.; Savino, E.; Girometta, C.; Brandalise, F.; et al. Hericium erinaceus improves mood and sleep disorders in patients affected by overweight or obesity: Could circulating Pro-BDNF and BDNF be potential biomarkers? Evid. Based Complement. Altern. Med. 2019, 2019, 7861297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, H.; Anno, N.; Tsuda, A.; Inokuchi, T.; Uchimura, N.; Inanaga, K. The effects of Hericium erinaceus on sleep quality and subjective well-being among female undergraduate students: A pilot study. Pers. Med. Universe 2015, 4, 76–78. [Google Scholar] [CrossRef]
- Park, S.K.; Oh, C.M.; Ryoo, J.H.; Jung, J.Y. The protective effect of mushroom consumption on depressive symptoms in Korean population. Sci. Rep. 2022, 12, 21914. [Google Scholar] [CrossRef]
- Bell, C.J.; Hood, S.D.; Nutt, D.J. Acute tryptophan depletion. Part II: Clinical Effects and Implications. Aust. N. Z. J. Psychiatry 2005, 39, 565–574. [Google Scholar] [CrossRef]
- Oladele, J.O.; Oladiji, A.T.; Oladele, O.T.; Oyeleke, O.M. Reactive oxygen species in neurodegenerative diseases: Implications in pathogenesis and treatment strategies. In Reactive Oxygen Species; Ahmad, R., Ed.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Pardridge, W.M. Blood-Brain Barrier carrier-mediated transport and brain metabolism of amino acids. Neurochem. Res. 1998, 23, 635–644. [Google Scholar] [CrossRef]
- Silber, B.Y.; Schmitt, J.A.J. Effects of tryptophan loading on human cognition, mood, and sleep. Neurosci. Biobehav. Rev. 2010, 34, 387–407. [Google Scholar] [CrossRef]
- Ruddick, J.P.; Evans, A.K.; Nutt, D.J.; Lightman, S.L.; Rook, G.A.W.; Lowry, C.A. Tryptophan metabolism in the Central Nervous System: Medical implications. Expert Rev. Mol. Med. 2006, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Fernstrom, J.D. Large neutral amino acids: Dietary effects on brain neurochemistry and function. Amino Acids 2013, 45, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Tovar, A.R.; Tews, J.K.; Torres, N.; Harper, A.E. Neutral amino acid transport into rat skeletal muscle: Competition, adaptive regulation, and effects of insulin. Metabolism 1991, 40, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357, eaaf9794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muszyńska, B.; Sułkowska–Ziaja, K.; Łojewski, M.; Opoka, W.; Zając, M.; Rojowski, J. Edible mushroom in prophylaxis and treatment of human diseases. Med. Int. Rev. 2013, 101, 170–183. [Google Scholar]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Indole compounds in some culinary-medicinal higher basidiomycetes from Poland. Int. J. Med. Mushrooms 2011, 13, 449–454. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Maślanka, A.; Rojowski, J.; Opoka, W.; Łojewski, M. Analysis of indole compounds quantity in biomass of edible Basidiomycota species from in vitro cultures and from cultures supplemented with L-tryptophan. Acta Chromatogr. 2016, 28, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Krakowska, A.; Zięba, P.; Włodarczyk, A.; Kała, K.; Sułkowska-Ziaja, K.; Bernaś, E.; Sękara, A.; Ostachowicz, B.; Muszyńska, B. Selected edible medicinal mushrooms from Pleurotus genus as an answer for human civilization diseases. Food Chem. 2020, 327, 127084. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Wójcik, A. Levels of physiologically active indole derivatives in the fruiting bodies of some edible mushrooms (Basidiomycota) before and after thermal processing. Mycoscience 2013, 54, 321–326. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K. Analysis of indole compounds in edible Basidiomycota species after thermal processing. Food Chem. 2012, 132, 455–459. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Muszyńska, B.; Szewczyk, A. Antioxidant components of selected indigenous edible mushrooms of the obsolete order Aphyllophorales. Rev. Iberoam. Micol. 2015, 32, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Indole compounds in fruiting bodies of some edible Basidiomycota species. Food Chem. 2011, 125, 1306–1308. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Indole compounds in fruiting bodies of some selected macromycetes species and in their mycelia cultured in vitro. Pharmazie 2009, 64, 479–480. [Google Scholar] [CrossRef]
- Çevikkalp, S.A.; Löker, G.B.; Yaman, M.; Amoutzopoulos, B. A Simplified HPLC method for determination of tryptophan in some cereals and legumes. Food Chem. 2016, 193, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Analysis, nutrition, and health benefits of tryptophan. Int. J. Tryptophan Res. 2018, 11, 1178646918802282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maffei, M.E. 5-Hydroxytryptophan (5-HTP): Natural occurrence, analysis, biosynthesis, biotechnology, physiology and toxicology. Int. J. Mol. Sci. 2020, 22, 181. [Google Scholar] [CrossRef]
- Vigliante, I.; Mannino, G.; Maffei, M.E. Chemical characterization and DNA fingerprinting of Griffonia simplicifolia Baill. Molecules 2019, 24, 1032. [Google Scholar] [CrossRef] [Green Version]
- Ly, D.; Kang, K.; Choi, J.Y.; Ishihara, A.; Back, K.; Lee, S.G. HPLC Analysis of serotonin, tryptamine, tyramine, and the hydroxycinnamic acid amides of serotonin and tyramine in food vegetables. J. Med. Food 2008, 11, 385–389. [Google Scholar] [CrossRef]
- Muszyńska, B.; Kała, K.; Firlej, A.; Sułkowska-Ziaja, K. Cantharellus cibarius–culinary-medicinal mushroom content and biological activity. Acta Pol. Pharm. 2016, 73, 589–598. [Google Scholar]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Hałaszczuk, P.; Krężałek, R.; Łojewski, M. Analysis of indole derivatives in methanolic extracts from mycelium of Agaricus bisporus cultured in vitro on liquid Oddoux medium. Folia Biol. Oecologica 2014, 10, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Sułkowska-Ziaja, K.; Muszyńska, B.; Ekiert, H. Analysis of indole compounds from the fruiting bodies and the culture mycelia of Sarcodon imbricatus. Mycoscience 2014, 55, 164–167. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K. Analysis of indole compounds in fruiting bodies and in mycelia from in vitro cultures of Calocera viscosa (Basidiomycota). Acta Mycol. 2012, 47, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Analysis of indole compounds in methanolic extracts from the fruiting bodies of Cantharellus cibarius (the Chanterelle) and from the mycelium of this species cultured in vitro. J. Food Sci. Technol. 2013, 50, 1233–1237. [Google Scholar] [CrossRef] [Green Version]
- Opoka, W.; Kała, K.; Krężałek, R.; Sułkowska-Ziaja, K.; Maślanka, A.; Muszyńska, B. TLC–densitometry analysis of indole compounds in mycelial culture of Imleria badia and Agaricus bisporus enriched with precursors—serine or anthranilic acid. Acta Chromatogr. 2018, 30, 236–242. [Google Scholar] [CrossRef]
- Muszyńska, B.; Kała, K.; Sułkowska-Ziaja, K.; Krakowska, A.; Opoka, W. Agaricus bisporus and its in vitro culture as a source of indole compounds released into artificial digestive juices. Food Chem. 2016, 199, 509–515. [Google Scholar] [CrossRef]
- Kała, K.; Sułkowska-Ziaja, K.; Rojowski, J.; Opoka, W.; Muszyńska, B. Tricholoma equestre species as a source of indole compounds and zinc released into artificial digestive juices. Med. Int. Rev. 2016, 106, 35–39. [Google Scholar]
- Kała, K.; Krakowska, A.; Sułkowska-Ziaja, K.; Szewczyk, A.; Reczyński, W.; Opoka, W.; Muszyńska, B. Kinetics of extracted bioactive components from mushrooms in artificial digestive juices. Int. J. Food Prop. 2016, 20, 1796–1817. [Google Scholar] [CrossRef] [Green Version]
- Saghir, S.A. Absorption. Encycl. Toxicol. Third Ed. 2014, 1–6. [Google Scholar] [CrossRef]
- Fang, Y.; Cao, W.; Xia, M.; Pan, S.; Xu, X. Study of structure and permeability relationship of flavonoids in Caco-2 Cells. Nutrients 2017, 9, 1301. [Google Scholar] [CrossRef] [Green Version]
- Kała, K.; Krakowska, A.; Gdula-Argasińska, J.; Opoka, W.; Muszyńska, B. Assessing the bioavailability of zinc and indole compounds from mycelial cultures of the Bay Mushroom Imleria badia (Agaricomycetes) using in vitro models. Int. J. Med. Mushrooms 2019, 21, 343–352. [Google Scholar] [CrossRef]
- Voineskos, D.; Daskalakis, Z.J.; Blumberger, D.M. Management of Treatment-Resistant Depression: Challenges and strategies. Neuropsychiatr. Dis. Treat. 2020, 16, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaynes, B.N.; Lux, L.; Gartlehner, G.; Asher, G.; Forman-Hoffman, V.; Green, J.; Boland, E.; Weber, R.P.; Randolph, C.; Bann, C.; et al. Defining Treatment-Resistant Depression. Depress. Anxiety 2020, 37, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Riggs, L.M.; Gould, T.D. Ketamine and the future of Rapid-Acting Antidepressants. Annu. Rev. Clin. Psychol. 2021, 17, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Witkin, J.M.; Martin, A.E.; Golani, L.K.; Xu, N.Z.; Smith, J.L. Rapid-Acting Antidepressants. Adv. Pharmacol. 2019, 86, 47–96. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Rosenblat, J.D.; Nemeroff, C.B.; Sanacora, G.; Murrough, J.W.; Berk, M.; Brietzke, E.; Dodd, S.; Gorwood, P.; Ho, R.; et al. Synthesizing the evidence for ketamine and esketamine in Treatment-Resistant Depression: An international expert opinion on the available evidence and implementation. Am. J. Psychiatry 2021, 178, 383–399. [Google Scholar] [CrossRef]
- Machado-Vieira, R.; Henter, I.D.; Zarate, C.A. New targets for Rapid Antidepressant Action. Prog. Neurobiol. 2017, 152, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Kosentka, P.; Sprague, S.L.; Ryberg, M.; Gartz, J.; May, A.L.; Campagna, S.R.; Matheny, P.B. Evolution of the toxins muscarine and psilocybin in a family of mushroom-forming fungi. PLoS ONE 2013, 8, e64646. [Google Scholar] [CrossRef] [Green Version]
- Kreisel, H.; Lindquest, U. Gymnopilus purpuratus, ein psilocybinhaltiger Pilz adventiv in Berzirk Rostock. Z. Mykol. 1988, 54, 73–76. [Google Scholar]
- Halama, M.; Poliwoda, A.; Jasicka-Misiak, I.; Wieczorek, P.P.; Rutkowski, R. Pholiotina cyanopus, a rare fungus producing psychoactive tryptamines. Open Life Sci. 2015, 10, 40–51. [Google Scholar] [CrossRef]
- Saupe, S.G. Occurrence of psilocybin/psilocin in Pluteus salicinus (Pluteaceae). Mycologia 2018, 73, 781–784. [Google Scholar] [CrossRef]
- Stijve, T. Psilocin, psilocybin, serotonin and urea in Panaeolus cyanescens from various origin. Pers. Mol. Phylogeny Evol. Fungi 1992, 15, 117–121. [Google Scholar]
- Froese, T.; Guzmán, G.; Guzmán-Dávalos, L. On the origin of the genus Psilocybe and its potential ritual use in ancient Africa and Europe. Econ. Bot. 2016, 70, 103–114. [Google Scholar] [CrossRef]
- Mahapatra, A.; Gupta, R. Role of psilocybin in the treatment of depression. Ther. Adv. Psychopharmacol. 2017, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Quednow, B.B.; Kometer, M.; Geyer, M.A.; Vollenweider, F.X. Psilocybin-induced deficits in automatic and controlled inhibition are attenuated by ketanserin in healthy human volunteers. Neuropsychopharmacology 2012, 37, 630–640. [Google Scholar] [CrossRef] [Green Version]
- Eison, A.S.; Mullins, U.L. Regulation of central 5-HT2A receptors: A review of in vivo studies. Behav. Brain Res. 1996, 73, 177–181. [Google Scholar] [CrossRef]
- Van Oekelen, D.; Luyten, W.H.M.L.; Leysen, J.E. 5-HT2A and 5-HT2C receptors and their atypical regulation properties. Life Sci. 2003, 72, 2429–2449. [Google Scholar] [CrossRef]
- Gill, H.; Gill, B.; Chen-Li, D.; El-Halabi, S.; Rodrigues, N.B.; Cha, D.S.; Lipsitz, O.; Lee, Y.; Rosenblat, J.D.; Majeed, A.; et al. The emerging role of psilocybin and MDMA in the treatment of mental illness. Expert Rev. Neurother. 2020, 20, 1263–1273. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Hallak, J.E.C. Therapeutic use of serotoninergic hallucinogens: A review of the evidence and of the biological and psychological mechanisms. Neurosci. Biobehav. Rev. 2020, 108, 423–434. [Google Scholar] [CrossRef]
- Nkadimeng, S.M.; Steinmann, C.M.L.; Eloff, J.N. Anti-inflammatory effects of four psilocybin-containing magic mushroom water extracts in vitro on 15-lipoxygenase activity and on lipopolysaccharide-induced cyclooxygenase-2 and inflammatory cytokines in human U937 macrophage cells. J. Inflamm. Res. 2021, 14, 3729–3738. [Google Scholar] [CrossRef]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef]
- Ko, K.; Kopra, E.I.; Cleare, A.J.; Rucker, J.J. Psychedelic therapy for depressive symptoms: A systematic review and meta-analysis. J. Affect. Disord. 2023, 322, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.M.; Aaronson, S.T.; Alvarez, O.; Arden, P.C.; Baker, A.; Bennett, J.C.; Bird, C.; Blom, R.E.; Brennan, C.; Brusch, D.; et al. Single-dose psilocybin for a treatment-resistant episode of major depression. N. Engl. J. Med. 2022, 387, 1637–1648. [Google Scholar] [CrossRef] [PubMed]
- Gotvaldová, K.; Borovička, J.; Hájková, K.; Cihlářová, P.; Rockefeller, A.; Kuchař, M. Extensive collection of psychotropic mushrooms with determination of their tryptamine alkaloids †. Int. J. Mol. Sci. 2022, 23, 14068. [Google Scholar] [CrossRef]
- Milne, N.; Thomsen, P.; Mølgaard Knudsen, N.; Rubaszka, P.; Kristensen, M.; Borodina, I. Metabolic engineering of Saccharomyces cerevisiae for the de novo production of psilocybin and related tryptamine derivatives. Metab. Eng. 2020, 60, 25–36. [Google Scholar] [CrossRef]
- Adams, A.M.; Kaplan, N.A.; Wei, Z.; Brinton, J.D.; Monnier, C.S.; Enacopol, A.L.; Ramelot, T.A.; Jones, J.A. In vivo production of psilocybin in E. coli. Metab. Eng. 2019, 56, 111–119. [Google Scholar] [CrossRef]
- Vollenweider, F.X.; Kometer, M. The neurobiology of psychedelic drugs: Implications for the treatment of mood disorders. Nat. Rev. Neurosci. 2010, 11, 642–651. [Google Scholar] [CrossRef]
- Hutten, N.R.P.W.; Mason, N.L.; Dolder, P.C.; Kuypers, K.P.C. Motives and side-effects of microdosing with psychedelics among users. Int. J. Neuropsychopharmacol. 2019, 22, 426. [Google Scholar] [CrossRef] [Green Version]
- Polito, V.; Stevenson, R.J. A systematic study of microdosing psychedelics. PLoS ONE 2019, 14, e0211023. [Google Scholar] [CrossRef] [Green Version]
- Prochazkova, L.; Lippelt, D.P.; Colzato, L.S.; Kuchar, M.; Sjoerds, Z.; Hommel, B. Exploring the effect of microdosing psychedelics on creativity in an open-label natural setting. Psychopharmacology 2018, 235, 3401–3413. [Google Scholar] [CrossRef] [Green Version]
- Cavanna, F.; Muller, S.; de la Fuente, L.A.; Zamberlan, F.; Palmucci, M.; Janeckova, L.; Kuchar, M.; Pallavicini, C.; Tagliazucchi, E. Microdosing with psilocybin mushrooms: A double-blind placebo-controlled study. Transl. Psychiatry 2022, 12, 307. [Google Scholar] [CrossRef]
- Nesić, Z.; Delić, D.; Prostran, M.; Stojanović, R.; Vucković, S.; Todorović, Z. Psychiatric adverse effects induced by recombinant interferon alfa in patients with chronic hepatitis C. Med. Pregl. 2004, 57, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, A.; Tam, W.W.; Zhang, M.W.; Ho, C.S.; Husain, S.F.; McIntyre, R.S.; Ho, R.C. IL-1β, IL-6, TNF- α and CRP in elderly patients with depression or Alzheimer’s disease: Systematic review and meta-analysis. Sci. Rep. 2018, 8, 12050. [Google Scholar] [CrossRef] [PubMed]
- Osimo, E.F.; Pillinger, T.; Rodriguez, I.M.; Khandaker, G.M.; Pariante, C.M.; Howes, O.D. Inflammatory markers in depression: A meta-analysis of mean differences and variability in 5,166 patients and 5,083 controls. Brain Behav. Immun. 2020, 87, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.A.; Torre, J.P.; Papadopoulos, A.S.; Poon, L.; Juruena, M.F.; Markopoulou, K.; Cleare, A.J.; Pariante, C.M. Lack of clinical therapeutic benefit of antidepressants is associated overall activation of the inflammatory system. J. Affect. Disord. 2013, 148, 136–140. [Google Scholar] [CrossRef]
- Varatharaj, A.; Galea, I. The Blood-Brain Barrier in systemic inflammation. Brain Behav. Immun. 2017, 60, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Widner, B.; Laich, A.; Sperner-Unterweger, B.; Ledochowski, M.; Fuchs, D. Neopterin production, tryptophan degradation, and mental depression—What is the link? Brain Behav. Immun. 2002, 16, 590–595. [Google Scholar] [CrossRef]
- Dunn, A.J.; Welch, J. Stress-and endotoxin-induced increases in brain tryptophan and serotonin metabolism depend on sympathetic nervous system activity. J. Neurochem. 1991, 57, 1615–1622. [Google Scholar] [CrossRef]
- O’Connor, J.C.; Lawson, M.A.; André, C.; Moreau, M.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Mol. Psychiatry 2009, 14, 511–522. [Google Scholar] [CrossRef] [Green Version]
- Raison, C.L.; Dantzer, R.; Kelley, K.W.; Lawson, M.A.; Woolwine, B.J.; Vogt, G.; Spivey, J.R.; Saito, K.; Miller, A.H. CSF concentrations of brain tryptophan and kynurenines during immune stimulation with IFN-alpha: Relationship to CNS immune responses and depression. Mol. Psychiatry 2010, 15, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.; Leonard, B.E.; Myint, A.M.; Kubera, M.; Verkerk, R. The new ‘5-HT’ hypothesis of depression: Cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to the onset of depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 702–721. [Google Scholar] [CrossRef]
- Guidetti, P.; Eastman, C.L.; Schwarcz, R. Metabolism of [5-3H]Kynurenine in the rat brain in vivo: Evidence for the existence of a functional kynurenine pathway. J. Neurochem. 1995, 65, 2621–2632. [Google Scholar] [CrossRef] [PubMed]
- Sanlioglu, S.; Williams, C.M.; Samavati, L.; Butler, N.S.; Wang, G.; McCray, P.B.; Ritchie, T.C.; Hunninghake, G.W.; Zandi, E.; Engelhardt, J.F. Lipopolysaccharide induces Rac1-dependent reactive oxygen species formation and coordinates Tumor Necrosis Factor-α secretion through IKK Regulation of NF-ΚB. J. Biol. Chem. 2001, 276, 30188–30198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemin, G.J.; Kerr, S.J.; Smythe, G.A.; Smith, D.G.; Kapoor, V.; Armati, P.J.; Croitoru, J.; Brew, B.J. Kynurenine pathway metabolism in human astrocytes: A paradox for neuronal protection. J. Neurochem. 2001, 78, 842–853. [Google Scholar] [CrossRef]
- Ogyu, K.; Kubo, K.; Noda, Y.; Iwata, Y.; Tsugawa, S.; Omura, Y.; Wada, M.; Tarumi, R.; Plitman, E.; Moriguchi, S.; et al. Kynurenine pathway in depression: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2018, 90, 16–25. [Google Scholar] [CrossRef]
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Jiang, P.; Zhao, J.; Shi, H.; Zhang, P.; Yang, X.; Biazik, J.; Hu, M.; Hua, H.; Ge, X.; et al. β-Glucan from Lentinula edodes prevents cognitive impairments in high-fat diet-induced obese mice: Involvement of colon-brain axis. J. Transl. Med. 2021, 19, 54. [Google Scholar] [CrossRef]
- Zhang, D.; Jia, Y.; Sun, X.; Li, H.; Yin, M.; Li, H.; Dai, L.; Han, L.; Wang, L.; Qian, M.; et al. The dectin-1 receptor signaling pathway mediates the remyelination effect of lentinan through suppression of neuroinflammation and conversion of microglia. J. Immunol. Res. 2022, 2022, 3002304. [Google Scholar] [CrossRef]
- Maza, P.A.M.A.; Lee, J.-H.; Kim, Y.-S.; Sun, G.-M.; Sung, Y.-J.; Ponomarenko, L.P.; Stonik, V.A.; Ryu, M.; Kwak, J.-Y. Inotodiol from Inonotus obliquus Chaga mushroom induces atypical maturation in dendritic cells. Front. Immunol. 2021, 12, 650841. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Guo, W.H.; Cao, C.Y.; Kou, R.W.; Xu, Y.Z.; Górecki, M.; di Bari, L.; Pescitelli, G.; Gao, J.M. Polyoxygenated cyathane diterpenoids from the mushroom Cyathus africanus, and their neurotrophic and anti-neuroinflammatory activities. Sci. Rep. 2018, 8, 2175. [Google Scholar] [CrossRef] [Green Version]
- Nguepi Tsopmejio, I.S.; Ding, M.; Wei, J.; Zhao, C.; Jiang, Y.; Li, Y.; Song, H. Auricularia polytricha and Flammulina velutipes ameliorate inflammation and modulate the gut microbiota via regulation of NF-ΚB and Keap1/Nrf2 signaling pathways on DSS-induced inflammatory bowel disease. Food Biosci. 2022, 47, 101426. [Google Scholar] [CrossRef]
- Sam, S.E.; Sim, K.S.; Rahman, S.N.S.A.; Tan, Y.S. Neuroprotective properties of wild medicinal mushroom, Sanguinoderma rugosum (Agaricomycetes), extracts against glutamate-induced hippocampal cells. Int. J. Med. Mushrooms 2022, 24, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.; Hsu, J.Y.; Chen, T.C.; Huang, C.C.; Wu, T.Y.; Chin, T.Y. Erinacine a prevents lipopolysaccharide-mediated glial cell activation to protect dopaminergic neurons against inflammatory factor-induced cell death in vitro and in vivo. Int. J. Mol. Sci. 2022, 23, 810. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Hernandez, S.; Blanco, J.; Garcia-Vallvé, S.; Pujadas, G.; Ojeda-Montes, M.J.; Gimeno, A.; Arola, L.; Minghetti, L.; Beltrán-Debón, R.; Mulero, M. Anti-inflammatory and immunomodulatory effects of the Grifola frondosa natural compound o-Orsellinaldehyde on LPS-challenged murine primary glial cells. Roles of NF-Κβ and MAPK. Pharmaceutics 2021, 13, 806. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.S.; Ooi, S.C.Y.; Lui, N.M.Y.; Qiong, C.; Ho, L.T.Y.; Cheah, I.K.M.; Halliwell, B.; Herr, D.R.; Ong, W.Y. Effect of ergothioneine on 7-ketocholesterol-induced endothelial injury. Neuromolecular Med. 2021, 23, 184. [Google Scholar] [CrossRef] [PubMed]
- He, M.T.; Park, C.H.; Cho, E.J. Caterpillar medicinal mushroom, Cordyceps militaris (Ascomycota), attenuates Aβ1–42–induced amyloidogenesis and inflammatory response by suppressing amyloid precursor protein progression and P38 MAPK/JNK Activation. Int. J. Med. Mushrooms 2021, 23, 71–83. [Google Scholar] [CrossRef]
- Nallathamby, N.; Lakshmanan, H.; Vidyadaran, S.; Malek, S.N.A.; Phan, C.W.; Sabaratnam, V. Lipid-rich fraction of the sclerotium of tiger milk mushroom Lignosus rhinocerotis (Agaricomycetes) attenuates LPS-induced inflammation in BV2 cells via Nrf2 Pathway. Braz. J. Pharm. Sci. 2020, 56. [Google Scholar] [CrossRef]
- Seow, S.L.S.; Naidu, M.; Sabaratnam, V.; Vidyadaran, S.; Wong, K.H. Tiger’s milk medicinal mushroom, Lignosus rhinocerotis (Agaricomycetes) sclerotium inhibits nitric oxide production in LPS-stimulated BV2 Microglia. Int. J. Med. Mushrooms 2017, 19, 405–418. [Google Scholar] [CrossRef]
- Scuto, M.; di Mauro, P.; Ontario, M.L.; Amato, C.; Modafferi, S.; Ciavardelli, D.; Salinaro, A.T.; Maiolino, L.; Calabrese, V. Nutritional mushroom treatment in Meniere’s disease with Coriolus versicolor: A rationale for therapeutic intervention in neuroinflammation and antineurodegeneration. Int. J. Mol. Sci. 2020, 21, 284. [Google Scholar] [CrossRef] [Green Version]
- Trovato-Salinaro, A.; Siracusa, R.; di Paola, R.; Scuto, M.; Fronte, V.; Koverech, G.; Luca, M.; Serra, A.; Toscano, M.A.; Petralia, A.; et al. Redox modulation of cellular stress response and lipoxin A4 expression by Coriolus versicolor in rat brain: Relevance to Alzheimer’s disease pathogenesis. Neurotoxicology 2016, 53, 350–358. [Google Scholar] [CrossRef]
- Sheng, F.; Zhang, L.; Wang, S.; Yang, L.; Li, P. Deacetyl ganoderic acid F inhibits LPS-induced neural inflammation via NF-ΚB pathway both in vitro and in vivo. Nutrients 2020, 12, 85. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Zhu, S.; Cheng, P.; Lu, Z.M.; Xu, H.Y.; Shi, J.S.; Xu, Z.H. Bioassay-guided fractionation of ethyl acetate extract from Armillaria mellea attenuates inflammatory response in lipopolysaccharide (LPS) stimulated BV-2 microglia. Phytomedicine 2017, 26, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Moresco, E.M.Y.; LaVine, D.; Beutler, B. Toll-like Receptors. Curr. Biol. 2011, 21, R488–R493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltschmidt, B.; Kaltschmidt, C. NF-B in the nervous system. Cold Spring Harb. Perspect. Biol. 2009, 1, a001271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahay, A.; Hen, R. Adult hippocampal neurogenesis in depression. Nat. Neurosci. 2007, 10, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M. Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch. Gen. Psychiatry 2000, 57, 925–935. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, M.C.; Yucel, K.; Nazarov, A.; MacQueen, G.M. A meta-analysis examining clinical predictors of hippocampal volume in patients with Major Depressive Disorder. J. Psych. Neurosci. 2009, 34, 41–54. [Google Scholar]
- Belleau, E.L.; Treadway, M.T.; Pizzagalli, D.A. The impact of stress and Major Depressive Disorder on hippocampal and medial prefrontal cortex morphology. Biol. Psychiatry 2019, 85, 443–453. [Google Scholar] [CrossRef]
- Amico, F.; Meisenzahl, E.; Koutsouleris, N.; Reiser, M.; Möller, H.J.; Frodl, T. Structural MRI correlates for vulnerability and resilience to Major Depressive Disorder. J. Psych. Neurosci. 2011, 36, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.C.; Hamilton, J.P.; Gotlib, I.H. Decreased hippocampal volume in healthy girls at risk of depression. Arch. Gen. Psychiatry 2010, 67, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Taylor, W.D.; McQuoid, D.R.; Payne, M.E.; Zannas, A.S.; MacFall, J.R.; Steffens, D.C. Hippocampus atrophy and the longitudinal course of late-life depression. Am. J. Geriatr. Psychiatry 2014, 22, 1504–1512. [Google Scholar] [CrossRef] [Green Version]
- Frodl, T.S.; Koutsoulcris, N.; Bottlender, R.; Born, C.; Jäger, M.; Scupin, I.; Reiser, M.; Möller, H.J.; Meiscnzahl, E.M. Depression-related variation in brain morphology over 3 years: Effects of stress? Arch. Gen. Psychiatry 2008, 65, 1156–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papagni, S.A.; Benetti, S.; Arulanantham, S.; McCrory, E.; McGuire, P.; Mechelli, A. Effects of stressful life events on human brain structure: A longitudinal voxel-based morphometry study. Stress 2011, 14, 227–232. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Rauskolb, S.; Polack, M.; Klose, J.; Kolbeck, R.; Korte, M.; Barde, Y.A. Biosynthesis and processing of endogenous BDNF: CNS neurons store and secrete BDNF, not pro-BDNF. Nat. Neurosci. 2008, 11, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Siao, C.J.; Nagappan, G.; Marinic, T.; Jing, D.; McGrath, K.; Chen, Z.Y.; Mark, W.; Tessarollo, L.; Lee, F.S.; et al. Neuronal release of ProBDNF. Nat. Neurosci. 2009, 12, 113–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, M. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Frade, J.M.; Rodriguez-Tebar, A.; Barde, Y.A. Induction of cell death by endogenous Nerve Growth Factor through its P75 receptor. Nature 1996, 383, 166–168. [Google Scholar] [CrossRef]
- Lee, R.; Kermani, P.; Teng, K.K.; Hempstead, B.L. Regulation of cell survival by secreted proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [Green Version]
- Drevets, W.C.; Price, J.L.; Furey, M.L. Brain structural and functional abnormalities in mood disorders: Implications for neurocircuitry models of depression. Brain Struct. Funct. 2008, 213, 93–118. [Google Scholar] [CrossRef] [Green Version]
- MacQueen, G.; Frodl, T. The hippocampus in major depression: Evidence for the convergence of the bench and bedside in psychiatric research? Mol. Psych. 2010, 16, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and Rapid-Acting Antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y. Brain-Derived Neurotrophic Factor: Role in depression and suicide. Neuropsychiatr. Dis. Treat. 2009, 5, 433–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinowich, K.; Manji, H.; Lu, B. New insights into BDNF function in depression and anxiety. Nat. Neurosci. 2007, 10, 1089–1093. [Google Scholar] [CrossRef]
- Zhao, S.; Rong, C.; Gao, Y.; Wu, L.; Luo, X.; Song, S.; Liu, Y.; Wong, J.H.; Wang, H.; Yi, L.; et al. Antidepressant-like effect of Ganoderma lucidum spore polysaccharide-peptide mediated by upregulation of prefrontal cortex Brain-Derived Neurotrophic Factor. Appl. Microbiol. Biotechnol. 2021, 105, 8675–8688. [Google Scholar] [CrossRef]
- Ma, B.-J.; Shen, J.-W.; Yu, H.-Y.; Ruan, Y.; Wu, T.-T.; Zhao, X. Hericenones and erinacines: Stimulators of Nerve Growth Factor (NGF) biosynthesis in Hericium erinaceus. Mycology 2010, 1, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Li, I.C.; Lee, L.Y.; Tzeng, T.T.; Chen, W.P.; Chen, Y.P.; Shiao, Y.J.; Chen, C.C. Neurohealth properties of Hericium erinaceus mycelia enriched with erinacines. Behav. Neurol. 2018, 2018, 5802634. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.F.; Chen, J.H.; Teng, C.C.; Shen, C.H.; Hsieh, M.C.; Lu, C.C.; Lee, K.C.; Lee, L.Y.; Chen, W.P.; Chen, C.C.; et al. Protective effects of Hericium erinaceus mycelium and its isolated erinacine A against ischemia-injury-induced neuronal cell death via the inhibition of INOS/P38 MAPK and nitrotyrosine. Int. J. Mol. Sci. 2014, 15, 15073–15089. [Google Scholar] [CrossRef] [Green Version]
- Chong, P.S.; Poon, C.H.; Roy, J.; Tsui, K.C.; Lew, S.Y.; Phang, M.W.L.; Tan, R.J.Y.; Cheng, P.G.; Fung, M.L.; Wong, K.H.; et al. Neurogenesis-dependent antidepressant-like activity of Hericium erinaceus in an animal model of depression. Chin. Med. 2021, 16, 132. [Google Scholar] [CrossRef]
- Ryu, S.H.; Hong, S.M.; Khan, Z.; Lee, S.K.; Vishwanath, M.; Turk, A.; Yeon, S.W.; Jo, Y.H.; Lee, D.H.; Lee, J.K.; et al. Neurotrophic isoindolinones from the fruiting bodies of Hericium erinaceus. Bioorg. Med. Chem. Lett. 2021, 31, 127714. [Google Scholar] [CrossRef]
- Nagano, M.; Shimizu, K.; Kondo, R.; Hayashi, C.; Sato, D.; Kitagawa, K.; Ohnuki, K. Reduction of depression and anxiety by 4 weeks Hericium erinaceus intake. Biomed. Res. 2010, 31, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.C.; Lin, T.C.; Chen, C.C.; Lee, L.Y.; Chen, W.P.; Liu, Y.Z.; Hwang, J.H. Effects of erinacine A-enriched Hericium erinaceus on elderly hearing-impaired patients: A double-blind, randomized, placebo-controlled clinical trial. J. Funct. Foods 2022, 97, 105220. [Google Scholar] [CrossRef]
- Wittstein, K.; Rascher, M.; Rupcic, Z.; Löwen, E.; Winter, B.; Köster, R.W.; Stadler, M. Corallocins A-C, Nerve Growth and Brain-Derived Neurotrophic Factor inducing metabolites from the mushroom Hericium coralloides. J. Nat. Prod. 2016, 79, 2264–2269. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.-W.; Xia, B.; Han, R.; Li, Z.-Q.; Yang, J.-R.; Yin, X.; Gao, Y.-Q.; Gao, J.-M. Neuroprotective effects of a new triterpenoid from edible mushroom on oxidative stress and apoptosis through the BDNF/TrkB/ERK/CREB and Nrf2 signaling pathway in vitro and in vivo. Food Funct. 2022, 13, 12121–12134. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhang, P.; Wang, R.; Zhou, M.; Pang, N.; Cui, X.; Ge, X.; Liu, X.; Huang, X.F.; Yu, Y. Three different types of β-glucans enhance cognition: The role of the gut-brain axis. Front. Nutr. 2022, 9, 312. [Google Scholar] [CrossRef]
- Hashim, H.; Azmin, S.; Razlan, H.; Yahya, N.W.; Tan, H.J.; Manaf, M.R.A.; Ibrahim, N.M. Eradication of Helicobacter pylori infection improves levodopa action, clinical symptoms and quality of life in patients with Parkinson’s Disease. PLoS ONE 2014, 9, e112330. [Google Scholar] [CrossRef] [Green Version]
- Rafii, F.; Sutherland, J.B.; Hansen, E.B.; Cerniglia, C.E. Reduction of nitrazepam by Clostridium leptum, a nitroreductase-producing bacterium isolated from the human intestinal tract. Clin. Infect. Dis. 1997, 25 (Suppl. 2), S121–S122. [Google Scholar] [CrossRef] [Green Version]
- Elmer, G.W.; Remmel, R.P. Role of the intestinal microflora in clonazepam metabolism in the rat. Xenobiotica 1984, 14, 829–840. [Google Scholar] [CrossRef]
- Kitamura, S.; Sugihara, K.; Kuwasako, M.; Tatsumi, K. The role of mammalian intestinal bacteria in the reductive metabolism of zonisamide. J. Pharm. Pharmacol. 1997, 49, 253–256. [Google Scholar] [CrossRef]
- Roy Sarkar, S.; Banerjee, S. Gut microbiota in neurodegenerative disorders. J. Neuroimmunol. 2019, 328, 98–104. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. New frontiers for treatment of metabolic diseases. Mediat. Inflamm. 2018, 2018, 2037838. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− Mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, C.; Ijaz, U.Z.; Gallagher, E.; Horton, F.; Ellis, R.J.; Jaiyeola, E.; Duparc, T.; Russell-Jones, D.; Hinton, P.; Cani, P.D.; et al. Fecal Enterobacteriales enrichment is associated with increased in vivo intestinal permeability in humans. Physiol. Rep. 2018, 6, e13649. [Google Scholar] [CrossRef] [PubMed]
- Bengmark, S. Gut microbiota, immune development and function. Pharmacol. Res. 2013, 69, 87–113. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Belzer, C.; Goossens, M.; Kleerebezem, M.; de Vos, W.M.; Thas, O.; de Weirdt, R.; Kerckhof, F.M.; van de Wiele, T. Butyrate-producing Clostridium Cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 2013, 7, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; de Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Sakon, H.; Nagai, F.; Morotomi, M.; Tanaka, R. Sutterella parvirubra Sp. Nov. and Megamonas funiformis Sp. Nov., Isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 970–975. [Google Scholar] [CrossRef] [Green Version]
- Stilling, R.M.; Dinan, T.G.; Cryan, J.F. Microbial genes, brain & behaviour–epigenetic regulation of the gut-brain axis. Genes Brain Behav. 2014, 13, 69–86. [Google Scholar] [CrossRef]
- Li, N.; Wang, Q.; Wang, Y.; Sun, A.; Lin, Y.; Jin, Y.; Li, X. Fecal microbiota transplantation from chronic unpredictable mild stress mice donors affects anxiety-like and depression-like behavior in recipient mice via the gut microbiota-inflammation-brain axis. Stress 2019, 22, 592–602. [Google Scholar] [CrossRef]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with Major Depressive Disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef]
- Li, C. Understanding interactions among diet, host and gut microbiota for personalized nutrition. Life Sci. 2023, 312, 121265. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, K.P.; Tuohy, K.M.; Mach -Istituto, F.E.; San, A.; All’adige, M.; Gibson, G.R.; Rastall, R.A.; Hotchkiss, A.; Dubert-Ferrandon, A.; Gareau, M.; et al. Bioactive food ingredients for safe and health-promoting functional foods view project neurodevelopmental impact of preterm birth in a porcine model view project dietary prebiotics: Current status and new definition. Food Sci. Technol. Bull. 2010, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lei, L.; Zhou, Y.; Ye, F.; Zhao, G. Roles of mushroom polysaccharides in chronic disease management. J. Integr. Agric. 2022, 21, 1839–1866. [Google Scholar] [CrossRef]
- Li, M.; Yu, L.; Zhai, Q.; Liu, B.; Zhao, J.; Chen, W.; Tian, F. Ganoderma lucidum ethanol extraction promotes dextran sulphate sodium induced colitis recovery and modulation in microbiota. Foods 2022, 11, 4023. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Wang, Z.; Jiang, H.; Yan, R.; Huang, Q.; Wang, Y.; Xie, H.; Zou, Y.; Yu, Y.; Lv, L. Ganoderma lucidum promotes sleep through a gut microbiota-dependent and serotonin-involved pathway in mice. Sci. Rep. 2021, 11, 13660. [Google Scholar] [CrossRef]
- Romero-Córdoba, S.L.; Salido-Guadarrama, I.; Meneses, M.E.; Cosentino, G.; Iorio, M.V.; Tagliabue, E.; Torres, N.; Sánchez-Tapia, M.; Bonilla, M.; Castillo, I.; et al. Mexican Ganoderma lucidum extracts decrease lipogenesis modulating transcriptional metabolic networks and gut microbiota in C57BL/6 mice fed with a high-cholesterol diet. Nutrients 2020, 13, 38. [Google Scholar] [CrossRef]
- Chen, M.; Xiao, D.; Liu, W.; Song, Y.; Zou, B.; Li, L.; Li, P.; Cai, Y.; Liu, D.; Liao, Q.; et al. Intake of Ganoderma lucidum polysaccharides reverses the disturbed gut microbiota and metabolism in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 155, 890–902. [Google Scholar] [CrossRef]
- Lu, C.-L.; Lee, B.H.; Ren, Y.-L.; Ji, D.; Rao, S.-Q.; Li, H.-X.; Yang, Z.-Q. Effects of exopolysaccharides from Antrodia cinnamomea on inflammation and intestinal microbiota disturbance induced by antibiotics in mice. Food Biosci. 2022, 50, 102116. [Google Scholar] [CrossRef]
- Ye, J.; Zhang, C.; Fan, Q.; Lin, X.; Wang, Y.; Azzam, M.; Alhotan, R.; Alqhtani, A.; Jiang, S. Antrodia cinnamomea polysaccharide improves liver antioxidant, anti-inflammatory capacity, and cecal flora structure of slow-growing broiler breeds challenged with lipopolysaccharide. Front. Vet. Sci. 2022, 9, 1559. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.U.; Khan, A.I.; Xin, Y.; Liang, W. Morchella esculenta polysaccharide attenuate obesity, inflammation and modulate gut microbiota. AMB Express 2022, 12, 114. [Google Scholar] [CrossRef]
- Wang, T.; Han, J.; Dai, H.; Sun, J.; Ren, J.; Wang, W.; Qiao, S.; Liu, C.; Sun, L.; Liu, S.; et al. Polysaccharides from Lyophyllum decastes reduce obesity by altering gut microbiota and increasing energy expenditure. Carbohydr. Polym. 2022, 295, 119862. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Yu, H.; Deng, H.; Fang, Q.; Lei, H.; Liu, L.; Wu, N.; Guo, X.; Song, C. Three main metabolites from Wolfiporia cocos (F. A. Wolf) Ryvarden & Gilb regulate the gut microbiota in mice: A comparative study using microbiome-metabolomics. Front. Pharmacol. 2022, 13, 2852. [Google Scholar] [CrossRef]
- Gao, X.; Feng, Y.; Xue, H.; Meng, M.; Qin, X. Antidepressant-like effect of triterpenoids extracts from Poria Cocos on the CUMS rats by 16S RRNA gene sequencing and LC–MS metabolomics. J. Liq. Chromatogr. Relat. Technol. 2020, 43, 494–507. [Google Scholar] [CrossRef]
- Huang, Y.; Gao, Y.; Pi, X.; Zhao, S.; Liu, W. In vitro hepatoprotective and human gut microbiota modulation of polysaccharide-peptides in Pleurotus citrinopileatus. Front. Cell Infect. Microbiol. 2022, 12, 892049. [Google Scholar] [CrossRef]
- Nakahara, D.; Nan, C.; Mori, K.; Hanayama, M.; Kikuchi, H.; Hirai, S.; Egashira, Y. Effect of mushroom polysaccharides from Pleurotus eryngii on obesity and gut microbiota in mice fed a high-fat diet. Eur. J. Nutr. 2020, 59, 3231–3244. [Google Scholar] [CrossRef]
- Hu, Q.; Yuan, B.; Wu, X.; Du, H.; Gu, M.; Han, Y.; Yang, W.; Song, M.; Xiao, H. Dietary intake of Pleurotus eryngii ameliorated dextran-sodium-sulfate-induced colitis in mice. Mol. Nutr. Food Res. 2019, 63, 1801265. [Google Scholar] [CrossRef]
- Hu, Y.; Xu, J.; Sheng, Y.; Liu, J.; Li, H.; Guo, M.; Xu, W.; Luo, Y.; Huang, K.; He, X. Pleurotus ostreatus ameliorates obesity by modulating the gut microbiota in obese mice induced by high-fat diet. Nutrients 2022, 14, 1868. [Google Scholar] [CrossRef]
- Adams, S.; Che, D.; Hailong, J.; Zhao, B.; Rui, H.; Danquah, K.; Qin, G. Effects of pulverized oyster mushroom (Pleurotus ostreatus) on diarrhea incidence, growth performance, immunity, and microbial composition in piglets. J. Sci. Food Agric. 2019, 99, 3616–3627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maheshwari, G.; Gessner, D.K.; Neuhaus, K.; Most, E.; Zorn, H.; Eder, K.; Ringseis, R. Influence of a biotechnologically produced oyster mushroom (Pleurotus Sajor-Caju) on the gut microbiota and microbial metabolites in obese Zucker rats. J. Agric. Food Chem. 2021, 69, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.; Wang, Q.; Gould, T.; Slavin, J. Impact of Agaricus bisporus mushroom consumption on gut health markers in healthy adults. Nutrients 2018, 10, 1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Sanmartín, J.; Bobadilla, M.; Mirpuri, E.; Grifoll, V.; Pérez-Clavijo, M.; Martínez, A. Agaricus mushroom-enriched diets modulate the microbiota-gut-brain axis and reduce brain oxidative stress in mice. Antioxidants 2022, 11, 695. [Google Scholar] [CrossRef]
- Yao, L.; Lv, J.; Duan, C.; An, X.; Zhang, C.; Li, D.; Li, C.; Liu, S. Armillaria mellea fermentation liquor ameliorates p-chlorophenylalanine-induced insomnia associated with the modulation of serotonergic system and gut microbiota in rats. J. Food Biochem. 2022, 46, e14075. [Google Scholar] [CrossRef]
- Yang, X.; Zheng, M.; Zhou, M.; Zhou, L.; Ge, X.; Pang, N.; Li, H.; Li, X.; Li, M.; Zhang, J.; et al. Lentinan supplementation protects the gut–liver axis and prevents steatohepatitis: The role of gut microbiota involved. Front. Nutr. 2022, 8, 1286. [Google Scholar] [CrossRef]
- Zou, M.; Wang, Y.; Liu, Y.; Xiong, S.; Zhang, L.; Wang, J. Huangshan floral mushroom polysaccharide ameliorates dextran sulfate sodium-induced colitis in mice by modulating Th17/Treg balance in a gut microbiota-dependent manner. Mol. Nutr. Food Res. 2022, 67, 2200408. [Google Scholar] [CrossRef]
- Cho, H.-W.; Choi, S.; Seo, K.; Kim, K.H.; Jeon, J.-H.; Kim, C.H.; Lim, S.; Jeong, S.; Chun, J.L. Gut microbiota profiling in aged dogs after feeding pet food contained Hericium erinaceus. J. Anim. Sci. Technol. 2022, 64, 937–949. [Google Scholar] [CrossRef]
- Xie, X.Q.; Geng, Y.; Guan, Q.; Ren, Y.; Guo, L.; Lv, Q.; Lu, Z.M.; Shi, J.S.; Xu, Z.H. Influence of short-term consumption of Hericium erinaceus on serum biochemical markers and the changes of the gut microbiota: A pilot study. Nutrients 2021, 13, 1008. [Google Scholar] [CrossRef]
- Lai, Y.; Fang, Q.; Guo, X.; Lei, H.; Zhou, Q.; Wu, N.; Song, C. Effect of polysaccharides from Dictyophora indusiata on regulating gut microbiota and short-chain fatty acids in mice. J. Food Meas. Charact. 2022, 17, 1–11. [Google Scholar] [CrossRef]
- Kanwal, S.; Aliya, S.; Xin, Y. Anti-obesity effect of Dictyophora indusiata mushroom polysaccharide (DIP) in high fat diet-induced obesity via regulating inflammatory cascades and intestinal microbiome. Front. Endocrinol. 2020, 11, 558874. [Google Scholar] [CrossRef] [PubMed]
- Abdureyim, Z.; Wang, L.; Tao, Q.; Xu, J.; Yimiti, D.; Gao, Q. Bachu mushroom polysaccharide alleviates colonic injury by modulating the gut microbiota. Comput. Math. Methods Med. 2022, 2022, 1353724. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xie, L.; Zhang, Z.; Zhang, W.; Tang, J.; He, X.; Zhou, J.; Peng, W. Tremella fuciformis polysaccharides inhibited colonic inflammation in dextran sulfate sodium-treated mice via Foxp3+ T cells, gut microbiota, and bacterial metabolites. Front. Immunol. 2021, 12, 948. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Guo, W.L.; Zhang, W.; Xu, J.X.; Qian, M.; Bai, W.D.; Zhang, Y.Y.; Rao, P.F.; Ni, L.; Lv, X.C. Grifola frondosa polysaccharides ameliorate lipid metabolic disorders and gut microbiota dysbiosis in high-fat diet fed rats. Food Funct. 2019, 10, 2560–2572. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, D.; Wang, D.; Lai, S.; Zhong, R.; Liu, Y.; Yang, C.; Liu, B.; Sarker, M.R.; Zhao, C. Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food Chem. Toxicol. 2019, 126, 295–302. [Google Scholar] [CrossRef]
- Finotello, F.; Mastrorilli, E.; di Camillo, B. Measuring the diversity of the human microbiota with targeted next-generation sequencing. Brief. Bioinform. 2018, 19, 679–692. [Google Scholar] [CrossRef]
- Di Pierro, F. Gut microbiota parameters potentially useful in clinical perspective. Microorganisms 2021, 9, 2402. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Iintestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Kerckhoffs, A.P.M.; Akkermans, L.M.A.; de Smet, M.B.M.; Besselink, M.G.H.; Hietbrink, F.; Bartelink, I.H.; Busschers, W.B.; Samsom, M.; Renooij, W. Intestinal permeability in irritable bowel syndrome patients: Effects of NSAIDs. Dig. Dis. Sci. 2010, 55, 716–723. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Madison, A.; Kiecolt-Glaser, J.K. Stress, depression, diet, and the gut microbiota: Human–bacteria interactions at the core of psychoneuroimmunology and nutrition. Curr. Opin. Behav. Sci. 2019, 28, 105. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, L.; Gustafsson, A.; Lavant, E.; Suneson, K.; Brundin, L.; Westrin, Å.; Ljunggren, L.; Lindqvist, D. Leaky gut biomarkers in depression and suicidal behavior. Acta Psychiatr. Scand. 2019, 139, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirce, J.M.; Alviña, K. The role of inflammation and the gut microbiome in depression and anxiety. J. Neurosci. Res. 2019, 97, 1223–1241. [Google Scholar] [CrossRef] [Green Version]
- Kris-Etherton, P.M.; Petersen, K.S.; Hibbeln, J.R.; Hurley, D.; Kolick, V.; Peoples, S.; Rodriguez, N.; Woodward-Lopez, G. Nutrition and behavioral health disorders: Depression and anxiety. Nutr. Rev. 2021, 79, 247–260. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Martínez-González, M.A.; Estruch, R.; Salas-Salvadó, J.; Corella, D.; Covas, M.I.; Arós, F.; Romaguera, D.; Gómez-Gracia, E.; Lapetra, J.; et al. Mediterranean dietary pattern and depression: The PREDIMED Randomized Trial. BMC Med. 2013, 11, 208. [Google Scholar] [CrossRef] [Green Version]
- Appleton, K.M.; Sallis, H.M.; Perry, R.; Ness, A.R.; Churchill, R. Omega-3 fatty acids for depression in adults. Cochrane Database Syst. Rev. 2015, 2015, CD004692. [Google Scholar] [CrossRef] [Green Version]
- Sofi, F.; Macchi, C.; Abbate, R.; Gensini, G.F.; Casini, A. Mediterranean diet and health. BioFactors 2013, 39, 335–342. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Source | L-Tryptophan (mg/100 g dw) | Tryptamine (mg/100 g dw) | 5-Hydroxy-L-tryptophan (mg/100 g dw) | Ref. |
|---|---|---|---|---|---|
| Pleurotus citrinopileatus | Fruiting bodies form commercial cultivation | 1.29 ± 0.08 | 1.29 ± 0.08 | 128.89 ± 10.67 | [39] |
| Pleurotus djamor | 24.84 ± 0.97 | 3.54 ± 0.18 | 193.95 ± 17.69 | ||
| Pleurotus ostreatus | 5.79 ± 0.06 | 1.04 ± 0.15 | 67.45 ± 7.94 | ||
| Auricularia cornea | 0.16 ± 0.050 | 2.77 ± 0.050 | 7.35 ± 0.200 | [40] | |
| Armillaria mellea | Fruiting bodies from natural habitats | 4.47 ± 0.01 | 2.74 ± 0.01 | - z | [41] |
| Imleria badia | 0.68 ± 0.047 | 0.18 ± 0.001 | - z | ||
| Boletus edulis | 0.39 ± 0.020 | 1.17 ± 0.003 | 0.18 ± 0.001 | ||
| Lactarius deliciosus | - z | - z | 0.25 ± 0.017 | ||
| Hydnum repandum | 0.37 ± 0.09 | 1.46 ± 1.05 | n/a | [42] | |
| Cantharellus cibarius | 0.01 ± 0.002 | 0.01 ± 0.002 | 0.02 ± 0.003 | [43] | |
| Agaricus bisporus | 0.39 ± 0.020 | 0.06 ± 0.003 | - z | ||
| Sarcodon imbricatus | 13.01 ± 0.01 | 22.12 ± 0.03 | n/a | ||
| Macrolepiota procera | 3.47 ± 0.050 | 0.92 ± 0.040 | 22.94 ± 0.500 | [40] | |
| Suillus bovinus | 25.90 ± 0.200 | 3.15 ± 0.050 | 15.83 ± 0.500 | ||
| Tricholoma equestre | 2.851 ± 0.242 | 2.011 ± 0.141 | 0.586 ± 0.041 | [44] | |
| Calocera viscosa | 1.26 ± 0.04 | n/a | 11.88 ± 0.19 |
| Species | L-Tryptophan (mg/100 g dw) | Tryptamine (mg/100 g dw) | 5-Hydroxy-L-tryptophan (mg/100 g dw) | Ref. |
|---|---|---|---|---|
| Pleurotus citrinopileatus | 7.82 ± 0.31 | 3.71 ± 0.34 | 368.67 ± 23.53 | [39] |
| Pleurotus djamor | 24.34 ± 1.44 | n/d | 703.56 ± 37.79 | |
| Pleurotus ostreatus | 1.89 ± 0.04 | 1.03 ± 0.15 | 120.11 ± 20.12 | |
| Agaricus bisporus | 14.00 ± 0.300 | 0.48 ± 0.050 | 12.50 ± 0.671 | [51] |
| Sarcodon imbricatus | 1.31 ± 0.6 | 4.11 ± 0.26 | n/a | [52] |
| Tricholoma equestre | 1.036 ± 0.093 | 0.598 ± 0.048 | 0.344 ± 0.031 | [44] |
| Imleria badia | 0.827 ± 0.049 | 0.409 ± 0.045 | - z | |
| Calocera viscosa | 1.79 ± 0.05 | n/a | 11.42 ± 0.20 | [53] |
| Cantharellus cibarius | 0.64 ± 0.013 | - z | 12.52 ± 0.671 | [54] |
| Species | Active Substance | Experience Model | Result | Ref. |
|---|---|---|---|---|
| Lentinula edodes | β-glucan | In vivo: C57BL/6J mice Inflammation: High-fat diet-induced (HFD) | ↓Neuroinflammation in hippocampus and prefrontal cortex
| [107] |
| Lentinan | In vitro: mice microglia BV–2 cell line Inflammation: LPS induced | ↓Microglia inflammation
| [108] | |
| Inonotus obliquus | Lanostane triterpenes: Inonotusols H-N | In vitro: mice microglia BV–2 cell line Inflammation: LPS induced | ↓Microglia inflammation
| [109] |
| Cyathus africanus | Cyanthane diterpenoids | In vitro: mice microglia BV–2 cell line, PC–12 cell line Inflammation: LPS induced | ↓Microglia inflammation
| [110] |
| Auricularia polytricha | Ethanol and hexane extracts | In vitro: mice microglia BV–2 cell line Inflammation: BPA induced | ↓Neuroinflammation by regulation of NG-kB signaling pathway ↑Antioxidant activity
| [111] |
| Sanguinoderma rugosum | Mycelial extracts | In vitro: mice hippocampal neuronal HT–22 cell line Inflammation: glutamate induced | ↑Cell viability in pretreated with mycelial extract cell line after glutamate inflammation induction The main constituents responsible for the action were: linoleic acid, ergosterol and ethyl linoleate (GC-MS analysis) | [112] |
| Hericium erinaceus | Erinacine A (EA)/H. erinaceus extract (HEM) | In vivo: male Sprague-Dawley rats Inflammation: LPS injection intranigrally In vitro: mice microglia BV–2 cell line, CTX TNA2 cell line Inflammation: LPS, IFN-γ induced | In vivo: Both, EA and HEM showed neuroprotective effect on motor dysfunction in rats with LPS-induced neuronal damage. ↓Neuroinflammation
In vitro: ↓Microglia inflammation
| [113] |
| Grifola frondosa | o-Orsellinaldehyde | In vitro: primary microglia and astrocytes from mice brain Inflammation: LPS induced | ↓Neuroinflammation
| [114] |
| - | Ergothioneine | In vitro: hCMEC/D3 human brain endothelial cell line Inflammation: 7-ketocholesterol induced | ↓Neuroinflammation ↓IL-1β, ↓IL-6, ↓IL-8, ↓TNF-α, ↓COX-2 mRNA expression after ergothioneine addition | [115] |
| Cordyceps militaris | C. militaris powdered fruiting bodies | In vivo: AD mice Inflammation: intracerebroventricular injection of Aβ1−42 | ↓Neuroinflammation
| [116] |
| Lignosus rhinocerotis | Lipid rich fraction of the sclerotium | In vitro: mice microglia BV–2 cell line Inflammation: LPS induced | ↓Neuroinflammation
| [117] |
| Hot aqueous extract (HAE), an ethanol extract (EE), fractions from the HAE and EE, crude polysaccharides | In vitro: mice microglia BV–2 cell line Inflammation: LPS induced | ↓Neuroinflammation 500 μg/mL of HAE → ↓NO production by 88.95% * 250 μg/mL of an n-butanol fraction → ↓NO production by 86.5% * 250 μg/mL of an ethyl acetate fraction of HAE → ↓NO production by 85.93% * * Compared to control group | [118] | |
| Coriolus versicolor | C. versicolor biomass (mycelium and primordia) | Clinical trial: 40 patients with Meniere’s disease (MD) | ↓Oxidation stress in group treated with Coriolus supplement ↑Hsp70, ↑HO-1 proteins level in lymphocytes and plasma ↑γ-GC liase activity ↑GSH/GSSG ratio in patients treated with Coriolus ↑GSH level in patient treated group | [119] |
| C. versicolor biomass (mycelium and primordia) | In vivo: male Sprague–Dawley rats Inflammation: no induction | ↑Neuroprotection ↑LXA4 level in rat brain, especially in cortex and hippocampus ↑Hsp72, ↑OH–1, ↑thioredoxin proteins level | [120] | |
| Ganoderma lucidum | Deacetyl ganoderic acid F (DeGA F) | In vivo: zebrafish and C57BL/6J mice Inflammation: LPS induced In vitro: mice microglia BV–2 cell line Inflammation: LPS induced | In vivo: ↓Inflammation in zebrafish
In vitro: ↓Neuroinflammation through NF-κB signaling pathway ↓p65 nuclear protein level and migration reduction ↓p-Akt, ↓p-IKKα/β, ↓p-IκBα expression | [121] |
| Armillaria mellea | Fr.2 fraction (5-hydroxymethylfurfural, vanillic acid, syringate) | In vitro: mice microglia BV–2 cell line Inflammation: LPS induced | ↓Neuroinflammation
| [122] |
| Medicinal Mushroom Species | Active Ingredient | Experimental Model | Gut Microbiota Alteration in Treated Group | Biological Activity | Ref. |
|---|---|---|---|---|---|
| Ganoderma lucidum | Ethanolic extract | In vivo: C57BL6/J mice Dextran sulphate sodium administration (DSS)-induced colitis | ↑Turicibacter ↑Bifidobacterium ↑Parabacteroides abudance ↓Escherichia/Shigella ↓Bacteroides ↓Staphylococcus abudance | Restoration of the intestinal barrier
| [178] |
| Ethanolic extract of G. lucidum mycelium (GLAA) | In vivo: ICR mice, SPF Gut microbiota depleted by antibiotic | ↑Bifidobacteriaceae Bidifobacterium ↑Lactobacillaceae Lactobacullus (L. reuteri) ↑Porphyromonadaceae Odoribacter ↑Erysipelotrichales Turicbacter ↓Ruminococcaceae Oscillibacter ↓Anaerotignum ↓Roseburia | Addition of GLAA reduced required time for sleep induction by 50% in pentobarbital-induced hypnosis model ↑5-HT in the hypothalamus in treated with GLAA group (50 and 100 mg/kg) | [179] | |
| Hydroalcoholic extracts | In vivo: C57BL/6 mice High-cholesterol diet (HCD) | Reversed gut dysbiosis caused by HCD ↑Lactobacillus ↓Bacteroides acidifaciens ↓Mucispirillum schaedleri ↓Parabacteroides distasonis | G. lucidum extracts revealed to be a novel transcriptome modulator—prevention of metabolic disorders associated with hypercholesterolemia | [180] | |
| Polysaccharides from hot water extraction | In vivo: Sprague-Dawley rats (SPF) HFD and streptozotocin-induced type 2 diabetes mellitus | ↑Blautia ↑Dehalobacterium ↑Parabacteroides ↑Bacteroides ↓Aerococcus ↓Ruminococcus ↓Corynebactrium ↓Proteus | Administration of mushroom polysaccharides showed antidiabetic effect | [181] | |
| Antrodia cinnamomea | Exopolysaccharides | In vivo: ICR mice Antibiotics induced gut dysbiosis (lincomycin) | ↑Lactobacillus ↑Roseburia ↑Ligilactobacillus ↑Lachnospiraceae_NK4A136 group abudance ↓Enterococcus Shigella abudance | Reduced peripheral inflammation ↓IL–6 and ↓TNF-a serum levels compared to control group | [182] |
| Isolated polysaccharides | In vivo: Lingnan yellow-heathered female chickens LPS induced liver inflammation | Restrained the decline of beneficial cecal microbiota (typically Lactobacillus, Faecalibacterium, and Christensenellaceae R-7 group) | Hepatoprotective effect—reversed LPS-induced liver inflammation ↓TLR4/NF-κB signaling pathway expression in the liver tissues | [183] | |
| Morchella esculenta | Isolated polysaccharides | In vivo: BALC/c mice (SPF) HFD | ↑Lactobacillus ↓Enterococcus | Improved intestinal barrier
| [184] |
| Lyophyllum decastes | Isolated polysaccharides | In vivo: C57BL/7J mice HFD | ↓Firmictues/Bacteroidetes ratio ↑Lactobacillus johnsonii Bacteroides intestinalis | Antiobestity effect of extracted polysaccharides due to microbiota alteration and secondary bile-acids production activation | [185] |
| Wolfiporia cocos | Including water-soluble polysaccharides (PCX), water-insoluble polysaccharides (PCY), and triterpenoid saponins (PCZ) | In vivo: healthy Kunming mice | The strongest altering gut microbiota activity was found in PCY fraction: ↑Lactobacillus In mice fed with PCX: ↑Deferribacterota | PCX: ↑IL–10 levels in the liver, spleen tissues and serum, PCY: ↓IFN–γ level in the liver, PCZ: ↓TNF-α level in liver and spleen PCX and PCZ: ↑IFN–γ | [186] |
| Triterpenoid fraction | In vivo: Spague–Daley rats Unpredictable mild stress model (CUMS) | Restoration of altered gut microbiota by CUMS: ↓Firmicutes/Bacteroidetes ratio | ↑BDNF, ↑NGF in the hippocampus | [187] | |
| Auricularia polytricha | Freeze dried ethanolic extracts of fruiting bodies | In vivo: ICR mice (SPF) DSS-induced intestinal dysbiosis | ↑Ruminococcaceae ↑Lachnospiraceae ↑Prevotellaceae ↑Bifidobacteriaceae ↑Erysipelotrichaceae | Anti-inflammatory activity: ↓F-κB and MAPK/ ERK1/2 signaling pathway ↑Keap1/Nrf2 signaling pathways | [111] |
| Flammulina velutipes | Freeze dried ethanolic extracts of fruiting bodies | ||||
| Pleurotus citrinopileatus | Isolated Polysaccharide–peptide I (PSI),Polysaccharide–peptide II (PSII) | In vitro batch culture: fecal samples from healthy human volunteers on YCFA medium | PSI: ↓Escherichia-Schigella PSII: ↑Bifidobacterium ↑Lactococcus ↑Lactobacillus ↑Desulfovibrionaceae ↑Lachnospiraceae ↑Odoribacter ↑Coriobacteriaceae ↑Blautia ↓Escherichia-Schigella | Administration of PSI resulted in higher production of SCFAs | [188] |
| Pleurotus eryngii | Water soluble polysaccharide fraction | In vivo: C57BL/6J mice HFD | ↑Anaerostipes ↑Clostridium ↑Lactococcus ↓Roseburia ↓Lactobacillus | [189] | |
| Powdered fruiting bodies | In vivo: CD–1 mice DSS-induced colitis | ↑Odoribacteraceae ↑Adlercreutzia ↑Akkermanisa ↑Lactobacillus ↑Anaerostipes ↑Allobaculum ↓Actinobacteria ↓Mollicutes ↓Desulfovibrionace ↓Enterococcaceae ↓Turicibacter ↓Dorea ↓Bacteroides ↓Prevotella | ↑Production of SFCAs in dose-depended manner ↓Local—colon inflammation
| [190] | |
| Pleurotus ostreatus | Powdered fruiting bodies | In vivo: C57BL/6J mice HFD-induced obesity | ↑Oscillospira ↑Lactobacillus group ↑Bifidobacterium ↑Anaerostipes ↑Anaerovorax ↑Anaerofustis ↑Ruminococcus ↑Coprococcus ↓Bacteroides ↓Roseburia ↓Acinetobacter ↓Agrobacterium ↓Microbacterium ↓Novosphingobium ↓Streptococcus ↓Prevotella ↓Sphingomonas ↓Macrococcus ↓Lactococcus Compared to HFD only | [191] | |
| Powdered fruiting bodies | In vivo: piglets (crossbred: Duroc × Large white × Landrace) | ↑Prevotella ↑Anaerovibrio ↑Veillonellaceae | ↑Production of SFCAs in group with supplementation of P. ostreatus, | [192] | |
| Pleurotus sajor-caju | Powdered mycelium | In vivo: Zucker rats | ↑Faecalibaculum ↑Bifidobacterium ↑Roseburia ↑Blautia ↓Escherichia-Shigella | ↑Production of SCFAs in the colon | [193] |
| Agaricus bisporus | Roasted fruiting bodies | Open label crossover study in healthy adults (n = 32) | ↑Bacteroides ↑Parabacteroides ↑Coprococcus ↑Sutterella ↑Anaerostipes Comparing to diet with meat | [194] | |
| Agaricus bisporus | Powdered fruiting bodies | In vivo: C57BL/J mice | Stable Firmicutes/Bacteroidetes ratio ↑Verrucomicrobia ↑Akkermansiaceae ↑Tannerellaceae ↓Prevotellaceae | ↓Inflammation related genes levels such as IL–6, Nox–2 (Portobello only), Hmox–1 | [195] |
| Agaricus bisporus brunnescens | |||||
| Armillaria mellea | A. mellea fermentation liquor | In vivo: Sprague–Dawley rats Insomnia model by the p-chlorophenylalanine (PCPA) induction | ↑Lachnospiraceae NK4A136 group ↑Lachnospiraceae ↑the [Eubacterium] xylanophilum group ↑Ruminococcus ↑Candidatus ↑Saccharimonas ↑the [Eubacterium] coprostanoligenes group ↓Lactobacillus ↓Maribaculum ↓Prevotellaceae UCG-001 | Administration of AFL resulted in ↑5-HT1A and 5-HT2A proteins in hippocampus comparing to model group ↓Peripherial inflammation reduced after AFL administration in dose-dependent manner
| [196] |
| Lentinula edodes | Isolated polysaccharide—lentinan | In vivo: C57BL/6J mice, HFD | ↑Actinobacteria ↑Firmicutes ↑Bifidobacterium ↓Proteobacteria ↓Epsilonbacteraeota | Addition of lentinan to HD diet resulted in improved intestinal barrier
| [197] |
| Isolated β–glucan | In vivo: C57BL/6J mice HFD | ↑Clostridiales ↑Lachnospiraceae ↑Ruminococcaceae | Addition of lentinan to HFD resulted in improved intestinal barrier
| [107] | |
| Polysaccharides | In vivo: C57BL/6J male mice DSS-induced colitis | In DDS-induced group: ↑Bacteroides ↑Helicobacter ↑Parasutterella ↓Firmicutes Addition of FMP to diet reversed negative gut microbiota alteration | Improved intestinal barrier after mushroom addition to diet:
| [198] | |
| Hericium erinaceus | Powderized H. erinaceus fruting bodies | In vivo: 11-years old dogs | ↑Bacteroidetes ↑Bacteroidales ↓Firmicutes ↓Streptococcus ↓Tyzzerella ↓Campylobacteraceae ↓Campylocaber | [199] | |
| Powdered fruiting bodies(as dietary supplement) | A pilot study with 13 healthy adults | ↑Roseburia faecis ↑Faecalibacterium prausnitzii ↑Eubacterium rectale ↑Fusicatenibacter saccharivorans ↑Kineothrix alysoides ↑Gemmiger formicilis ↑Dorea longicatena ↓Streptococcus thermophilus ↓Roseburia intestinalis ↓Bacteroides caccae ↓Anaerostipes hadrus | Short supplementation (7 days) resulted in increased alpha and beta diversity—it may produce short term effect retention. | [200] | |
| Dictyophora indusiata | Different fractions of polysaccharides: soluble (DIPX) and insoluble in water (DIPY) | In vivo: Kunming mice | In DIPY group: ↓Lachnospiraceae ↑Lactobacillus Gut microbiota composition in DIPX group did not differ to the control group | ↑Production of SCFAs in DIPY group Production of SCFAs in DIPX groups did not differ comparing to the control group | [201] |
| Isolated polysaccharide | In vivo: BALB/c mice HFD | Addition of polysaccharide resulted in reversion of HFD-induced gut alteration: ↓Firmicutes/Bacteroidetes ratio | Improved intestinal barrier after polysaccharide supplementation to diet: ↑claudin-1, ↑occludin, and ↑zonula occludens (ZO-1) ↓Serum LPS → ↓peripheral inflammation | [202] | |
| Helvella leucopus | Isolated polysaccharide (p-HLP) | In vivo: C57BL/6 DSS-induced colitis | ↑Bacteroidaceae ↑Prevotellaceae ↑Akkermansiaceae | ↓Local inflammation
| [203] |
| Tremella fuciformis | Isolated polysaccharides | In vivo: C57BL/6 DSS-induced colitis | ↑Lactobacillaceae ↑Lactobacillus ↑Marinifilaceae ↓Helicobacter ↓Ruminococcaceae | ↓ Local inflammation
| [204] |
| Grifola frondosa | Isolated polysaccharides | In vivo: Wistar rats HFD | ↑Helicobacter ↑Intestinimonas ↑Barnesiella ↑Parasutterella ↑Ruminococcus ↑Flavonifracter ↓Clostridium-XVIII ↓Turicibacter | [205] | |
| Isolated heteropolysaccharide (GFP-N) | In vivo: ICR mice T2DM induced | ↑Porphyromonas gingivalis ↑Akkermansia muciniphila ↑Lactobacillus acidophilus ↑Tannerella forsythia ↑Bacteroides acidifaciens ↑Roseburia intestinalis | Improved oral glucose test, alleviated insulin resistance, decreased the fast blood glucose level | [206] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazur, J.; Hnatyk, K.; Kała, K.; Sułkowska-Ziaja, K.; Muszyńska, B. Discovering the Potential Mechanisms of Medicinal Mushrooms Antidepressant Activity: A Review. Antioxidants 2023, 12, 623. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030623
Lazur J, Hnatyk K, Kała K, Sułkowska-Ziaja K, Muszyńska B. Discovering the Potential Mechanisms of Medicinal Mushrooms Antidepressant Activity: A Review. Antioxidants. 2023; 12(3):623. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030623
Chicago/Turabian StyleLazur, Jan, Kamil Hnatyk, Katarzyna Kała, Katarzyna Sułkowska-Ziaja, and Bożena Muszyńska. 2023. "Discovering the Potential Mechanisms of Medicinal Mushrooms Antidepressant Activity: A Review" Antioxidants 12, no. 3: 623. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030623