
Development of a Viral-Like Particle Candidate Vaccine Against Novel Variant Infectious Bursal Disease Virus

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Animals
2.3. Construction of Recombinant Expression Plasmid
2.4. Expression and Purification of SHG19-VP2-466
2.5. Analysis of SDS-PAGE and Western-Blot
2.6. Transmission Electron Microscopy (TEM)
2.7. Agar-Gel Precipitation (AGP)
2.8. Preparation of SHG19-VLP Vaccine
2.9. Evaluation of the Immune Effect of SHG19-VLP Vaccine Against nVarIBDV
2.10. Evaluation of Immune Effect of SHG19-VLP Vaccine Against vvIBDV
2.11. Virus-Neutralization Assay
2.12. Statistical Analyses
3. Results
3.1. Expression, Purification, and Identification of SHG19-VLP
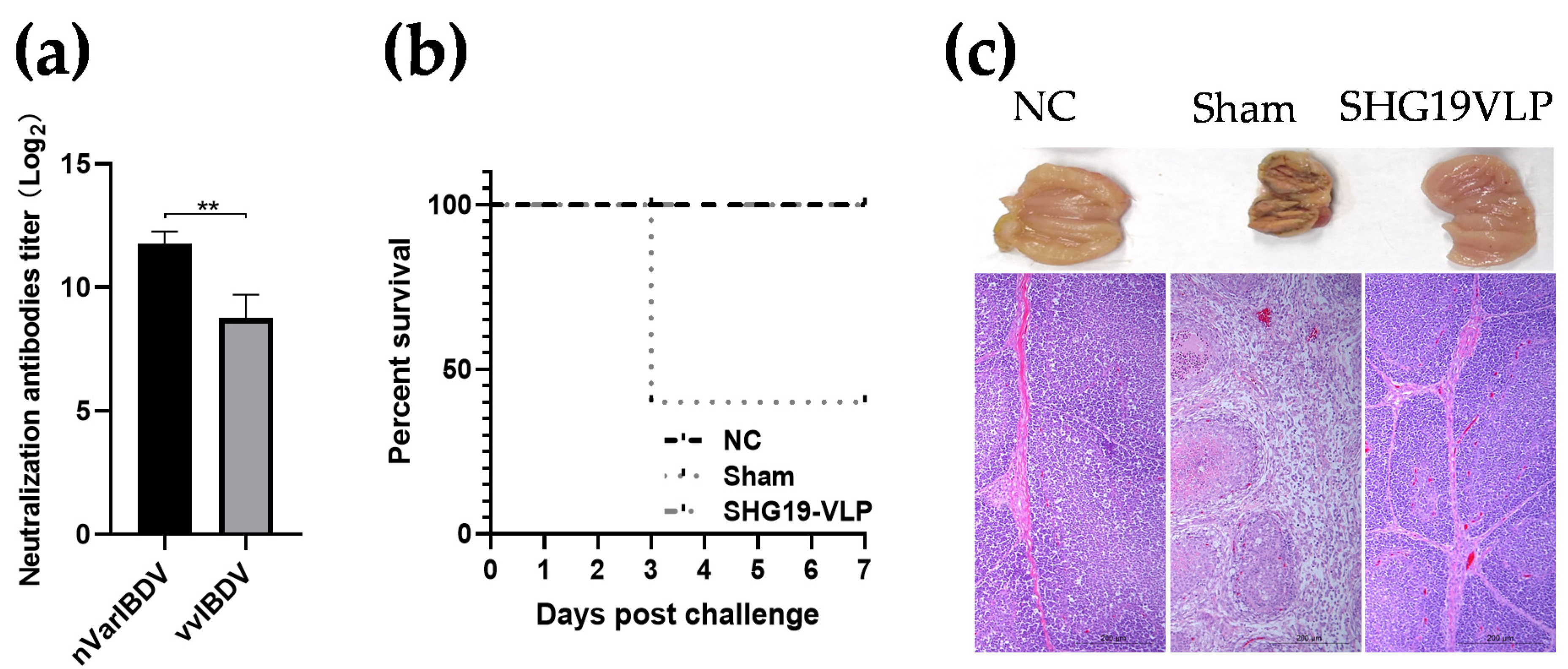
3.2. SHG19-VLP Vaccine Provided Protection against nVarIBDV Challenge
3.3. SHG19-VLP Vaccine Induced Protection against vvIBDV Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Müller, H.; Islam, M.R.; Raue, R. Research on infectious bursal disease—the past, the present and the future. Vet. Microbiol. 2003, 97, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.D.; Skinner, M.A. Coding sequences of both genome segments of a European ‘very virulent’ infectious bursal disease virus. Virus Res. 1996, 40, 1–15. [Google Scholar] [CrossRef]
- Lombardo, E.; Maraver, A.; Espinosa, I.; Fernandez-Arias, A.; Rodriguez, J.F. VP5, the nonstructural polypeptide of infectious bursal disease virus, accumulates within the host plasma membrane and induces cell lysis. Virology 2000, 277, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Petit, S.; Lejal, N.; Huet, J.C.; Delmas, B. Active residues and viral substrate cleavage sites of the protease of the birnavirus infectious pancreatic necrosis virus. J. Virol. 2000, 74, 2057–2066. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, A.B.; Rodriguez, J.F. Proteolytic processing in infectious bursal disease virus: Identification of the polyprotein cleavage sites by site-directed mutagenesis. Virology 1999, 262, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, C.; Lepault, J.; Erk, I.; Da Costa, B.; Delmas, B. The maturation process of pVP2 requires assembly of infectious bursal disease virus capsids. J. Virol. 2002, 76, 2384–2392. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Pathak, D.C.; Ramamurthy, N.; Maity, H.K.; Chellappa, M.M. Infectious bursal disease virus in chickens: Prevalence, impact, and management strategies. Vet. Med. 2019, 10, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackwood, D.J. Advances in vaccine research against economically important viral diseases of food animals: Infectious bursal disease virus. Vet. Microbiol. 2017, 206, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Letzel, T.; Coulibaly, F.; Rey, F.A.; Delmas, B.; Jagt, E.; Van Loon, A.A.; Mundt, E. Molecular and structural bases for the antigenicity of VP2 of infectious bursal disease virus. J. Virol. 2007, 81, 12827–12835. [Google Scholar] [CrossRef] [Green Version]
- Brandt, M.; Yao, K.; Liu, M.; Heckert, R.A.; Vakharia, V.N. Molecular determinants of virulence, cell tropism, and pathogenic phenotype of infectious bursal disease virus. J. Virol. 2001, 75, 11974–11982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackwood, D.J.; Sreedevi, B.; LeFever, L.J.; Sommer-Wagner, S.E. Studies on naturally occurring infectious bursal disease viruses suggest that a single amino acid substitution at position 253 in VP2 increases pathogenicity. Virology 2008, 377, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, C.; Wang, Y.; Zhang, E.; Han, X.; Yu, Z.; Liu, H. VP1 and VP3 are required and sufficient for translation initiation of uncapped infectious bursal disease virus genomic double-stranded RNA. J. Virol. 2018, 92, e01345-17. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Shi, L.; Zhang, Y.; Peng, X.; Zheng, T.; Li, Y.; Hu, B.; Zheng, X.; Zhou, J. Ubiquitination is essential for Avibirnavirus replication by supporting VP1 polymerase activity. J. Virol. 2019, 93, e01899-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardo, E.; Maraver, A.; Castón, J.R.; Rivera, J.; Fernández-Arias, A.; Serrano, A.; Carrascosa, J.L.; Rodriguez, J.F. VP1, the putative RNA-dependent RNA polymerase of infectious bursal disease virus, forms complexes with the capsid protein VP3, leading to efficient encapsidation into virus-like particles. J. Virol. 1999, 73, 6973–6983. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, Y.; Ye, C. Rapid generation of attenuated infectious bursal disease virus from dual-promoter plasmids by reduction of viral ribonucleoprotein activity. J. Virol. 2020, 94, e01569. [Google Scholar] [CrossRef]
- Cosgrove, A.S. An apparently new disease of chickens: Avian nephrosis. Avian Dis. 1962, 6, 385–389. [Google Scholar] [CrossRef]
- Jackwood, D.H.; Saif, Y.M. Antigenic diversity of infectious bursal disease viruses. Avian Dis. 1987, 31, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Chettle, N.; Stuart, J.C.; Wyeth, P.J. Outbreak of virulent infectious bursal disease in East Anglia. Vet. Rec. 1989, 125, 271–272. [Google Scholar] [CrossRef]
- Prabhu, S.N.; Singh, A.P.; Varghese, B.P.; Dhama, K.; Singh, S.D.; Singh, R. A comparative study of pathology and host immune response induced by very virulent infectious bursal disease virus in experimentally infected chickens of Aseel and white Leghorn breeds. Vaccines 2020, 8, 627. [Google Scholar] [CrossRef]
- Fan, L.; Wu, T.; Hussain, A.; Gao, Y.; Zeng, X.; Wang, Y.; Gao, L.; Li, K.; Wang, Y.; Liu, C. Novel variant strains of infectious bursal disease virus isolated in China. Vet. Microbiol. 2019, 230, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Pei, Y.; Zhang, K.; Xue, J.; Ruan, S.; Zhang, G. Phylogenetic analyses and pathogenicity of a variant infectious bursal disease virus strain isolated in China. Virus Res. 2019, 276, 197833. [Google Scholar] [CrossRef]
- Fan, L.; Wu, T.; Wang, Y.; Hussain, A.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Liu, C.; Cui, H.; et al. Novel variants of infectious bursal disease virus can severely damage the bursa of fabricius of immunized chickens. Vet. Microbiol. 2020, 240, 108507. [Google Scholar] [CrossRef]
- Myint, O.; Suwanruengsri, M.; Araki, K.; Izzati, U.Z.; Pornthummawat, A.; Nueangphuet, P.; Fuke, N.; Hirai, T.; Jackwood, D.J.; Yamaguchi, R. The bursa atrophy at 28 days old by the variant infectious bursal disease virus makes a negative economic impact on broiler farms in Japan. Avian Pathol. 2020, 50, 1–41. [Google Scholar] [CrossRef]
- Muller, H.; Mundt, E.; Eterradossi, N.; Islam, M.R. Current status of vaccines against infectious bursal disease. Avian Pathol. 2012, 41, 133–139. [Google Scholar] [CrossRef]
- Li, G.; Kuang, H.; Guo, H.; Cai, L.; Chu, D.; Wang, X.; Hu, J.; Rong, J. Development of a recombinant VP2 vaccine for the prevention of novel variant strains of infectious bursal disease virus. Avian Pathol. 2020, 49, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Gao, L.; Qin, L.; Deng, X.; Wu, G.; Zhang, L.; Yu, F.; Ren, X.; Gao, Y.; Gao, H.; et al. Genomic sequencing and molecular characteristics of a very virulent strain of infectious bursal disease virus isolated in China. Agric. Sci. Technol. 2011, 12, 1946–1949. [Google Scholar]
- Fan, L.; Wang, Y.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Cui, H.; Pan, Q.; Liu, C.; Zhang, Y.; et al. A reassortment vaccine candidate of the novel variant infectious bursal disease virus. Vet. Microbiol. 2020, 251, 108905. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Qi, X.; Li, K.; Gao, H.; Gao, Y.; Qin, L.; Wang, Y.; Wang, X. Development of a tailored vaccine against challenge with very virulent infectious bursal disease virus of chickens using reverse genetics. Vaccine 2011, 29, 5550–5557. [Google Scholar] [CrossRef]
- Saugar, I.; Irigoyen, N.; Luque, D.; Carrascosa, J.L.; Rodríguez, J.F.; Castón, J.R. Electrostatic interactions between capsid and scaffolding proteins mediate the structural polymorphism of a double-stranded RNA virus. J. Biol. Chem. 2010, 285, 3643–3650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, S.; Chellappa, M.M.; Pathak, D.C.; Gaikwad, S.; Yadav, K.; Ramakrishnan, S.; Vakharia, V.N. Newcastle disease virus vectored bivalent vaccine against virulent infectious bursal disease and Newcastle disease of chickens. Vaccines 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Wang, Y.; Li, H.; Fan, L.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Liu, C.; Cui, H.; et al. Naturally occurring homologous recombination between novel variant infectious bursal disease virus and intermediate vaccine strain. Vet. Microbiol. 2020, 245, 108700. [Google Scholar] [CrossRef]
- Huo, S.; Zhang, J.; Fan, J.; Wang, X.; Wu, F.; Zuo, Y.; Zhong, F. Co-expression of chicken IL-2 and IL-7 enhances the immunogenicity and protective efficacy of a VP2-eexpressing DNA vaccine against IBDV in chickens. Viruses 2019, 11, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Pan, Q.; Lu, Z.; Li, K.; Gao, H.; Qi, X.; Gao, Y.; Wang, X. An optimized, highly efficient, self-assembled, subvirus-like particle of infectious bursal disease virus (IBDV). Vaccine 2016, 34, 3508–3514. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Durairaj, V.; Mundt, E.; Schulze, K.; Breunig, K.D.; Behrens, S.E. Protective vaccination against infectious bursal disease virus with whole recombinant Kluyveromyces lactis yeast expressing the viral VP2 subunit. PLoS ONE 2012, 7, e42870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackwood, D.J. Multivalent virus-like–particle vaccine protects against classic and variant infectious bursal disease viruses. Avian Dis. 2013, 57, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lucero, M.S.; Richetta, M.; Chimeno Zoth, S.; Jaton, J.; Pinto, S.; Canet, Z.; Berinstein, A.; Gómez, E. Plant-based vaccine candidate against Infectious bursal disease: An alternative to inactivated vaccines for breeder hens. Vaccine 2019, 37, 5203–5210. [Google Scholar] [CrossRef]
- Rage, E.; Drissi Touzani, C.; Marusic, C.; Lico, C.; Göbel, T.; Bortolami, A.; Bonfante, F.; Salzano, A.M.; Scaloni, A.; Fellahi, S.; et al. Functional characterization of a plant-produced infectious bursal disease virus antigen fused to the constant region of avian IgY immunoglobulins. Appl. Microbiol. Biotechnol. 2019, 103, 7491–7504. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Liu, Y.; Wang, A.; Zhang, G.; Yang, G.; Chen, Y.; Ji, P.; Liu, C.; Song, Y.; Su, Y.; et al. High level soluble expression and one-step purification of IBDV VP2 protein in Escherichia coli. Biotechnol. Lett. 2016, 38, 901–908. [Google Scholar] [CrossRef]
- Mustafa, S.; Abd-Aziz, N.; Saw, W.T.; Liew, S.Y.; Yusoff, K.; Shafee, N. Recombinant enterovirus 71 viral protein 1 fused to a truncated Newcastle disease virus NP (NPt) carrier protein. Vaccines 2020, 8, 742. [Google Scholar] [CrossRef] [PubMed]
- Feodorova, V.A.; Lyapina, A.M.; Zaitsev, S.S.; Khizhnyakova, M.A.; Sayapina, L.V.; Ulianova, O.V.; Ulyanov, S.S.; Motin, V.L. New promising targets for synthetic Omptin-based peptide vaccine against gram-negative pathogens. Vaccines 2019, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Irigoyen, N.; Castón, J.R.; Rodríguez, J.F. Host proteolytic activity is necessary for infectious bursal disease virus capsid protein assembly. J. Biol. Chem. 2012, 287, 24473–24482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saugar, I.; Luque, D.; Ona, A.; Rodriguez, J.F.; Carrascosa, J.L.; Trus, B.L.; Caston, J.R. Structural polymorphism of the major capsid protein of a double-stranded RNA virus: An amphipathic alpha helix as a molecular switch. Structure 2005, 13, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eterradossi, N.; Saif, Y.M. Infectious Bursal Disease. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S., Grimes, T., Johnson, D., Kromm, M., Eds.; John Wiley & Sons, Inc: Hoboken, USA, 2020; Volume 1, pp. 257–283. [Google Scholar]
- Ismail, N.M.; Saif, Y.M. Immunogenicity of infectious bursal disease viruses in chickens. Avian Dis. 1991, 35, 460–469. [Google Scholar] [CrossRef]
- He, X.; Wang, W.; Chen, G.; Jiao, P.; Ji, Z.; Yang, L.; Wei, P. Serological study reveal different antigenic IBDV strains prevalent in southern China during the years 2000–2017 and also the antigenic differences between the field strains and the commonly used vaccine strains. Vet. Microbiol. 2019, 239, 108458. [Google Scholar] [CrossRef]
- Le, X.T.K.; Doan, H.T.T.; Do, R.T.; Le, T.H. Molecular characterization of field isolates of infectious bursal disease virus from three decades, 1987-2018, reveals a distinct genotypic subgroup in Vietnam. Arch. Virol. 2019, 164, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Boudaoud, A.; Mamache, B.; Tombari, W.; Ghram, A. Virus mutations and their impact on vaccination against infectious bursal disease (Gumboro disease). Rev. Sci. Tech. 2016, 35, 875–897. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Jiang, N.; Fan, L.; Gao, L.; Li, K.; Gao, Y.; Niu, X.; Zhang, W.; Cui, H.; Liu, A.; et al. Development of a Viral-Like Particle Candidate Vaccine Against Novel Variant Infectious Bursal Disease Virus. Vaccines 2021, 9, 142. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020142
Wang Y, Jiang N, Fan L, Gao L, Li K, Gao Y, Niu X, Zhang W, Cui H, Liu A, et al. Development of a Viral-Like Particle Candidate Vaccine Against Novel Variant Infectious Bursal Disease Virus. Vaccines. 2021; 9(2):142. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020142
Chicago/Turabian StyleWang, Yulong, Nan Jiang, Linjin Fan, Li Gao, Kai Li, Yulong Gao, Xinxin Niu, Wenying Zhang, Hongyu Cui, Aijing Liu, and et al. 2021. "Development of a Viral-Like Particle Candidate Vaccine Against Novel Variant Infectious Bursal Disease Virus" Vaccines 9, no. 2: 142. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020142