
Immune Escape Mechanism and Vaccine Research Progress of African Swine Fever Virus
by
 ,
,
Zhaoyang Wang
1,2,3,4 ,
,
Qiangyun Ai
1,2,3,4,
Shenglin Huang
1,2,3,4,
Yating Ou
1,2,3,4,
Yinze Gao
1,2,3,4,
Tiezhu Tong
5,* and
Huiying Fan
1,2,3,4,* 1
College of Veterinary Medicine, South China Agricultural University, Guangzhou 510642, China
2
National and Regional Joint Engineering Laboratory for Medicament of Zoonosis Prevention and Control, Guangzhou 510642, China
3
Key Laboratory of Animal Vaccine Development, Ministry of Agriculture, Guangzhou 510642, China
4
Key Laboratory of Zoonosis Prevention and Control of Guangdong Province, Guangzhou 510642, China
5
Guangzhou Customs Technology Center, Guangzhou 510623, China
*
Authors to whom correspondence should be addressed.
Vaccines 2022, 10(3), 344; https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10030344
Submission received: 19 January 2022
/
Revised: 16 February 2022
/
Accepted: 17 February 2022
/
Published: 22 February 2022
(This article belongs to the Special Issue African Swine Fever Immunity and Vaccines)
Abstract
:African swine fever virus (ASFV) is the causative agent of the epidemic of African swine fever (ASF), with virulent strains having a mortality rate of up to 100% and presenting devastating impacts on animal farming. Since ASF was first reported in China in 2018, ASFV still exists and poses a potential threat to the current pig industry. Low-virulence and genotype I strains of ASFV have been reported in China, and the prevention and control of ASF is more complicated. Insufficient understanding of the interaction of ASFV with the host immune system hinders vaccine development. Physical barriers, nonspecific immune response and acquired immunity are the three barriers of the host against infection. To escape the innate immune response, ASFV invades monocytes/macrophages and dendritic cells, thereby inhibiting IFN expression, regulating cytokine expression and the body’s inflammatory response process. Meanwhile, in order to evade the adaptive immune response, ASFV inhibits antigen presentation, induces the production of non-neutralizing antibodies, and inhibits apoptosis. Recently, significant advances have been achieved in vaccine development around the world. Live attenuated vaccines (LAVs) based on artificially deleting specific virulence genes can achieve 100% homologous protection and partial heterologous protection. The key of subunit vaccines is identifying the combination of antigens that can effectively provide protection and selecting carriers that can effectively deliver the antigens. In this review, we introduce the epidemic trend of ASF and the impact on the pig industry, analyze the interaction mechanism between ASFV and the body’s immune system, and compare the current status of potential vaccines in order to provide a reference for the development of effective ASF vaccines.
1. Introduction
In 1921, ASFV was first reported in Kenya, which caused about an 100% mortality in infected domestic pigs [1]. Subsequently, ASFV spread to most African countries south of the Sahara [2]. In 1957 and 1960, the African swine fever epidemic broke out in Portugal which was the first time to demonstrate that ASFV spread across continents and spread to other European countries, the Caribbean, and Brazil [3]. Except in Sardinia, the genotype I ASFV was eradicated in the 1990’s [4]. However, the genotype II ASFV was introduced to Georgia in the Caucasus region and started a new round of transmission in 2007 [5]. Afterwards, it spread to the Russian Federation, Ukraine and Belarus, and in 2014 it spread to Eastern European countries [4,5]. In 2018, the epidemic spread to Belgium, Hungary, Czech Republic, Romania, Bulgaria, Slovakia, and Serbia, as well as China and other parts of Asia (Mongolia, Korea, Vietnam, Laos, Cambodia, Myanmar, the Philippines, Hong Kong, and Indonesia) [3]. ASF outbreaks are still trending regionally, with new regional reports of ASF outbreaks. In September 2019, Timor-Leste reported the first outbreak of African swine fever in Oceania, followed by Papua New Guinea (March 2020) [6,7]. In July 2021, ASF reappeared in the Americas, first in the Dominican Republic and later in Haiti [8,9]. In January 2022, ASF genotype II was notified in mainland Italy [9]. In January 2022, two new countries also reported their first cases of the disease: North Macedonia in Europe and Thailand in Asia [9]. According to OIE, from January 2020 to January 2022, ASF outbreaks were reported in 35 countries or regions around the world, including 4767 cases (1043334 animals lost) in domestic pigs and 18,262 cases (29970 animals lost) in wild boars [9]. It is worth noting that wild boar cases in Europe accounted for the vast majority (83.3%, 16743/20107), while wild boar cases in Asia accounted for a lower proportion of 59.4% (1519/2559) [9]. The wild boar reservoir of ASFV will present challenges to the eradication of ASF by vaccination programs.
In August 2018, the first ASF case in China was identified in Shenyang, Liaoning Province [10]. Since most pig farms in China are small-scale pig farms with very low biosecurity levels, the rapid spread of ASFV across the country has led to a serious reduction in the number of live pigs and caused heavy losses to the pig industry [11]. The decline in the number of live pigs has led to a rapid rise in the price of pork in a short period of time, affecting the normal life of residents in China. After the outbreak of ASF, with the joint efforts of Chinese government leaders and industry practitioners, various pig farms tried to resume production. By improving the level of biosecurity, including optimizing the location of pig farms, strict disinfection measures, and strengthening ASFV testing on pig farms, ASFV-positive pigs were culled at designated locations, and the source of the infection was eliminated. Most pig farms finally successfully resumed breeding [12]. However, in order to improve the level of biosecurity, the measures taken by pig farms have objectively increased the cost of breeding. Therefore, there is an urgent need for vaccines for the effective control and eradication of ASF.
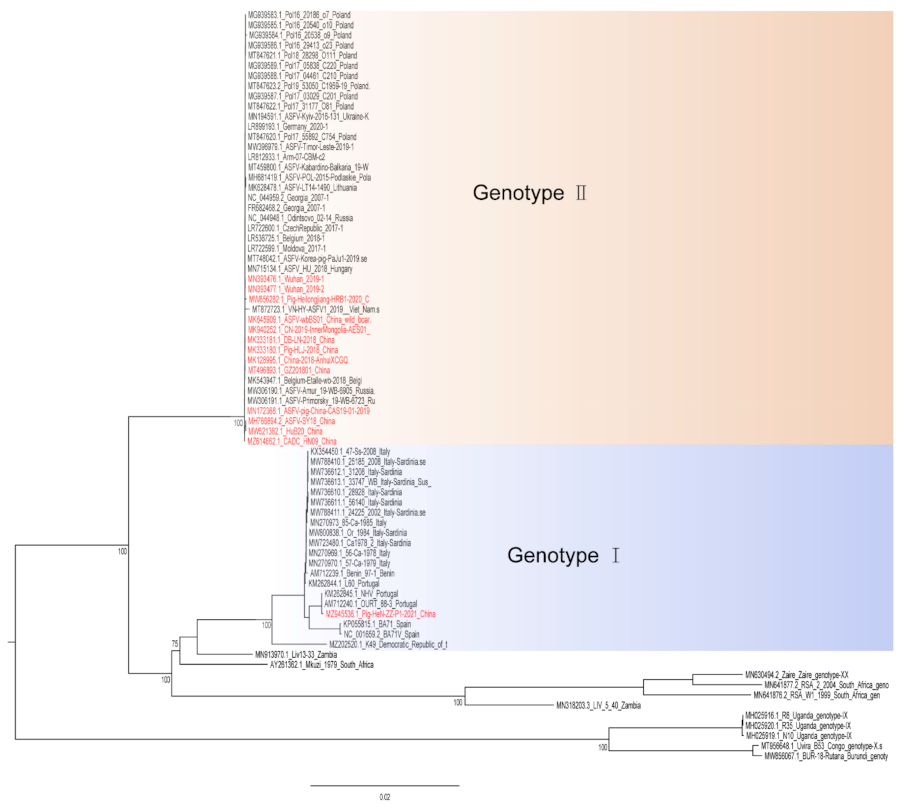
Hemadsorption (HAD) refers to the ability of ASFV-infected macrophages to adsorb erythrocytes, and non-hemadsorption (non-HAD) refers to the inability of ASFV-infected macrophages to adsorb erythrocytes. ASFV has spread for more than three years in China, and its epidemic trend has become more complicated with the emergence of non-hemadsorbing (non-HAD) naturally attenuated strains of ASFV [13,14,15]. The initial clinical symptoms of domestic pigs infected with low-viral strains are not obvious, which is difficult to diagnose [13,14]. Comparative analysis showed that the genome of the low-virulence strains had mutations, deletions, insertions or substitutions of short fragments, including the deletion or mutation of the CD2v (EP402R) gene, which is related to the HAD phenotype [16]. In addition, genotype I strains have also been reported in China (Figure 1) [14]. The potential for endemic ASF outbreaks is increasing due to the widespread distribution of wild boars in China. Researchers are actively conducting ASF vaccine-related research, including the evaluation of live attenuated vaccines (LAVs) vaccines by knocking out virulence genes, and the screening of antigens with protective effects to develop subunit vaccines, vector vaccines, etc. [17,18,19,20,21,22,23,24,25,26,27,28,29,30]. ASFV has evolved the ability to manipulate host immune responses by encoding many immune escape genes, including regulation of IFN expression, inhibition of autophagy, and apoptosis. In addition, ASFV affects the antigen presentation and activation of lymphocytes by invading monocytes-macrophages and DCs, thereby affecting the immune system of the entire body [31]. This review summarizes the escape mechanism of ASFV from the immune response and the progress of ASF vaccines, hoping to provide help for future ASF vaccines research.
2. The Major Defense Mechanisms and ASFV Interactions
2.1. Physical Barriers
The skin barrier is the most effective physical defense of the host to block entry of pathogens. The physical barriers of the body’s defenses include intact skin, surfaces of the respiratory and gastrointestinal tracts and include processes of self-cleaning (coughing, sneezing and mucus flow in the respiratory tract, vomiting, diarrhea, and urination of the urinary system, etc.), and normal flora on the skin surface and in the gut. The transmission of ASFV among pigs is mainly direct contact transmission, including transmission through mouth-nose contact and short-distance aerosol transmission [34]. In addition, the presence of soft tick bites in pig farms can also cause the transmission of ASFV [35]. Therefore, it is necessary to improve animal welfare in the farm, and increase the resistance of animals by reducing the breeding density. Pig farms should strengthen environmental sanitation and disinfection to block the spread of the virus. On the contrary, for infected animals, the self-cleaning of the body is also conducive to virus spread. It is necessary to regularly collect and detect the presence of ASFV in pig herds, and to ensure the removal and safe destruction of positive pig herds to eliminate the source of infection [12].
2.2. Innate Immunity
Sentinel cells, such as macrophages, dendritic cells, and mast cells, elicit inflammatory responses by recognizing pathogen-associated molecular patterns (PAMPs) of microorganisms through pattern recognition receptors. The inflammatory response involves the activation and directed migration of various cells from the blood to the site of invasion, especially neutrophils and macrophages. The pattern recognition receptors that recognize ASFV mainly include TLR3 that recognizes viral dsRNA and cGAS-STING that recognizes viral DNA in the cytoplasm [36,37,38]. When TLR3 binds to dsRNA, it transmits signals to cells, resulting in increased expression of nuclear factor kappa-B (NF-κB), which in turn activates IL-1, IL-6 and TNF-α and other cytokine expression [36]. The cGAS-STING system recognizes virus DNA, and the virus-infected (mainly plasmacytoid dendritic cells) cells produce IFN-I [38]. IFN-I acts on virus-infected cells to inhibit virus growth.
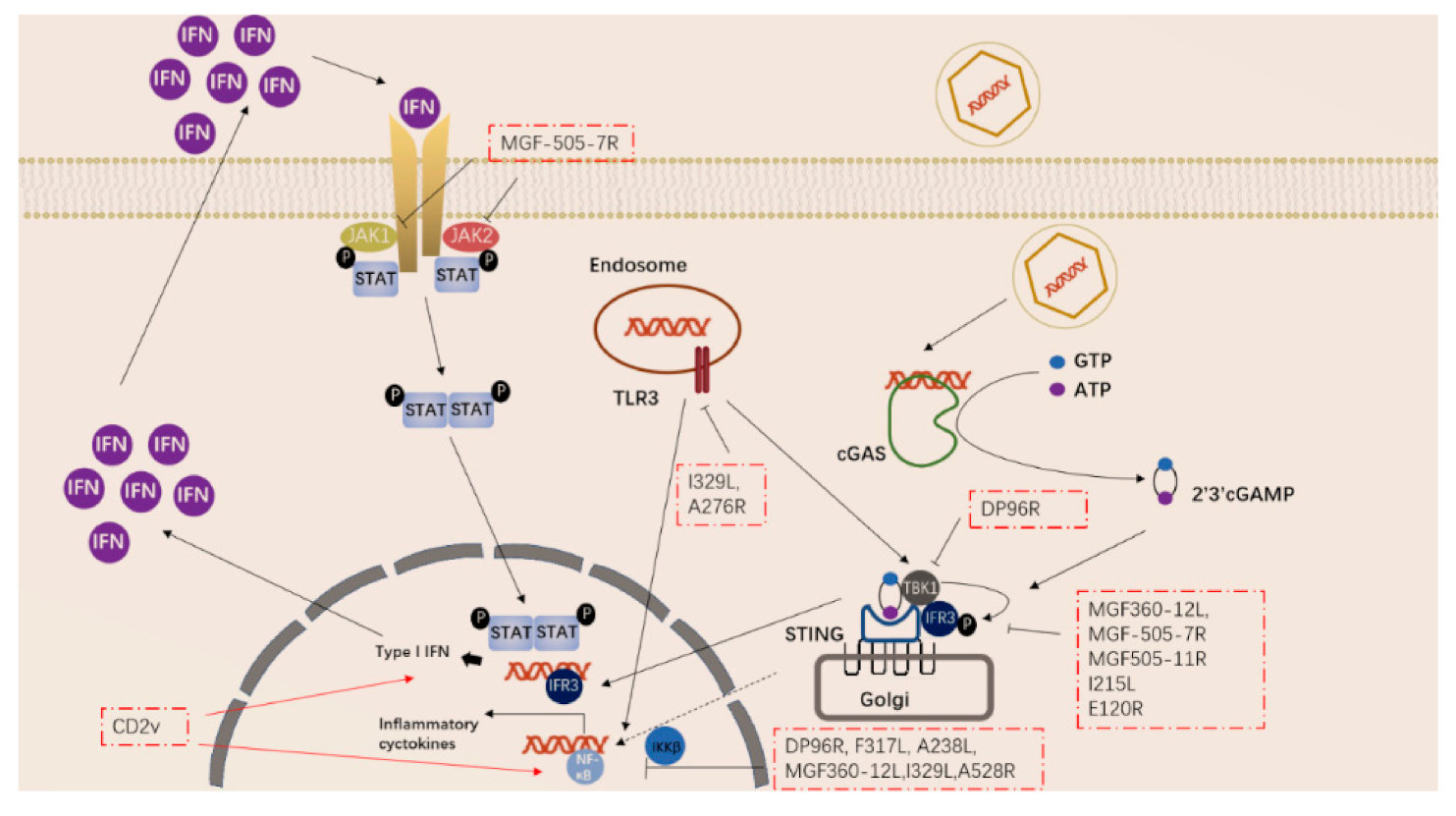
ASFV has also acquired many genes in evolution that can regulate the inflammatory response in the early stage of the body. ASFV-encoded genes also primarily inhibit the actions of TLR3 and the cGAS-STING system (Figure 2). Studies have shown that the I329L and A276R proteins encoded by ASFV have immunosuppressive effects [39,40]. I329L (the homologous protein of TLR3) interacts with the adaptor protein TRIF to inhibit the activation of NF-κB and IRF3, while A276R can also inhibit the recognition of TLR3 [37,39]. In addition, another study showed that TLR expression was decreased after ASFV infection of macrophages [40].
ASFV encodes multiple genes involved in suppressing the cGAS-STING signaling pathway. In 2013, Correia et al. reported that A528R (MGF530) could inhibit the expression of IFN-β by targeting IRF3 and NF-κB or through the JAK-STAT pathway [39,41]. In 2018, Wang et al. found that DP96R targets TBK1 and IκB kinase beta (IKKβ) to negatively regulate IFN-I expression and induction of NF-κB signaling [42]. In 2020, Zhuo et al. demonstrated that MGF360-12L can block the interaction between p65 and KPNA2, KPNA3, and KPNA4, and can interfere with the nuclear translocation of NF-κB [43]. Five immunosuppressive genes were reported in 2021, including E120R, F317L, MGF505-7R, MGF505-11R, and I215L [44,45,46,47,48,49]. E120R can interact and block the activation of IRF3, thereby inhibiting the expression of IFN-β [44]. F317L inhibits IKKβ phosphorylation, thereby reducing the phosphorylation and ubiquitination of IkBα, thereby inhibiting the activation of the NF-κB pathway [45]. Li et al. reported that MGF-505-7R promoted the expression of autophagy-related protein ULK1 to degrade STING, and inhibited IFN-γ-mediated JAK1 and JAK2-mediated signaling pathways [46,48]. Similarly, Yang et al. reported that MGF505-11R interacts with STING, degrades STING expression through lysosome, ubiquitin-proteasome and autophagy pathways, and can inhibit the phosphorylation of TBK1 and IRF3 stimulated by cGAS/STING overexpression [47]. pI215L recruits RNF138 to inhibit K63-related TBK1 ubiquitination, which has an inhibitory effect on IFN-I production [49]. It is worth noting that CD2V can activate NF-κB and induce activation of IFN signaling pathways in porcine lymphocytes/macrophages [50]. These immunosuppressive genes are potential target genes for the preparation of live attenuated vaccines.
2.3. Acquired Immunity
2.3.1. Antigen Processing
Dendritic cells and macrophages are both sentinel cells of innate immunity and efficient antigen processing cells. DCs are the only antigen-presenting cells capable of activating T cells that have never been exposed to an antigen (naive T cells). ASFV invasion of DCs may seriously affect their antigen processing and delivery, resulting in immunosuppressive responses [51]. Antigen processing breaks down antigens into small polypeptides, which are subsequently attached to professional antigen receptor-major histocompatibility complex molecules (MHCs).
Exogenous antigen processing begins with phagocytosis of the antigen to form endosomes, which are gradually acidified during intracellular migration and maturation, followed by fusion with protease-containing lysosomes [52,53]. The ingested peptides are degraded into fragments of different lengths by proteases, and some peptide fragments are combined with MHC-II molecules and presented to CD4+ T cells. The invasion process of ASFV utilizes the exogenous processing pathway of the cell. After acidification of the endosome, ASFV sheds its capsid, fuses its inner capsule with the restrictive endosomal membrane, releases its core capsid into the cytoplasm, and then releases its DNA [54].
Ubiquitin molecules bind to lysine of endogenous abnormal proteins. Ubiquitin chains are recognized by the proteasome and cleaved into fragments of 8 and 15 amino acids in length. In addition, the activity of the proteasome is regulated by cytokines such as IFN-γ and caspases. In fact, ubiquitination modifications have been used in ASFV vaccine research. A DNA vaccine fused to the extracellular domain (sHA) of ASFV p30, p54, and CD2v induced potent humoral and specific T cell responses in pigs, but was not protective [55]. In addition, ubiquitination of the above three genes may provide partial protection by improving the presentation of major histocompatibility complex (MHC) class I antigens [55]. This property of ubiquitination could be applied to the design of future ASF vaccines.
2.3.2. Antigen Presentation
The antigen presentation process includes endogenous presentation pathway (MHC-I pathway), exogenous presentation pathway (MHC-II pathway) and cross antigen presentation pathway. MHC-I is present in all nucleated cells and presents endogenous antigens to CD8+ T cells. MHC-II exists only in professional antigen-presenting cells, dendritic cells, macrophages and B cells, and its function is to present exogenous antigens to CD4+ T cells.
Exogenous molecules elicit adaptive immunity by binding to the antigen-binding groove of at least one MHC molecule. MHC-class Ia and class II molecules are polymorphic, and one MHC molecule is estimated to bind to about 2500 different polypeptides. Therefore, MHC alleles can determine the susceptibility of the body to infectious diseases based on the adaptive immune response to the infecting pathogen. Screening for specific MHC haplotypes can be used to develop ASFV-resistant domestic swine lines. In March 2020, a Chinese research team reported that ASFV-resistant domestic pigs LS-2 were significantly resistant to oral infection with the 106.0 TCID50 ASFV SY18 strain. There were significant improvements in the rate of infection, viremia, clinical symptoms, and antibody response after challenge, and there were also significant differences in the expression of inflammatory factors, which is of great significance for the prevention and control of ASFV [56]. However, MHC polymorphism is an evolutionary conserved feature making it difficult for pathogens to evade from immune recognition; if one resistant MHC haplotype is clonally expanded, this may bear risks for the other epizootic pathogens to come.
EP153R of ASFV can affect the trafficking of MHC-I from the endoplasmic reticulum to the cell membrane [51]. In addition, ASFV infection of cells resulted in increased expression of SLA-DOA and SLA-DOB, which inhibited the binding of antigenic peptides to MHC-II [40]. However, ASFV can inhibit the expression of MHC-I, NK cells will not receive inhibitory signals, and cells infected with ASFV may be killed [57]. Similar to T cells, NK cells induce apoptosis by activating intracellular caspases cascades through both exogenous and endogenous pathways. Interestingly, Franzoni et al. showed that attenuated or avirulent strains, but not virulent strains, were able to downregulate the expression of MHC-I molecules on MoDCs [58]. Similar results were observed in experiments infected with un-activated (moMΦ) and alternatively (moM2) activated monocyte-derived macrophages [59]. These studies of virulence ASFV have evolved mechanisms for covert replication, which may escape the killing effect of NK cells [58,59].
2.3.3. Antibody Induction
Activation of B cells requires the stimulation of multiple signals. Antigen binding to the BCR is a necessary first step in triggering a B cell response, but is often insufficient to trigger antibody formation. Activation of B cells requires co-stimulation of helper T cells and cytokines. However, for helper T cells to function, they need to first present antigens to the helper T cells before they can provide costimulatory signals. The antigen can be captured and processed by DCs, macrophages and B cells and presented to T cells. The target cells of ASFV are mainly macrophages and DCs, and their antigen-presenting ability may be reduced after a virus infection [60,61,62]. The body may activate B cells through other pathways. After capturing the antigen, B cells process the antigen and present it to helper T cells. The antigen-presenting ability of B cells is much smaller in comparison with DC cells.
Responding B cells can differentiate into plasma cells, which secrete large amounts of antibodies. The main functions of antibodies include blocking the adsorption of viruses to target cells, stimulating the phagocytosis of macrophages, and stimulating complement-mediated viral decomposition. However, antibodies elicited by ASFV infection cannot completely neutralize ASFV, and about 4–13% of the virus cannot be neutralized [63,64]. An experiment revealed that the incomplete neutralization of ASFV may be caused by virus aggregation [64]. In addition, non-neutralizing antibodies in the ASFV infection may result in Antibody-Dependent Enhancement (ADE) [65]. Therefore, ASFV vaccines need to eliminate those antigens or epitopes that induce non-neutralizing antibodies.
2.3.4. Cellular Immune
Cell-mediated immunity clears abnormal cells and intracellular pathogens. Endogenous antigens are first cleaved into small polypeptides and then inserted into the antigen-binding groove of the major histocompatibility complex (MHC) class I. When presented to the cell surface, the antigen binds to the antigen receptor of T cells. After CD8+ T cells are fully activated, they leave the lymphoid organs and search for infected cells by themselves. When activated CD8+ T cells recognize MHC-antigen complexes expressed on another cell, and cytotoxic T cells force the apoptosis of virus-infected target cells. In addition, NK cells fight against viral infection by directly killing virus-infected cells, inducing antibody-dependent cell-mediated cytotoxicity (ADCC), or secreting cytokines (such as IFN and TNF-α). A study has shown that asymptomatic pig herds infected with NH/P68 strain significantly enhanced NK cell cytotoxicity and resisted virulent ASFV Lisbon 60 challenge [66]. The genes currently known to inhibit apoptosis are A224L, EP153R, A179L, and DP71L, mainly by inhibiting the death receptor pathway, the mitochondrial apoptotic pathway and protein kinase RNA-like ER kinase (PERK) pathways (endoplasmic reticulum stress response) [67,68,69,70]. Granzyme B is one of the main effector molecules, and can directly act on caspase-3 to induce apoptosis. ASFV-encoded EP153R and A224L can inhibit the activity of caspase-3 [67,68]. However, granzyme B can trigger apoptosis through ICAD/CAD [71]. In addition, it has been reported that ASFV can inhibit the expression of cytotoxic T cells perforin at 5 dpi [72]. ASFV may partially inhibit perforin/granzyme-induced apoptosis. Furthermore, when the host cell senses abnormal signals, eIF2 is phosphorylated, which shuts down the protein synthesis system. DP71L of ASFV promotes the replication of the virus in cells by recruiting PP1 to dephosphorylate eIF2 [70]. When ASFV replication is completed, the mitochondrial apoptosis pathway is activated through the structural protein p54 encoded by E183L, which promotes cell lysis to start a new round of infection and replication [73]. ASFV encodes related proteins to regulate apoptosis and promote the replication, proliferation and spread of ASFV, which is also one of the strategies to achieve immune escape.
In conclusion, effective ASF vaccines can induce the production of neutralizing antibodies that block the entry of ASFV into target cells or induce ADCC. Therefore, screening for target epitopes of neutralizing antibodies is required. In addition, vaccination induces the body to produce Cytotoxic T lymphocyte cells (CTLs) that can specifically recognize abnormal cells. Similarly, it is also necessary to screen for antigenic epitopes that can induce specific cellular immunity. In fact, this is also one of the current research directions [74,75,76,77,78]. However, whether the effective combination of antigenic epitopes is effective still needs much experimental verification.
3. ASF Vaccines
3.1. Live Attenuated Vaccines (LAVs)
LAVs can generally be divided into natural attenuated strains and artificial gene deletion strains, and the latter is made by deleting certain virulence genes. Compared with other types of ASF vaccines, LAVs can provide complete homologous and partial heterologous protection [79]. The most promising LAVs vaccine candidates are summarized in Table 1. Chen et al. reported that a seven-gene deletion LAV had a good protective effect [17]. In addition, Zhang et al. reported that deleting L7L-L11L attenuates ASFV. In vaccination experiments, these attenuated strains conferred 100% protection against homologous challenge [18]. LAVs have experienced a trend from multiple gene deletions to single gene deletions. Protection induced by immunization with LAVs correlates with the level of replication of the LAVs in addition to the number of immunogenic genes expressed. If the replication of LAVs is severely attenuated, then the LAVs will not be immunogenic. LAVs with single gene deletions express more genes and may cause better protection. ASFV-G-∆I177L could achieve 100% protection through oral and injection routes, and field trials have been completed in Vietnam [19,20,21]. Similarly, ASFV-G-ΔA137R and SY18ΔI226R were proven to confer 100% protection [22,23].
Historically, attenuated strains were once used in Spain and Portugal, but widely caused chronic ASF infections in vaccinated pigs. Therefore, adequate experiments should be conducted to verify the effectiveness and safety of LAVs before extensive promotion, for example, future ASF vaccines should be distinguish between infected animals and vaccinated animals (DIVA) [80]. Ramirez-Medina et al. constructed a deletion strain of E184L to distinguish between infected animals and vaccinated animals (DIVA), but the deletion strain cannot provide complete protection [80]. LAVs are still far from commercialization due to the safety issue. LAVs may have the risk of virulence reversion during large-scale vaccination. Therefore, adequate experiments should be conducted to verify the effectiveness and safety of LAVs before extensive promotion.
In addition, the lack of cell lines for large-scale LAVs production is another difficulty. ASFV subculture can adapt to cell lines such as 293 and Vero, but the virulence and antigenicity of the adapted strains are weakened, and adapted strains cannot provide effective protection after immunizing pigs [81,82]. The adapted cells cultured in LAVs are primary cells, mainly including porcine alveolar macrophages (PAM) and porcine bone marrow cells (PBMS). Primary cells are not sufficient for large-scale production of LAVs. Borca et al. reported that ASFV-G-ΔI177L/ΔLVR could replicate efficiently in stable porcine cell lines with potent protection [83]. In addition, Takenouchi et al. reported a novel immortalized porcine macrophage cell line, IPKM, which was generated by transduction of primary PKM with lentiviral vectors encoding SV40LT and pTERT [84]. However, it is necessary to carefully test it before using IPKM as a production cell line for a live-attenuated vaccine strain.
Finally, the cross-protective ability of LAVs needs to be further evaluated. ASFV has mutations, duplications, and loss of certain sequences in the genomes of different genotypes or even different strains of the same genotype, which may lead to changes in virulence [85].
3.2. Subunit Vaccines
Subunit vaccines are generally in the laboratory research stage, and there is still a huge gap between practical applications. It has been confirmed that some ASFV antigens have a certain protective effect [24,26]. Antibodies against p72 and p54 can block the adsorption of ASFV, and antibodies against p30 can block the internalization of the virus [87]. The key to subunit vaccines is to screen for those antigens or those epitopes of antigens that have definite protective effects [88]. In the presence of non-neutralizing antibodies, the virus-antibody complex formed is infectious. The virus bound by the non-neutralizing antibody is phagocytosed by macrophages, and the virus can still grow in the macrophage at this time, and the virus bound by the non-neutralizing antibody accelerates the replication of the virus [89,90].
Previous studies have noted a critical role for CD8+ T cells in protection, as demonstrated by using the virulent Portuguese ASFV isolate OUR/T88/1 [91]. DNA vaccines as well as vector vaccines have been attempted as alternative ASF vaccine platforms. In theory, DNA vaccines as well as vectors have better immunogenicity because the antigen can be expressed intracellularly and via MHC I, which is important for CD8+ T cell activation. Neither DNA vaccine, vector vaccine, nor prime-boost immunization strategies can provide complete protection [56,92,93,94]. Luckily, Goatley et al. cloned B602L, p72, p30, p54, E199L, EP153R, F317L, and MGF505-5R into adenovirus vectors for primary immunization, and into MVA vectors for booster immunization, then found the protective function up to 100% in pigs [30]. However, low levels of infectious virus remained in the immunized pigs until the end of the trial [30]. Future experiments should determine whether pigs can clear the infection and whether these vaccinated animals shed infectious virus [30].
Similar to screening for antigens for neutralizing antibodies, screening is also required to identify antigenic epitopes capable of inducing ASFV-specific T cells. Bosch-Camós used in silico prediction of ASFV protein CD8+ T cell epitopes and predicted a 19-mer peptide from MGF100-1L [76]. Netherton et al. predicted peptides corresponding to 133 proteins encoded by OUR T88/3, and screened 18 viral proteins that could be recognized by lymphocytes of ASF-immunized pigs [78]. The specific antigenic epitopes are determined by software analysis and verification in combination with relevant experiments, which guides the selection of antigens in the development of ASF vaccines.
Recently, some research teams have conducted large-scale screening of subunit vaccine protective antigens and identified a group of antigens that can effectively block virus replication in pigs and protect cohabiting pigs from infection [95,96]. Part of the data showed that CD2V protein and p72 protein containing trimers make subunit vaccine compositions that can provide complete and safe protection to pigs [95].
3.3. Other Types of Vaccines
Inactivated vaccine is a safe and low-cost vaccine that is used in the prevention of many diseases [97,98,99]. However, the ASF inactivated vaccine does not provide protection [100,101]. Co-immunization of inactivated vaccines and adjuvants that stimulate cellular immunity also failed to provide protection [102].
The ASF outbreaks from 2016 to 2020 are dominated by wild boar populations in Europe, and wild boar infections have also been reported in Asia [103]. ASFV spreads in wild boar herds and may spread to domestic pigs. The use of oral vaccines in ASFV endemic areas may be a feasible measure to prevent ASFV in wild boars. Previous studies have demonstrated that oral live-attenuated vaccines could achieve effective protection in in the laboratory stage [20,104].
Critical progress in mRNA vaccines technology has occurred over the past two decades [105,106], which have recently presented distinguished effectiveness against COVID-19 [107,108]. mRNA vaccines are divided into non-replicating and self-amplifying (Alpha virus replicons). The mRNA expression of exogenous protein in the cytoplasm does not need to enter the nucleus, so it can be expressed in non-dividing cells or slowly dividing cells (such as dendritic cells). Compared with DNA vaccine, mRNA vaccines have higher safety and stronger immunogenic response in humans and large animals [109,110,111]. And for viral vector vaccines, mRNA vaccines have superior safety and applicability, and will not affect the effect of boosting immunity [112,113]. However, mRNA vaccines are also expensive to manufacture and transport, and there may be potential unknown risks. Compared to mRNA vaccines, self-amplifying RNA-replicons represent a cost-effective alternative that could become an interesting approach also in veterinary medicine.
4. Conclusions
ASFV has a complex five-layer structure and encodes 150 to 200 proteins, including many unknown functional proteins. Low-virulence and genotype I strains of ASFV have been reported in China, and the prevention and control of ASF is more complicated. The major defense mechanisms of animals have three lines of defense against the invasion of microorganisms. The first line of defense is a simple physical barrier that can resist most pathogens. The second line of defense is the non-specific immune response, which mainly includes neutrophils, macrophages, phagocytosing foreign bodies, and secreting cytokines, causing an inflammatory response and also initiating an adaptive immune response. The third line of defense is the acquired immune line of defense, including antigen capture and processing, antigen presentation, activation of B lymphocytes, and secretion of specific antibodies. At the same time, T lymphocytes are activated to kill abnormal cells. However, the target cells of ASFV are macrophages and DCs, and they encode a variety of proteins that inhibit the function of IFN. The inhibition of ASFV on the body’s immunity is reflected in various aspects, including the regulation of inflammatory response, the inhibition of antigen presentation, and the inhibition of apoptosis. Therefore, the development of ASF vaccine has a long way to go. It is necessary to fully understand the interaction between ASFV and the body’s immunity, and to find the key genes that can play a protective role. Currently, different vaccines have been tried against ASF. ASF LAVs can provide 100% protection against homologous strains, but there are still potential safety concerns. ASF vaccine needs more relevant basic research. In addition, subunit vaccines are also one of the current research priorities. In conclusion, screening ASF immunogenic proteins, identifying immune-related genes, and understanding the protective mechanism of ASF vaccines will facilitate the development of new ASF vaccines.
Author Contributions
Manuscript writing, Z.W. and S.H.; review and editing, Q.A., Y.O., Y.G. and H.F.; conception and design of manuscript, Z.W., Q.A., T.T. and H.F. All authors contributed to the article and approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Key-Area Research and Development Program of Guangdong Province (2019B020211004) and National Natural Sciences Foundation of China (No.31941013).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge Jiahao Zhang for technical support of the sequence analysis in this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulumba-Mfumu, L.K.; Saegerman, C.; Dixon, L.K.; Madimba, K.C.; Kazadi, E.; Mukalakata, N.T.; Oura, C.A.L.; Chenais, E.; Masembe, C.; Stahl, K.; et al. African swine fever: Update on Eastern, Central and Southern Africa. Transbound. Emerg. Dis. 2019, 66, 1462–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African Swine Fever Epidemiology and Control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, E.G.; Perez-Nunez, D.; Revilla, Y. Development of vaccines against African swine fever virus. Virus Res. 2019, 265, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Niemi, J.K. Impacts of African Swine Fever on Pigmeat Markets in Europe. Front Vet. Sci 2020, 7, 634. [Google Scholar] [CrossRef] [PubMed]
- Mileto, P.; da Conceicao, F.; Stevens, V.; Cummins, D.; Certoma, A.; Neave, M.J.; Bendita da Costa Jong, J.; Williams, D.T. Complete Genome Sequence of African Swine Fever Virus Isolated from a Domestic Pig in Timor-Leste, 2019. Microbiol. Resour. Announc. 2021, 10, e0026321. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Truong, A.D.; Dang, A.K.; Ly, D.V.; Nguyen, C.T.; Chu, N.T.; Nguyen, H.T.; Dang, H.V. Genetic characterization of African swine fever viruses circulating in North Central region of Vietnam. Transbound. Emerg. Dis. 2021, 68, 1697–1699. [Google Scholar] [CrossRef]
- Goonewardene, K.B.; Onyilagha, C.; Goolia, M.; Le, V.P.; Blome, S.; Ambagala, A. Superficial Inguinal Lymph Nodes for Screening Dead Pigs for African Swine Fever. Viruses 2022, 14, 83. [Google Scholar] [CrossRef]
- African Swine Fever (ASF)—Situation Report 4. Available online: https://www.oie.int/app/uploads/2022/01/asf-situation-report-4.pdf (accessed on 18 January 2022).
- Zhao, D.; Liu, R.; Zhang, X.; Li, F.; Wang, J.; Zhang, J.; Liu, X.; Wang, L.; Zhang, J.; Wu, X.; et al. Replication and virulence in pigs of the first African swine fever virus isolated in China. Emerg. Microbes Infect. 2019, 8, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Liu, J.; Wang, L.; Fan, S.; Li, Z.; Li, Y.; Yi, L.; Ding, H.; Zhao, M.; Chen, J. Current State of Global African Swine Fever Vaccine Development under the Prevalence and Transmission of ASF in China. Vaccines 2020, 8, 531. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Qi, W.; Yang, Y.; Liu, Z.; An, T.; Wu, X.; Chen, J. Prevention and Control Strategies of African Swine Fever and Progress on Pig Farm Repopulation in China. Viruses 2021, 13, 2552. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Zhang, Z.; Wang, Z.; He, X.; Zhang, X.; Wang, L.; Wang, W.; Huang, L.; Xi, F.; Huangfu, H.; et al. Emergence and prevalence of naturally occurring lower virulent African swine fever viruses in domestic pigs in China in 2020. Sci. China Life Sci. 2021, 64, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Huang, L.; Zhang, X.; Zhang, J.; Shen, D.; Zhang, Z.; Wang, Z.; Huo, H.; Wang, W.; Huangfu, H.; et al. Genotype I African swine fever viruses emerged in domestic pigs in China and caused chronic infection. Emerg. Microbes Infect. 2021, 10, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Kutish, G.F.; Afonso, C.L.; Irusta, P.; Carrillo, C.; Brun, A.; Sussman, M.; Rock, D.L. An African swine fever virus gene with similarity to the T-lymphocyte surface antigen CD2 mediates hemadsorption. Virology 1994, 199, 463–468. [Google Scholar] [CrossRef]
- Yáñez, R.J.; Rodríguez, J.M.; Nogal, M.L.; Yuste, L.; Enríquez, C.; Rodriguez, J.F.; Viñuela, E. Analysis of the complete nucleotide sequence of African swine fever virus. Virology 1995, 208, 249–278. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhao, D.; He, X.; Liu, R.; Wang, Z.; Zhang, X.; Li, F.; Shan, D.; Chen, H.; Zhang, J.; et al. A seven-gene-deleted African swine fever virus is safe and effective as a live attenuated vaccine in pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Chen, T.; Yang, J.; Yue, H.; Wang, L.; Zhou, X.; Qi, Y.; Han, X.; Ke, J.; et al. Deletion of the L7L-L11L Genes Attenuates ASFV and Induces Protection against Homologous Challenge. Viruses 2021, 13, 255. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017. [Google Scholar] [CrossRef]
- Borca, M.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Gay, C.; Gladue, D. ASFV-G-∆I177L as an Effective Oral Nasal Vaccine against the Eurasia Strain of Africa Swine Fever. Viruses 2021, 13, 765. [Google Scholar] [CrossRef]
- Hanh, T.; Phuong, L.; Huy, N.; Thuy, D.; Van Dung, N.; Gay, C.; Borca, M.; Gladue, D. African swine fever virus vaccine candidate ASFV-G-DI177L efficiently protects European and native pig breeds against circulating Vietnamese field strain. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. ASFV bearing an I226R gene-deletion elicits a robust immunity in pigs to African swine fever. J. Virol. 2021, 95, e0119921. [Google Scholar] [CrossRef]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of A137R gene from the pandemic strain of African swine fever virus is attenuated and offers protection against virulent pandemic virus. J. Virol. 2021, 95, e0113921. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Puertas, P.; Rodríguez, F.; Oviedo, J.; Brun, A.; Alonso, C.; Escribano, J. The African swine fever virus proteins p54 and p30 are involved in two distinct steps of virus attachment and both contribute to the antibody-mediated protective immune response. Virology 1998, 243, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilan, J.; Zsak, L.; Lu, Z.; Burrage, T.; Kutish, G.; Rock, D. Neutralizing antibodies to African swine fever virus proteins p30, p54, and p72 are not sufficient for antibody-mediated protection. Virology 2004, 319, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Gonzalvo, F.; Rodríguez, F.; Escribano, J. Functional and immunological properties of the baculovirus-expressed hemagglutinin of African swine fever virus. Virology 1996, 218, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yang, J.; Ji, Y.; Okoth, E.; Liu, B.; Li, X.; Yin, H.; Zhu, Q. Recombinant Newcastle disease virus expressing African swine fever virus protein 72 is safe and immunogenic in mice. Virol. Sin. 2016, 31, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Chen, J.; Liang, W.; Chen, W.; Li, Z.; Chen, Q.; Cai, S. The recombinant pseudorabies virus expressing African swine fever virus CD2v protein is safe and effective in mice. Virol. J. 2020, 17, 180. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.V.; Mogler, M.; Certoma, A.; Green, D.; Monaghan, P.; Williams, D.T.; Rowland, R.R.R.; Gaudreault, N.N. Evaluation of an African swine fever (ASF) vaccine strategy incorporating priming with an alphavirus-expressed antigen followed by boosting with attenuated ASF virus. Arch. Virol. 2019, 164, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Goatley, L.C.; Reis, A.L.; Portugal, R.; Goldswain, H.; Shimmon, G.L.; Hargreaves, Z.; Ho, C.S.; Montoya, M.; Sanchez-Cordon, P.J.; Taylor, G.; et al. A Pool of Eight Virally Vectored African Swine Fever Antigens Protect Pigs Against Fatal Disease. Vaccines 2020, 8, 234. [Google Scholar] [CrossRef]
- Wang, Y.; Kang, W.; Yang, W.; Zhang, J.; Li, D.; Zheng, H. Structure of African Swine Fever Virus and Associated Molecular Mechanisms Underlying Infection and Immunosuppression: A Review. Front. Immunol. 2021, 12, 715582. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Olesen, A.S.; Lohse, L.; Boklund, A.; Halasa, T.; Gallardo, C.; Pejsak, Z.; Belsham, G.J.; Rasmussen, T.B.; Botner, A. Transmission of African swine fever virus from infected pigs by direct contact and aerosol routes. Vet. Microbiol. 2017, 211, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira De Oliveira, R.; Hutet, E.; Duhayon, M.; Guionnet, J.M.; Paboeuf, F.; Vial, L.; Le Potier, M.F. Successful Infection of Domestic Pigs by Ingestion of the European Soft Tick, O. Erraticus That Fed on African Swine Fever Virus Infected Pig. Viruses 2020, 12, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Shen, C.; Zhang, D.; Zhang, T.; Shi, X.; Yang, J.; Hao, Y.; Zhao, D.; Cui, H.; Yuan, X.; et al. Mechanism of interaction between virus and host is inferred from the changes of gene expression in macrophages infected with African swine fever virus CN/GS/2018 strain. Virol. J. 2021, 18, 170. [Google Scholar] [CrossRef]
- De Oliveira, V.; Almeida, S.; Soares, H.; Crespo, A.; Marshall-Clarke, S.; Parkhouse, R. A novel TLR3 inhibitor encoded by African swine fever virus (ASFV). Arch. Virol. 2011, 156, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Belmonte, R.; Perez-Nunez, D.; Pittau, M.; Richt, J.A.; Revilla, Y. African Swine Fever Virus Armenia/07 Virulent Strain Controls Interferon Beta Production through the cGAS-STING Pathway. J. Virol. 2019, 93, e02298-18. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Ventura, S.; Parkhouse, R.M. Identification and utility of innate immune system evasion mechanisms of ASFV. Virus Res. 2013, 173, 87–100. [Google Scholar] [CrossRef]
- Zhu, J.J.; Ramanathan, P.; Bishop, E.A.; O’Donnell, V.; Gladue, D.P.; Borca, M.V. Mechanisms of African swine fever virus pathogenesis and immune evasion inferred from gene expression changes in infected swine macrophages. PLoS ONE 2019, 14, e0223955. [Google Scholar] [CrossRef]
- Liu, X.; Ao, D.; Jiang, S.; Xia, N.; Xu, Y.; Shao, Q.; Luo, J.; Wang, H.; Zheng, W.; Chen, N.; et al. African Swine Fever Virus A528R Inhibits TLR8 Mediated NF-kappaB Activity by Targeting p65 Activation and Nuclear Translocation. Viruses 2021, 13, 2046. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Wu, Y.; Chen, H.; Zhang, S.; Li, J.; Xin, T.; Jia, H.; Hou, S.; Jiang, Y.; et al. Inhibition of cGAS-STING-TBK1 signaling pathway by DP96R of ASFV China 2018/1. Biochem. Biophys. Res. Commun. 2018, 506, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Guo, Z.; Ba, T.; Zhang, C.; He, L.; Zeng, C.; Dai, H. African Swine Fever Virus MGF360-12L Inhibits Type I Interferon Production by Blocking the Interaction of Importin alpha and NF-kappaB Signaling Pathway. Virol. Sin. 2020, 36, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, Z.; Feng, T.; Ma, Z.; Xue, Q.; Wu, P.; Li, P.; Li, S.; Yang, F.; Cao, W.; et al. African swine fever virus E120R protein inhibits interferon-β production by interacting with IRF3 to block its activation. J. Virol. 2021, 95, e0082421. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, S.; Feng, T.; Zhang, X.; Yang, F.; Cao, W.; Chen, H.; Liu, H.; Zhang, K.; Zhu, Z.; et al. African Swine Fever Virus F317L Protein Inhibits NF-kappaB Activation To Evade Host Immune Response and Promote Viral Replication. mSphere 2021, 6, e0065821. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, W.; Li, L.; Li, P.; Ma, Z.; Zhang, J.; Qi, X.; Ren, J.; Ru, Y.; Niu, Q.; et al. African Swine Fever Virus MGF-505-7R Negatively Regulates cGAS-STING-Mediated Signaling Pathway. J. Immunol. 2021, 206, 1844–1857. [Google Scholar] [CrossRef]
- Yang, K.; Huang, Q.; Wang, R.; Zeng, Y.; Cheng, M.; Xue, Y.; Shi, C.; Ye, L.; Yang, W.; Jiang, Y.; et al. African swine fever virus MGF505-11R inhibits type I interferon production by negatively regulating the cGAS-STING-mediated signaling pathway. Vet. Microbiol. 2021, 263, 109265. [Google Scholar] [CrossRef]
- Li, D.; Zhang, J.; Yang, W.; Li, P.; Ru, Y.; Kang, W.; Li, L.; Ran, Y.; Zheng, H. African swine fever virus protein MGF-505-7R promotes virulence and pathogenesis by inhibiting JAK1- and JAK2-mediated signaling. J. Biol. Chem. 2021, 297, 101190. [Google Scholar] [CrossRef]
- Huang, L.; Xu, W.; Liu, H.; Xue, M.; Liu, X.; Zhang, K.; Hu, L.; Li, J.; Liu, X.; Xiang, Z.; et al. African Swine Fever Virus pI215L Negatively Regulates cGAS-STING Signaling Pathway through Recruiting RNF138 to Inhibit K63-Linked Ubiquitination of TBK1. J. Immunol. 2021, 207, 2754–2769. [Google Scholar] [CrossRef]
- Chaulagain, S.; Delhon, G.A.; Khatiwada, S.; Rock, D.L. African Swine Fever Virus CD2v Protein Induces beta-Interferon Expression and Apoptosis in Swine Peripheral Blood Mononuclear Cells. Viruses 2021, 13, 1480. [Google Scholar] [CrossRef]
- Hurtado, C.; Bustos, M.J.; Granja, A.G.; de Leon, P.; Sabina, P.; Lopez-Vinas, E.; Gomez-Puertas, P.; Revilla, Y.; Carrascosa, A.L. The African swine fever virus lectin EP153R modulates the surface membrane expression of MHC class I antigens. Arch. Virol. 2011, 156, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Malerød, L.; Berg, T.; Kjeken, R. Clathrin-dependent endocytosis. Biochem. J. 2004, 377, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mercer, J.; Helenius, A. Virus entry by macropinocytosis. Nat. Cell Bio. 2009, 11, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.G.; Perez-Nunez, D.; Revilla, Y. Mechanisms of Entry and Endosomal Pathway of African Swine Fever Virus. Vaccines 2017, 5, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argilaguet, J.M.; Perez-Martin, E.; Nofrarias, M.; Gallardo, C.; Accensi, F.; Lacasta, A.; Mora, M.; Ballester, M.; Galindo-Cardiel, I.; Lopez-Soria, S.; et al. DNA vaccination partially protects against African swine fever virus lethal challenge in the absence of antibodies. PLoS ONE 2012, 7, e40942. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Chen, Q.; Ge, L.; Liu, Z.; Zhou, X.; Qi, Y.; Miao, F.; Wu, T.; Wang, L.; Yang, J.; et al. Characterizing Lansibai-2 pigs, a special breed in China, resistant to African swine fever. Chin. J. Vet. Sci. 2020, 40, 665–672. [Google Scholar]
- Karmakar, S.; Pal, P.; Lal, G. Key Activating and Inhibitory Ligands Involved in the Mobilization of Natural Killer Cells for Cancer Immunotherapies. Immunotargets Ther. 2021, 10, 387–407. [Google Scholar] [CrossRef]
- Franzoni, G.; Graham, S.P.; Sanna, G.; Angioi, P.; Fiori, M.S.; Anfossi, A.; Amadori, M.; Dei Giudici, S.; Oggiano, A. Interaction of porcine monocyte-derived dendritic cells with African swine fever viruses of diverse virulence. Vet. Microbiol. 2018, 216, 190–197. [Google Scholar] [CrossRef]
- Franzoni, G.; Graham, S.P.; Giudici, S.D.; Bonelli, P.; Pilo, G.; Anfossi, A.G.; Pittau, M.; Nicolussi, P.S.; Laddomada, A.; Oggiano, A. Characterization of the interaction of African swine fever virus with monocytes and derived macrophage subsets. Vet. Microbiol. 2017, 198, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Franzoni, G.; Graham, S.P.; Dei Giudici, S.; Oggiano, A. Porcine Dendritic Cells and Viruses: An Update. Viruses 2019, 11, 445. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Cordón, P.; Romero-Trevejo, J.; Pedrera, M.; Sánchez-Vizcaíno, J.; Bautista, M.; Gómez-Villamandos, J. Role of hepatic macrophages during the viral haemorrhagic fever induced by African Swine Fever Virus. Histolo. Histopathol. 2008, 23, 683–691. [Google Scholar] [CrossRef]
- Franzoni, G.; Dei Giudici, S.; Oggiano, A. Infection, modulation and responses of antigen-presenting cells to African swine fever viruses. Virus Res. 2018, 258, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Gonzalvo, F.; Caballero, C.; Martinez, J.; Carnero, M. Neutralization of African swine fever virus by sera from African swine fever-resistant pigs. Am. J. Vet. Res. 1986, 47, 1858–1862. [Google Scholar] [PubMed]
- Zsak, L.; Onisk, D.; Afonso, C.; Rock, D. Virulent African swine fever virus isolates are neutralized by swine immune serum and by monoclonal antibodies recognizing a 72-kDa viral protein. Virology 1993, 196, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Escribano, J.M.; Galindo, I.; Alonso, C. Antibody-mediated neutralization of African swine fever virus: Myths and facts. Virus Res. 2013, 173, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Leitão, A.; Cartaxeiro, C.; Coelho, R.; Cruz, B.; Parkhouse, R.; Portugal, F.; Vigário, J.; Martins, C. The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response. J. Gen. Virol. 2001, 82, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Nogal, M.L.; Gonzalez de Buitrago, G.; Rodriguez, C.; Cubelos, B.; Carrascosa, A.L.; Salas, M.L.; Revilla, Y. African swine fever virus IAP homologue inhibits caspase activation and promotes cell survival in mammalian cells. J. Virol. 2001, 75, 2535–2543. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, C.; Granja, A.G.; Bustos, M.J.; Nogal, M.L.; Gonzalez de Buitrago, G.; de Yebenes, V.G.; Salas, M.L.; Revilla, Y.; Carrascosa, A.L. The C-type lectin homologue gene (EP153R) of African swine fever virus inhibits apoptosis both in virus infection and in heterologous expression. Virology 2004, 326, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Banjara, S.; Caria, S.; Dixon, L.K.; Hinds, M.G.; Kvansakul, M. Structural Insight into African Swine Fever Virus A179L-Mediated Inhibition of Apoptosis. J. Virol. 2017, 91, e02228-16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Moon, A.; Childs, K.; Goodbourn, S.; Dixon, L.K. The African swine fever virus DP71L protein recruits the protein phosphatase 1 catalytic subunit to dephosphorylate eIF2alpha and inhibits CHOP induction but is dispensable for these activities during virus infection. J. Virol. 2010, 84, 10681–10689. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.A.; Du, C.; Xu, M.; Wang, X.; Ley, T.J. DFF45/ICAD can be directly processed by granzyme B during the induction of apoptosis. Immunity 2000, 12, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, A.; Zani, L.; Pikalo, J.; Hühr, J.; Sehl, J.; Mettenleiter, T.C.; Breithaupt, A.; Blome, S.; Blohm, U. T-cell responses in domestic pigs and wild boar upon infection with the moderately virulent African swine fever virus strain ‘Estonia2014’. Transbound. Emerg. Dis. 2021, 68, 2733–2749. [Google Scholar] [CrossRef] [PubMed]
- Hernaez, B.; Diaz-Gil, G.; Garcia-Gallo, M.; Ignacio Quetglas, J.; Rodriguez-Crespo, I.; Dixon, L.; Escribano, J.M.; Alonso, C. The African swine fever virus dynein-binding protein p54 induces infected cell apoptosis. FEBS Lett. 2004, 569, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Burmakina, G.; Malogolovkin, A.; Tulman, E.R.; Xu, W.; Delhon, G.; Kolbasov, D.; Rock, D.L. Identification of T-cell epitopes in African swine fever virus CD2v and C-type lectin proteins. J. Gen. Virol. 2019, 100, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Camos, L.; Lopez, E.; Navas, M.J.; Pina-Pedrero, S.; Accensi, F.; Correa-Fiz, F.; Park, C.; Carrascal, M.; Dominguez, J.; Salas, M.L.; et al. Identification of Promiscuous African Swine Fever Virus T-Cell Determinants Using a Multiple Technical Approach. Vaccines 2021, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Camos, L.; Lopez, E.; Collado, J.; Navas, M.J.; Blanco-Fuertes, M.; Pina-Pedrero, S.; Accensi, F.; Salas, M.L.; Mundt, E.; Nikolin, V.; et al. M448R and MGF505-7R: Two African Swine Fever Virus Antigens Commonly Recognized by ASFV-Specific T-Cells and with Protective Potential. Vaccines 2021, 9, 508. [Google Scholar] [CrossRef] [PubMed]
- Netherton, C.L.; Goatley, L.C.; Reis, A.L.; Portugal, R.; Nash, R.H.; Morgan, S.B.; Gault, L.; Nieto, R.; Norlin, V.; Gallardo, C.; et al. Identification and Immunogenicity of African Swine Fever Virus Antigens. Front. Immunol. 2019, 10, 1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jancovich, J.K.; Chapman, D.; Hansen, D.T.; Robida, M.D.; Loskutov, A.; Craciunescu, F.; Borovkov, A.; Kibler, K.; Goatley, L.; King, K.; et al. Immunization of Pigs by DNA Prime and Recombinant Vaccinia Virus Boost To Identify and Rank African Swine Fever Virus Immunogenic and Protective Proteins. J. Virol. 2018, 92, e02219-17. [Google Scholar] [CrossRef] [Green Version]
- Teklue, T.; Sun, Y.; Abid, M.; Luo, Y.; Qiu, H.J. Current status and evolving approaches to African swine fever vaccine development. Transbound. Emerg. Dis. 2020, 67, 529–542. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Pina-Pedrero, S.; Zhu, J.; Rodriguez, F.; Borca, M.V.; et al. Deletion of E184L, a putative DIVA target from the pandemic strain of African swine fever virus, produces a reduction in virulence and protection against virulent challenge. J. Virol. 2021, 96, e01419-21. [Google Scholar] [CrossRef]
- Krug, P.W.; Holinka, L.G.; O’Donnell, V.; Reese, B.; Sanford, B.; Fernandez-Sainz, I.; Gladue, D.P.; Arzt, J.; Rodriguez, L.; Risatti, G.R.; et al. The progressive adaptation of a georgian isolate of African swine fever virus to vero cells leads to a gradual attenuation of virulence in swine corresponding to major modifications of the viral genome. J. Virol. 2015, 89, 2324–2332. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wang, L.; Han, Y.; Pan, L.; Yang, J.; Sun, M.; Zhou, P.; Sun, Y.; Bi, Y.; Qiu, H.J. Adaptation of African swine fever virus to HEK293T cells. Transbound. Emerg. Dis. 2021, 68, 2853–2866. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.; Rai, A.; Ramirez-Medina, E.; Silva, E.; Velazquez-Salinas, L.; Vuono, E.; Pruitt, S.; Espinoza, N.; Gladue, D. A Cell Culture-Adapted Vaccine Virus against the Current African Swine Fever Virus Pandemic Strain. J. Virol. 2021, 95, e0012321. [Google Scholar] [CrossRef] [PubMed]
- Takenouchi, T.; Kitani, H.; Suzuki, S.; Nakai, M.; Fuchimoto, D.I.; Tsukimoto, M.; Shinkai, H.; Sato, M.; Uenishi, H. Immortalization and Characterization of Porcine Macrophages That Had Been Transduced with Lentiviral Vectors Encoding the SV40 Large T Antigen and Porcine Telomerase Reverse Transcriptase. Front. Vet. Sci. 2017, 4, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.; Wang, Q.; Lin, P.; Liu, C.; Li, L.; Wu, X.; Chi, T.; Xu, T.; Ge, S.; Liu, Y.; et al. Genome comparison of African swine fever virus China/2018/AnhuiXCGQ strain and related European p72 Genotype II strains. Transbound. Emerg. Dis. 2019, 66, 1167–1176. [Google Scholar] [CrossRef]
- Monteagudo, P.; Lacasta, A.; López, E.; Bosch, L.; Collado, J.; Pina-Pedrero, S.; Correa-Fiz, F.; Accensi, F.; Navas, M.; Vidal, E.; et al. BA71ΔCD2: A New Recombinant Live Attenuated African Swine Fever Virus with Cross-Protective Capabilities. J. Virol. 2017, 91, e01058-17. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Puertas, P.; Rodriguez, F.; Oviedo, J.M.; Ramiro-Ibanez, F.; Ruiz-Gonzalvo, F.; Alonso, C.; Escribano, J.M. Neutralizing antibodies to different proteins of African swine fever virus inhibit both virus attachment and internalization. J. Virol. 1996, 70, 5689–5694. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.; de la Torre, A.; Dixon, L.; Gallardo, C.; Jori, F.; Laddomada, A.; Martins, C.; Parkhouse, R.M.; Revilla, Y.; Rodriguez, F.A.J.; et al. Approaches and Perspectives for Development of African Swine Fever Virus Vaccines. Vaccines 2017, 5, 35. [Google Scholar] [CrossRef]
- Argilaguet, J.M.; Perez-Martin, E.; Gallardo, C.; Salguero, F.J.; Borrego, B.; Lacasta, A.; Accensi, F.; Diaz, I.; Nofrarias, M.; Pujols, J.; et al. Enhancing DNA immunization by targeting ASFV antigens to SLA-II bearing cells. Vaccine 2011, 29, 5379–5385. [Google Scholar] [CrossRef]
- Lokhandwala, S.; Petrovan, V.; Popescu, L.; Sangewar, N.; Elijah, C.; Stoian, A.; Olcha, M.; Ennen, L.; Bray, J.; Bishop, R.P.; et al. Adenovirus-vectored African Swine Fever Virus antigen cocktails are immunogenic but not protective against intranasal challenge with Georgia 2007/1 isolate. Vet. Microbiol. 2019, 235, 10–20. [Google Scholar] [CrossRef]
- Oura, C.; Denyer, M.; Takamatsu, H.; Parkhouse, R. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus. J. Gen. Virol. 2005, 86, 2445–2450. [Google Scholar] [CrossRef]
- Sunwoo, S.Y.; Perez-Nunez, D.; Morozov, I.; Sanchez, E.G.; Gaudreault, N.N.; Trujillo, J.D.; Mur, L.; Nogal, M.; Madden, D.; Urbaniak, K.; et al. DNA-Protein Vaccination Strategy Does Not Protect from Challenge with African Swine Fever Virus Armenia 2007 Strain. Vaccines 2019, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacasta, A.; Ballester, M.; Monteagudo, P.L.; Rodriguez, J.M.; Salas, M.L.; Accensi, F.; Pina-Pedrero, S.; Bensaid, A.; Argilaguet, J.; Lopez-Soria, S.; et al. Expression library immunization can confer protection against lethal challenge with African swine fever virus. J. Virol. 2014, 88, 13322–13332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopera-Madrid, J.; Osorio, J.E.; He, Y.; Xiang, Z.; Adams, L.G.; Laughlin, R.C.; Mwangi, W.; Subramanya, S.; Neilan, J.; Brake, D.; et al. Safety and immunogenicity of mammalian cell derived and Modified Vaccinia Ankara vectored African swine fever subunit antigens in swine. Vet. Immunol. Immunop. 2017, 185, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Qian, H.; Wu, Y.; Wu, S.; Che, Y. Subunit Vaccine Composition for African Swine Fever, and Preparation Therefor and Use Thereof. CN113940992A, 18 January 2022. [Google Scholar]
- Zheng, H.; Mao, R.; Yang, F.; Zhou, X.; Sun, D.; Liu, H.; Zhang, K.; Liu, X. Polypeptides for Promoting Swine Body to Generate African Swine Fever Virus (ASFV) Antigen-Specific Immune Response and Application of Polypeptides. CN113896772A, 7 January 2022. [Google Scholar]
- Nolan, T.; Fortanier, A.; Leav, B.; Põder, A.; Bravo, L.; Szymański, H.; Heeringa, M.; Vermeulen, W.; Matassa, V.; Smolenov, I.; et al. Efficacy of a Cell-Culture-Derived Quadrivalent Influenza Vaccine in Children. N. Engl. J. Med. 2021, 385, 1485–1495. [Google Scholar] [CrossRef]
- Trabelsi, K.; Ben Zakour, M.; Kallel, H. Purification of rabies virus produced in Vero cells grown in serum free medium. Vaccine 2019, 37, 7052–7060. [Google Scholar] [CrossRef]
- Satchidanandam, V. Japanese Encephalitis Vaccines. Curr. Treat. Options Infect. Dis. 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Stone, S.; Hess, W. Antibody response to inactivated preparations of African swine fever virus in pigs. Am. J. Vet. Res. 1967, 28, 475–481. [Google Scholar]
- Bidet, K.; Dadlani, D.; Garcia-Blanco, M.A. Correction: G3BP1, G3BP2 and CAPRIN1 Are Required for Translation of Interferon Stimulated mRNAs and Are Targeted by a Dengue Virus Non-coding RNA. PLoS Pathog. 2017, 13, e1006295. [Google Scholar] [CrossRef] [Green Version]
- Blome, S.; Gabriel, C.; Beer, M. Modern adjuvants do not enhance the efficacy of an inactivated African swine fever virus vaccine preparation. Vaccine 2014, 32, 3879–3882. [Google Scholar] [CrossRef]
- Global Situation of African Swine Fever 2016–2020. Available online: https://www.oie.int/app/uploads/2021/03/report-47-global-situation-asf.pdf (accessed on 28 December 2021).
- Barasona, J.A.; Gallardo, C.; Cadenas-Fernandez, E.; Jurado, C.; Rivera, B.; Rodriguez-Bertos, A.; Arias, M.; Sanchez-Vizcaino, J.M. First Oral Vaccination of Eurasian Wild Boar Against African Swine Fever Virus Genotype II. Front. Vet. Sci. 2019, 6, 137. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelzaher, H.M.; Gabr, A.S.; Saleh, B.M.; Abdel Gawad, R.M.; Nour, A.A.; Abdelanser, A. RNA Vaccines against Infectious Diseases: Vital Progress with Room for Improvement. Vaccines 2021, 9, 1211. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Pascolo, S. Vaccination with messenger RNA (mRNA). Handb. Exp. Pharmacol. 2008, 183, 221–235. [Google Scholar] [CrossRef]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccin. Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, T.; Ulbert, S. Improvement of DNA vaccination by adjuvants and sophisticated delivery devices: Vaccine-platforms for the battle against infectious diseases. Clin. Exp. Vaccine Res. 2015, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pollard, C.; De Koker, S.; Saelens, X.; Vanham, G.; Grooten, J. Challenges and advances towards the rational design of mRNA vaccines. Trends Mol. Med. 2013, 19, 705–713. [Google Scholar] [CrossRef]
- Fuchs, J.D.; Frank, I.; Elizaga, M.L.; Allen, M.; Frahm, N.; Kochar, N.; Li, S.; Edupuganti, S.; Kalams, S.A.; Tomaras, G.D.; et al. First-in-Human Evaluation of the Safety and Immunogenicity of a Recombinant Vesicular Stomatitis Virus Human Immunodeficiency Virus-1 gag Vaccine (HVTN 090). Open Forum Infect. Dis. 2015, 2, ofv082. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
A phylogenetic tree was constructed based on the whole genome sequences of 74 strains in the GenBank database, after which datasets for the sequences were aligned using MAFFT (version 7.149) program [32]. Maximum likelihood (ML) phylogenies for the codon alignment of the genome sequences were estimated using the GTRGAMMA nucleotide substitution model in the IQ-TREE 1.68 software [33]. Node support was determined by nonparametric bootstrapping with 1000 replicates, and the phylogenetic tree was visualized in the Figtree (version 1.4.3) program (http://tree.bio.ed.ac.uk/software/Figtree/) (accessed on 18 January 2022). Types written in red indicate Chinese isolates.
Figure 1.
A phylogenetic tree was constructed based on the whole genome sequences of 74 strains in the GenBank database, after which datasets for the sequences were aligned using MAFFT (version 7.149) program [32]. Maximum likelihood (ML) phylogenies for the codon alignment of the genome sequences were estimated using the GTRGAMMA nucleotide substitution model in the IQ-TREE 1.68 software [33]. Node support was determined by nonparametric bootstrapping with 1000 replicates, and the phylogenetic tree was visualized in the Figtree (version 1.4.3) program (http://tree.bio.ed.ac.uk/software/Figtree/) (accessed on 18 January 2022). Types written in red indicate Chinese isolates.

Figure 2.
ASFV genes regulate innate immune signaling pathway.


{kind=link}
{kind=link}
Table 1.
Summary of the most promising LAV candidates.
| Genes | Strains | Genotype | Minimal Protective Dose | Route | Challenge | Gene Function | References |
|---|---|---|---|---|---|---|---|
| I177L | Georgia2007/1 | II | 102HAD50 | IM | Georgia2007/1 | unknown | [19] |
| 106HAD50 | ON | Georgia2007/1 | [20] | ||||
| 102HAD50 | IM | TTKN/ASFV/DN/2019 | [21] | ||||
| A137R | Georgia2007/1 | II | 102HAD50 | IM | Georgia2007/1 | unknown | [23] |
| I226R I226R | SY18 | II | 104HAD50 | IM | SY18 | unknown | [22] |
| L7L-L11L | SY18 | II | 103HAD50 | IM | SY18 | unknown | [18] |
| MGF505/360(6) 1 and EP402R | HLJ/18 | II | 103HAD50 | IM | HLJ/18 | hemadsorbing and inhibition of type I interferon responses | [17] |
| 105HAD50 | ON | ||||||
| EP402R | Ba71V | I | 104HAD50 | IM | Ba71V | hemadsorbing | [86] |
| E75 | |||||||
| Georgia2007/1 |
1: MGF505-1R, MGF505-2R, MGF505-3R, MGF360-12L, MGF360-13L, MGF360-14L.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Z.; Ai, Q.; Huang, S.; Ou, Y.; Gao, Y.; Tong, T.; Fan, H. Immune Escape Mechanism and Vaccine Research Progress of African Swine Fever Virus. Vaccines 2022, 10, 344. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10030344
AMA Style
Wang Z, Ai Q, Huang S, Ou Y, Gao Y, Tong T, Fan H. Immune Escape Mechanism and Vaccine Research Progress of African Swine Fever Virus. Vaccines. 2022; 10(3):344. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10030344
Chicago/Turabian StyleWang, Zhaoyang, Qiangyun Ai, Shenglin Huang, Yating Ou, Yinze Gao, Tiezhu Tong, and Huiying Fan. 2022. "Immune Escape Mechanism and Vaccine Research Progress of African Swine Fever Virus" Vaccines 10, no. 3: 344. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10030344
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.