Chimeric Hemagglutinin-Based Live-Attenuated Vaccines Confer Durable Protective Immunity against Influenza A Viruses in a Preclinical Ferret Model


 , , and
, , and 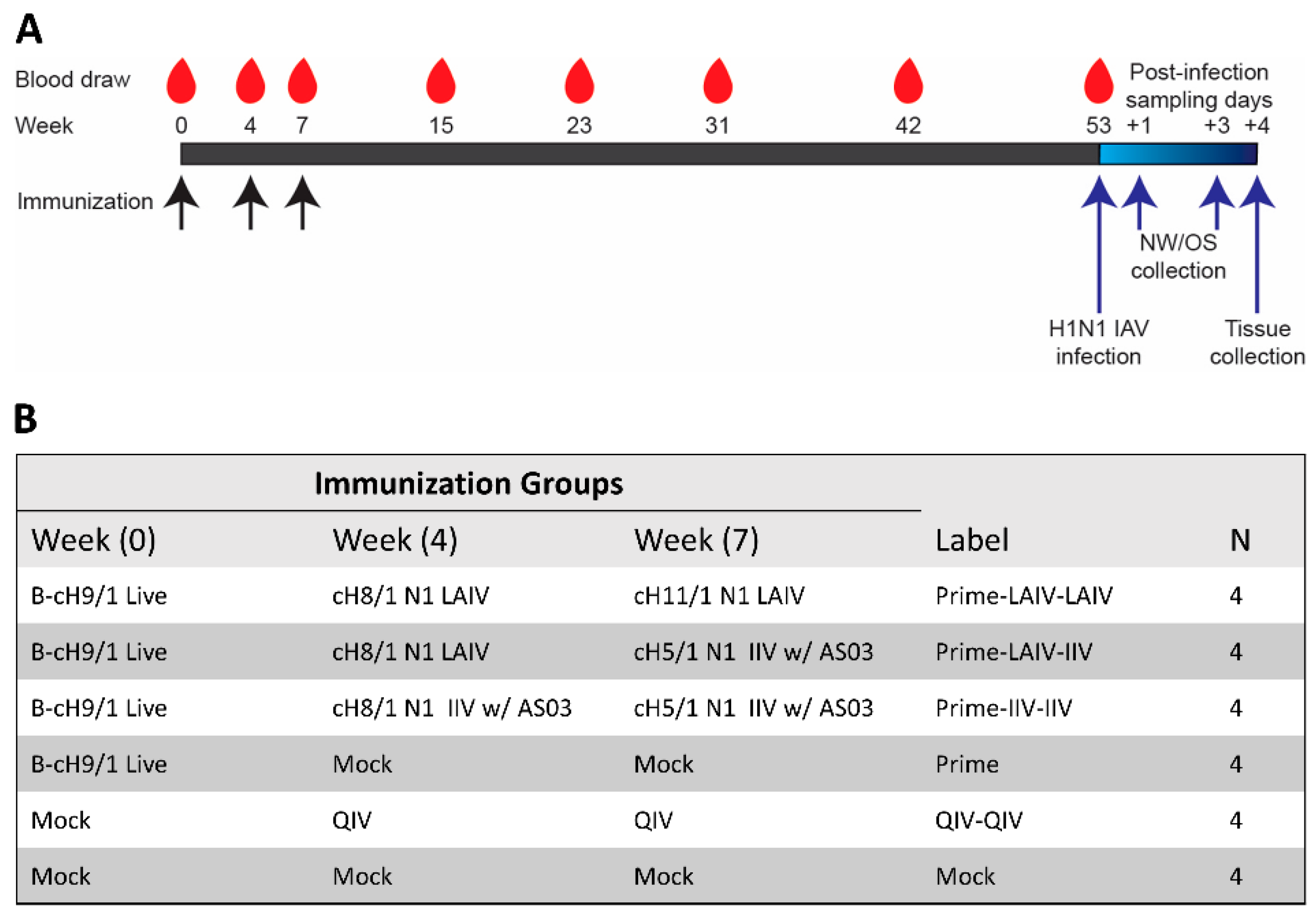{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells, Viruses, and Proteins
2.3. cHA-Based Universal Vaccine and QIV Vaccine Preparations
2.4. Ferret Immunization and Challenge
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Enzyme-Linked Lectin Assay (ELLA)
2.7. Hemagglutination Inhibition (HI) Assays
2.8. Preparation of Single-Cell Suspensions from Ferret Tissues and Peripheral Blood Mononuclear Cells (PBMCs)
2.9. Peptide Stimulation and Flow Cytometry Analyses
2.10. Statistical Analysis
3. Results
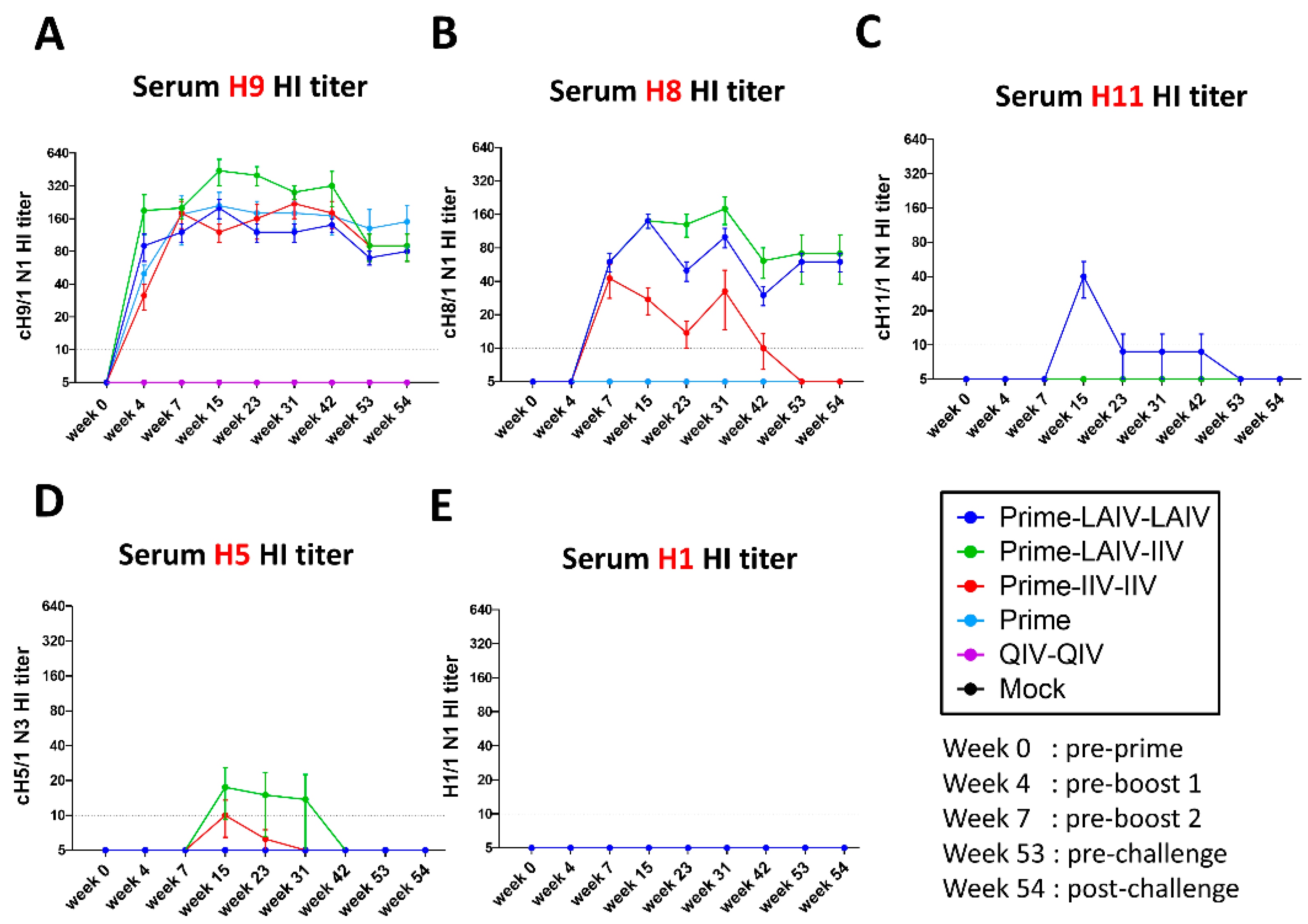
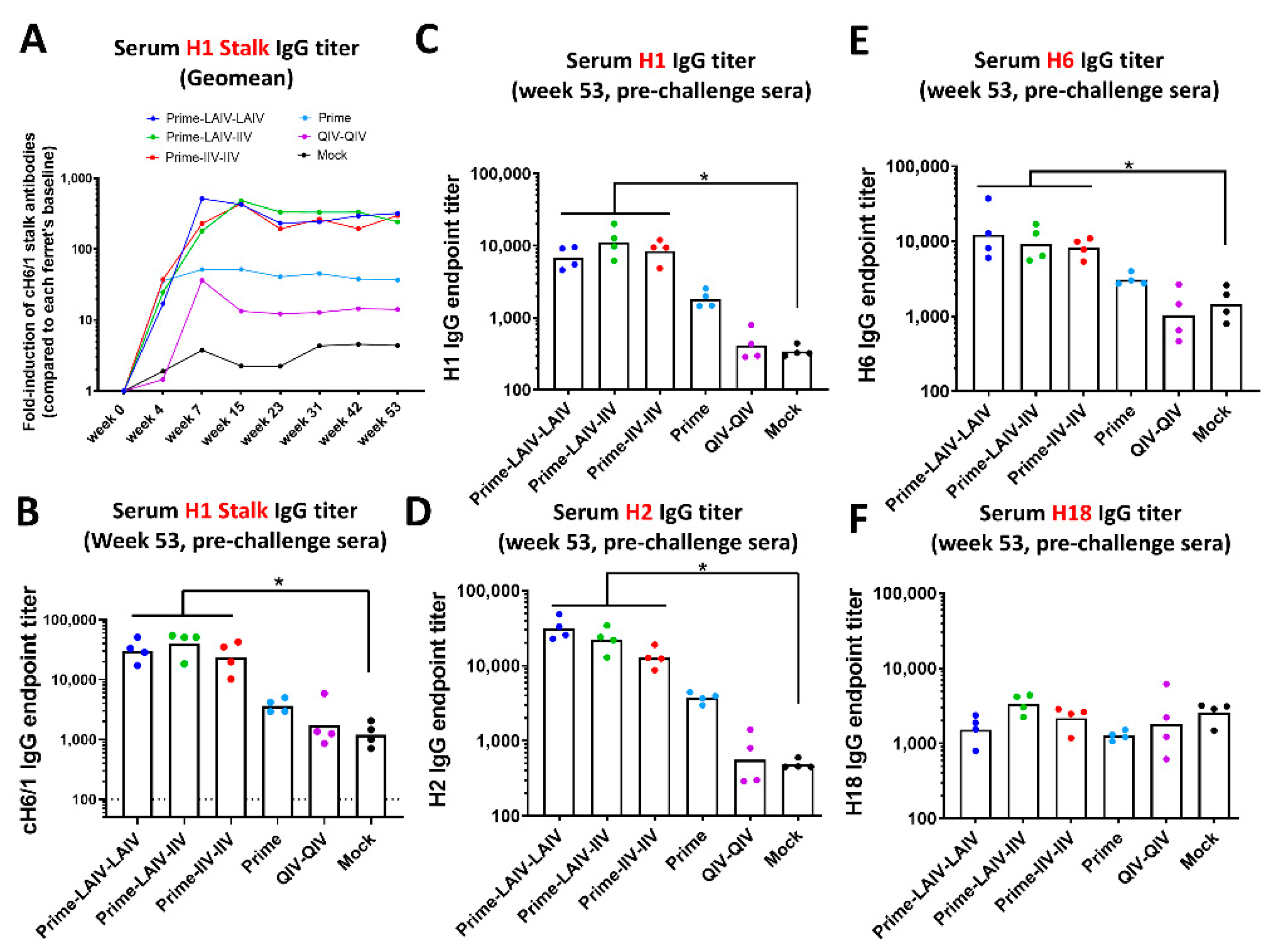
3.1. Chimeric HA-Based Influenza Vaccination Regimens Induce Durable Stalk-Reactive Immunity in Ferrets

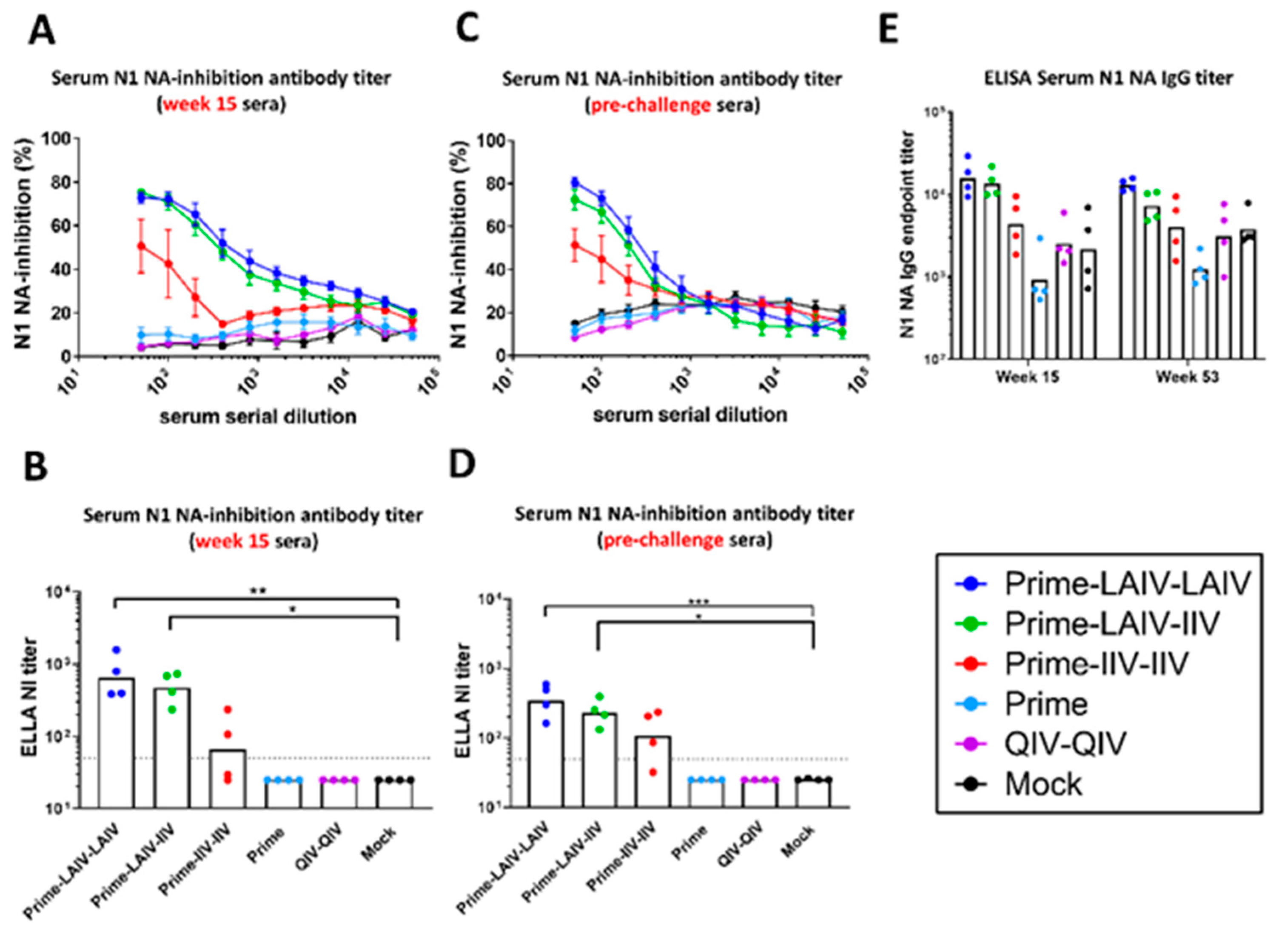
3.2. cHA-Based LAIV-LAIV Vaccine Regimen Induced Long-Lived N1 Neuraminidase-Inhibiting Antibodies
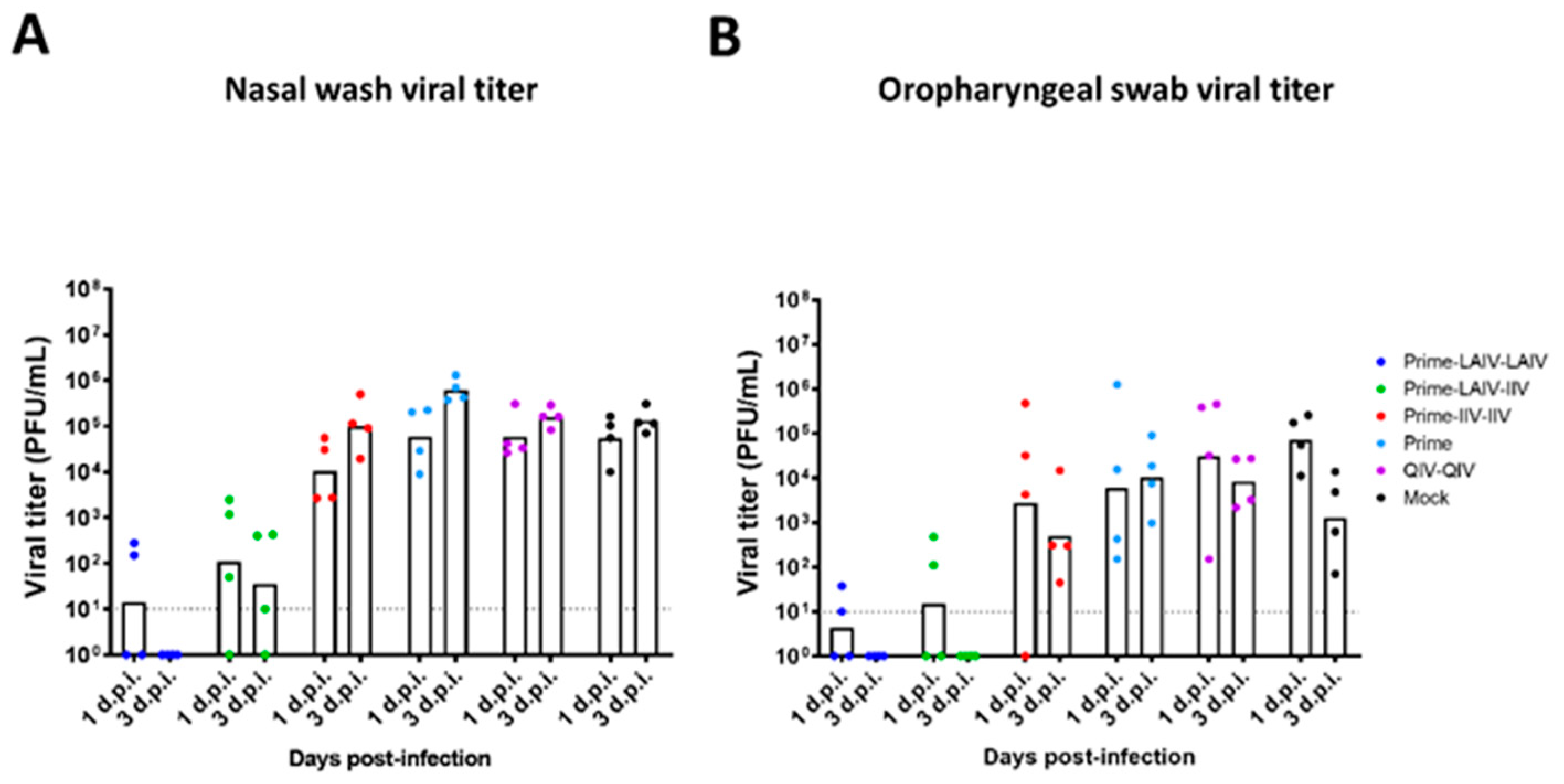
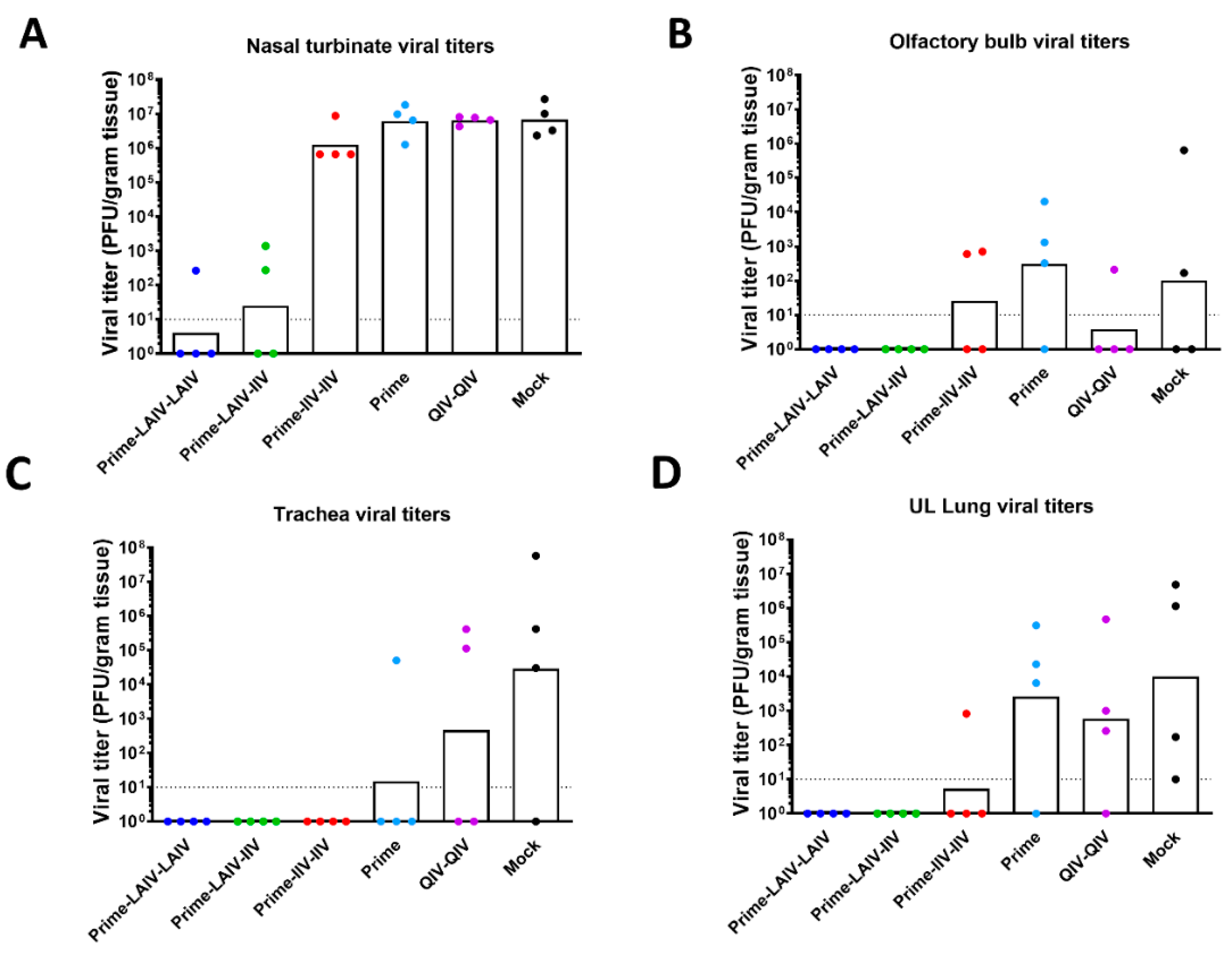
3.3. Vaccination with an LAIV-LAIV Sequential Immunization Regimen Induces Long-Lived Protective Immunity against pH1N1 Viral Challenge
3.4. cHA-Based Vaccination Regimens Stimulated Activation of T Cell Immune Responses in Ferrets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Animal Care and Use Committee Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grohskopf, L.A.; Alyanak, E.; Broder, K.R.; Walter, E.B.; Fry, A.M.; Jernigan, D.B. Prevention and Control of Seasonal Influenza with Vaccines: Recommendations of the Advisory Committee on Immunization Practices—United States, 2019–2020 Influenza Season. MMWR Recomm. Rep. 2019, 68, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, E.J.; Davis, C.T.; Abd Elal, A.I.; Alabi, N.; Azziz-Baumgartner, E.; Barnes, J.; Blanton, L.; Brammer, L.; Budd, A.P.; Burns, E.; et al. Update: Influenza Activity—United States and Worldwide, 20 May to 13 October 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1178–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Blanton, L.; Elal, A.I.A.; Alabi, N.; Barnes, J.; Biggerstaff, M.; Brammer, L.; Budd, A.P.; Burns, E.; Cummings, C.N.; et al. Update: Influenza Activity in the United States During the 2018–2019 Season and Composition of the 2019–2020 Influenza Vaccine. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Yamayoshi, S.; Kawaoka, Y. Current and future influenza vaccines. Nat. Med. 2019, 25, 212–220. [Google Scholar] [CrossRef]
- Belongia, E.A.; Sundaram, M.E.; McClure, D.L.; Meece, J.K.; Ferdinands, J.; VanWormer, J.J. Waning vaccine protection against influenza A (H3N2) illness in children and older adults during a single season. Vaccine 2015, 33, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Ferdinands, J.M.; Fry, A.M.; Reynolds, S.; Petrie, J.; Flannery, B.; Jackson, M.L.; Belongia, E.A. Intraseason waning of influenza vaccine protection: Evidence from the US Influenza Vaccine Effectiveness Network, 2011–2012 through 2014–2015. Clin. Infect. Dis. 2017, 64, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Kissling, E.; Nunes, B.; Robertson, C.; Valenciano, M.; Reuss, A.; Larrauri, A.; Cohen, J.M.; Oroszi, B.; Rizzo, C.; Machado, A.; et al. I-MOVE multicentre case-control study 2010/11 to 2014/15: Is there within-season waning of influenza type/subtype vaccine effectiveness with increasing time since vaccination? Euro. Surveill. 2016, 21. [Google Scholar] [CrossRef] [Green Version]
- Kissling, E.; Valenciano, M.; Larrauri, A.; Oroszi, B.; Cohen, J.M.; Nunes, B.; Pitigoi, D.; Rizzo, C.; Rebolledo, J.; Paradowska-Stankiewicz, I.; et al. Low and decreasing vaccine effectiveness against influenza A(H3) in 2011/12 among vaccination target groups in Europe: Results from the I-MOVE multicentre case-control study. Eur. Surveill. 2013, 18. [Google Scholar] [CrossRef]
- Petrie, J.G.; Ohmit, S.E.; Johnson, E.; Truscon, R.; Monto, A.S. Persistence of Antibodies to Influenza Hemagglutinin and Neuraminidase Following One or Two Years of Influenza Vaccination. J. Infect. Dis. 2015, 212, 1914–1922. [Google Scholar] [CrossRef] [Green Version]
- Petrie, J.G.; Ohmit, S.E.; Truscon, R.; Johnson, E.; Braun, T.M.; Levine, M.Z.; Eichelberger, M.C.; Monto, A.S. Modest Waning of Influenza Vaccine Efficacy and Antibody Titers During the 2007–2008 Influenza Season. J. Infect. Dis. 2016, 214, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Puig-Barbera, J.; Mira-Iglesias, A.; Tortajada-Girbes, M.; Lopez-Labrador, F.X.; Librero-Lopez, J.; Diez-Domingo, J.; Carballido-Fernandez, M.; Carratala-Munuera, C.; Correcher-Medina, P.; Gil-Guillen, V.; et al. Waning protection of influenza vaccination during four influenza seasons, 2011/2012 to 2014/2015. Vaccine 2017, 35, 5799–5807. [Google Scholar] [CrossRef] [PubMed]
- Zelner, J.; Petrie, J.G.; Trangucci, R.; Martin, E.T.; Monto, A.S. Effects of Sequential Influenza A(H1N1)pdm09 Vaccination on Antibody Waning. J. Infect. Dis. 2019, 220, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.; Kondor, R.J.G.; Chung, J.R.; Gaglani, M.; Reis, M.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; et al. Spread of Antigenically Drifted Influenza A(H3N2) Viruses and Vaccine Effectiveness in the United States During the 2018-2019 Season. J. Infect. Dis. 2020, 221, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Chit, A.; Soobiah, C.; Hallett, D.; Meier, G.; Chen, M.H.; Tashkandi, M.; Bauch, C.T.; Loeb, M. Comparing influenza vaccine efficacy against mismatched and matched strains: A systematic review and meta-analysis. BMC Med. 2013, 11, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdil, C. The annual production cycle for influenza vaccine. Vaccine 2003, 21, 1776–1779. [Google Scholar] [CrossRef]
- Houser, K.; Subbarao, K. Influenza vaccines: Challenges and solutions. Cell Host. Microbe. 2015, 17, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Hampson, A.; Barr, I.; Cox, N.; Donis, R.O.; Siddhivinayak, H.; Jernigan, D.; Katz, J.; McCauley, J.; Motta, F.; Odagiri, T.; et al. Improving the selection and development of influenza vaccine viruses—Report of a WHO informal consultation on improving influenza vaccine virus selection, Hong Kong SAR, China, 18-20 November 2015. Vaccine 2017, 35, 1104–1109. [Google Scholar] [CrossRef]
- Weir, J.P.; Gruber, M.F. An overview of the regulation of influenza vaccines in the United States. Influenza Other Respir. Viruses 2016, 10, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Erbelding, E.J.; Post, D.J.; Stemmy, E.J.; Roberts, P.C.; Augustine, A.D.; Ferguson, S.; Paules, C.I.; Graham, B.S.; Fauci, A.S. A Universal Influenza Vaccine: The Strategic Plan for the National Institute of Allergy and Infectious Diseases. J. Infect. Dis. 2018, 218, 347–354. [Google Scholar] [CrossRef]
- Ortiz, J.R.; Hickling, J.; Jones, R.; Donabedian, A.; Engelhardt, O.G.; Katz, J.M.; Madhi, S.A.; Neuzil, K.M.; Rimmelzwaan, G.F.; Southern, J.; et al. Report on eighth WHO meeting on development of influenza vaccines that induce broadly protective and long-lasting immune responses: Chicago, USA, 23–24 August 2016. Vaccine 2018, 36, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.I.; Marston, H.D.; Eisinger, R.W.; Baltimore, D.; Fauci, A.S. The Pathway to a Universal Influenza Vaccine. Immunity 2017, 47, 599–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paules, C.I.; Sullivan, S.G.; Subbarao, K.; Fauci, A.S. Chasing Seasonal Influenza—The Need for a Universal Influenza Vaccine. N. Engl. J. Med. 2018, 378, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Boonnak, K.; Matsuoka, Y.; Wang, W.; Suguitan, A.L., Jr.; Chen, Z.; Paskel, M.; Baz, M.; Moore, I.; Jin, H.; Subbarao, K. Development of Clade-Specific and Broadly Reactive Live Attenuated Influenza Virus Vaccines against Rapidly Evolving H5 Subtype Viruses. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, L.; Deng, W.; Bao, L.; Lv, Q.; Ma, C.; Li, F.; Xu, L.; Qin, C. Higher Immunological Protection of Pandemic 2009 H1N1 Influenza Live Virus Infection than Split Vaccine Against the Homologous Virus for Long Term Immunization in Ferret. Indian J. Virol. 2012, 23, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Elbahesh, H.; Saletti, G.; Gerlach, T.; Rimmelzwaan, G.F. Broadly protective influenza vaccines: Design and production platforms. Curr. Opin. Virol. 2019, 34, 1–9. [Google Scholar] [CrossRef]
- Estrada, L.D.; Schultz-Cherry, S. Development of a Universal Influenza Vaccine. J. Immunol. 2019, 202, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, L.; Palese, P. Overcoming Barriers in the Path to a Universal Influenza Virus Vaccine. Cell Host. Microbe 2018, 24, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Hai, R.; Krammer, F.; Tan, G.S.; Pica, N.; Eggink, D.; Maamary, J.; Margine, I.; Albrecht, R.A.; Palese, P. Influenza viruses expressing chimeric hemagglutinins: Globular head and stalk domains derived from different subtypes. J. Virol. 2012, 86, 5774–5781. [Google Scholar] [CrossRef] [Green Version]
- Krammer, F.; Hai, R.; Yondola, M.; Tan, G.S.; Leyva-Grado, V.H.; Ryder, A.B.; Miller, M.S.; Rose, J.K.; Palese, P.; Garcia-Sastre, A.; et al. Assessment of influenza virus hemagglutinin stalk-based immunity in ferrets. J. Virol. 2014, 88, 3432–3442. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Nachbagauer, R.; Stadlbauer, D.; Solorzano, A.; Berlanda-Scorza, F.; Garcia-Sastre, A.; Palese, P.; Krammer, F.; Albrecht, R.A. Sequential Immunization with Live-Attenuated Chimeric Hemagglutinin-Based Vaccines Confers Heterosubtypic Immunity Against Influenza A Viruses in a Preclinical Ferret Model. Front. Immunol. 2019, 10, 756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachbagauer, R.; Krammer, F.; Albrecht, R.A. A Live-Attenuated Prime, Inactivated Boost Vaccination Strategy with Chimeric Hemagglutinin-Based Universal Influenza Virus Vaccines Provides Protection in Ferrets: A Confirmatory Study. Vaccines 2018, 6, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachbagauer, R.; Liu, W.C.; Choi, A.; Wohlbold, T.J.; Atlas, T.; Rajendran, M.; Solorzano, A.; Berlanda-Scorza, F.; Garcia-Sastre, A.; Palese, P.; et al. A universal influenza virus vaccine candidate confers protection against pandemic H1N1 infection in preclinical ferret studies. NPJ. Vaccines 2017, 2, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachbagauer, R.; Miller, M.S.; Hai, R.; Ryder, A.B.; Rose, J.K.; Palese, P.; Garcia-Sastre, A.; Krammer, F.; Albrecht, R.A. Hemagglutinin Stalk Immunity Reduces Influenza Virus Replication and Transmission in Ferrets. J. Virol. 2015, 90, 3268–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isakova-Sivak, I.; Matyushenko, V.; Kotomina, T.; Kiseleva, I.; Krutikova, E.; Donina, S.; Rekstin, A.; Larionova, N.; Mezhenskaya, D.; Sivak, K.; et al. Sequential Immunization with Universal Live Attenuated Influenza Vaccine Candidates Protects Ferrets against a High-Dose Heterologous Virus Challenge. Vaccines 2019, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Hurt, A.C. Using the Ferret as an Animal Model for Investigating Influenza Antiviral Effectiveness. Front. Microbiol. 2016, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Margine, I.; Palese, P.; Krammer, F. Expression of functional recombinant hemagglutinin and neuraminidase proteins from the novel H7N9 influenza virus using the baculovirus expression system. J. Vis. Exp. 2013, e51112. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhu, X.; Dwek, R.A.; Stevens, J.; Wilson, I.A. Structural characterization of the 1918 influenza virus H1N1 neuraminidase. J. Virol. 2008, 82, 10493–10501. [Google Scholar] [CrossRef] [Green Version]
- Nachbagauer, R.; Kinzler, D.; Choi, A.; Hirsh, A.; Beaulieu, E.; Lecrenier, N.; Innis, B.L.; Palese, P.; Mallett, C.P.; Krammer, F. A chimeric haemagglutinin-based influenza split virion vaccine adjuvanted with AS03 induces protective stalk-reactive antibodies in mice. NPJ. Vaccines 2016, 1. [Google Scholar] [CrossRef]
- Krammer, F.; Pica, N.; Hai, R.; Margine, I.; Palese, P. Chimeric hemagglutinin influenza virus vaccine constructs elicit broadly protective stalk-specific antibodies. J. Virol. 2013, 87, 6542–6550. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Nachbagauer, R.; Krammer, F.; Albrecht, R.A. Assessment of Influenza Virus Hemagglutinin Stalk-Specific Antibody Responses. Methods Mol. Biol. 2018, 1836, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Eckert, A.M.; Tumpey, T.M.; Maines, T.R. Complexities in Ferret Influenza Virus Pathogenesis and Transmission Models. Microbiol Mol. Biol. Rev. 2016, 80, 733–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, J.A.; Katz, J.M.; Tumpey, T.M. The ferret as a model organism to study influenza A virus infection. Dis. Model. Mech. 2011, 4, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Hoa, L.N.M.; Mai, L.Q.; Bryant, J.E.; Thai, P.Q.; Hang, N.L.K.; Yen, N.T.T.; Duong, T.N.; Thoang, D.D.; Horby, P.; Werheim, H.F.L.; et al. Association between Hemagglutinin Stem-Reactive Antibodies and Influenza A/H1N1 Virus Infection during the 2009 Pandemic. J. Virol. 2016, 90, 6549–6556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, J.; Sheehan, J.; Hwang, W.C.; Bankston, L.A.; Burchett, S.K.; Huang, C.Y.; Liddington, R.C.; Beigel, J.H.; Marasco, W.A. Wide prevalence of heterosubtypic broadly neutralizing human anti-influenza A antibodies. Clin. Infect. Dis. 2011, 52, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- Leroux-Roels, I.; Roman, F.; Forgus, S.; Maes, C.; De Boever, F.; Drame, M.; Gillard, P.; van der Most, R.; Van Mechelen, M.; Hanon, E.; et al. Priming with AS03 A-adjuvanted H5N1 influenza vaccine improves the kinetics, magnitude and durability of the immune response after a heterologous booster vaccination: An open non-randomised extension of a double-blind randomised primary study. Vaccine 2010, 28, 849–857. [Google Scholar] [CrossRef]
- Bernstein, D.I.; Guptill, J.; Naficy, A.; Nachbagauer, R.; Berlanda-Scorza, F.; Feser, J.; Wilson, P.C.; Solorzano, A.; Van der Wielen, M.; Walter, E.B.; et al. Immunogenicity of chimeric haemagglutinin-based, universal influenza virus vaccine candidates: Interim results of a randomised, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis. 2020, 20, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Jansen, J.M.; Gerlach, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Influenza virus-specific CD4+ and CD8+ T cell-mediated immunity induced by infection and vaccination. J. Clin. Virol. 2019, 119, 44–52. [Google Scholar] [CrossRef]
- Choi, A.; Bouzya, B.; Cortes Franco, K.D.; Stadlbauer, D.; Rajabhathor, A.; Rouxel, R.N.; Mainil, R.; Van der Wielen, M.; Palese, P.; Garcia-Sastre, A.; et al. Chimeric Hemagglutinin-Based Influenza Virus Vaccines Induce Protective Stalk-Specific Humoral Immunity and Cellular Responses in Mice. Immunohorizons 2019, 3, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.M.; Li, O.T.W.; Poh, C.M.; Perera, R.; Valkenburg, S.A.; Peiris, M.; Poon, L.L.M. Combined use of live-attenuated and inactivated influenza vaccines to enhance heterosubtypic protection. Virology 2018, 525, 73–82. [Google Scholar] [CrossRef]
- Isakova-Sivak, I.; Korenkov, D.; Smolonogina, T.; Kotomina, T.; Donina, S.; Matyushenko, V.; Mezhenskaya, D.; Krammer, F.; Rudenko, L. Broadly protective anti-hemagglutinin stalk antibodies induced by live attenuated influenza vaccine expressing chimeric hemagglutinin. Virology 2018, 518, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Feser, J.; Naficy, A.; Bernstein, D.I.; Guptill, J.; Walter, E.B.; Berlanda-Scorza, F.; Stadlbauer, D.; Wilson, P.C.; Aydillo, T.; et al. A chimeric hemagglutinin-based universal influenza virus vaccine approach induces broad and long-lasting immunity in a randomized, placebo-controlled phase I trial. Nat. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Choi, A.; Hirsh, A.; Margine, I.; Iida, S.; Barrera, A.; Ferres, M.; Albrecht, R.A.; Garcia-Sastre, A.; Bouvier, N.M.; et al. Defining the antibody cross-reactome directed against the influenza virus surface glycoproteins. Nat. Immunol. 2017, 18, 464–473. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-C.; Nachbagauer, R.; Stadlbauer, D.; Strohmeier, S.; Solórzano, A.; Berlanda-Scorza, F.; Innis, B.L.; García-Sastre, A.; Palese, P.; Krammer, F.; et al. Chimeric Hemagglutinin-Based Live-Attenuated Vaccines Confer Durable Protective Immunity against Influenza A Viruses in a Preclinical Ferret Model. Vaccines 2021, 9, 40. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9010040
Liu W-C, Nachbagauer R, Stadlbauer D, Strohmeier S, Solórzano A, Berlanda-Scorza F, Innis BL, García-Sastre A, Palese P, Krammer F, et al. Chimeric Hemagglutinin-Based Live-Attenuated Vaccines Confer Durable Protective Immunity against Influenza A Viruses in a Preclinical Ferret Model. Vaccines. 2021; 9(1):40. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9010040
Chicago/Turabian StyleLiu, Wen-Chun, Raffael Nachbagauer, Daniel Stadlbauer, Shirin Strohmeier, Alicia Solórzano, Francesco Berlanda-Scorza, Bruce L. Innis, Adolfo García-Sastre, Peter Palese, Florian Krammer, and et al. 2021. "Chimeric Hemagglutinin-Based Live-Attenuated Vaccines Confer Durable Protective Immunity against Influenza A Viruses in a Preclinical Ferret Model" Vaccines 9, no. 1: 40. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9010040