1. Introduction
Grape pomace (GP) predominantly comprises grape seeds, skins, and stalks, containing high levels of carbohydrates and proteins [
1]. Specifically, grape seeds are abundant in polyunsaturated fatty acids, contributing to animal health when consumed in moderation. Additionally, GP is a rich source of polyphenols, known for their potent antioxidant properties [
2,
3]. With the increasing demand for wine and concerns over the environmental impact of the traditional treatments, there is a pressing need to explore efficient resource utilization strategies for GP.
Grape pomace is rich in diverse polyphenolic compounds, including tannins, proanthocyanidins, anthocyanidins, flavonoids, and phenolic acids, attributable to the extraction conditions, efficiency, and chemical reactions inherent in the wine-making process [
1,
4,
5]. Voicu et al. [
6] showed that the growth performance and digestive metabolism of beef cattle are not adversely affected by many of these polyphenolic compounds. Notably, condensed tannins (CT) stand out as the foremost bioactive constituents, constituting 58.7% of the total phenolic compounds in GP [
7]. Condensed tannins have more hydroxyl groups and a higher polymerization of polyphenols, making them stronger antioxidants than simple phenolics [
8]. Increasing evidence suggests that polyphenolic compounds can be utilized as dietary additives in adult ruminant diets [
9,
10]. Furthermore, the structure–activity relationships and chemical environment of the polyphenol compounds in GP affect the antioxidant efficiency and balance [
11]. Studies have shown that feeding 25 kg lambs with 10% GP and feed increased the total antioxidant capacity (T-AOC) and Glutathione Peroxidase 4 (GPX4) and superoxide dismutase (SOD) activity in the lambs [
12].
Condensed tannins have the ability to bind and precipitate proteins. Due to their complex structure and large relative molecular mass, hydrolysis of CT can occur only under strong oxidation and acidic conditions. Therefore, CT have a strong antibacterial ability, which can destroy microbial membranes by inhibiting the substrate of enzymes and inactivating membrane-binding proteins [
13]. In a previous experiment [
14], it was found that GP can improve the anti-inflammatory effect of Caco-2 cells. In addition, the addition of GP can inhibit the growth of pathogenic bacteria such as
Enterobacteriaceae and
E. coli. in feces [
15].
In the late fattening period of beef cattle, long-term feeding with a high-concentrate diet can stimulate the release of proinflammatory cytokines, which leads to oxidative stress in the body [
16]. We can use feed rich in polyphenols, and GP is one of the best choices; however, dietary polyphenols can have toxic effects on animals’ bodies when fed in large doses. The evidence indicates that dietary polyphenols can have adverse effects on animals’ bodies through their antioxidant action [
17]. A high dose of GP may over-protect proteins and hinder the digestion and absorption of nutrients by reducing the activity of rumen microorganisms [
8]. Therefore, this evidence suggests that although wine grains can be used as an antioxidant diet for ruminants, we need to explore the appropriate replacement ratio to prevent a high concentration reducing the stability of ruminal and rectal microbial communities, which is detrimental to the health of beef cattle.
At present, the animal studies on GP mainly focus on growth performance [
6,
7] and nitrogen utilization efficiency (NUE) [
18], and the effects of GP on the antioxidant activity and health of beef cattle are still unclear. Therefore, the purpose of this study was to investigate the effects of different proportions of GP on the growth performance, antioxidant activity, and rumen and rectal microbiota of beef cattle through feeding experiments. The purpose is to understand the use of GP and provide a theoretical basis for its application in diets.
4. Discussion
The condensed tannins and lignins in GP emerge as pivotal factors influencing digestive processes [
27,
28]. However, recent studies have demonstrated that CT from various sources and with distinct chemical structures exhibit clear dose effects but above a certain threshold can cause negative effects [
29]. Total tannins can be beneficial bioactive compounds in ruminant diets when included at levels below 3% of dietary dry matter [
30]. In this study, the total tannin content within the 20% dried grape pomace diet did not surpass 2 g/kg. Despite this, the observed ADG and apparent digestibility of the beef cattle were lower compared to those in the control group. These findings suggest that the limitations observed in the diet were attributed to more than just the tannin content in the GP. Lignin acts as a physical barrier against rumen microorganisms, inhibiting the degradation of cell walls. This hindrance in the digestive processes results in a decrease in the apparent digestibility of nutrients [
31,
32,
33]. Abarghuei et al. [
32] observed that even after eliminating the influence of CT, the apparent digestibility of nutrients remained significantly reduced. This finding underscores that lignin independently exerts a detrimental impact on the growth performance of beef cattle. In this study, the lignin content (ADL = 7.88% DM) observed in the G20 group may have negatively influenced the growth performance of the beef cattle. Consequently, it is imperative to consider both the tannin and lignin content when incorporating GP into the diet.
Upon entering the rumen, dietary lipids transform processes such as lipolysis and fatty acid biohydrogenation (BH). A portion is hydrolyzed by microbial lipases, yielding nutrients such as glycerol for bacterial utilization [
34]. The remaining lipids proceed through the rumen and are digested and absorbed in the small intestine. The microbial results of this experiment showed that
Butyrivibrio (
Figure 3) was the dominant genus in group G0, which was shown to be involved in further steps of BH after lipolysis [
10]. After the addition of GP, the dominant genera changed to
unclassified-Christensenellaceae and
Pantoea in the rumen. Additionally, research indicates that tannins can diminish the release of free fatty acids by inhibiting lipase activity, and they can impede lipid digestion by binding to bile salts [
35]. In this experiment, the EED of the 20% group exhibited a decrease, aligning with the findings observed by Molosse [
36]. This decline may be attributed to the inhibitory effects of the tannins in GP on lipid absorption.
Our observations revealed a linear decrease in urinary nitrogen with an increasing addition of GP, a phenomenon likely attributed to the concurrent rise in tannin content. This correlation suggests the direct impact of the tannin content in GP on ruminal protein metabolism. Relevant studies indicate that supplementing feed with the appropriate tannins can alter the nitrogen pathway, facilitating nitrogen transfer from the urine to the feces [
37,
38]. This, in turn, contributes to a reduction in the loss of highly volatile forms of nitrogen. On the one hand, tannins bind with a portion of the protein in the rumen, leading to decreased utilization of dietary protein within the rumen. On the other hand, tannins interact with proteins, causing a shift in the location of nitrogen metabolism and digestion. Additionally, this diminishes microbial enzyme activity, thereby slowing down the growth rate of hydrolytic bacteria and reducing the production of ruminal NH
3-N. Rumen microorganisms absorb excess nitrogen into NH
3-N and convert it into urea and then excrete it through the urine or circulate it in the intestine. Consequently, the interaction between tannins and proteins in the rumen can mitigate the conversion of nitrogen in the diet, reducing the urinary nitrogen content [
39]. Therefore, the utilization of GP holds significant potential for promoting the green and low-carbon development of beef cattle. Previous studies have established a positive correlation between fecal nitrogen excretion and urinary nitrogen and nitrogen intake [
40]. The nitrogen in feces predominantly exists as organic nitrogen, which exhibits lower volatility. Conversely, the nitrogen in urine primarily exists in the form of urea, which undergoes rapid oxidation into NO
3− and nitrite (N
2O
−) through soil nitrification [
41]. This process generates N
2O, an intermediate product recognized as a potent greenhouse gas. In contrast, the addition of GP allows for a reduction in the degradation and deamination rate of dietary protein in the rumen [
37,
42,
43]. This alteration in the nitrogen pathways helps mitigate N
2O emissions, thereby contributing to the goal of reducing greenhouse gas emissions during both production and manure treatment.
During times of oxidative stress, cells generate a significant amount of reactive oxygen species (ROS). This increase in ROS levels can lead to inflammation, prompting the body to rely on antioxidant enzymes and antioxidants for the purpose of counteracting the sudden rise in ROS. Examples include SOD and CAT, which act to safeguard cells against the damage caused by ROS [
44,
45]. High-concentrate diets promote oxidative stress in beef cattle, so we can promote increases in endogenous antioxidants (SOD and CAT) by supplementing exogenous antioxidant molecules to offset the ROS of beef cattle. SOD is the first line of defense, catalyzing superoxides to produce oxygen and hydrogen peroxide, while CAT neutralizes the decomposition of hydrogen peroxide into oxygen and water [
46]. Veras [
47] showed that tannic acid in feed promoted an increased CAT concentration in mice. Feed including 10% GP was found to increase the concentration of CAT, similar to the results of previous studies. However, a high proportion of GP in the diet did not improve the antioxidant levels in beef cattle. This could be due to the pro-oxidation effect of a high proportion of polyphenol compounds in beef cattle. Studies have shown that the polyphenols in bilberry extract lead to apoptosis in cancer cells by increasing intracellular ROS levels [
48]. Combined with heatmap analysis, although the SOD concentration in the serum did not significantly change in the 20% group in this experiment, the abundance of
Bifidobacterium decreased after they were fed 20% GP, and
Bifidobacterium was negatively correlated with T-AOC. Therefore, we speculated that 20% added GP may be the counterbalance of antioxidant and pro-oxidation effects in the body, but further studies are needed to confirm this.
The digestion and utilization of feed in ruminants are primarily linked to the capacity of the rumen microbial community to convert potentially digestible feed into metabolizable nutrients [
49]. Rumen microorganisms play a dual role, satisfying the majority of the nutritional requirements for ruminants and accounting for up to 90% of their metabolic needs [
50,
51]. The results of this experiment indicate that the diversity of rumen microorganisms in beef cattle remains unaltered. This suggests that including GP in the diet does not induce microbial disorders in the rumen of beef cattle during the fattening period. Generally, the rumen exhibits the highest proportion of Bacteroidetes, followed by Firmicutes [
52,
53,
54]. It is worth noting that the body weight of beef cattle in the 20% group decreased in this trial, suggesting a potential association with alterations in rumen species distribution (
Figure 1) and the characteristics of diet (
Table 2). Bacteroidetes have exhibited precise mechanisms of orchestrating the expression of polysaccharide-degrading enzymes, thereby regulating carbohydrate digestion and absorption at the genetic level [
55,
56]. Firmicutes play a role in promoting the digestion and absorption of proteins within animal organisms. Additionally, they can produce free enzymes, cellulosomes, or multi-enzyme complexes during the process of fiber digestion [
56]. This collaborative action facilitates the degradation of polysaccharides and the production of lactic acid and butyric acid, which can be utilized by the liver and muscle tissues [
57]. Furthermore, an investigation into the gastrointestinal tract of humans and mice indicated a positive association between the Firmicutes/Bacteroidetes ratios and obesity. The study suggested that Firmicutes/Bacteroidetes influence fat storage by regulating the energy absorption efficiency [
58,
59]. This aligns with our observation that the relative abundance of Firmicutes in the G20 group was diminished, consequently impacting the energy absorption from the diet in the beef cattle and reducing the FCR.
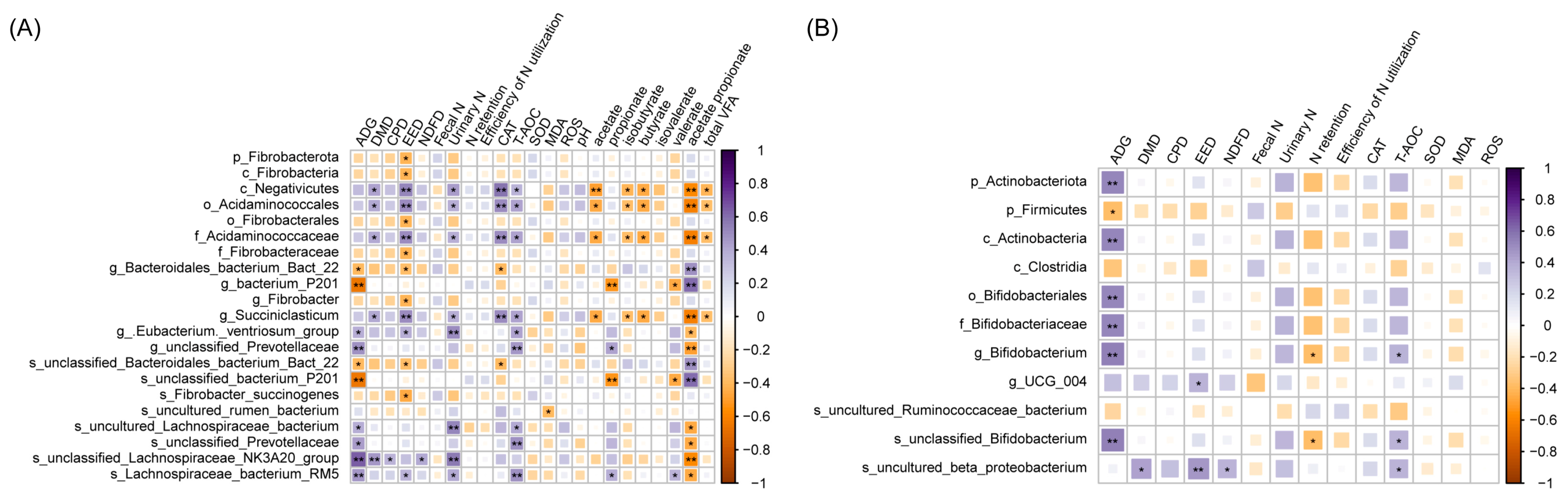
Additionally, the proportion of added GP will impact the composition of the rumen microbiota. Interestingly, including 20% GP in the diet increased the abundance of Fibrobacterium in the rumen. This observation may signify a metabolic strategy beef cattle employ to adapt to higher-lignocellulose diets.
F. succinogenes belongs to Fibrobacterota. The protein in the outer membrane that
Succinogenes possesses has a distinctive ability to bind cellulose, which plays a crucial role in fiber degradation [
56,
60,
61]. An elevated lignocellulose content promotes the growth of Fibrobacteria, facilitating their increased production of VFA, essential to the host animal’s maintenance, reproduction, and growth [
62]. However, the proliferation of fibrolytic bacteria may increase the acetate/propionate ratio, potentially negatively affecting the efficiency of dietary energy utilization. In this study, adding 10% GP promoted the enrichment of
Succiniclasticum.
Succiniclasticum, classified within the Negativicutes class, is the primary bacterium engaged in the succinic acid pathway, producing propionic acid. The relative abundance of
Succiniclasticum showed a positive correlation with propionic acid production [
63]. Hence, the addition of 10% GP can partially facilitate an increase in the acetate-to-propionate ratio in beef cattle, thereby contributing to a balance in the impact of
Succinogenes. The heatmap showed that the antioxidant index (CAT and T-AOC) had a significant positive correlation with
Succiniclasticum, indicating that the addition of 10% GP is conducive to improving the genes of
Succiniclasticum, thus improving the health of beef cattle.
Nevertheless, when the GP is increased to 20%, the stability of the microbial structure in the rumen diminishes. This could be attributed to the excessive addition of GP, disrupting the composition of the rumen microbial community and causing alterations in the keystone taxa of the rumen microorganisms. Keystone taxa significantly influence microbial communities and functions, irrespective of the abundance of microbiota across spatial and temporal dimensions [
64]. Katharine et al. [
65] argue that although positive interactions improve the overall metabolic efficiency, they reduce ecological stability. Thus, a higher neg/pos ratio indicates weaker microbial competition and a more stable network structure. In combination with other indicators, the network diagram became more complex with an increase in GP, but the microbial network diagram did not become more stable with an increase in GP. The stability of the microbial network and the antioxidant indexes both increased first and then decreased. These results suggested that the stability of the rumen microbiota may be affected by the antioxidant properties of GP after feeding, thus affecting the overall metabolic efficiency, which may also explain the decrease in body weight and the apparent digestibility of nutrients observed in G20.
In this study, rectum Firmicutes showed a relative abundance increase with an elevated GP intake, while Bacteroidetes linearly decreased. It is a pity that there is a paucity of studies on the rectal microbiota of ruminants. A study of human feces revealed a correlation between this trend and bacteria aiding the further breakdown of tannins into phenolic acids [
66]. However, whether the breakdown products in beef cattle align with those in the human gastrointestinal tract remains to be explored.
We observed an increase in the relative abundance of
Lachnospiraceae after feeding with 20% GP, potentially attributable to an elevation in polyphenols in the diet. Studies have demonstrated that polyphenols can modulate the relative abundance of
Lachnospiraceae in the rectum, aligning with the findings of this study [
67]. Moreover,
Lachnospiraceae have been associated with maintaining healthy immune homeostasis and preventing human inflammatory diseases [
68]. Including GP may play a role in modulating hindgut health in beef cattle. In this study, the relative abundance of UCG_004 decreased after adding 20% GP. UCG_004, a member of
Erysipelatoclostridiaceae, is a potential pathogen associated with colon diseases [
69]. Previous reports have indicated that polyphenols can enhance ruminants’ antioxidant activity by constraining pathogenic bacteria’s growth [
15,
70], and the results of this study suggest the potential that adding a certain proportion of GP may contribute to reducing inflammation in diseases.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}