
Immunogenicity of a Candidate DTacP-sIPV Combined Vaccine and Its Protection Efficacy against Pertussis in a Rhesus Macaque Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Use and Welfare
2.2. Vaccine Preparation
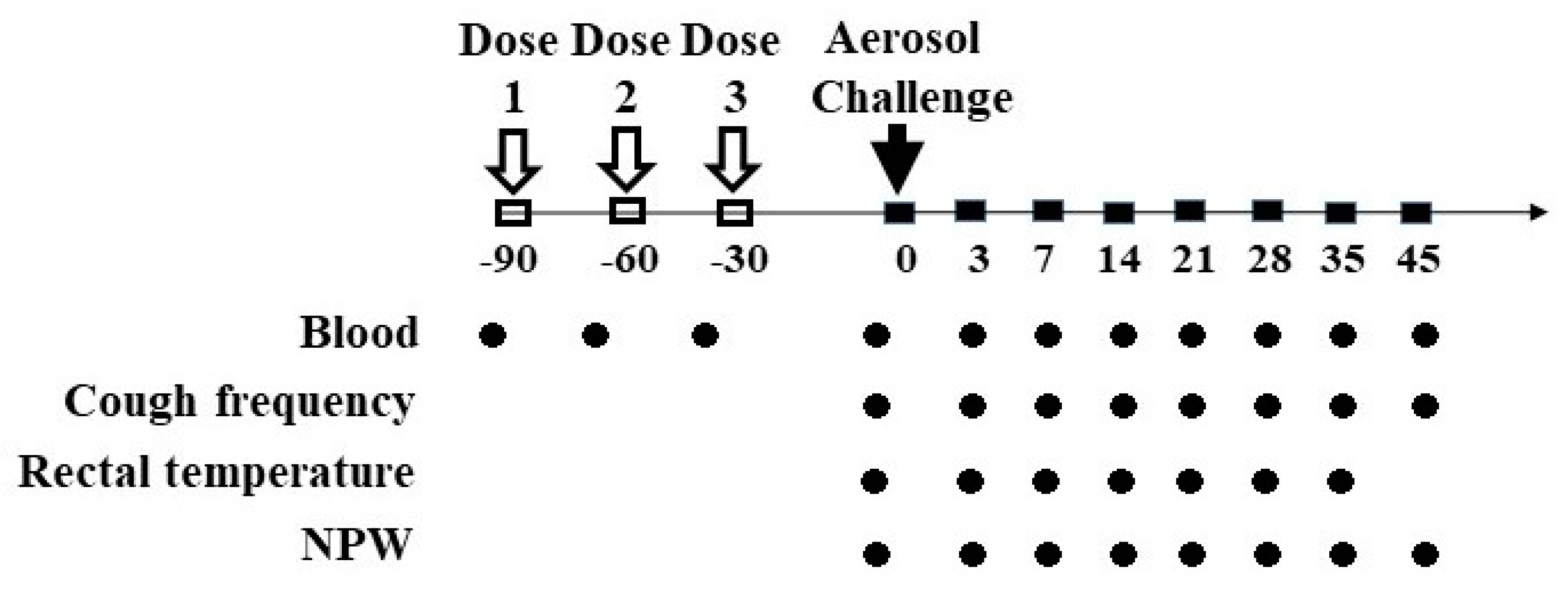
2.3. Grouping, Vaccination, and Pertussis Challenge
2.4. Evaluation of Animal
2.5. Serology Antibody
2.6. Measurement of Cytokines
2.7. Histopathological Observations of a Rhesus Macaque after Challenge
2.8. Statistics
3. Results
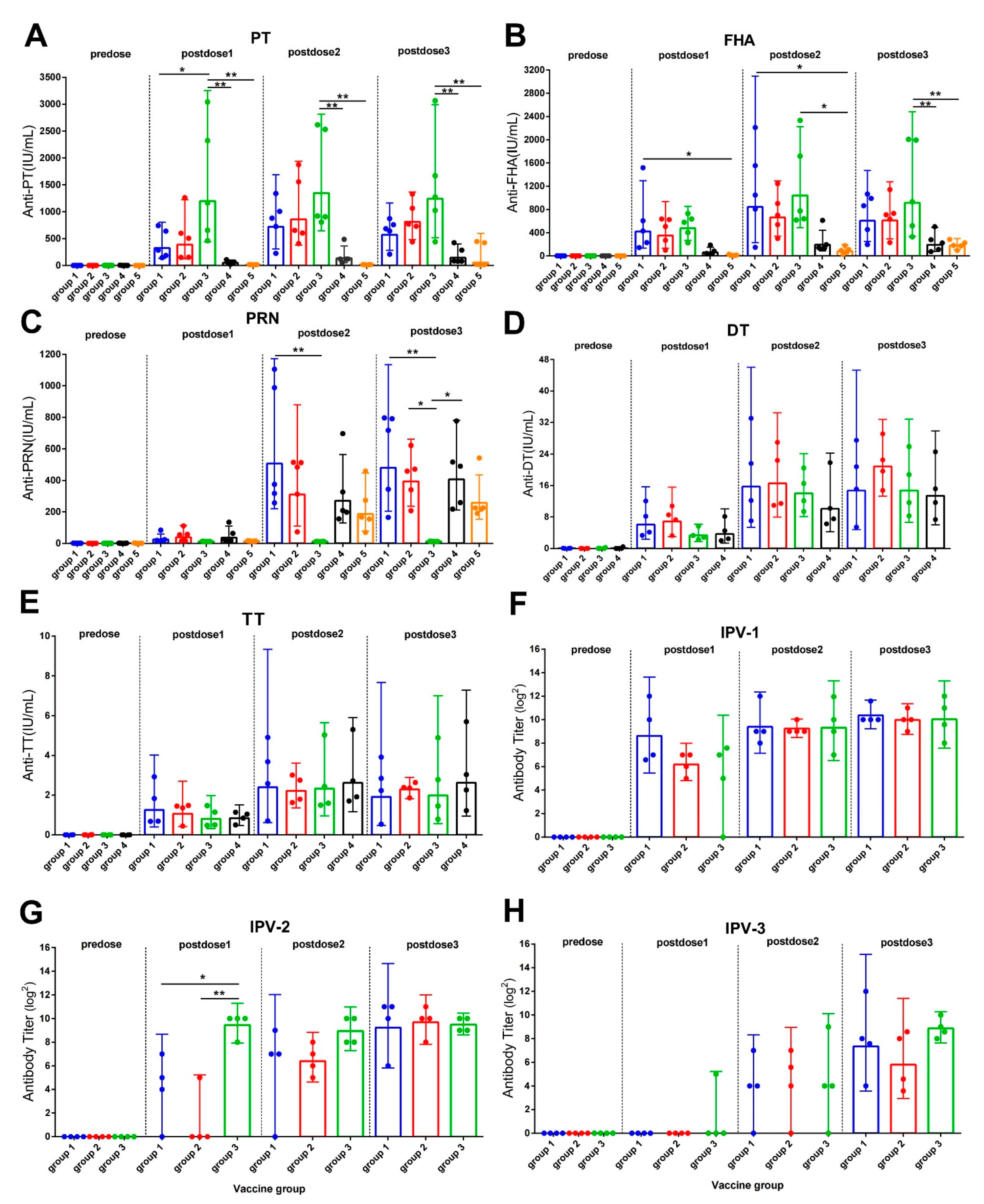
3.1. Antibody Response after Vaccine Immunization
3.2. Antibody Responses after Aerosol Challenge
3.3. Changes in Cytokine Levels of Rhesus Macaques after Challenge
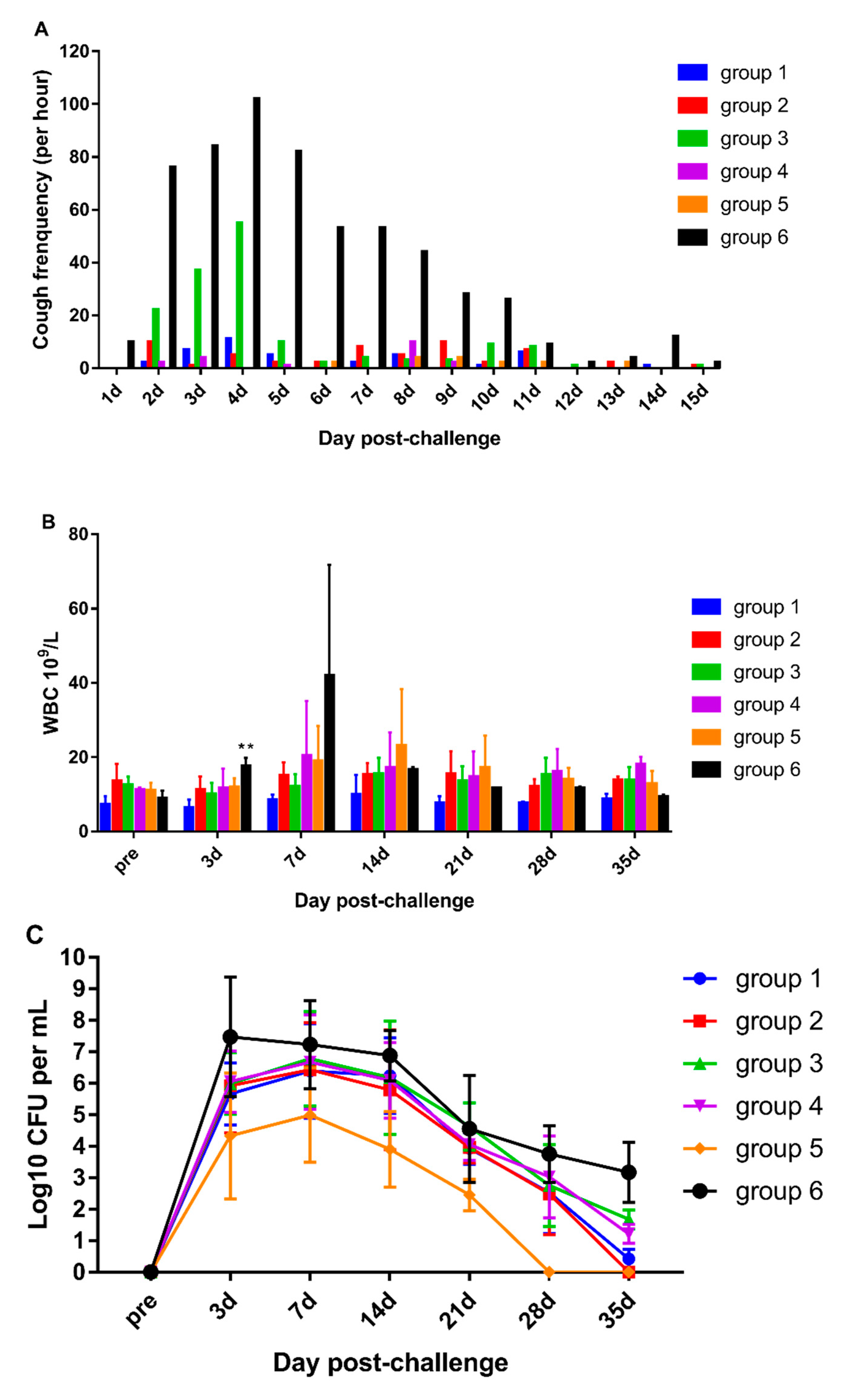
3.4. DTacP-sIPV Protection Efficacy against Pertussis
3.5. Pathological Characteristics of the Rhesus Macaque from the Negative Control Group after Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maman, K.; Zollner, Y.; Greco, D.; Duru, G.; Sendyona, S.; Remy, V. The value of childhood combination vaccines: From beliefs to evidence. Hum. Vaccines Immunother. 2015, 11, 2132–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidor, E.; Soubeyrand, B. Manufacturing DTaP-based combination vaccines: Industrial challenges around essential public health tools. Expert Rev. Vaccines 2016, 15, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Johns, T.L.; Hutter, G.E. New combination vaccines: DTaP-IPV (Kinrix) and DTaP-IPV/Hib (Pentacel). Ann. Pharmacother. 2010, 44, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Black, S.; Friedland, L.R.; Schuind, A.; Howe, B. Immunogenicity and safety of a combined DTaP-IPV vaccine compared with separate DTaP and IPV vaccines when administered as pre-school booster doses with a second dose of MMR vaccine to healthy children aged 4–6 years. Vaccine 2006, 24, 6163–6171. [Google Scholar] [CrossRef] [PubMed]
- Weston, W.M.; Klein, N.P. Kinrix: A new combination DTaP-IPV vaccine for children aged 4-6 years. Expert Rev. Vaccines 2008, 7, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Polio vaccines and polio immunization in the pre-eradication era: WHO position paper--recommendations. Vaccine 2010, 28, 6943–6944. [Google Scholar] [CrossRef]
- World Health Organization. Polio eradication & endgame strategic plan 2013-2018. Glob. Polio. Erad. Initiat. Work Draft. 2013, 23, 1–99. [Google Scholar]
- Melvin, J.A.; Scheller, E.V.; Miller, J.F.; Cotter, P.A. Bordetella pertussis pathogenesis: Current and future challenges. Nat. Rev. Microbiol. 2014, 12, 274. [Google Scholar] [CrossRef] [Green Version]
- Rohani, P.; Drake, J.M. The decline and resurgence of pertussis in the US. Epidemics 2011, 3, 183. [Google Scholar] [CrossRef]
- Gill, C.; Rohani, P.; Thea, D.M. The relationship between mucosal immunity, nasopharyngeal carriage, asymptomatic transmission and the resurgence of Bordetella pertussis. F1000 Res. 2017, 6, 1568. [Google Scholar] [CrossRef]
- Ikematsu, H.; Kawai, N.; Yajima, S. A cross sectional survey measuring sero-incidence of pertussis infection among Japanese junior and senior high school students in 2013 and 2014. Vaccine 2017, 35, 3859–3864. [Google Scholar] [CrossRef]
- Pramono, R.X.; Imtiaz, S.A.; Rodriguez-Villegas, E. A Cough-Based Algorithm for Automatic Diagnosis of Pertussis. PLoS ONE 2016, 11, e0162128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ausiello, C.M.; Cassone, A. Acellular Pertussis Vaccines and Pertussis Resurgence: Revise or Replace? mBio 2014, 5, 01339-14. [Google Scholar] [CrossRef] [Green Version]
- Tinnion, O.N.; Hanlon, M. Acellular Vaccines for Preventing Whooping Cough in Children; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1999; p. CD001478. [Google Scholar]
- Warfel, J.M.; Zimmerman, L.I.; Merkel, T.J. Acellular pertussis vaccines protect against disease but fail to prevent infection and transmission in a nonhuman primate model. Proc. Natl. Acad. Sci. USA 2014, 111, 787–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Wei, C.; Mou, D.; Zuo, W.; Liang, J.; Ma, X.; Wang, L.; Gao, N.; Gu, Q.; Luo, P.; et al. Infant rhesus macaques as a non-human primate model of Bordetella pertussis infection. BMC Infect. Dis. 2021, 21, 407. [Google Scholar] [CrossRef]
- Mingbo, S.; Yan, M.; Yinhua, X.; Huijuan, Y.; Li, S.; Yanchun, C.; Guoyang, L.; Shude, J.; Shumin, Z.; Qihan, L. Dynamic profiles of neutralizing antibody responses elicited in rhesus monkeys immunized with a combined tetravalent DTaP-Sabin IPV candidate vaccine. Vaccine 2014, 32, 1100–1106. [Google Scholar]
- World Health Organization. Polio Laboratory Manual. Available online: https://apps.who.int/iris/handle/10665/68762 (accessed on 30 May 2020).
- Sato, Y.; Kimura, M.; Fukumi, H. Development of a pertussis component vaccine in Japan. Lancet 1984, 1, 122–126. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.C.; Ye, Q.; Li, C.; Liu, Y.P.; Ma, X.; Li, Y.; Zhao, H.; Chen, X.; Assudani, D.; et al. Safety, immunogenicity and persistence of immune response to the combined diphtheria, tetanus, acellular pertussis, poliovirus and Haemophilus influenzae type b conjugate vaccine (DTPa-IPV/Hib) administered in Chinese infants. Hum. Vaccines Immunother. 2017, 13, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zhang, H.; Zhou, W.; Ye, Q.; Li, F.; Wang, H.; Hou, Q.; Xu, Y.; Ma, X.; Tan, Y.; et al. Safety and immunogenicity of a diphtheria, tetanus, acellular pertussis and Haemophilus influenzae Type b combination vaccine compared with separate administration of licensed equivalent vaccines in Chinese infants and toddlers for primary and booster immunization. Vaccine 2010, 28, 4215–4223. [Google Scholar] [CrossRef]
- Cherry, J.D. Pertussis: Challenges today and for the future. PLoS Pathog. 2013, 9, e1003418. [Google Scholar] [CrossRef] [Green Version]
- Dewan, K.K.; Linz, B.; DeRocco, S.E.; Harvill, E.T. Acellular Pertussis Vaccine Components: Today and Tomorrow. Vaccines 2020, 8, 217. [Google Scholar] [CrossRef] [PubMed]
- Warfel, J.M.; Beren, J.; Kelly, V.K.; Lee, G.; Merkel, T.J. Nonhuman primate model of pertussis. Infect. Immun. 2012, 80, 1530–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saso, A.; Kampmann, B.; Roetynck, S. Vaccine-Induced Cellular Immunity against Bordetella pertussis: Harnessing Lessons from Animal and Human Studies to Improve Design and Testing of Novel Pertussis Vaccines. Vaccines 2021, 9, 877. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Component | Group1 (DTacP-sIPV Formulation1) | Group2 (DTacP-sIPV Formulation2) | Group3 (DTacP-wIPV/Hib) | Group4 (DTaP/Hib) | Group5 (wP) | Group6 (Negative Control) |
|---|---|---|---|---|---|---|
| DT (Lf/dose) | 25 | 12.5 | 15 | 12.5 | — | — |
| TT (Lf/dose) | 10 | 3.5 | 5 | 3.5 | — | — |
| PT (μg/dose) | 25 | 25 | 25 | 9 µg PN * | — | — |
| PRN (μg/dose) | 8 | 8 | — | — | — | |
| FHA (μg/dose) | 25 | 25 | 25 | — | — | |
| Whole cell pertussis (IU/dose) | — | — | — | — | 4 | — |
| IPV type I (DU/dose) | 30 (Sabin) | 15 (Sabin) | 40 (Mahoney) | — | — | — |
| IPV type II (DU/dose) | 32 (Sabin) | 16 (Sabin) | 8 (MEF-1) | — | — | — |
| IPV type III (DU/dose) | 45 (Pfizer) | 22.5 (Pfizer) | 32 (Saukett) | — | — | — |
| Aluminum (mg/dose) | 0.60–0.75 | 0.60–0.75 | 0.60–0.75 | 0.60–0.75 | — | 0.60–0.75 |
| Group 1 | Group 2 | Group 3 | Group 4 | Group 5 | Group 6 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose 1 | Dose 2 | Dose 3 | Dose 1 | Dose 2 | Dose 3 | Dose 1 | Dose 2 | Dose 3 | Dose 1 | Dose 2 | Dose 3 | Dose 1 | Dose 2 | Dose 3 | Dose 1 | Dose 2 | Dose 3 | |
| PT | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 0% | 0% | 60% | - | - | - |
| FHA | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 40% | 100% | 100% | - | - | - |
| PRN | 40% | 100% | 100% | 60% | 100% | 100% | - | - | - | 60% | 100% | 100% | 0% | 100% | 100% | - | - | - |
| DT | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | - | - | - | - | - | - |
| TT | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | - | - | - | - | - | - |
| Polio I | 100% | 100% | 100% | 100% | 100% | 100% | 75% | 100% | 100% | - | - | - | - | - | - | - | - | - |
| Polio II | 75% | 75% | 100% | 25% | 100% | 100% | 100% | 100% | 100% | - | - | - | - | - | - | - | - | - |
| Polio III | 0% | 75% | 100% | 0% | 75% | 100% | 25% | 75% | 100% | - | - | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Gao, N.; Wen, J.; Li, J.; Ma, Y.; Sun, M.; Liang, J.; Shi, L. Immunogenicity of a Candidate DTacP-sIPV Combined Vaccine and Its Protection Efficacy against Pertussis in a Rhesus Macaque Model. Vaccines 2022, 10, 47. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10010047
Wang X, Gao N, Wen J, Li J, Ma Y, Sun M, Liang J, Shi L. Immunogenicity of a Candidate DTacP-sIPV Combined Vaccine and Its Protection Efficacy against Pertussis in a Rhesus Macaque Model. Vaccines. 2022; 10(1):47. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10010047
Chicago/Turabian StyleWang, Xiaoyu, Na Gao, Jiana Wen, Jingyan Li, Yan Ma, Mingbo Sun, Jiangli Liang, and Li Shi. 2022. "Immunogenicity of a Candidate DTacP-sIPV Combined Vaccine and Its Protection Efficacy against Pertussis in a Rhesus Macaque Model" Vaccines 10, no. 1: 47. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10010047