
Investigating the Role of Antigen Orientation on the Immune Response Elicited by Neisseria meningitidis Factor H Binding Protein on GMMA

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
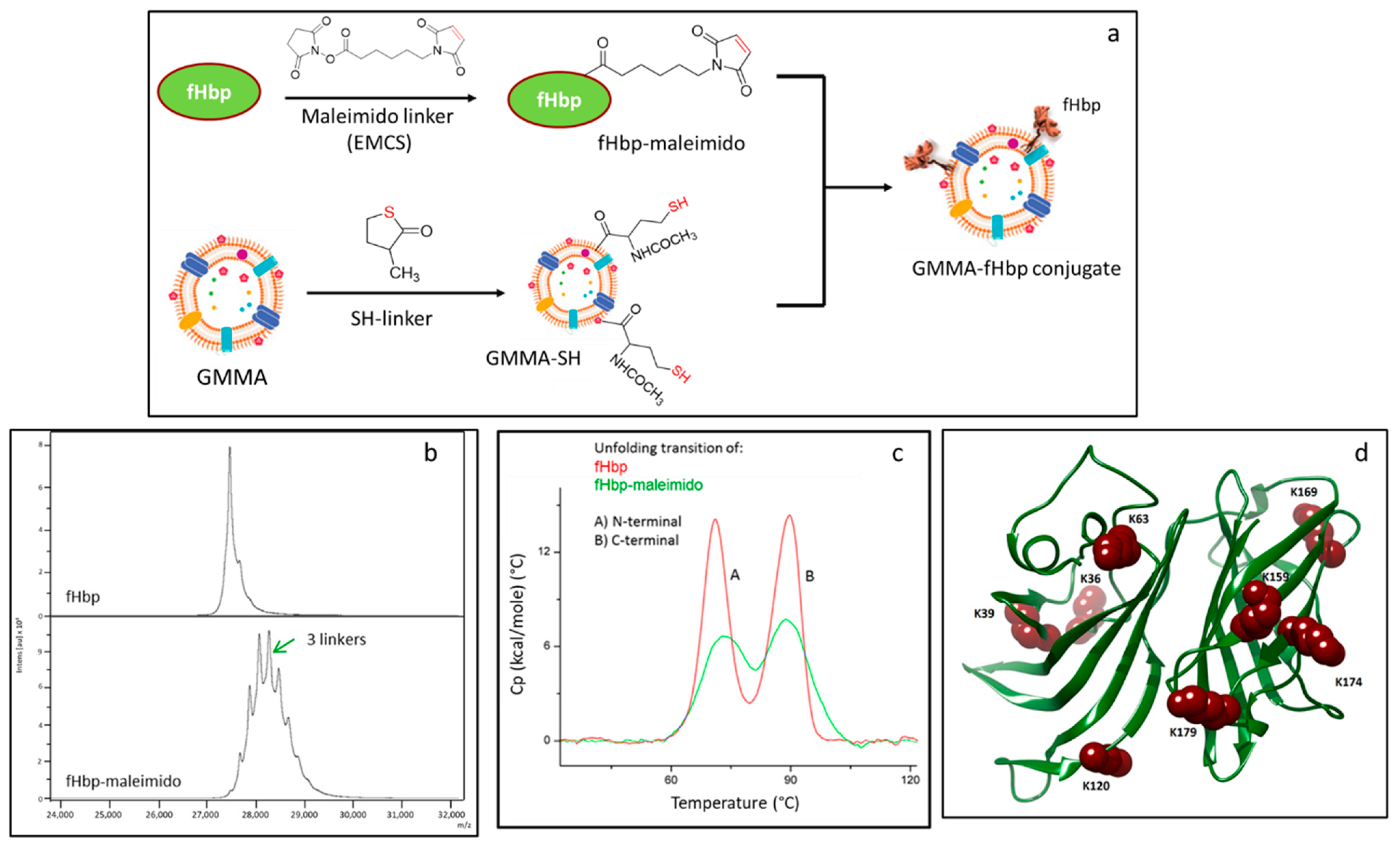
2.1. Synthesis and Characterization of the Conjugates
2.1.1. Characterization of fHbp Randomly Conjugated to GMMA
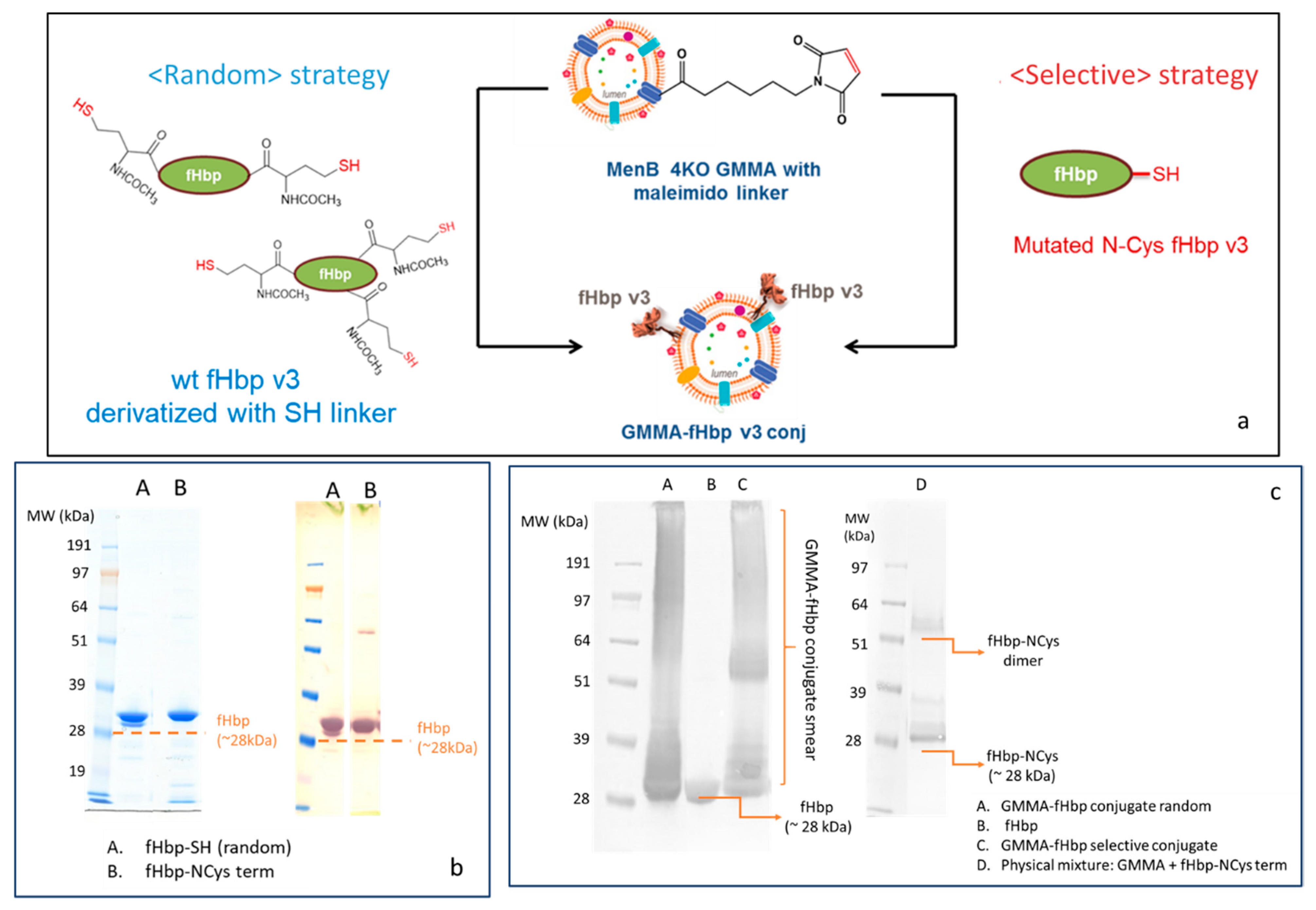
2.1.2. Synthesis of Selective and Random fHbp-GMMA Conjugates
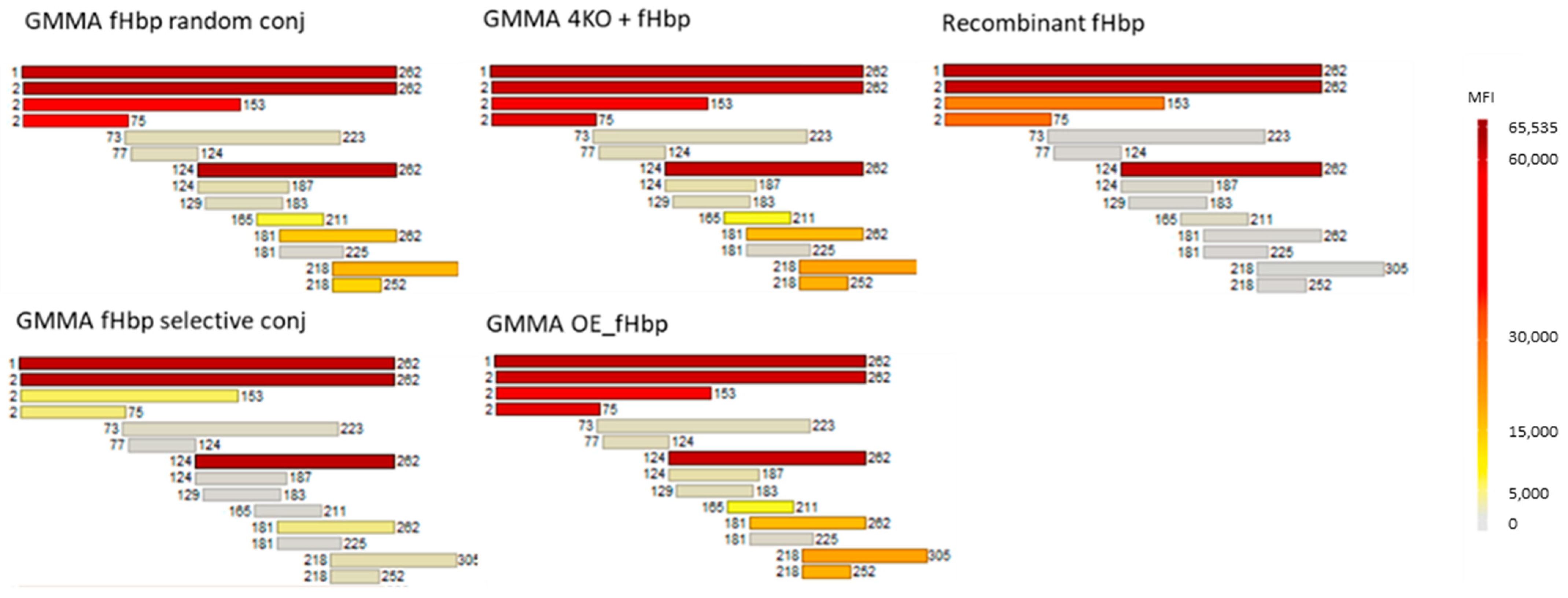
2.1.3. Quantification of fHbp in Random or Selective GMMA Conjugates
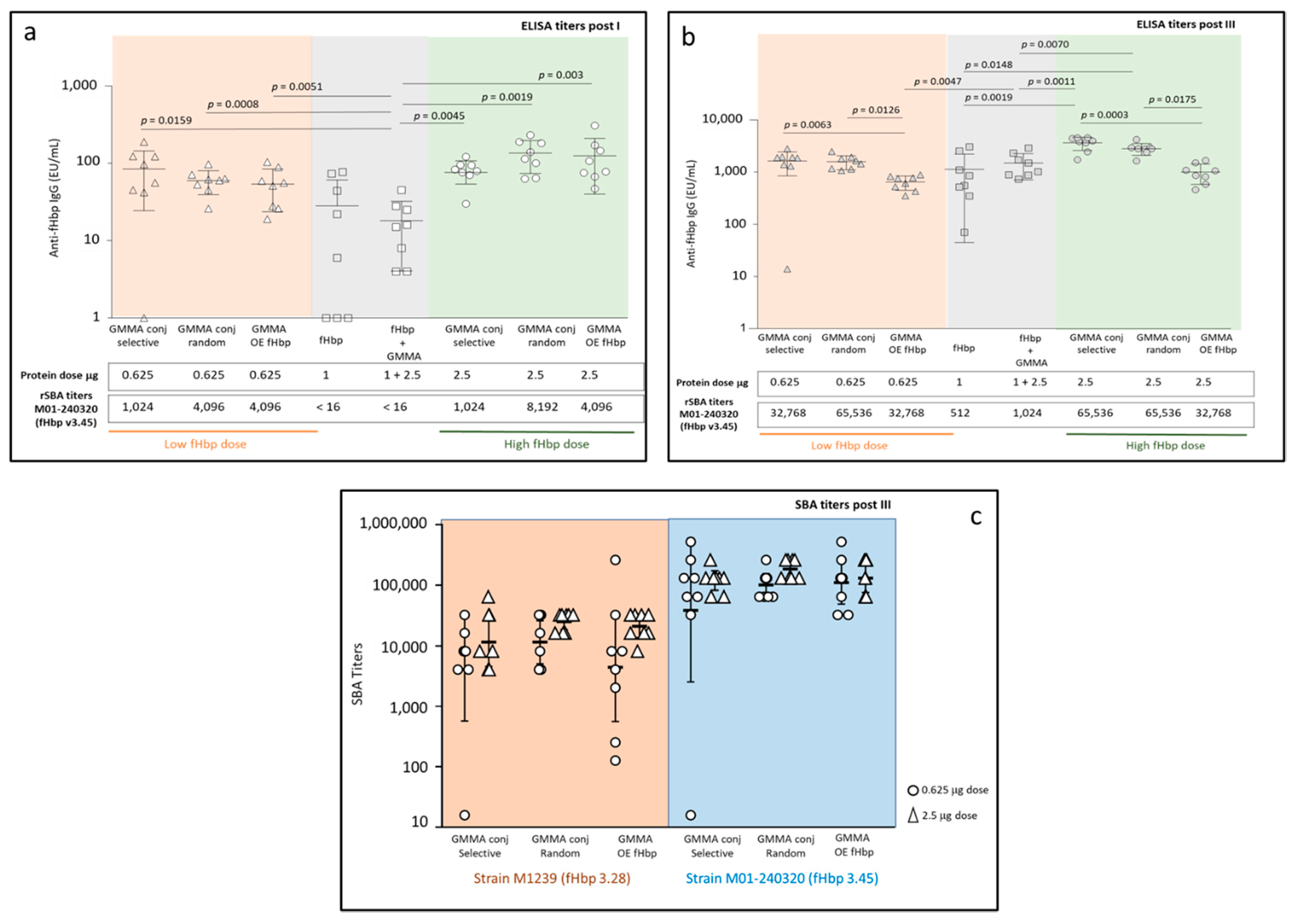
2.2. Immunogenicity in Mice: Random and Selective Conjugates Compared to MenB GMMA Overexpressing fHbp
3. Discussion
4. Materials and Methods
4.1. GMMA Production and Purification
4.2. fHbp and fHbp-Cys Term Production, Purification and Characterization
4.3. Synthesis and Characterization of the GMMA Conjugates
4.3.1. fHbp Derivatization with N-ε-malemidocaproyl-oxysuccinimde Ester (EMCS) Linker
4.3.2. fHbp Derivatization with SH Linker (N-acetyl-DL-homocysteine Thiolactone)
4.3.3. MenB GMMA Derivatization with N-ε-malemidocaproyl-oxysuccinimde Ester (EMCS) Linker
4.3.4. Conjugations
4.4. Differential Scanning Calorimetry (DSC)
4.4.1. Immunogenicity Studies in Mice
4.4.2. Statistical Analysis
4.5. Protein Array Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef] [PubMed]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, e1800376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, F.; Rossi, O.; Necchi, F.; Micoli, F. OMV Vaccines and the Role of TLR Agonists in Immune Response. Int. J. Mol. Sci. 2020, 21, 4416. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar] [CrossRef]
- Benne, N.; van Duijn, J.; Kuiper, J.; Jiskoot, W.; Slütter, B. Orchestrating immune responses: How size, shape and rigidity affect the immunogenicity of particulate vaccines. J. Control. Release 2016, 234, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Moyer, T.J.; Zmolek, A.C.; Irvine, D.J. Beyond antigens and adjuvants: Formulating future vaccines. J. Clin. Investig. 2016, 126, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef]
- Gerritzen, M.J.H.; Martens, D.E.; Wijffels, R.H.; van der Pol, L.; Stork, M. Bioengineering bacterial outer membrane vesicles as vaccine platform. Biotechnol. Adv. 2017, 35, 565–574. [Google Scholar] [CrossRef]
- Piccioli, D.; Bartolini, E.; Micoli, F. GMMA as a ‘plug and play’ technology to tackle infectious disease to improve global health: Context and perspectives for the future. Expert. Rev. Vaccines 2022, 21, 163–172. [Google Scholar] [CrossRef]
- van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer Membrane Vesicles: Current Status and Future Direction of These Novel Vaccine Adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holst, J.; Martin, D.; Arnold, R.; Huergo, C.C.; Oster, P.; O’Hallahan, J.; Rosenqvist, E. Properties and clinical performance of vaccines containing outer membrane vesicles from Neisseria meningitidis. Vaccine 2009, 27 (Suppl. S2), B3–B12. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R.; Pizza, M.; Masignani, V.; Vadivelu, K. Meningococcal B vaccine (4CMenB): The journey from research to real world experience. Expert. Rev. Vaccines 2018, 17, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, N.G.; Stephens, D.S. Neisseria meningitidis: Biology, microbiology, and epidemiology. Methods Mol. 2012, 799, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Purmohamad, A.; Abasi, E.; Azimi, T.; Hosseini, S.; Safari, H.; Nasiri, M.J.; Imani Fooladi, A.A. Global estimate of Neisseria meningitidis serogroups proportion in invasive meningococcal disease: A systematic review and meta-analysis. Microb. Pathog. 2019, 134, 103571. [Google Scholar] [CrossRef] [PubMed]
- Committee On Infectious Diseases; Byington, C.L.; Maldonado, Y.A.; Barnett, E.D.; Dele Davies, H.; Edwards, K.M.; Lynfield, R.; Munoz, F.M.; Nolt, D.L.; Nyquist, A.-C.; et al. Recommendations for Serogroup B Meningococcal Vaccine for Persons 10 Years and Older. Pediatrics 2016, 138, e20161890. [Google Scholar]
- Gandhi, A.; Balmer, P.; York, L.J. Characteristics of a new meningococcal serogroup B vaccine, bivalent rLP2086 (MenB-FHbp; Trumenba®). Postgrad. Med. 2016, 128, 548–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masignani, V.; Comanducci, M.; Giuliani, M.M.; Bambini, S.; Adu-Bobie, J.; Arico, B.; Brunelli, B.; Pieri, A.; Santini, L.; Savino, S.; et al. Vaccination against Neisseria meningitidis using three variants of the lipoprotein GNA1870. J. Exp. Med. 2003, 197, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.C.; Exley, R.M.; Chan, H.; Feavers, I.; Kang, Y.H.; Sim, R.B.; Tang, C.M. Functional significance of factor H binding to Neisseria meningitidis. J. Immunol. 2006, 176, 7566–7575. [Google Scholar] [CrossRef] [Green Version]
- Madico, G.; Welsch, J.A.; Lewis, L.A.; McNaughton, A.; Perlman, D.H.; Costello, C.E.; Ngampasutadol, J.; Vogel, U.; Granoff, D.M.; Ram, S. The meningococcal vaccine candidate GNA1870 binds the complement regulatory protein factor H and enhances serum resistance. J. Immunol. 2006, 177, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.Q.; Hoiseth, S.K.; Harris, S.L.; McNeil, L.K.; Zhu, D.; Tan, C.; Scott, A.A.; Alexander, K.; Mason, K.; Miller, L.; et al. Broad vaccine coverage predicted for a bivalent recombinant factor H binding protein based vaccine to prevent serogroup B meningococcal disease. Vaccine 2010, 28, 6086–6093. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.D.; Bernfield, L.; Barniak, V.; Farley, J.E.; Howell, A.; Knauf, M.; Ooi, P.; Smith, R.P.; Weise, P.; Wetherell, M.; et al. Vaccine potential of the Neisseria meningitidis 2086 lipoprotein. Infect. Immun. 2004, 72, 2088–2100. [Google Scholar] [CrossRef] [Green Version]
- Welsch, J.A.; Rossi, R.; Comanducci, M.; Granoff, D.M. Protective activity of monoclonal antibodies to genome-derived neisserial antigen 1870, a Neisseria meningitidis candidate vaccine. J. Immunol. 2004, 172, 5606–5615. [Google Scholar] [CrossRef] [Green Version]
- Welsch, J.A.; Ram, S.; Koeberling, O.; Granoff, D.M. Complement-dependent synergistic bactericidal activity of antibodies against factor H-binding protein, a sparsely distributed meningococcal vaccine antigen. J. Infect. Dis. 2008, 197, 1053–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granoff, D.M.; Welsch, J.A.; Ram, S. Binding of complement factor H (fH) to Neisseria meningitidis is specific for human fH and inhibits complement activation by rat and rabbit sera. Infect. Immun. 2009, 77, 764–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeberling, O.; Ispasanie, E.; Hauser, J.; Rossi, O.; Pluschke, G.; Caugant, D.A.; Saul, A.; MacLennan, C.A. A broadly-protective vaccine against meningococcal disease in sub-Saharan Africa based on generalized modules for membrane antigens (GMMA). Vaccine 2014, 32, 2688–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajon, R.; Fergus, A.M.; Granoff, D.M. Mutant Native Outer Membrane Vesicles Combined with a Serogroup A Polysaccharide Conjugate Vaccine for Prevention of Meningococcal Epidemics in Africa. PLoS ONE 2013, 8, e66536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Palmieri, E.; Balocchi, C.; Gasperini, G.; et al. GMMA Is a Versatile Platform to Design Effective Multivalent Combination Vaccines. Vaccines 2020, 8, 540. [Google Scholar] [CrossRef]
- Di Benedetto, R.; Alfini, R.; Carducci, M.; Aruta, M.G.; Lanzilao, L.; Acquaviva, A.; Palmieri, E.; Giannelli, C.; Necchi, F.; Saul, A.; et al. Novel Simple Conjugation Chemistries for Decoration of GMMA with Heterologous Antigens. Int. J. Mol. Sci. 2021, 22, 10180. [Google Scholar] [CrossRef]
- Seib, K.L.; Scarselli, M.; Comanducci, M.; Toneatto, D.; Masignani, V. Neisseria meningitidis factor H-binding protein fHbp: A key virulence factor and vaccine antigen. Expert. Rev. Vaccines 2015, 14, 841–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedetto, G.; Salvini, L.; Gotta, S.; Cescutti, P.; Micoli, F. Investigation on Sugar-Protein Connectivity in Salmonella O-Antigen Glycoconjugate Vaccines. Bioconjug. Chem. 2018, 29, 1736–1747. [Google Scholar] [CrossRef]
- Cantini, F.; Veggi, D.; Dragonetti, S.; Savino, S.; Scarselli, M.; Romagnoli, G.; Pizza, M.; Banci, L.; Rappuoli, R. Solution structure of the factor H-binding protein, a survival factor and protective antigen of Neisseria meningitidis. J. Biol. Chem. 2009, 284, 9022–9026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Necchi, F.; Stefanetti, G.; Alfini, R.; Palmieri, E.; Carducci, M.; Di Benedetto, R.; Schiavo, F.; Aruta, M.G.; Giusti, F.; Ferlenghi, I.; et al. Neisseria meningitidis Factor H Binding Protein Surface Exposure on Salmonella Typhimurium GMMA Is Critical to Induce an Effective Immune Response against Both Diseases. Pathogens 2021, 10, 726. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Micoli, F.; Necchi, F.; Pizza, M.; Berlanda Scorza, F.; Rossi, O. GMMA-Based Vaccines: The Known and The Unknown. Front. Immunol. 2021, 12, 715393. [Google Scholar] [CrossRef]
- Marini, A.; Rossi, O.; Aruta, M.G.; Micoli, F.; Rondini, S.; Guadagnuolo, S.; Delany, I.; Henderson, I.R.; Cunningham, A.F.; Saul, A.; et al. Contribution of factor H-Binding protein sequence to the cross-reactivity of meningococcal native outer membrane vesicle vaccines with over-expressed fHbp variant group 1. PLoS ONE 2017, 12, e0181508. [Google Scholar] [CrossRef] [Green Version]
- Faleri, A.; Santini, L.; Brier, S.; Pansegrau, W.; Lo Surdo, P.; Scarselli, M.; Buricchi, F.; Volpini, G.; Genovese, A.; van der Veen, S.; et al. Two cross-reactive monoclonal antibodies recognize overlapping epitopes on Neisseria meningitidis factor H binding protein but have different functional properties. FASEB J. 2014, 28, 1644–1653. [Google Scholar] [CrossRef]
- Vu, D.M.; Pajon, R.; Reason, D.C.; Granoff, D.M. A broadly cross-reactive monoclonal antibody against an epitope on the n-terminus of meningococcal fHbp. Sci. Rep. 2012, 2, 341. [Google Scholar] [CrossRef] [Green Version]
- Malito, E.; Faleri, A.; Lo Surdo, P.; Veggi, D.; Maruggi, G.; Grassi, E.; Cartocci, E.; Bertoldi, I.; Genovese, A.; Santini, L.; et al. Defining a protective epitope on factor H binding protein, a key meningococcal virulence factor and vaccine antigen. Proc. Natl. Acad. Sci. USA 2013, 110, 3304–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beernink, P.T.; Welsch, J.A.; Bar-Lev, M.; Koeberling, O.; Comanducci, M.; Granoff, D.M. Fine antigenic specificity and cooperative bactericidal activity of monoclonal antibodies directed at the meningococcal vaccine candidate factor h-binding protein. Infect. Immun. 2008, 76, 4232–4240. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, M.; Bartolini, E.; Galli, B.; Santini, L.; Lo Surdo, P.; Buricchi, F.; Bruttini, M.; Benucci, B.; Pacchiani, N.; Alleri, L.; et al. Human protective response induced by meningococcus B vaccine is mediated by the synergy of multiple bactericidal epitopes. Sci. Rep. 2018, 8, 3700. [Google Scholar] [CrossRef] [PubMed]
- López-Sagaseta, J.; Malito, E.; Rappuoli, R.; Bottomley, M.J. Self-assembling protein nanoparticles in the design of vaccines. Comput. Struct. Biotechnol. J. 2016, 14, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.G.; Taylor, J.A.; Sibilo, M.Q.; Miura, K.; Mallory, K.L.; Mann, C.; Karch, C.; Beck, Z.; Matyas, G.R.; Long, C.A.; et al. Orientation of Antigen Display on Self-Assembling Protein Nanoparticles Influences Immunogenicity. Vaccines 2021, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Arigita, C.; Kersten, G.F.; Hazendonk, T.; Hennink, W.E.; Crommelin, D.J.; Jiskoot, W. Restored functional immunogenicity of purified meningococcal PorA by incorporation into liposomes. Vaccine 2003, 21, 950–960. [Google Scholar] [CrossRef]
- Muralinath, M.; Kuehn, M.J.; Roland, K.L.; Curtiss, R., 3rd. Immunization with Salmonella enterica serovar Typhimurium-derived outer membrane vesicles delivering the pneumococcal protein PspA confers protection against challenge with Streptococcus pneumoniae. Infect. Immun. 2011, 79, 887–894. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Wang, S.; Yao, Y.; Xia, Y.; Yang, X.; Li, K.; Sun, P.; Liu, C.; Sun, W.; Bai, H.; et al. Employing Escherichia coli-derived outer membrane vesicles as an antigen delivery platform elicits protective immunity against Acinetobacter baumannii infection. Sci. Rep. 2016, 6, 37242. [Google Scholar] [CrossRef]
- Bartolini, E.; Ianni, E.; Frigimelica, E.; Petracca, R.; Galli, G.; Berlanda Scorza, F.; Norais, N.; Laera, D.; Giusti, F.; Pierleoni, A.; et al. Recombinant outer membrane vesicles carrying Chlamydia muridarum HtrA induce antibodies that neutralize chlamydial infection in vitro. J. Extracell. Vesicles 2013, 2, 20181. [Google Scholar] [CrossRef]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Oldrini, D.; Pitirollo, O.; Gasperini, G.; et al. Generalized Modules for Membrane Antigens as Carrier for Polysaccharides: Impact of Sugar Length, Density, and Attachment Site on the Immune Response Elicited in Animal Models. Front. Immunol. 2021, 12, 719315. [Google Scholar] [CrossRef]
- Micoli, F.; Alfini, R.; Giannelli, C. Methods for Assessment of OMV/GMMA Quality and Stability. Methods Mol. Biol. 2022, 2414, 227–279. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Santini, L.; Brunelli, B.; Biolchi, A.; Arico, B.; Di Marcello, F.; Cartocci, E.; Comanducci, M.; Masignani, V.; Lozzi, L.; et al. The region comprising amino acids 100 to 255 of Neisseria meningitidis lipoprotein GNA 1870 elicits bactericidal antibodies. Infect. Immun. 2005, 73, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, E.; Borgogni, E.; Bruttini, M.; Muzzi, A.; Giuliani, M.; Iozzi, S.; Petracca, R.; Martinelli, M.; Bonacci, S.; Marchi, S.; et al. Immunological fingerprint of 4CMenB recombinant antigens via protein microarray reveals key immunosignatures correlating with bactericidal activity. Nat. Commun. 2020, 11, 4994. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfini, R.; Brunelli, B.; Bartolini, E.; Carducci, M.; Luzzi, E.; Ferlicca, F.; Buccato, S.; Galli, B.; Lo Surdo, P.; Scarselli, M.; et al. Investigating the Role of Antigen Orientation on the Immune Response Elicited by Neisseria meningitidis Factor H Binding Protein on GMMA. Vaccines 2022, 10, 1182. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10081182
Alfini R, Brunelli B, Bartolini E, Carducci M, Luzzi E, Ferlicca F, Buccato S, Galli B, Lo Surdo P, Scarselli M, et al. Investigating the Role of Antigen Orientation on the Immune Response Elicited by Neisseria meningitidis Factor H Binding Protein on GMMA. Vaccines. 2022; 10(8):1182. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10081182
Chicago/Turabian StyleAlfini, Renzo, Brunella Brunelli, Erika Bartolini, Martina Carducci, Enrico Luzzi, Francesca Ferlicca, Scilla Buccato, Barbara Galli, Paola Lo Surdo, Maria Scarselli, and et al. 2022. "Investigating the Role of Antigen Orientation on the Immune Response Elicited by Neisseria meningitidis Factor H Binding Protein on GMMA" Vaccines 10, no. 8: 1182. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10081182