
Obesogenic Diet in Mice Leads to Inflammation and Oxidative Stress in the Mother in Association with Sex-Specific Changes in Fetal Development, Inflammatory Markers and Placental Transcriptome
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Tissue Collection
2.3. Plasma and Serum Samples Analysis
2.4. Maternal Liver Assessments
2.4.1. Fat Content Assay
2.4.2. Glycogen Content Assay
2.4.3. Liver Steatosis
2.4.4. Liver Protein Levels
2.5. Labyrinth Zone Structure Analysis
2.6. Placental Labyrinth Zone RNA-Seq
2.7. Statistics
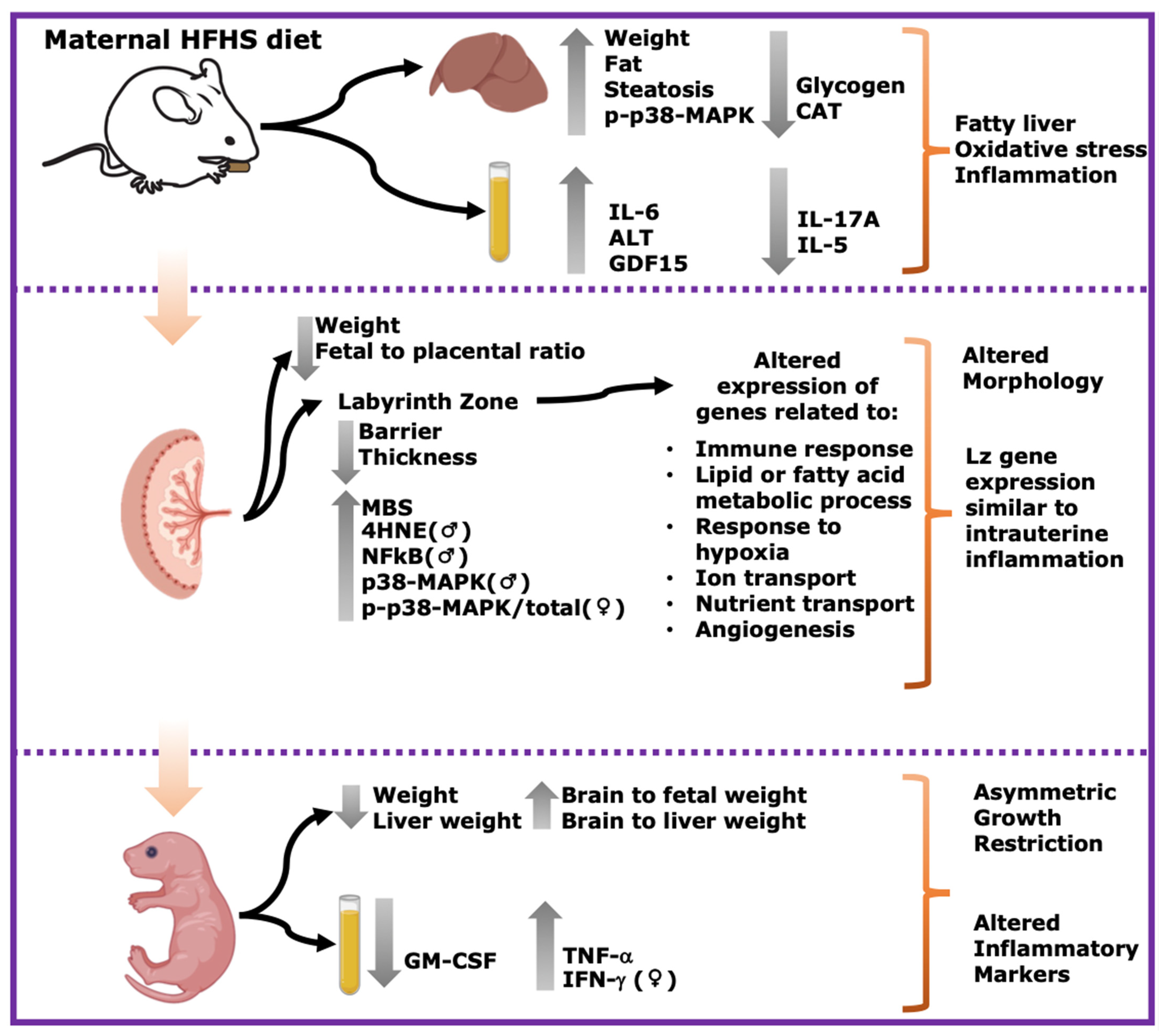
3. Results
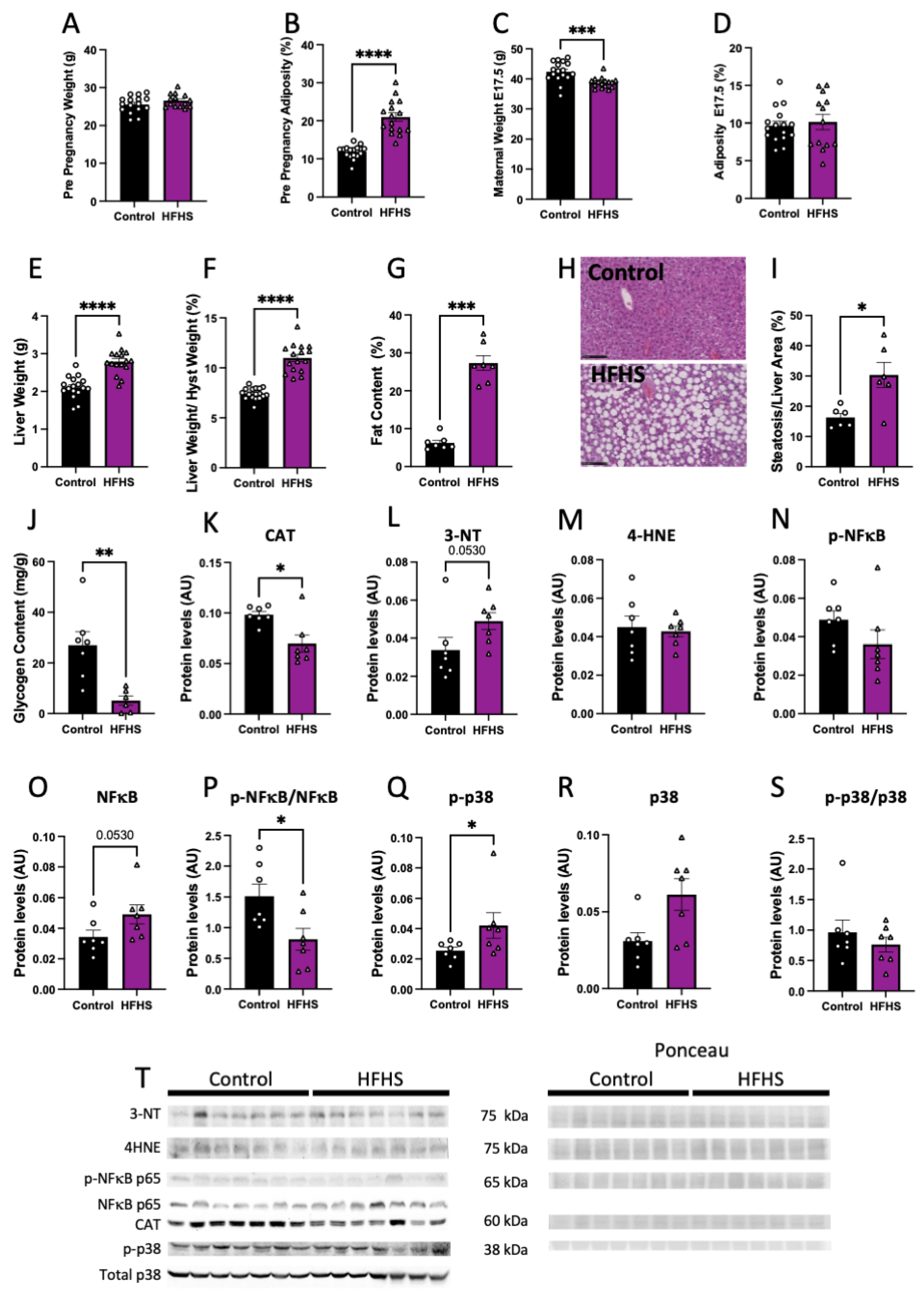
3.1. Maternal Body and Liver Tissue Composition, Oxidative Stress and Inflammatory Signaling
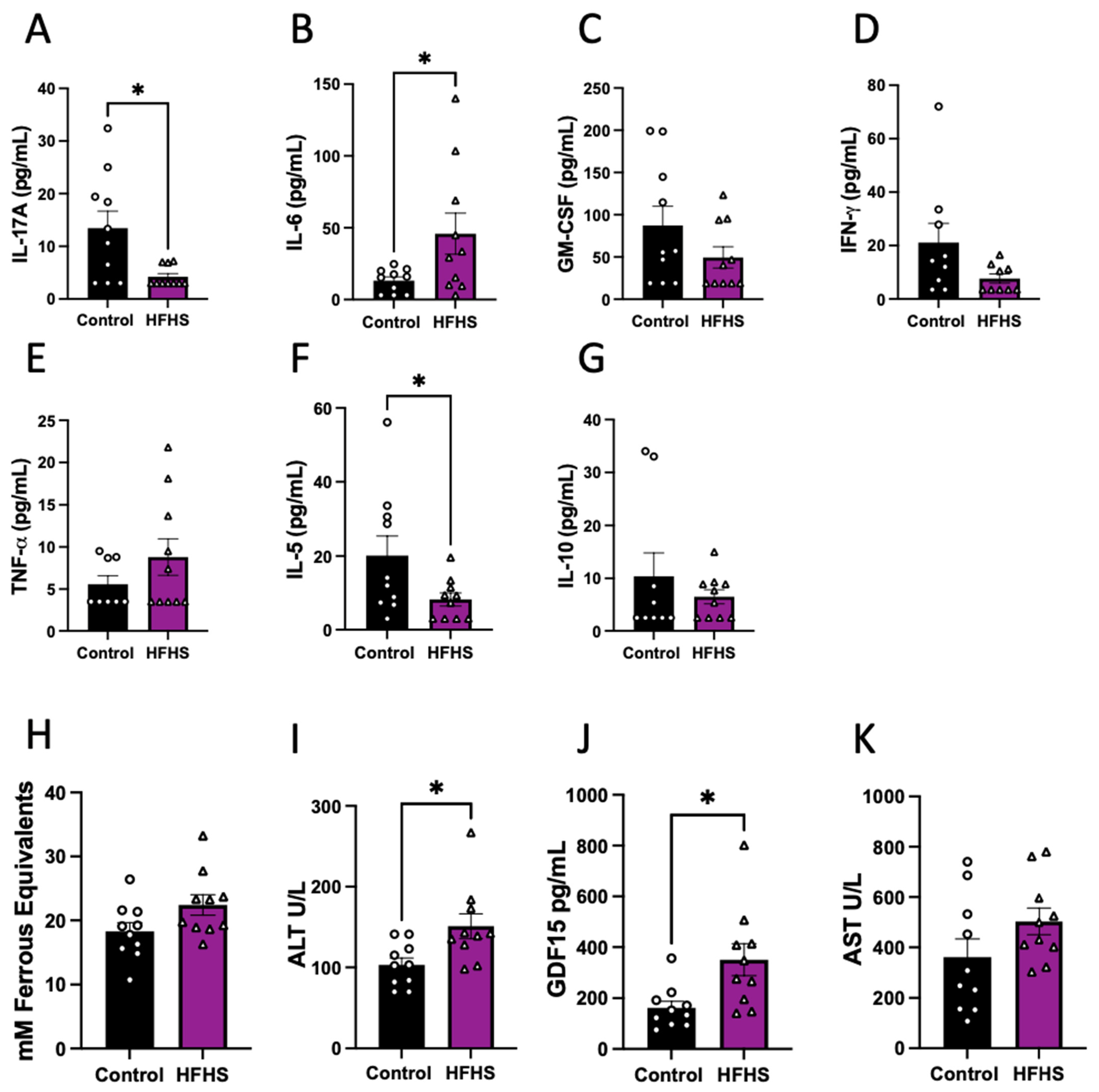
3.2. Maternal Plasma Levels of Damage Markers
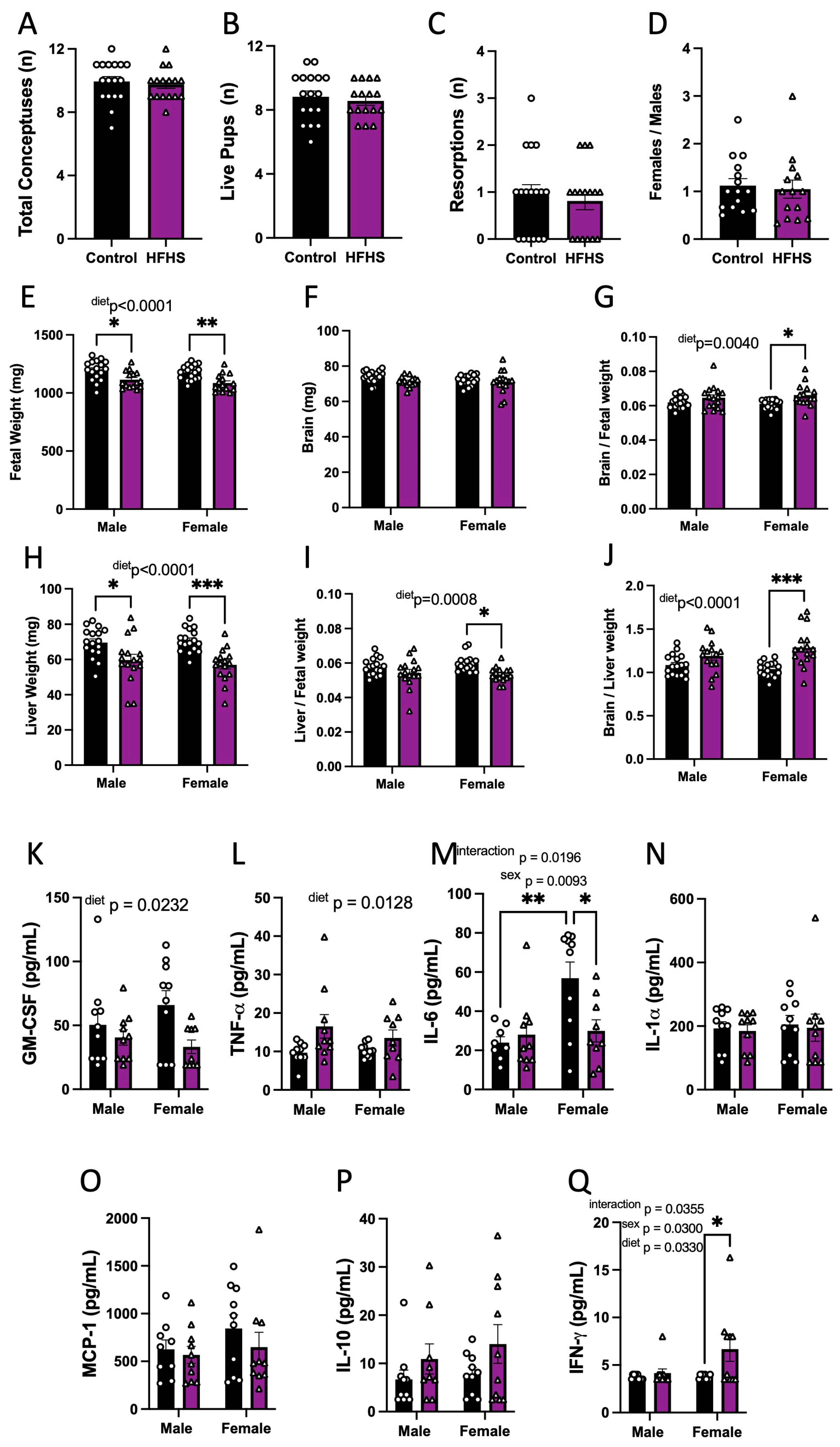
3.3. Fetal Viability and Growth
3.4. Fetal Circulating Inflammatory Markers
3.5. Placental Structural Development
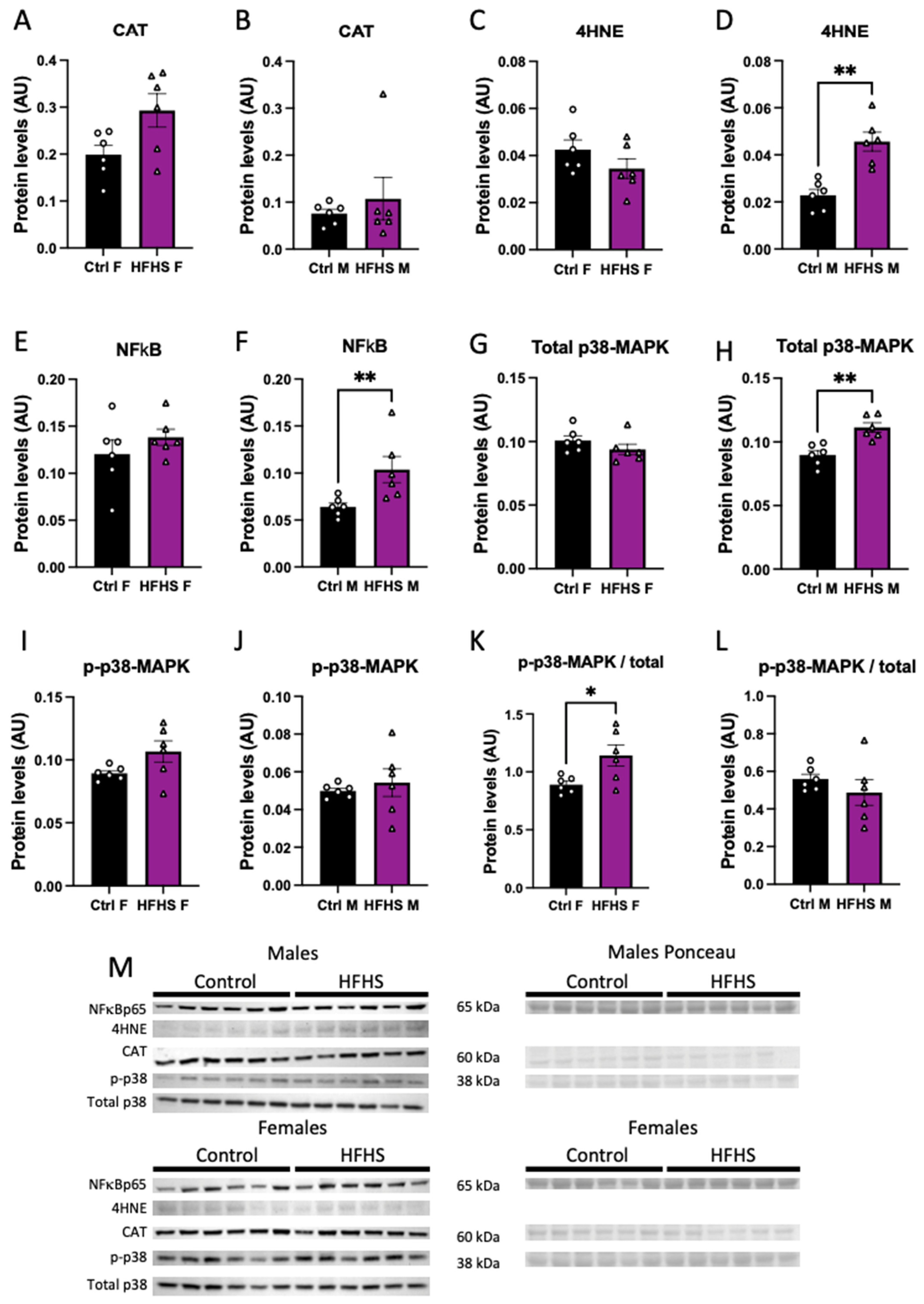
3.6. Placental Oxidative Stress, Inflammation, and Stress Signaling
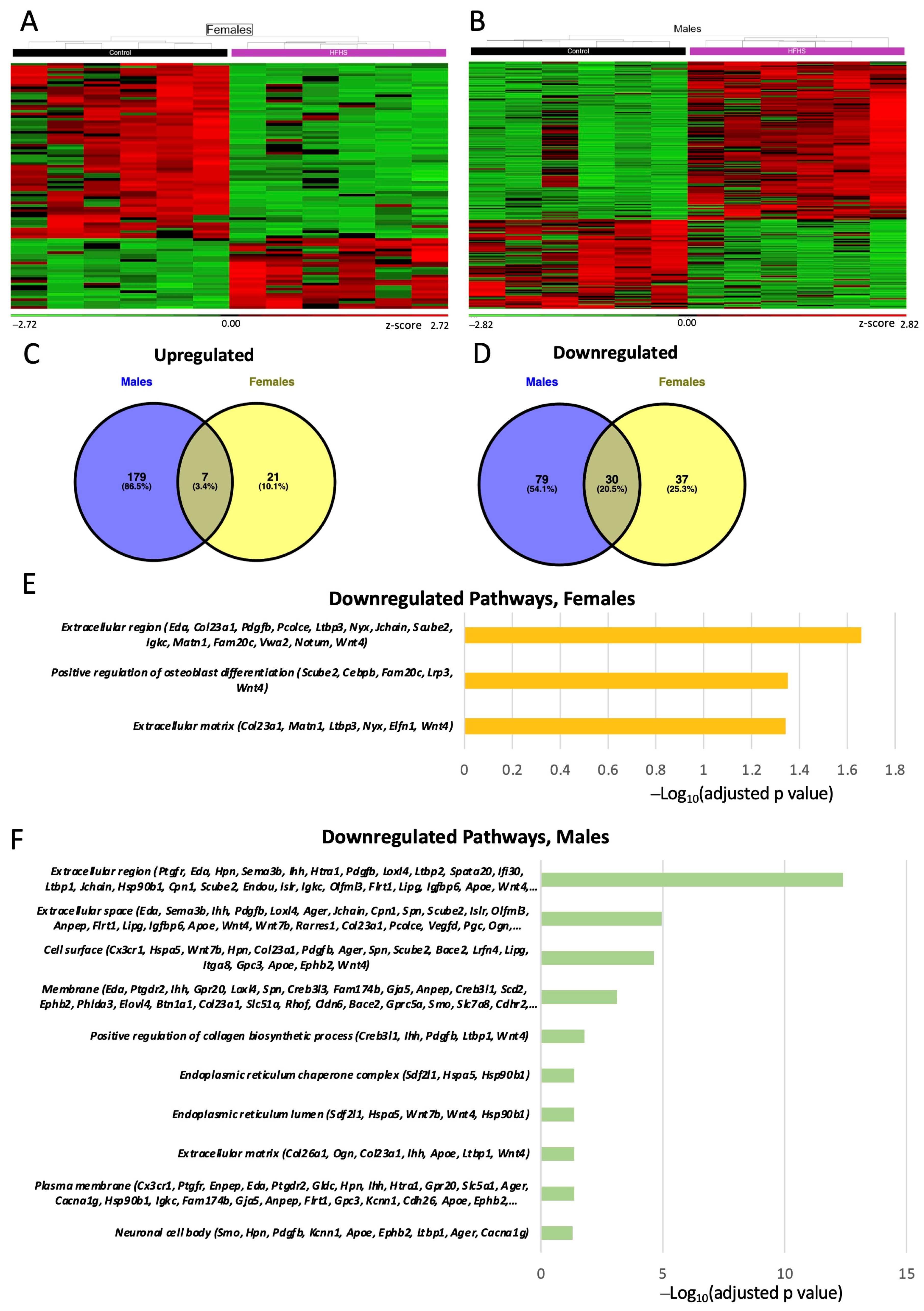
3.7. Placental Transcriptome
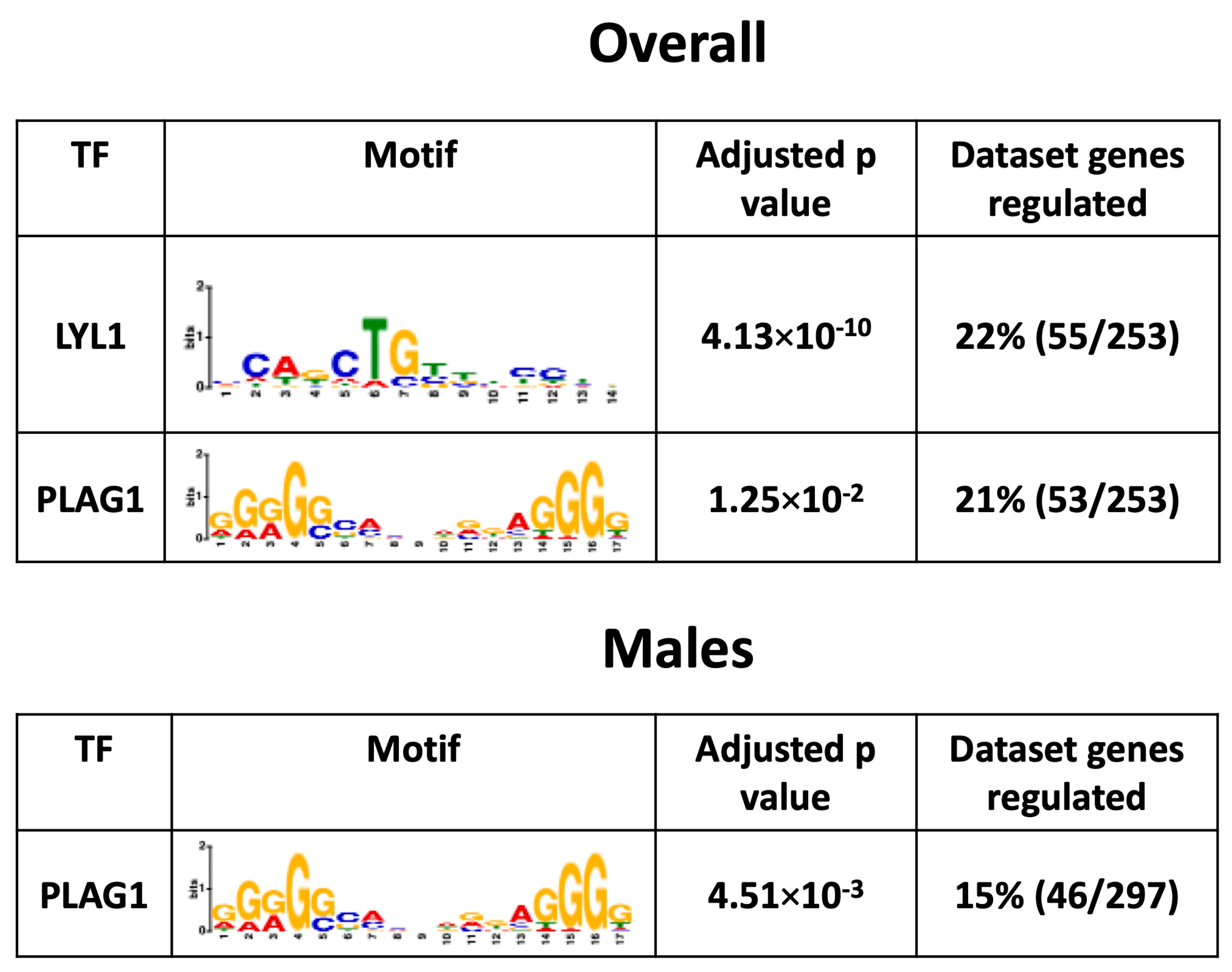
3.8. Placental Transcription Factors Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poston, L.; Caleyachetty, R.; Cnattingius, S.; Corvalán, C.; Uauy, R.; Herring, S.; Gillman, M.W. Preconceptional and Maternal Obesity: Epidemiology and Health Consequences. Lancet Diabetes Endocrinol. 2016, 4, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Marchi, J.; Berg, M.; Dencker, A.; Olander, E.K.; Begley, C. Risks Associated with Obesity in Pregnancy, for the Mother and Baby: A Systematic Review of Reviews. Obes. Rev. 2015, 16, 621–638. [Google Scholar] [CrossRef] [PubMed]
- Sasson, I.E.; Vitins, A.P.; Mainigi, M.A.; Moley, K.H.; Simmons, R.A. Pre-Gestational vs Gestational Exposure to Maternal Obesity Differentially Programs the Offspring in Mice. Diabetologia 2015, 58, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M. Maternal Obesity and Risk of Low Birth Weight, Fetal Growth Restriction, and Macrosomia: Multiple Analyses. Nutrients 2021, 13, 1213. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Kina, E.; Hivert, M.F. Maternal/Fetal Determinants of Insulin Resistance in Women during Pregnancy and in Offspring over Life. Curr. Diab. Rep. 2013, 13, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Musial, B.; Vaughan, O.R.; Fernandez-Twinn, D.S.; Voshol, P.; Ozanne, S.E.; Fowden, A.L.; Sferruzzi-Perri, A.N. A Western-Style Obesogenic Diet Alters Maternal Metabolic Physiology with Consequences for Fetal Nutrient Acquisition in Mice. J. Physiol. 2017, 595, 4875–4892. [Google Scholar] [CrossRef] [PubMed]
- Son, J.S.; Liu, X.; Tian, Q.; Zhao, L.; Chen, Y.; Hu, Y.; Chae, S.A.; de Avila, J.M.; Zhu, M.J.; Du, M. Exercise Prevents the Adverse Effects of Maternal Obesity on Placental Vascularization and Fetal Growth. J. Physiol. 2019, 597, 3333–3347. [Google Scholar] [CrossRef]
- Wallace, J.G.; Bellissimo, C.J.; Yeo, E.; Fei Xia, Y.; Petrik, J.J.; Surette, M.G.; Bowdish, D.M.E.; Sloboda, D.M. Obesity during Pregnancy Results in Maternal Intestinal Inflammation, Placental Hypoxia, and Alters Fetal Glucose Metabolism at Mid-Gestation. Sci. Rep. 2019, 9, 17621. [Google Scholar] [CrossRef]
- Reilly, S.M.; Saltiel, A.R. Adapting to Obesity with Adipose Tissue Inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jönsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary Factors and Low-Grade Inflammation in Relation to Overweight and Obesity. Br. J. Nutr. 2011, 106, S5–S78. [Google Scholar] [CrossRef]
- Stewart, F.M.; Freeman, D.J.; Ramsay, J.E.; Greer, I.A.; Caslake, M.; Ferrell, W.R. Longitudinal Assessment of Maternal Endothelial Function and Markers of Inflammation and Placental Function throughout Pregnancy in Lean and Obese Mothers. J. Clin. Endocrinol. Metab. 2007, 92, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Christian, L.M.; Porter, K. Longitudinal Changes in Serum Proinflammatory Markers across Pregnancy and Postpartum: Effects of Maternal Body Mass Index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Sferruzzi-Perri, A.N.; Camm, E.J. The Programming Power of the Placenta. Front. Physiol. 2016, 7, 181639. [Google Scholar] [CrossRef] [PubMed]
- Frias, A.E.; Morgan, T.K.; Evans, A.E.; Rasanen, J.; Oh, K.Y.; Thornburg, K.L.; Grove, K.L. Maternal High-Fat Diet Disturbs Uteroplacental Hemodynamics and Increases the Frequency of Stillbirth in a Nonhuman Primate Model of Excess Nutrition. Endocrinology 2011, 152, 2456–2464. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.J.; Du, M.; Nathanielsz, P.W.; Ford, S.P. Maternal Obesity Up-Regulates Inflammatory Signaling Pathways and Enhances Cytokine Expression in the Mid-Gestation Sheep Placenta. Placenta 2010, 31, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Farley, D.; Tejero, M.E.; Comuzzie, A.G.; Higgins, P.B.; Cox, L.; Werner, S.L.; Jenkins, S.L.; Li, C.; Choi, J.; Dick, E.J.; et al. Feto-Placental Adaptations to Maternal Obesity in the Baboon. Placenta 2009, 30, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-P.; Chen, X.; Li, M.-Q. Gestational Diabetes Induces Chronic Hypoxia Stress and Excessive Inflammatory Response in Murine Placenta. Int. J. Clin. Exp. Pathol. 2013, 6, 650–659. [Google Scholar]
- Kim, D.W.; Young, S.L.; Grattan, D.R.; Jasoni, C.L. Obesity during Pregnancy Disrupts Placental Morphology, Cell Proliferation, and Inflammation in a Sex-Specific Manner across Gestation in the Mouse. Biol. Reprod. 2014, 90, 1–11. [Google Scholar] [CrossRef]
- Gohir, W.; Kennedy, K.M.; Wallace, J.G.; Saoi, M.; Bellissimo, C.J.; Britz-McKibbin, P.; Petrik, J.J.; Surette, M.G.; Sloboda, D.M. High-fat Diet Intake Modulates Maternal Intestinal Adaptations to Pregnancy and Results in Placental Hypoxia, as Well as Altered Fetal Gut Barrier Proteins and Immune Markers. J. Physiol. 2019, 597, 3029–3051. [Google Scholar] [CrossRef]
- Nam, J.; Greenwald, E.; Jack-Roberts, C.; Ajeeb, T.T.; Malysheva, O.V.; Caudill, M.A.; Axen, K.; Saxena, A.; Semernina, E.; Nanobashvili, K.; et al. Choline Prevents Fetal Overgrowth and Normalizes Placental Fatty Acid and Glucose Metabolism in a Mouse Model of Maternal Obesity. J. Nutr. Biochem. 2017, 49, 80–88. [Google Scholar] [CrossRef]
- Kretschmer, T.; Schulze-edinghausen, M.; Turnwald, E.M.; Janoschek, R.; Bae-gartz, I.; Zentis, P.; Handwerk, M.; Wohlfarth, M.; Schauss, A.; Hucklenbruch-rother, E.; et al. Effect of Maternal Obesity in Mice on IL-6 Levels and Placental Endothelial Cell Homeostasis. Nutrients 2020, 12, 296. [Google Scholar] [CrossRef]
- D’Archivio, M.; Annuzzi, G.; Varì, R.; Filesi, C.; Giacco, R.; Scazzocchio, B.; Santangelo, C.; Giovannini, C.; Rivellese, A.A.; Masella, R. Predominant Role of Obesity/Insulin Resistance in Oxidative Stress Development. Eur. J. Clin. Investig. 2012, 42, 70–78. [Google Scholar] [CrossRef]
- Malti, N.; Merzouk, H.; Merzouk, S.A.; Loukidi, B.; Karaouzene, N.; Malti, A.; Narce, M. Oxidative Stress and Maternal Obesity: Feto-Placental Unit Interaction. Placenta 2014, 35, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-Associated Oxidative Stress: Strategies Finalized to Improve Redox State. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef]
- Alcala, M.; Gutierrez-Vega, S.; Castro, E.; Guzman-Gutiérrez, E.; Ramos-Álvarez, M.P.; Viana, M. Antioxidants and Oxidative Stress: Focus in Obese Pregnancies. Front. Physiol. 2018, 9, 1569. [Google Scholar] [CrossRef]
- Kubes, P.; Jenne, C.; Snyder, J. Immune Responses in the Liver. Annu. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human Fatty Liver Disease: Old Questions and New Insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef]
- Arisqueta, L.; Navarro-Imaz, H.; Labiano, I.; Rueda, Y.; Fresnedo, O. High-Fat Diet Overfeeding Promotes Nondetrimental Liver Steatosis in Female Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Sattar, N.; Forrest, E.; Preiss, D. Non-Alcoholic Fatty Liver Disease. BMJ Online 2014, 349, 45. [Google Scholar] [CrossRef]
- Azzaroli, F.; Mazzella, G.; Marchesini, G.; Brodosi, L.; Petroni, M.L. Fatty Liver in Pregnancy: A Narrative Review of Two Distinct Conditions. Expert Rev. Gastroenterol. Hepatol. 2020, 14, 127–135. [Google Scholar] [CrossRef]
- Hagström, H.; Höijer, J.; Ludvigsson, J.F.; Bottai, M.; Ekbom, A.; Hultcrantz, R.; Stephansson, O.; Stokkeland, K. Adverse Outcomes of Pregnancy in Women with Non-Alcoholic Fatty Liver Disease. Liver Int. 2016, 36, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Christoforou, E.R.; Sferruzzi-Perri, A.N. Molecular Mechanisms Governing Offspring Metabolic Programming in Rodent Models of in Utero Stress. Cell. Mol. Life Sci. 2020, 77, 4861–4898. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, L.M.; Nagao, M.; Kusinski, L.C.; Fernandez-Twinn, D.S.; Eliasson, L.; Ozanne, S.E. Exposure to Maternal Obesity Programs Sex Differences in Pancreatic Islets of the Offspring in Mice. Diabetologia 2020, 63, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Barke, T.L.; Money, K.M.; Du, L.; Serezani, A.; Gannon, M.; Mirnics, K.; Aronoff, D.M. Sex Modifies Placental Gene Expression in Response to Metabolic and Inflammatory Stress. Placenta 2019, 78, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Napso, T.; Lean, S.C.; Lu, M.; Mort, E.J.; Desforges, M.; Moghimi, A.; Bartels, B.; El-Bacha, T.; Fowden, A.L.; Camm, E.J.; et al. Diet-induced Maternal Obesity Impacts Feto-placental Growth and Induces Sex-specific Alterations in Placental Morphology, Mitochondrial Bioenergetics, Dynamics, Lipid Metabolism and Oxidative Stress in Mice. Acta Physiol. 2022, 234, e13795. [Google Scholar] [CrossRef] [PubMed]
- Roberts, V.H.J.; Smith, J.; McLea, S.A.; Heizer, A.B.; Richardson, J.L.; Myatt, L. Effect of Increasing Maternal Body Mass Index on Oxidative and Nitrative Stress in The Human Placenta. Placenta 2009, 30, 169–175. [Google Scholar] [CrossRef]
- Saben, J.; Lindsey, F.; Zhong, Y.; Thakali, K.; Badger, T.M.; Andres, A.; Gomez-Acevedo, H.; Shankar, K. Maternal Obesity Is Associated with a Lipotoxic Placental Environment. Placenta 2014, 35, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.A.; Riley, S.C.; Reynolds, R.M.; Barr, S.; Evans, M.; Statham, A.; Hor, K.; Jabbour, H.N.; Norman, J.E.; Denison, F.C. Placental Structure and Inflammation in Pregnancies Associated with Obesity. Placenta 2011, 32, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Twinn, D.S.; Gascoin, G.; Musial, B.; Carr, S.; Duque-Guimaraes, D.; Blackmore, H.L.; Alfaradhi, M.Z.; Loche, E.; Sferruzzi-Perri, A.N.; Fowden, A.L.; et al. Exercise Rescues Obese Mothers’ Insulin Sensitivity, Placental Hypoxia and Male Offspring Insulin Sensitivity. Sci. Rep. 2017, 7, 44650. [Google Scholar] [CrossRef]
- Appel, S.; Schulze-Edinghausen, M.; Kretschmer, T.; Storck, S.; Janoschek, R.; Bae-Gartz, I.; Handwerk, M.; Wohlfarth, M.; Nüsken, K.D.; Hucklenbruch-Rother, E.; et al. Maternal Obesity Attenuates Predelivery Inflammatory Reaction in C57BL/6N Mice. J. Reprod. Immunol. 2017, 122, 10–13. [Google Scholar] [CrossRef]
- Hoch, D.; Gauster, M.; Hauguel-de Mouzon, S.; Desoye, G. Diabesity-Associated Oxidative and Inflammatory Stress Signalling in the Early Human Placenta. Mol. Asp. Med. 2019, 66, 21–30. [Google Scholar] [CrossRef]
- Sferruzzi-Perri, A.N.; Vaughan, O.R.; Haro, M.; Cooper, W.N.; Musial, B.; Charalambous, M.; Pestana, D.; Ayyar, S.; Ferguson-Smith, A.C.; Burton, G.J.; et al. An Obesogenic Diet during Mouse Pregnancy Modifies Maternal Nutrient Partitioning and the Fetal Growth Trajectory. FASEB J. 2013, 27, 3928–3937. [Google Scholar] [CrossRef]
- Piao, D.; Ritchey, J.W.; Reed Holyoak, G.; Wall, C.R.; Sultana, N.; Murray, J.K.; Bartels, K.E. In Vivo Percutaneous Reflectance Spectroscopy of Fatty Liver Development in Rats Suggests That the Elevation of the Scattering Power Is an Early Indicator of Hepatic Steatosis. J. Innov. Opt. Health Sci. 2018, 11, 1850019. [Google Scholar] [CrossRef]
- Romero-Calvo, I.; Ocón, B.; Martínez-Moya, P.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; De Medina, F.S. Reversible Ponceau Staining as a Loading Control Alternative to Actin in Western Blots. Anal. Biochem. 2010, 401, 318–320. [Google Scholar] [CrossRef] [PubMed]
- Lean, S.C.; Candia, A.A.; Gulacsi, E.; Lee, G.C.L.; Sferruzzi-Perri, A.N. Obesogenic Diet in Mice Compromises Maternal Metabolic Physiology and Lactation Ability Leading to Reductions in Neonatal Viability. Acta Physiol. 2022, 236, e13861. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Dreos, R.; Ambrosini, G.; Groux, R.; Cavin Périer, R.; Bucher, P. The Eukaryotic Promoter Database in Its 30th Year: Focus on Non-Vertebrate Organisms. Nucleic Acids Res. 2017, 45, D51–D55. [Google Scholar] [CrossRef] [PubMed]
- McLeay, R.C.; Bailey, T.L. Motif Enrichment Analysis: A Unified Framework and an Evaluation on ChIP Data. BMC Bioinform. 2010, 11, 165. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Lien, Y.-C.; Zhang, Z.; Barila, G.; Green-Brown, A.; Elovitz, M.A.; Simmons, R.A. Intrauterine Inflammation Alters the Transcriptome and Metabolome in Placenta. Front. Physiol. 2020, 11, 592689. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front. Endocrinol. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- Crispi, F.; Miranda, J.; Gratacós, E. Long-Term Cardiovascular Consequences of Fetal Growth Restriction: Biology, Clinical Implications, and Opportunities for Prevention of Adult Disease. Am. J. Obstet. Gynecol. 2018, 218, S869–S879. [Google Scholar] [CrossRef] [PubMed]
- Inkster, A.M.; Fernández-Boyano, I.; Robinson, W.P. Sex Differences Are Here to Stay: Relevance to Prenatal Care. J. Clin. Med. 2021, 10, 3000. [Google Scholar] [CrossRef] [PubMed]
- Irimia, J.M.; Meyer, C.M.; Segvich, D.M.; Surendran, S.; DePaoli-Roach, A.A.; Morral, N.; Roach, P.J. Lack of Liver Glycogen Causes Hepatic Insulin Resistance and Steatosis in Mice. J. Biol. Chem. 2017, 292, 10455–10464. [Google Scholar] [CrossRef]
- Krähenbühl, L.; Lang, C.; Lüdes, S.; Seiler, C.; Schäfer, M.; Zimmermann, A.; Krähenbühl, S. Reduced Hepatic Glycogen Stores in Patients with Liver Cirrhosis. Liver Int. Off. J. Int. Assoc. Study Liver 2003, 23, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Yarrington, C.D. Obstetric Outcomes in Women with Nonalcoholic Fatty Liver Disease. Metab. Syndr. Relat. Disord. 2017, 15, 387–392. [Google Scholar] [CrossRef]
- Serviddio, G.; Bellanti, F.; Vendemiale, G.; Altomare, E. Mitochondrial Dysfunction in Nonalcoholic Steatohepatitis. Expert Rev. Gastroenterol. Hepatol. 2011, 5, 233–244. [Google Scholar] [CrossRef]
- Muriel, P. Role of Free Radicals in Liver Diseases. Hepatol. Int. 2009, 3, 526–536. [Google Scholar] [CrossRef]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The Role of Oxidative Stress during Inflammatory Processes. Biol. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef]
- Westenberger, G.; Sellers, J.; Fernando, S.; Junkins, S.; Han, S.M.; Min, K.; Lawan, A. Function of Mitogen-Activated Protein Kinases in Hepatic Inflammation. J. Cell. Signal. 2021, 2, 172–180. [Google Scholar] [PubMed]
- Hwang, S.; Wang, X.; Rodrigues, R.M.; Ma, J.; He, Y.; Seo, W.; Park, S.H.; Kim, S.-J.; Feng, D.; Gao, B. Protective and Detrimental Roles of P38α Mitogen-Activated Protein Kinase in Different Stages of Nonalcoholic Fatty Liver Disease. Hepatology 2020, 72, 873–891. [Google Scholar] [CrossRef]
- Luedde, T.; Schwabe, R.F. NF-κB in the Liver--Linking Injury, Fibrosis and Hepatocellular Carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Gaffen, S.L. IL-17 in Obesity and Adipogenesis. Cytokine Growth Factor Rev. 2010, 21, 449–453. [Google Scholar] [CrossRef]
- de Lima Kaminski, V.; Ellwanger, J.H.; Matte, M.C.C.; Savaris, R.F.; Vianna, P.; Chies, J.A.B. IL-17 Blood Levels Increase in Healthy Pregnancy but Not in Spontaneous Abortion. Mol. Biol. Rep. 2018, 45, 1565–1568. [Google Scholar] [CrossRef]
- Hountohotegbe, T.; Gbedande, K.; Agbota, G.; Ibitokou, S.; Massougbodji, A.; Deloron, P.; Fievet, N.; Luty, A.J.F. Circulating Cytokines Associated with Poor Pregnancy Outcomes in Beninese Exposed to Infection with Plasmodium Falciparum. Infect. Immun. 2020, 88, 10-1128. [Google Scholar] [CrossRef]
- Chêne, A.; Briand, V.; Ibitokou, S.; Dechavanne, S.; Massougbodji, A.; Deloron, P.; Luty, A.J.F.; Gamain, B.; Fievet, N. Placental Cytokine and Chemokine Profiles Reflect Pregnancy Outcomes in Women Exposed to Plasmodium Falciparum Infection. Infect. Immun. 2014, 82, 3783–3789. [Google Scholar] [CrossRef]
- Connor, K.L.; Chehoud, C.; Altrichter, A.; Chan, L.; Desantis, T.Z.; Lye, S.J. Maternal Metabolic, Immune, and Microbial Systems in Late Pregnancy Vary with Malnutrition in Mice. Biol. Reprod. 2018, 98, 579–592. [Google Scholar] [CrossRef]
- Prins, J.R.; Gomez-Lopez, N.; Robertson, S.A. Interleukin-6 in Pregnancy and Gestational Disorders. J. Reprod. Immunol. 2012, 95, 1–14. [Google Scholar] [CrossRef]
- Barron, A.; McCarthy, C.M.; O’Keeffe, G.W. Preeclampsia and Neurodevelopmental Outcomes: Potential Pathogenic Roles for Inflammation and Oxidative Stress? Mol. Neurobiol. 2021, 58, 2734–2756. [Google Scholar] [CrossRef]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The Maternal Interleukin-17a Pathway in Mice Promotes Autism-like Phenotypes in Offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Edlow, A.G. Maternal Obesity and Neurodevelopmental and Psychiatric Disorders in Offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Mahany, E.B.; Han, X.; Borges, B.C.; Da Silveira Cruz-Machado, S.; Allen, S.J.; Garcia-Galiano, D.; Hoenerhoff, M.J.; Bellefontaine, N.H.; Elias, C.F. Obesity and High-Fat Diet Induce Distinct Changes in Placental Gene Expression and Pregnancy Outcome. Endocrinology 2018, 159, 1718–1733. [Google Scholar] [CrossRef] [PubMed]
- Mayor, R.S.; Finch, K.E.; Zehr, J.; Morselli, E.; Neinast, M.D.; Frank, A.P.; Hahner, L.D.; Wang, J.; Rakheja, D.; Palmer, B.F.; et al. Maternal High-Fat Diet Is Associated with Impaired Fetal Lung Development. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, 360–368. [Google Scholar] [CrossRef]
- Musial, B.; Fernandez-Twinn, D.S.; Duque-Guimaraes, D.; Carr, S.K.; Fowden, A.L.; Ozanne, S.E.; Sferruzzi-Perri, A.N. Exercise Alters the Molecular Pathways of Insulin Signaling and Lipid Handling in Maternal Tissues of Obese Pregnant Mice. Physiol. Rep. 2019, 7, e14202. [Google Scholar] [CrossRef]
- Stuart, T.J.; O’Neill, K.; Condon, D.; Sasson, I.; Sen, P.; Xia, Y.; Simmons, R.A. Diet-Induced Obesity Alters the Maternal Metabolome and Early Placenta Transcriptome and Decreases Placenta Vascularity in the Mouse. Biol. Reprod. 2018, 98, 795–809. [Google Scholar] [CrossRef]
- Panchenko, P.E.; Voisin, S.; Jouin, M.; Jouneau, L.; Prézelin, A.; Lecoutre, S.; Breton, C.; Jammes, H.; Junien, C.; Gabory, A. Expression of Epigenetic Machinery Genes Is Sensitive to Maternal Obesity and Weight Loss in Relation to Fetal Growth in Mice. Clin. Epigenetics 2016, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Edlow, A.G.; Glass, R.M.; Smith, C.J.; Tran, P.K.; James, K.; Bilbo, S. Placental Macrophages: A Window Into Fetal Microglial Function in Maternal Obesity. Int. J. Dev. Neurosci. 2019, 77, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Fornes, R.; Manti, M.; Qi, X.; Vorontsov, E.; Sihlbom, C.; Nyström, J.; Jerlhag, E.; Maliqueo, M.; Hirschberg, A.L.; Carlström, M.; et al. Mice Exposed to Maternal Androgen Excess and Diet-Induced Obesity Have Altered Phosphorylation of Catechol-O-Methyltransferase in the Placenta and Fetal Liver. Int. J. Obes. 2019, 43, 2176–2188. [Google Scholar] [CrossRef]
- Robertson, S.A.; Sjöblom, C.; Jasper, M.J.; Norman, R.J.; Seamark, R.F. Granulocyte-Macrophage Colony-Stimulating Factor Promotes Glucose Transport and Blastomere Viability in Murine Preimplantation Embryos1. Biol. Reprod. 2001, 64, 1206–1215. [Google Scholar] [CrossRef]
- Sferruzzi-Perri, A.N.; Macpherson, A.M.; Roberts, C.T.; Robertson, S.A. Csf2 Null Mutation Alters Placental Gene Expression and Trophoblast Glycogen Cell and Giant Cell Abundance in Mice1. Biol. Reprod. 2009, 81, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, M.; Leszczyńska-Gorzelak, B.; Laskowska, K.; Oleszczuk, J. Evaluation of Maternal and Umbilical Serum TNFα Levels in Preeclamptic Pregnancies in the Intrauterine Normal and Growth-Restricted Fetus. J. Matern. Fetal Neonatal Med. 2006, 19, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Neta, G.I.; Von Ehrenstein, O.S.; Goldman, L.R.; Lum, K.; Sundaram, R.; Andrews, W.; Zhang, J. Umbilical Cord Serum Cytokine Levels and Risks of Small-for-Gestational-Age and Preterm Birth. Am. J. Epidemiol. 2010, 171, 859–867. [Google Scholar] [CrossRef]
- Ødegård, R. Umbilical Cord Plasma Interleukin-6 and Fetal Growth Restriction in Preeclampsia: A Prospective Study in Norway. Obstet. Gynecol. 2001, 98, 289–294. [Google Scholar] [CrossRef]
- Dosch, N.C.; Guslits, E.F.; Weber, M.B.; Murray, S.E.; Ha, B.; Coe, C.L.; Auger, A.P.; Kling, P.J. Maternal Obesity Affects Inflammatory and Iron Indices in Umbilical Cord Blood. J. Pediatr. 2016, 172, 20–28. [Google Scholar] [CrossRef]
- Wilson, R.M.; Marshall, N.E.; Jeske, D.R.; Purnell, J.Q.; Thornburg, K.; Messaoudi, I. Maternal Obesity Alters Immune Cell Frequencies and Responses in Umbilical Cord Blood Samples. Pediatr. Allergy Immunol. 2015, 26, 344–351. [Google Scholar] [CrossRef]
- Musa, E.; Salazar-Petres, E.; Arowolo, A.; Levitt, N.; Matjila, M.; Sferruzzi-Perri, A.N. Obesity and Gestational Diabetes Independently and Collectively Induce Specific Effects on Placental Structure, Inflammation and Endocrine Function in a Cohort of South African Women. J. Physiol. 2023, 601, 1287–1306. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiao, X.; Frank, S.J.; Lin, H.Y.; Xia, Y. Distinct Mechanisms of Induction of Hepatic Growth Hormone Resistance by Endogenous IL-6, TNF-α, and IL-1β. Am. J. Physiol.-Endocrinol. Metab. 2014, 307, E186–E198. [Google Scholar] [CrossRef]
- Gurzov, E.N.; Stanley, W.J.; Pappas, E.G.; Thomas, H.E.; Gough, D.J. The JAK/STAT Pathway in Obesity and Diabetes. FEBS J. 2016, 283, 3002–3015. [Google Scholar] [CrossRef]
- Monaco-Brown, M.; Lawrence, D.A. Obesity and Maternal-Placental-Fetal Immunology and Health. Front. Pediatr. 2022, 10, 859885. [Google Scholar] [CrossRef]
- Cordner, Z.A.; Khambadkone, S.G.; Boersma, G.J.; Song, L.; Summers, T.N.; Moran, T.H.; Tamashiro, K.L.K. Maternal High-Fat Diet Results in Cognitive Impairment and Hippocampal Gene Expression Changes in Rat Offspring. Exp. Neurol. 2019, 318, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.M.; Messaoudi, I. The Impact of Maternal Obesity during Pregnancy on Offspring Immunity. Mol. Cell. Endocrinol. 2015, 418 Pt 2, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Aaltonen, R.; Heikkinen, T.; Hakala, K.; Laine, K.; Alanen, A. Transfer of Proinflammatory Cytokines across Term Placenta. Obstet. Gynecol. 2005, 106, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Zaretsky, M.V.; Alexander, J.M.; Byrd, W.; Bawdon, R.E. Transfer of Inflammatory Cytokines across the Placenta. Obstet. Gynecol. 2004, 103, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, J.; Samuelsson, A.-M.; Jansson, T.; Holmäng, A. Interleukin-6 in the Maternal Circulation Reaches the Rat Fetus in Mid-Gestation. Pediatr. Res. 2006, 60, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.P.; Tayade, C.; Ashkar, A.A.; Hatta, K.; Zhang, J.; Croy, B.A. Interferon Gamma in Successful Pregnancies. Biol. Reprod. 2009, 80, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Sferruzzi-Perri, A.N.; Robertson, S.A.; Dent, L.A. Interleukin-5 Transgene Expression and Eosinophilia Are Associated with Retarded Mammary Gland Development in Mice1. Biol. Reprod. 2003, 69, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.; Mau, V.; Young, I.; Matthaei, K. Uterine Eosinophils and Reproductive Performance in Interleukin 5-Deficient Mice. Reproduction 2000, 120, 423–432. [Google Scholar] [CrossRef]
- Wu, H.-X.; Jin, L.-P.; Xu, B.; Liang, S.-S.; Li, D.-J. Decidual Stromal Cells Recruit Th17 Cells into Decidua to Promote Proliferation and Invasion of Human Trophoblast Cells by Secreting IL-17. Cell. Mol. Immunol. 2014, 11, 253–262. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Lv, X.; Liu, H. Interleukin-17 Promotes Proliferation, Migration, and Invasion of Trophoblasts via Regulating PPAR-γ/RXR-α/Wnt Signaling. Bioengineered 2022, 13, 1224–1234. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; Patterson, P.H. Activation of the Maternal Immune System Induces Endocrine Changes in the Placenta via IL-6. Brain Behav. Immun. 2011, 25, 604–615. [Google Scholar] [CrossRef]
- Jones, H.N.; Jansson, T.; Powell, T.L. IL-6 Stimulates System A Amino Acid Transporter Activity in Trophoblast Cells through STAT3 and Increased Expression of SNAT2. Am. J. Physiol.-Cell Physiol. 2009, 297, C1228–C1235. [Google Scholar] [CrossRef]
- Lager, S.; Jansson, N.; Olsson, A.L.; Wennergren, M.; Jansson, T.; Powell, T.L. Effect of IL-6 and TNF-α on Fatty Acid Uptake in Cultured Human Primary Trophoblast Cells. Placenta 2011, 32, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.P.; Chen, Y.H.; Gao, L.; Wang, P.; Wang, X.L.; Luo, B.; Li, J.; Xu, D.X. Differential Effects of High-Fat Diets before Pregnancy and/or during Pregnancy on Fetal Growth Development. Life Sci. 2018, 212, 241–250. [Google Scholar] [CrossRef]
- Evans, L.S.; Myatt, L. Sexual Dimorphism in the Effect of Maternal Obesity on Antioxidant Defense Mechanisms in the Human Placenta. Placenta 2017, 51, 64–69. [Google Scholar] [CrossRef]
- Mudgett, J.S.; Ding, J.; Guh-Siesel, L.; Chartrain, N.A.; Yang, L.; Gopal, S.; Shen, M.M. Essential Role for P38α Mitogen-Activated Protein Kinase in Placental Angiogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 10454–10459. [Google Scholar] [CrossRef] [PubMed]
- Gabory, A.; Ferry, L.; Fajardy, I.; Jouneau, L.; Gothié, J.D.; Vigé, A.; Fleur, C.; Mayeur, S.; Gallou-Kabani, C.; Gross, M.S.; et al. Maternal Diets Trigger Sex-Specific Divergent Trajectories of Gene Expression and Epigenetic Systems in Mouse Placenta. PLoS ONE 2012, 7, e47986. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Xie, Y.; Zhao, Q.; Peng, W.; Guo, C.; Zhang, J.; Zhang, L. Polymorphisms and Gene-Gene Interaction in AGER/IL6 Pathway Might Be Associated with Diabetic Ischemic Heart Disease. J. Pers. Med. 2022, 12, 392. [Google Scholar] [CrossRef]
- Zhu, H.; Meissner, L.E.; Byrnes, C.; Tuymetova, G.; Tifft, C.J.; Proia, R.L. The Complement Regulator Susd4 Influences Nervous-System Function and Neuronal Morphology in Mice. iScience 2020, 23, 100957. [Google Scholar] [CrossRef]
- Konishi, H. Advanced Glycation End Products Induce Secretion of Chemokines and Apoptosis in Human First Trimester Trophoblasts. Hum. Reprod. 2004, 19, 2156–2162. [Google Scholar] [CrossRef]
- Shirasuna, K.; Seno, K.; Ohtsu, A.; Shiratsuki, S.; Ohkuchi, A.; Suzuki, H.; Matsubara, S.; Nagayama, S.; Iwata, H.; Kuwayama, T. AGEs and HMGB1 Increase Inflammatory Cytokine Production from Human Placental Cells, Resulting in an Enhancement of Monocyte Migration. Am. J. Reprod. Immunol. 2016, 75, 557–568. [Google Scholar] [CrossRef]
- Alexander, K.L.; Mejia, C.A.; Jordan, C.; Nelson, M.B.; Howell, B.M.; Jones, C.M.; Reynolds, P.R.; Arroyo, J.A. Differential Receptor for Advanced Glycation End Products Expression in Preeclamptic, Intrauterine Growth Restricted, and Gestational Diabetic Placentas. Am. J. Reprod. Immunol. 2016, 75, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, Z.; Zhang, L.; Shi, Y.; Qi, J.; Chang, A.; Gao, J.; Feng, Y.; Yang, X. HMGB1–RAGE Signaling Pathway in Severe Preeclampsia. Placenta 2015, 36, 1148–1152. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Y.; Yue, C.; Ye, Y.; Chen, P.; Peng, W.; Wang, Y. Accumulation of Advanced Glycation End Products Involved in Inflammation and Contributing to Severe Preeclampsia, in Maternal Blood, Umbilical Blood and Placental Tissues. Gynecol. Obstet. Investig. 2017, 82, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, R.; Falck, P.; Hellström, M.; Lindahl, P.; Boström, H.; Franklin, G.; Ährlund-Richter, L.; Pollard, J.; Soriano, P.; Betsholtz, C. PDGFB Regulates the Development of the Labyrinthine Layer of the Mouse Fetal Placenta. Dev. Biol. 1999, 212, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, A.; Lechner, A.J.; Ueno, M.; Acharya, A.; Van Handel, B.; Wang, Y.; Iruela-Arispe, M.L.; Tallquist, M.D.; Mikkola, H.K.A. Trophoblasts Regulate the Placental Hematopoietic Niche through PDGF-B Signaling. Dev. Cell 2012, 22, 651–659. [Google Scholar] [CrossRef] [PubMed]
- De Barros Mucci, D.; Kusinski, L.C.; Wilsmore, P.; Loche, E.; Pantaleão, L.C.; Ashmore, T.J.; Blackmore, H.L.; Fernandez-Twinn, D.S.; Carmo, M.D.G.T.D.; Ozanne, S.E. Impact of Maternal Obesity on Placental Transcriptome and Morphology Associated with Fetal Growth Restriction in Mice. Int. J. Obes. 2020, 44, 1087–1096. [Google Scholar] [CrossRef]
- Natale, B.V.; Mehta, P.; Vu, P.; Schweitzer, C.; Gustin, K.; Kotadia, R.; Natale, D.R.C. Reduced Uteroplacental Perfusion Pressure (RUPP) Causes Altered Trophoblast Differentiation and Pericyte Reduction in the Mouse Placenta Labyrinth. Sci. Rep. 2018, 8, 17162. [Google Scholar] [CrossRef]
- Plows, J.F.; Vickers, M.H.; Ganapathy, T.P.; Bridge-Comer, P.E.; Stanley, J.L.; Reynolds, C.M. Interleukin-1 Receptor-1 Deficiency Impairs Metabolic Function in Pregnant and Non-Pregnant Female Mice. Mol. Nutr. Food Res. 2021, 65, 1900770. [Google Scholar] [CrossRef]
- Dawson, P.A.; Rakoczy, J.; Simmons, D.G. Placental, Renal, and Ileal Sulfate Transporter Gene Expression in Mouse Gestation. Biol. Reprod. 2012, 87, 43. [Google Scholar] [CrossRef]
- Rakoczy, J.; Zhang, Z.; Bowling, F.G.; Dawson, P.A.; Simmons, D.G. Loss of the Sulfate Transporter Slc13a4 in Placenta Causes Severe Fetal Abnormalities and Death in Mice. Cell Res. 2015, 25, 1273–1276. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.L.; Leemaqz, S.Y.; Goh, Z.; McAninch, D.; Jankovic-Karasoulos, T.; Leghi, G.E.; Phillips, J.A.; Colafella, K.M.; Tran, C.; O’Leary, S.; et al. Zinc Is a Critical Regulator of Placental Morphogenesis and Maternal Hemodynamics during Pregnancy in Mice. Sci. Rep. 2017, 7, 15137. [Google Scholar] [CrossRef] [PubMed]
- Altmäe, S.; Segura, M.T.; Esteban, F.J.; Bartel, S.; Brandi, P.; Irmler, M.; Beckers, J.; Demmelmair, H.; López-Sabater, C.; Koletzko, B.; et al. Maternal Pre-Pregnancy Obesity Is Associated with Altered Placental Transcriptome. PLoS ONE 2017, 12, e0169223. [Google Scholar] [CrossRef]
- Starks, R.R.; Abu Alhasan, R.; Kaur, H.; Pennington, K.A.; Schulz, L.C.; Tuteja, G. Transcription Factor PLAGL1 Is Associated with Angiogenic Gene Expression in the Placenta. Int. J. Mol. Sci. 2020, 21, 8317. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Platas, I.; Martin-Trujillo, A.; Petazzi, P.; Guillaumet-Adkins, A.; Esteller, M.; Monk, D. Altered Expression of the Imprinted Transcription Factor PLAGL1 Deregulates a Network of Genes in the Human IUGR Placenta. Hum. Mol. Genet. 2014, 23, 6275–6285. [Google Scholar] [CrossRef] [PubMed]
- Pirot, N.; Deleuze, V.; El-Hajj, R.; Dohet, C.; Sablitzky, F.; Couttet, P.; Mathieu, D.; Pinet, V. LYL1 Activity Is Required for the Maturation of Newly Formed Blood Vessels in Adulthood. Blood 2010, 115, 5270–5279. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ren, D.; Arkoun, B.; Kaushik, A.-L.; Matherat, G.; Lécluse, Y.; Filipp, D.; Vainchenker, W.; Raslova, H.; Plo, I.; et al. Lyl-1 Regulates Primitive Macrophages and Microglia Development. Commun. Biol. 2021, 4, 1382. [Google Scholar] [CrossRef] [PubMed]
- Kadakia, R.; Zheng, Y.; Zhang, Z.; Zhang, W.; Josefson, J.L.; Hou, L. Association of Cord Blood Methylation with Neonatal Leptin: An Epigenome Wide Association Study. PLoS ONE 2019, 14, e0226555. [Google Scholar] [CrossRef] [PubMed]
- Czogała, W.; Czogała, M.; Strojny, W.; Wątor, G.; Wołkow, P.; Wójcik, M.; Bik Multanowski, M.; Tomasik, P.; Wędrychowicz, A.; Kowalczyk, W.; et al. Methylation and Expression of FTO and PLAG1 Genes in Childhood Obesity: Insight into Anthropometric Parameters and Glucose–Lipid Metabolism. Nutrients 2021, 13, 1683. [Google Scholar] [CrossRef]
- Lopes-Ramos, C.M.; Chen, C.-Y.; Kuijjer, M.L.; Paulson, J.N.; Sonawane, A.R.; Fagny, M.; Platig, J.; Glass, K.; Quackenbush, J.; DeMeo, D.L. Sex Differences in Gene Expression and Regulatory Networks across 29 Human Tissues. Cell Rep. 2020, 31, 107795. [Google Scholar] [CrossRef]
- Howerton, C.L.; Bale, T.L. Targeted Placental Deletion of OGT Recapitulates the Prenatal Stress Phenotype Including Hypothalamic Mitochondrial Dysfunction. Proc. Natl. Acad. Sci. USA 2014, 111, 9639–9644. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Cruz, M.; Palatnik, A.; Olivier-Van Stichelen, S. O-GlcNAc Transferase Contributes to Sex-Specific Placental Deregulation in Gestational Diabetes. Placenta 2023, 131, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Sex-Specific Placental Responses in Fetal Development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [PubMed]
- Kalisch-Smith, J.I.; Simmons, D.G.; Pantaleon, M.; Moritz, K.M. Sex Differences in Rat Placental Development: From Pre-Implantation to Late Gestation. Biol. Sex Differ. 2017, 8, 17. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | HFHS | p Value | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | ||||||||||||
| Structure or Measurement | Mean | ± | SEM | Mean | ± | SEM | Mean | ± | SEM | Mean | ± | SEM | Diet | Sex | Interaction |
| Placental Weight (g) | 0.10 | ± | 0.0023 | 0.09 | ± | 0.0012 | 0.09 | ± | 0.0026 | 0.09 | ± | 0.0018 | 0.0018 | <0.0001 | 0.2049 |
| Fetal to Placental Ratio | 11.91 | ± | 0.3451 | 12.81 | ± | 0.3555 | 11.82 | ± | 0.3263 | 12.40 | ± | 0.3394 | 0.2474 | 0.0089 | 0.3822 |
| Db, mm3 | 17.09 | ± | 1.3630 | 12.65 | ± | 1.2330 | 15.62 | ± | 1.5460 | 11.79 | ± | 0.9515 | 0.3895 | 0.0051 | 0.8213 |
| Jz, mm3 | 32.87 | ± | 2.2400 | 29.86 | ± | 4.1160 | 30.61 | ± | 2.7510 | 32.92 | ± | 2.1070 | 0.8902 | 0.9035 | 0.3634 |
| Lz, mm3 | 46.34 | ± | 2.2340 | 39.56 | ± | 1.2790 | 44.24 | ± | 1.0410 | 40.83 | ± | 2.1900 | 0.8160 | 0.0084 | 0.3496 |
| Lz—FC, mm3 | 19.93 | ± | 1.7370 | 16.38 | ± | 0.9625 | 16.65 | ± | 0.9153 | 17.03 | ± | 1.5020 | 0.3403 | 0.2512 | 0.1578 |
| Lz—MBS, mm3 | 11.30 | ± | 0.5420 | 8.53 | ± | 0.2379 | 12.70 | ± | 0.8470 | 10.48 | ± | 0.7709 | 0.0190 | 0.0010 | 0.6786 |
| Lz—TB, mm3 | 15.11 | ± | 0.7809 | 14.65 | ± | 0.9512 | 14.90 | ± | 0.7750 | 13.31 | ± | 1.1350 | 0.4019 | 0.2718 | 0.5404 |
| Capillary Length, µm | 67.62 | ± | 2.0190 | 60.81 | ± | 1.6510 | 65.70 | ± | 4.6930 | 63.97 | ± | 6.1490 | 0.8794 | 0.3014 | 0.5353 |
| Barrier Thickness, µm | 3.24 | ± | 0.0905 | 3.63 | ± | 0.1157 | 3.10 | ± | 0.1389 | 3.18 | ± | 0.1912 | 0.0424 | 0.1014 | 0.2900 |
| TDC, cm2*min−1*kPA−1 | 0.03 | ± | 0.0021 | 0.01 | ± | 0.0011 | 0.03 | ± | 0.0026 | 0.01 | ± | 0.0006 | 0.9389 | <0.0001 | 0.8935 |
| Surface Area, cm2 | 0.05 | ± | 0.0022 | 0.02 | ± | 0.0008 | 0.04 | ± | 0.0023 | 0.02 | ± | 0.0011 | 0.4452 | <0.0001 | 0.3730 |
| Capillary Diameter, µm | 19.23 | ± | 0.6491 | 18.48 | ± | 0.3735 | 18.11 | ± | 0.7099 | 18.49 | ± | 0.7150 | 0.3969 | 0.7729 | 0.3905 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candia, A.A.; Lean, S.C.; Zhang, C.X.W.; McKeating, D.R.; Cochrane, A.; Gulacsi, E.; Herrera, E.A.; Krause, B.J.; Sferruzzi-Perri, A.N. Obesogenic Diet in Mice Leads to Inflammation and Oxidative Stress in the Mother in Association with Sex-Specific Changes in Fetal Development, Inflammatory Markers and Placental Transcriptome. Antioxidants 2024, 13, 411. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040411
Candia AA, Lean SC, Zhang CXW, McKeating DR, Cochrane A, Gulacsi E, Herrera EA, Krause BJ, Sferruzzi-Perri AN. Obesogenic Diet in Mice Leads to Inflammation and Oxidative Stress in the Mother in Association with Sex-Specific Changes in Fetal Development, Inflammatory Markers and Placental Transcriptome. Antioxidants. 2024; 13(4):411. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040411
Chicago/Turabian StyleCandia, Alejandro A., Samantha C. Lean, Cindy X. W. Zhang, Daniel R. McKeating, Anna Cochrane, Edina Gulacsi, Emilio A. Herrera, Bernardo J. Krause, and Amanda N. Sferruzzi-Perri. 2024. "Obesogenic Diet in Mice Leads to Inflammation and Oxidative Stress in the Mother in Association with Sex-Specific Changes in Fetal Development, Inflammatory Markers and Placental Transcriptome" Antioxidants 13, no. 4: 411. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040411