
Camphene as a Protective Agent in Myocardial Ischemia/Reperfusion Injury

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Administration of Camphene and Experimental Procedures for I/R in Isolated Hearts
2.3. Infarct Size (IS) Determination
2.4. LDH Determination
2.5. Determination of Protein Carbonyls
2.6. Glutathione Determination
2.7. Measurement of Citrate Synthase (CS) Activity
2.8. Determination of Antioxidant Enzymes Activity
2.8.1. Catalase (CAT) Activity Assay
2.8.2. Superoxide Dismutase (Mn-SOD) Activity Assay
2.8.3. Glutathione Peroxidase (GPx) Activity Assay
2.8.4. Glutathione Reductase (GR) Activity Assay
2.9. RNA Preparation and Quantitative Real-Time PCR (qPCR)
2.10. Lipid Peroxidation
2.11. Statistical Analysis
3. Results
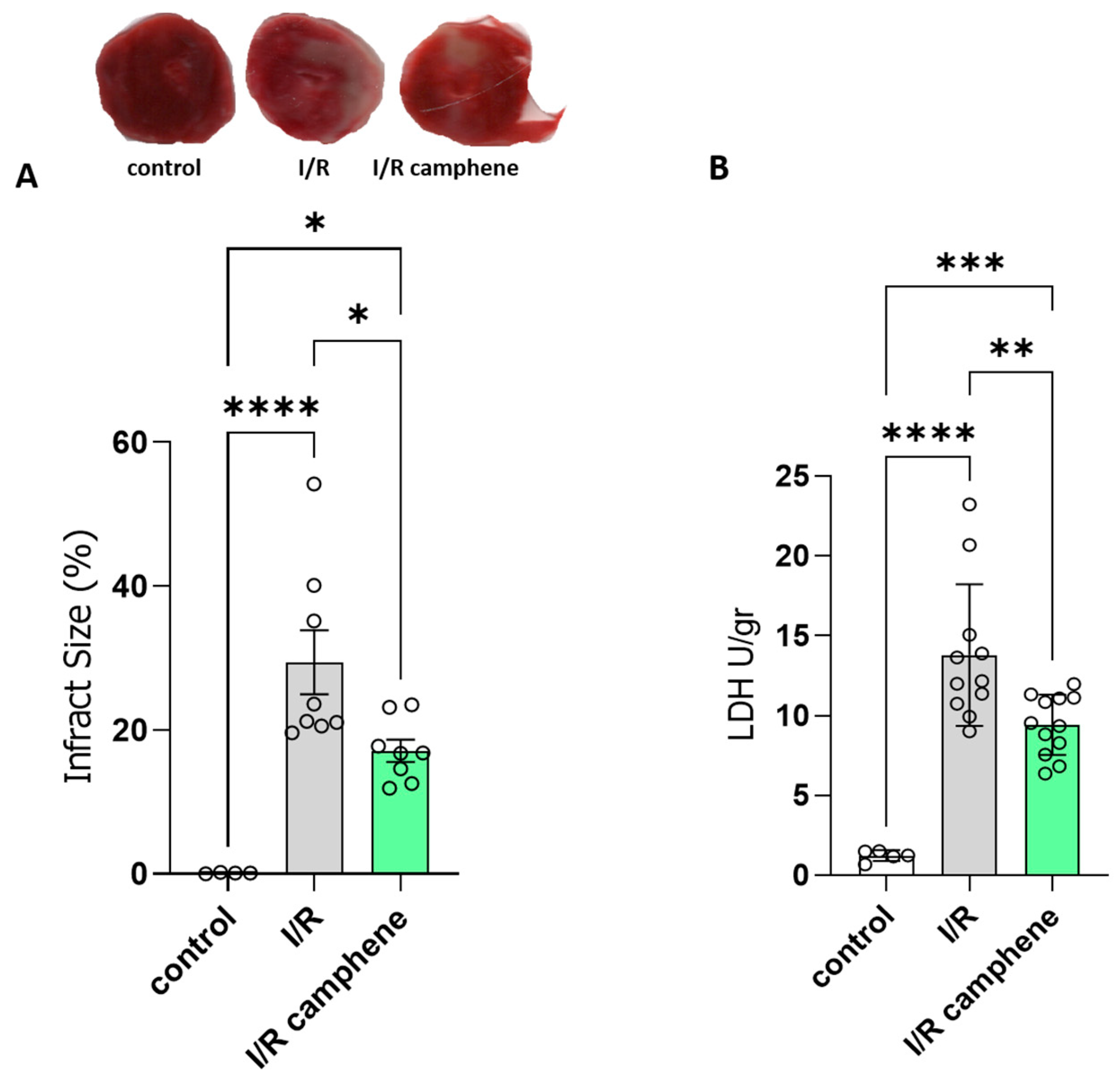
3.1. Camphene Pretreatment Reduces Infarct Size and Ameliorates Myocardial I/R Injury
3.2. Camphene Attenuates I/R-Induced Oxidative Stress and Increases Mitochondrial Content
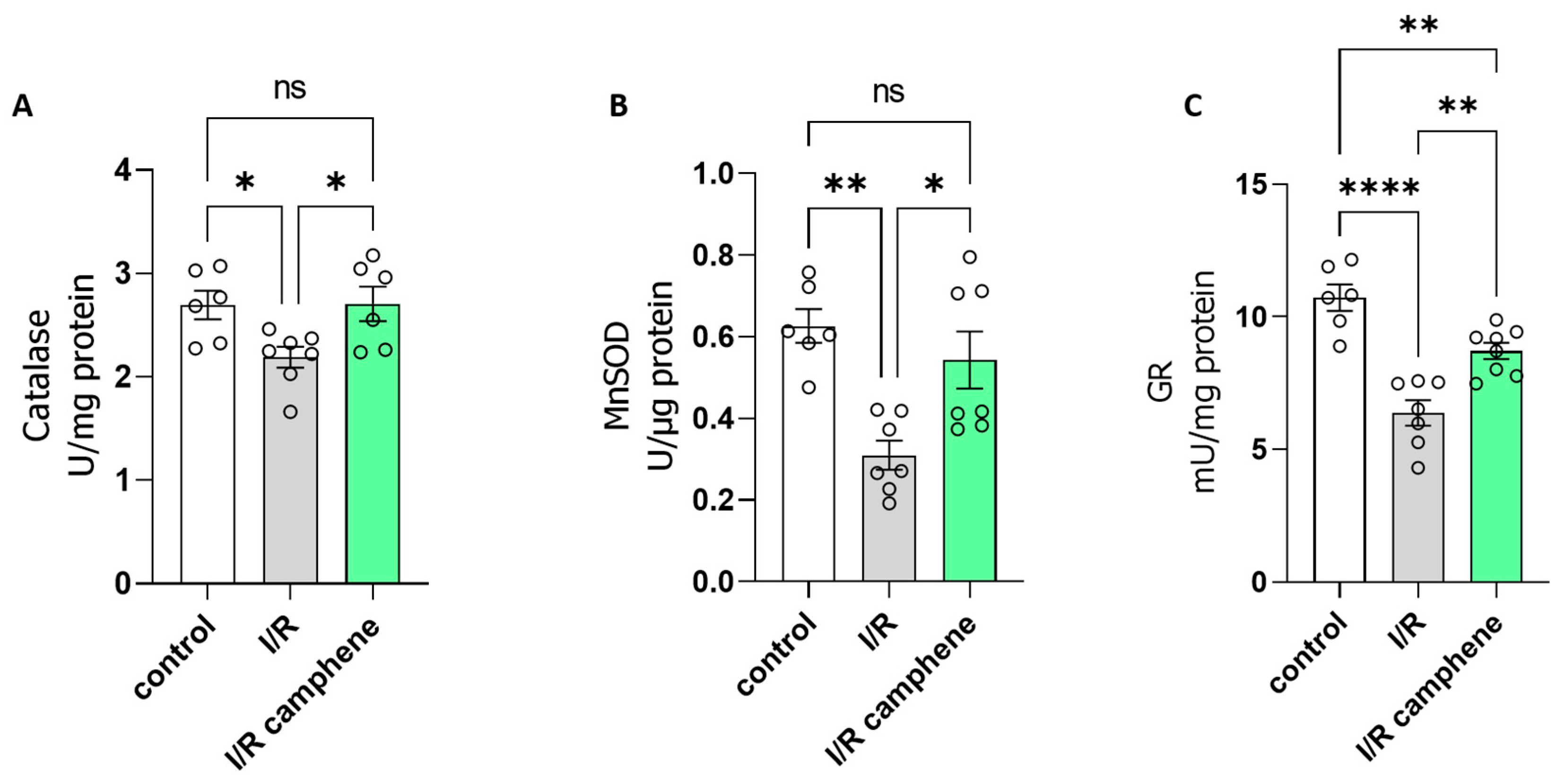
3.3. Camphene Pretreatment Activates Antioxidant Mechanisms in the Heart
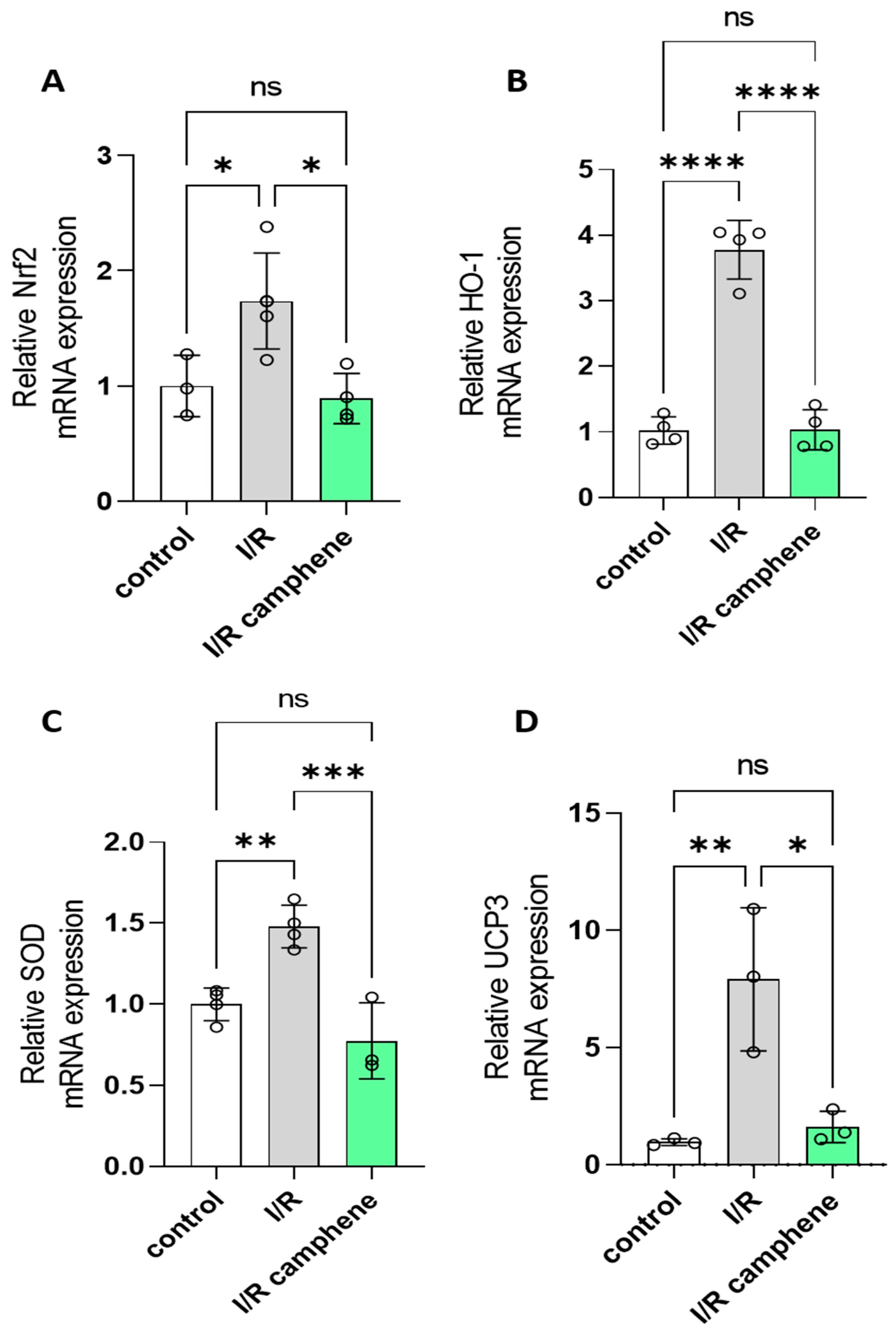
3.4. Camphene Pretreatment Decreases Nrf2, HO-1, SOD, and UCP3 mRNA
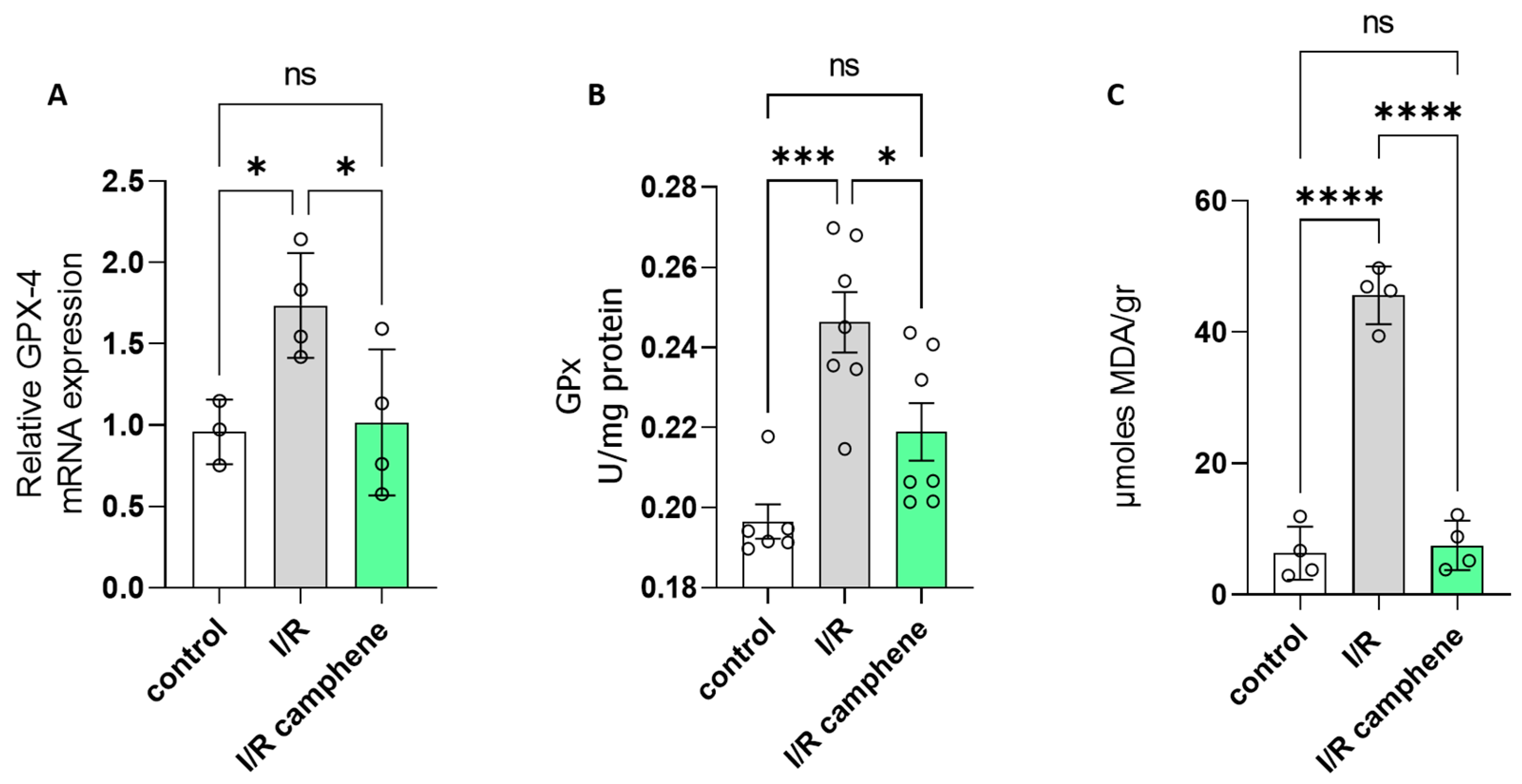
3.5. Camphene Inhibits Ferroptosis Induced by I/R
4. Discussion
5. Conclusions
Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Andreadou, I.; Daiber, A.; Baxter, G.F.; Brizzi, M.F.; Di Lisa, F.; Kaludercic, N.; Lazou, A.; Varga, Z.V.; Zuurbier, C.J.; Schulz, R.; et al. Influence of cardiometabolic comorbidities on myocardial function, infarction, and cardioprotection: Role of cardiac redox signaling. Free Radic. Biol. Med. 2021, 166, 33–52. [Google Scholar] [CrossRef] [PubMed]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS). Redox Biol. 2017, 13, 94–162, Erratum in Redox Biol. 2018, 14, 694–696. [Google Scholar] [CrossRef]
- Davidson, S.M.; Adameová, A.; Barile, L.; Cabrera-Fuentes, H.A.; Lazou, A.; Pagliaro, P.; Stensløkken, K.O.; Garcia-Dorado, D. EU-CARDIOPROTECTION COST Action (CA16225). Mitochondrial and mitochondrial-independent pathways of myocardial cell death during ischaemia and reperfusion injury. J. Cell Mol. Med. 2020, 24, 3795–3806. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fan, H.; Wang, S.; Tang, G.; Zhai, C.; Shen, L. Ferroptosis: A Novel Therapeutic Target for Ischemia-Reperfusion Injury. Front. Cell Dev. Biol. 2021, 10, 688605. [Google Scholar] [CrossRef] [PubMed]
- Pizzorno, J. Glutathione! Integr. Med. 2014, 13, 8–12. [Google Scholar]
- Barteková, M.; Adameová, A.; Görbe, A.; Ferenczyová, K.; Pecháňová, O.; Lazou, A.; Dhalla, N.S.; Ferdinandy, P.; Giricz, Z. Natural and synthetic antioxidants targeting cardiac oxidative stress and redox signaling in cardiometabolic diseases. Free Radic. Biol. Med. 2021, 169, 446–477. [Google Scholar] [CrossRef] [PubMed]
- Matuz-Mares, D.; Riveros-Rosas, H.; Vilchis-Landeros, M.M.; Vázquez-Meza, H. Glutathione Participation in the Prevention of Cardiovascular Diseases. Antioxidants 2021, 10, 1220. [Google Scholar] [CrossRef]
- Mata, A.; Cadenas, S. The Antioxidant Transcription Factor Nrf2 in Cardiac Ischemia–Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 11939. [Google Scholar] [CrossRef]
- López-Bernardo, E.; Anedda, A.; Sánchez-Pérez, P.; Acosta-Iborra, B.; Cadenas, S. 4-Hydroxynonenal induces Nrf2-mediated UCP3 upregulation in mouse cardiomyocytes. Free Radic. Biol. Med. 2015, 88 Pt B, 427–438. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Galatou, E.; Panagiotidis, G.-D.; Ravingerová, T.; Lazou, A. Cardioprotective Effects of PPARβ/δ Activation against Ischemia/Reperfusion Injury in Rat Heart Are Associated with ALDH2 Upregulation, Amelioration of Oxidative Stress and Preservation of Mitochondrial Energy Production. Int. J. Mol. Sci. 2021, 22, 6399. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, I.; Makrecka-Kuka, M.; Kuka, J.; Liepinsh, E.; Dambrova, M.; Lazou, A. Pharmacological activation of PPARβ/δ preserves mitochondrial respiratory function in ischemia/reperfusion via stimulation of fatty acid oxidation-linked respiration and PGC-1α/NRF-1 signaling. Front. Endocrinol. 2022, 15, 941822. [Google Scholar] [CrossRef]
- Barlaka, E.; Galatou, E.; Mellidis, K.; Ravingerova, T.; Lazou, A. Role of Pleiotropic Properties of Peroxisome Proliferator-Activated Receptors in the Heart: Focus on the Nonmetabolic Effects in Cardiac Protection. Cardiovasc. Ther. 2016, 34, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Tsoumani, M.; Georgoulis, A.; Nikolaou, P.E.; Kostopoulos, I.V.; Dermintzoglou, T.; Papatheodorou, I.; Zoga, A.; Efentakis, P.; Konstantinou, M.; Gikas, E.; et al. Acute administration of the olive constituent, oleuropein, combined with ischemic postconditioning increases myocardial protection by modulating oxidative defense. Free Radic. Biol. Med. 2021, 166, 18–32. [Google Scholar] [CrossRef]
- Sharmeen, J.B.; Mahomoodally, F.M.; Zengin, G.; Maggi, F. Essential Oils as Natural Sources of Fragrance Compounds for Cosmetics and Cosmeceuticals. Molecules 2021, 26, 666. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, H.C.; Ieque, A.L.; Valverde, T.L.; Baldin, V.P.; Meneguello, J.E.; Campanerut-Sá, P.A.Z.; Vandresen, F.; Ghi-raldi Lopes, L.D.; Passos Souza, M.R.; Santos, N.C.S.; et al. Activity of (-)-Camphene Derivatives Against Mycobacterium tuberculosis in Acidic pH. Med. Chem. 2021, 17, 485–492. [Google Scholar] [CrossRef]
- de Freitas, B.C.; Queiroz, P.A.; Baldin, V.P.; do Amaral, P.H.; Rodrigues, L.L.; Vandresen, F.; RCaleffi-Ferracioli, K.; de LScodro, R.B.; Cardoso, R.F.; Siqueira, V.L. (-)-Camphene-based derivatives as potential antibacterial agents against Staphylococcus aureus and Enterococcus spp. Future Microbiol. 2020, 15, 1527–1534. [Google Scholar] [CrossRef]
- Yamaguchi, M.U.; Barbosa da Silva, A.P.; Ueda-Nakamura, T.; Dias Filho, B.P.; Conceição da Silva, C.; Naka-mura, C.V. Effects of a Thiosemicarbazide Camphene Derivative on Trichophyton mentagrophytes. Molecules 2009, 14, 1796–1807. [Google Scholar] [CrossRef]
- Gadotti, V.M.; Huang, S.; Zamponi, G.W. The terpenes camphene and alpha-bisabolol inhibit inflammatory and neuropathic pain via Cav3.2 T-type calcium channels. Mol. Brain 2021, 14, 166. [Google Scholar] [CrossRef]
- Lin, C.T.; Chen, C.J.; Lin, T.Y.; Tung, J.C.; Wang, S.Y. Anti-inflammation activity of fruit essential oil from Cinnamomum inslarimontanum Hayata. Bioresour. Technol. 2008, 99, 8783–8787. [Google Scholar] [CrossRef]
- Boyd, E.M.; Sheppard, P. Nutmeg Oil and Camphene as Inhaled Expectorants. Arch. Otolaryngol. 1970, 92, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Quintans-Júnior, L.; Moreira, J.C.F.; Pasquali MA, B.; Rabie SM, S.; Pires, A.S.; Schröder, R.; Rabelo, T.K.; Santos, J.P.A.; Lima PS, S.; Cavalcanti, S.C.H.; et al. Antinociceptive Activity and Redox Profile of the Monoterpenes (+)-Camphene, p-Cymene, and Geranyl Acetate in Experimental Models. Int. Sch. Res. Not. 2013, 2013, 459530. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, I.; Hadzopoulou-Cladaras, M. Camphene, a plant-derieved monoterpene, exerts its hypolipidemic action by affecting SREBP-1 and MTP expression. PLoS ONE 2016, 11, e0147117. [Google Scholar] [CrossRef] [PubMed]
- Chroni, A.; Liu, T.; Gorshkova, I.; Kan, H.Y.; Uehara, Y.; Von Eckardstein, A.; Zannis, V.I. The central helices of ApoA-I can promote ATP-binding cassette transporter A1 (ABCA1)-mediated lipid efflux. Amino acid residues 220–231 of the wild-type ApoAI are required for lipid efflux in vitro and high density lipoprotein formation in vivo. J. Biol. Chem. 2003, 278, 6719–6730. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, H.; Xia, D.; Wang, S. Antioxidant Properties of Camphene-Based Thiosemicarbazones: Experimental and Theoretical Evaluation. Molecules 2020, 25, 1192. [Google Scholar] [CrossRef] [PubMed]
- Suji, B.; Jisu, K.; Byung, M. Camphene Attenuates Skeletal Muscle Atrophy by Regulating Oxidative Stress and Lipid Metabolism in Rats. Nutrients 2020, 12, 3731. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, I.; Peroulis, N.; Pantazis, P.; Hadzopoulou-Cladaras, M. Camphene, a plant-derived monoterpene, reduces plasma cholesterol and triglycerides in hyperlipidemic rats independently of HMG-CoA reductase activity. PLoS ONE 2011, 6, e20516. [Google Scholar] [CrossRef] [PubMed]
- Ravingerová, T.; Matejíková, J.; Neckár, J.; Andelová, E.; Kolár, F. Differential role of PI3K/Akt pathway in the infarct size limitation and antiarrhythmic protection in the rat heart. Mol. Cell Biochem. 2007, 297, 111–120. [Google Scholar] [CrossRef]
- Burd, J.F.; Usategui-Gomez, M. Immunochemical studies on lactate dehydrogenase. Biochim. Biophys. Acta 1973, 310, 238–247. [Google Scholar] [CrossRef]
- Colombo, G.; Clerici, M.; Garavaglia, M.E.; Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. A step-by-step protocol for assaying protein carbonylation in biological samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1019, 178–190. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- Eigentler, A. Laboratory protocol: Citrate synthase, Mitochondrial marker enzyme. Mitochondr. Physiol. Netw. 2012, 17, 1–12. [Google Scholar]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase activity in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.T. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Mancilla, R.; Pava-Mejia, D.; van Polanen, N.; de Wit, V.; Bergman, M.; Grevendonk, L.; Jorgensen, J.; Kornips, E.; Hoeks, J.; Hesselink, M.K.C.; et al. Invasive and noninvasive markers of human skeletal muscle mitochondrial function. Physiol. Rep. 2023, 11, e15734. [Google Scholar] [CrossRef]
- Chen, Z.; Yan, Y.; Qi, C.; Liu, J.; Li, L.; Wang, J. Ferroptosis in cardiovascular diseases: Role and mechanism. Cell Biosci. 2023, 13, 226. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [CrossRef]
- Dhalla, N.S.; Elmoselhi, A.B.; Hata, T.; Makino, N. Status of myocardial antioxidants in ischemia–reperfusion injury. Cardiovasc. Res. 2000, 47, 446–456. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Yellon, D.M. Myocardial ischemia-reperfusion injury: A neglected therapeutic target. J. Clin. Investig. 2013, 123, 92–100. [Google Scholar] [CrossRef]
- Davidson, S.M.; Ferdinandy, P.; Andreadou, I.; Bøtker, H.E.; Heusch, G.; Ibáñez, B.; Ovize, M.; Schulz, R.; Yellon, D.M.; Hausenloy, D.J.; et al. Multitarget Strategies to Reduce Myocardial Ischemia/Reperfusion Injury: JACC Review Topic of the Week. J. Am. Coll. Cardiol. 2019, 73, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Lu, Y.; Xin, L.; Gao, J.; Shang, C.; Jiang, Z.; Lin, H.; Fang, X.; Qu, Y.; Wang, Y.; et al. Role of Oxidative Stress in Reperfusion following Myocardial Ischemia and Its Treatments. Oxid. Med. Cell. Longev. 2021, 2021, 6614009. [Google Scholar] [CrossRef]
- Bradic, J.; Zivkovic, V.; Srejovic, I.; Jakovljevic, V.; Petkovic, A.; Turnic, T.N.; Jeremic, J.; Jeremic, N.; Mitrovic, S.; Sobot, T.; et al. Protective Effects of Galium verum L. Extract against Cardiac Ischemia/Reperfusion Injury in Spontaneously Hypertensive Rats. Oxid. Med. Cell. Longev 2019, 2019, 4235405. [Google Scholar] [CrossRef] [PubMed]
- Draginic, N.; Milosavljevic, I.; Andjic, M.; Jeremic, J.; Nikolic, M.; Sretenovic, J.; Kocovic, A.; Srejovic, I.; Zivkovic, V.; Bolevich, S.; et al. Short-Term Administration of Lemon Balm Extract Ameliorates Myocardial Ischemia/Reperfusion Injury: Focus on Oxidative Stress. Pharmaceuticals 2022, 15, 840. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Zhang, T.; Zhang, W.; Zhao, Z.; Sun, J. Natural Drugs as a Treatment Strategy for Cardiovascular Disease through the Regulation of Oxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020, 5430407. [Google Scholar] [CrossRef]
- Tan, M.; Yin, Y.; Ma, X.; Zhang, J.; Pan, W.; Tan, M.; Zhao, Y.; Yang, T.; Jiang, T.; Li, H. Glutathione system enhancement for cardiac protection: Pharmacological options against oxidative stress and ferroptosis. Cell Death Dis. 2023, 14, 131. [Google Scholar] [CrossRef]
- Daiber, A.; Andreadou, I.; Oelze, M.; Davidson, S.M.; Hausenloy, D.J. Discovery of new therapeutic redox tar-gets for cardioprotection against ischemia/reperfusion injury and heart failure. Free. Radic. Biol. Med. 2021, 163, 325–343. [Google Scholar] [CrossRef]
- Bertero, E.; Maack, C. Ins and Outs of Glutathione in Cardiac Ischemia/Reperfusion Injury. J. Circ. Res. 2023, 133, 877–879. [Google Scholar] [CrossRef]
- Fedorova, M.; Griesser, E.; Vemula, V.; Weber, D.; Ni, Z.; Hoffmann, R. Protein and lipid carbonylation in cellular model of nitrosative stress: Mass spectrometry, biochemistry and microscopy study. Free Radic. Biol. Med. 2014, 75 (Suppl. S1), S15. [Google Scholar] [CrossRef]
- Haramaki, N.; Stewart, D.B.; Aggarwal, S.; Ikeda, H.; Reznick, A.Z.; Packer, L. Networking antioxidants in the isolated rat heart are selectively depleted by ischemia-reperfusion. Free Radic. Biol. Med. 1998, 25, 329–339. [Google Scholar] [CrossRef]
- Leichtweis, S.; Ji, L.L. Glutathione deficiency intensifies ischaemia-reperfusion induced cardiac dysfunction and oxidative stress. Acta Physiol. Scand. 2001, 172, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marczin, N.; El-Habashi, N.; Hoare, G.S.; Bundy, R.E.; Yacoub, M. Antioxidants in myocardial ischemia–reperfusion injury: Therapeutic potential and basic mechanisms. Arch. Biochem. Biophys. 2003, 420, 2222–2236. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Bohn, T. Exogenous antioxidants--Double-edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Aceto, A.; Mezzetti, A.; Di Ilio, C.; Calafiore, A.M.; De Cesare, D.; Bosco, G.; Acciai, N.; Cappelletti, L.; Federici, G.; Cuccurullo, F. Effect of Ischaemia-Reperfusion on Glutathione Peroxidase, Glutathione Reductase and Glutathione Transferase Activities in Human Heart Protected by Hypothermic Cardioplegia, Free Radical. Res. Commun. 1990, 8, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Ravingerová, T.; Kindernay, L.; Barteková, M.; Ferko, M.; Adameová, A.; Zohdi, V.; Bernátová, I.; Ferenczyová, K.; Lazou, A. The Molecular Mechanisms of Iron Metabolism and Its Role in Cardiac Dysfunction and Cardioprotection. Int. J. Mol. Sci. 2020, 21, 7889. [Google Scholar] [CrossRef]
- Bøtker, H.E.; Hausenloy, D.; Andreadou, I.; Antonucci, S.; Boengler, K.; Davidson, S.M.; Deshwal, S.; Devaux, Y.; Di Lisa, F.; Di Sante, M.; et al. Practical guidelines for rigor and reproducibility in preclinical and clinical studies on cardioprotection. Basic. Res. Cardiol. 2018, 113, 39. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stamatiou, R.; Anagnostopoulou, M.; Ioannidou-Kabouri, K.; Rapti, C.; Lazou, A. Camphene as a Protective Agent in Myocardial Ischemia/Reperfusion Injury. Antioxidants 2024, 13, 405. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040405
Stamatiou R, Anagnostopoulou M, Ioannidou-Kabouri K, Rapti C, Lazou A. Camphene as a Protective Agent in Myocardial Ischemia/Reperfusion Injury. Antioxidants. 2024; 13(4):405. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040405
Chicago/Turabian StyleStamatiou, Rodopi, Maria Anagnostopoulou, Konstantina Ioannidou-Kabouri, Chrysa Rapti, and Antigone Lazou. 2024. "Camphene as a Protective Agent in Myocardial Ischemia/Reperfusion Injury" Antioxidants 13, no. 4: 405. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040405