
Protection against Different Genotypes of Newcastle Disease Viruses (NDV) Afforded by an Adenovirus-Vectored Fusion Protein and Live NDV Vaccines in Chickens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chickens
2.2. Viruses
2.3. Adenovirus Construct
2.4. Immunofluorescent Assay (IFA)
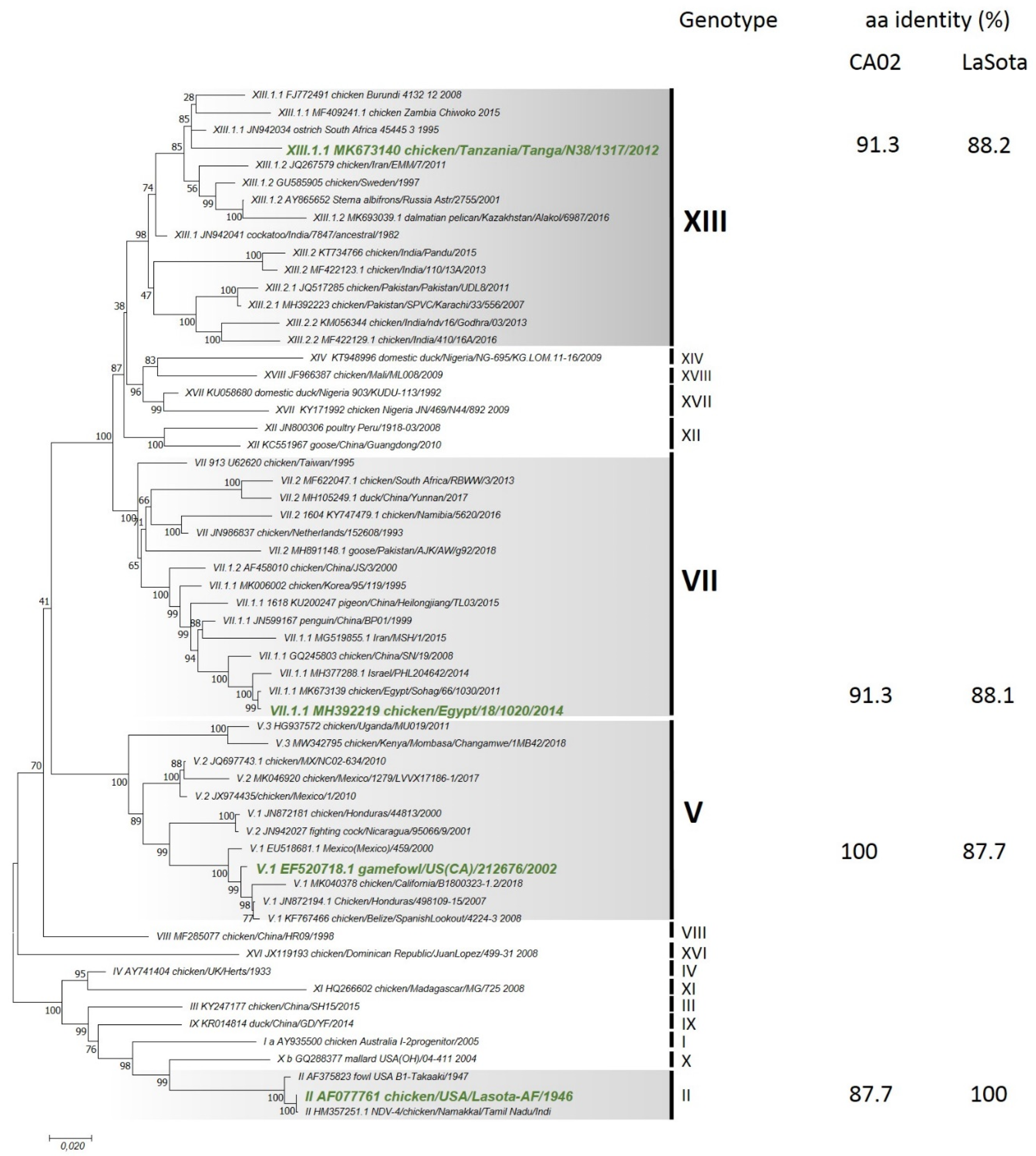
2.5. Phylogenetic Analysis
2.6. Experimental Design
2.7. Virus Shedding
2.8. Hemagglutination Inhibition (HI) Test
2.9. Indirect ELISA for Detection of Antibodies Specific to NDV
2.10. Statistical Analysis
3. Results
3.1. IFA
3.2. Phylogenetic Analysis
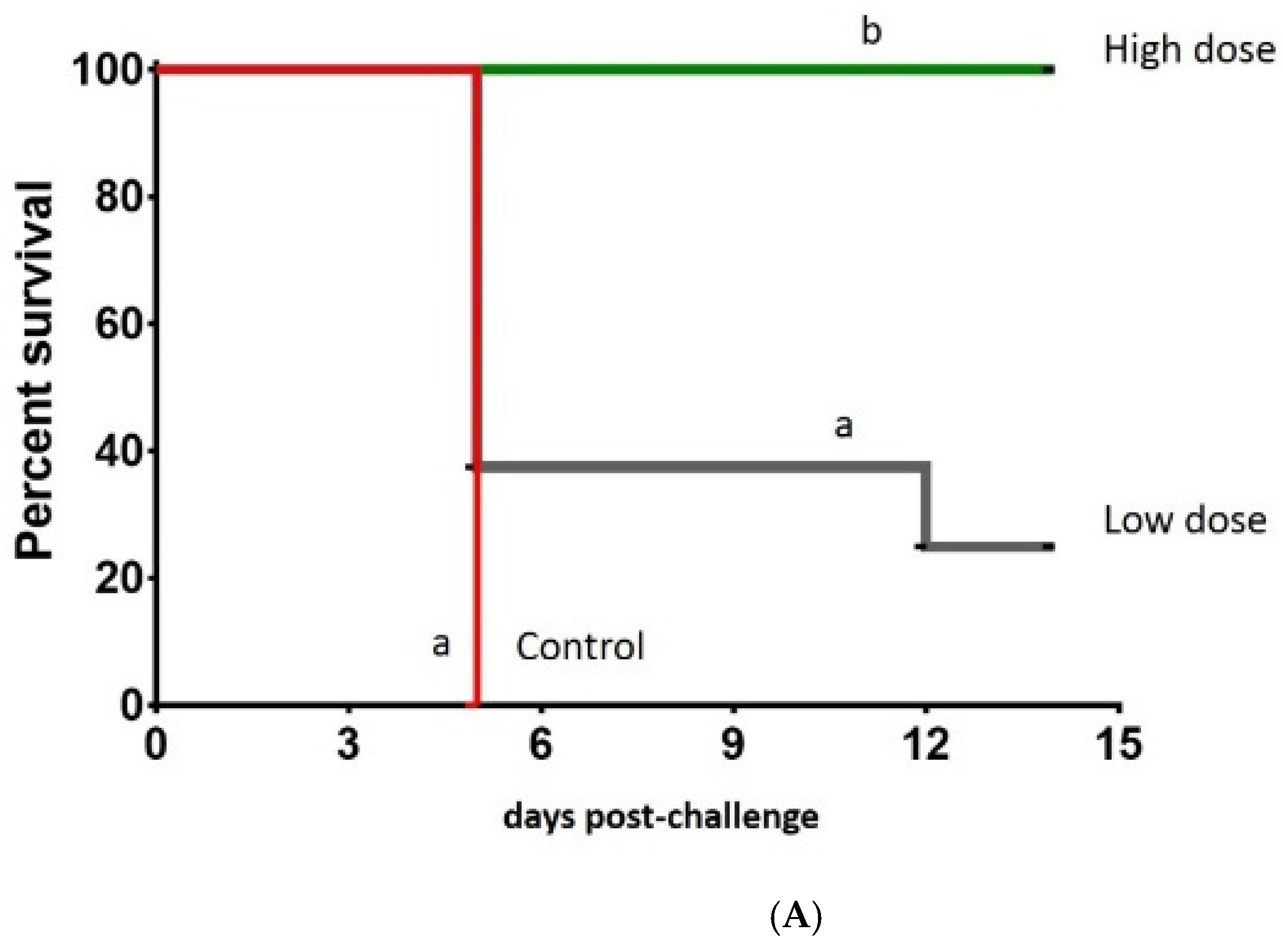
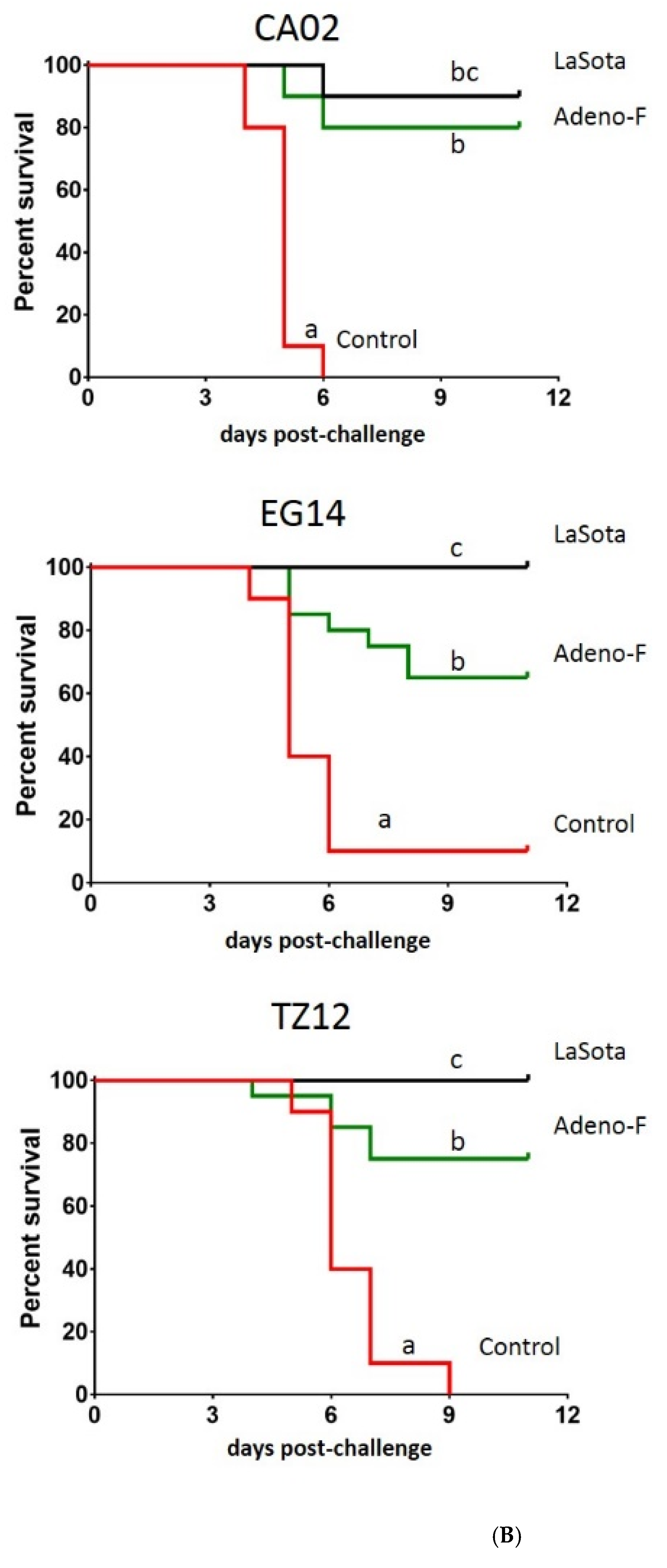
3.3. Clinical Protection
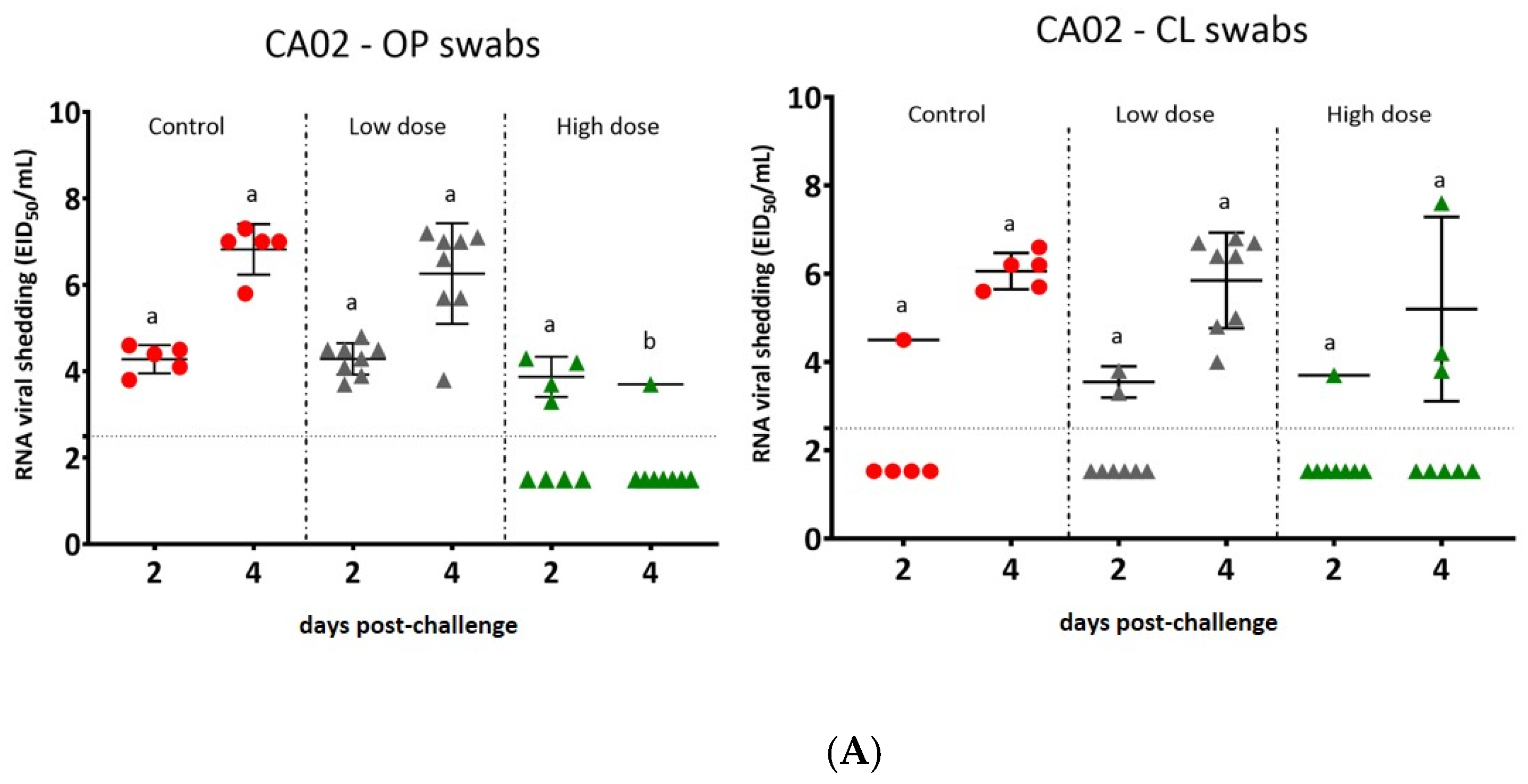
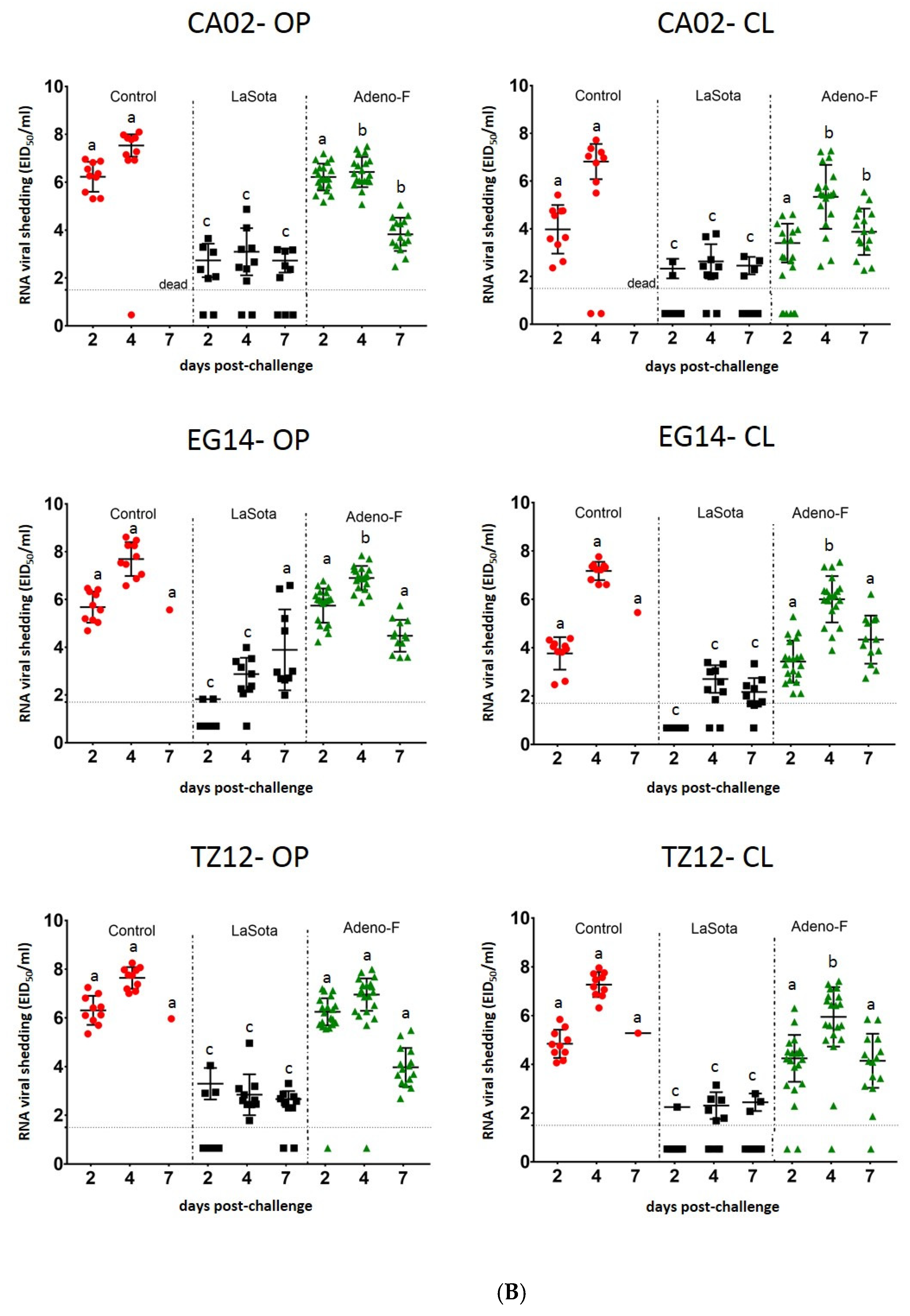
3.4. Virus Shedding
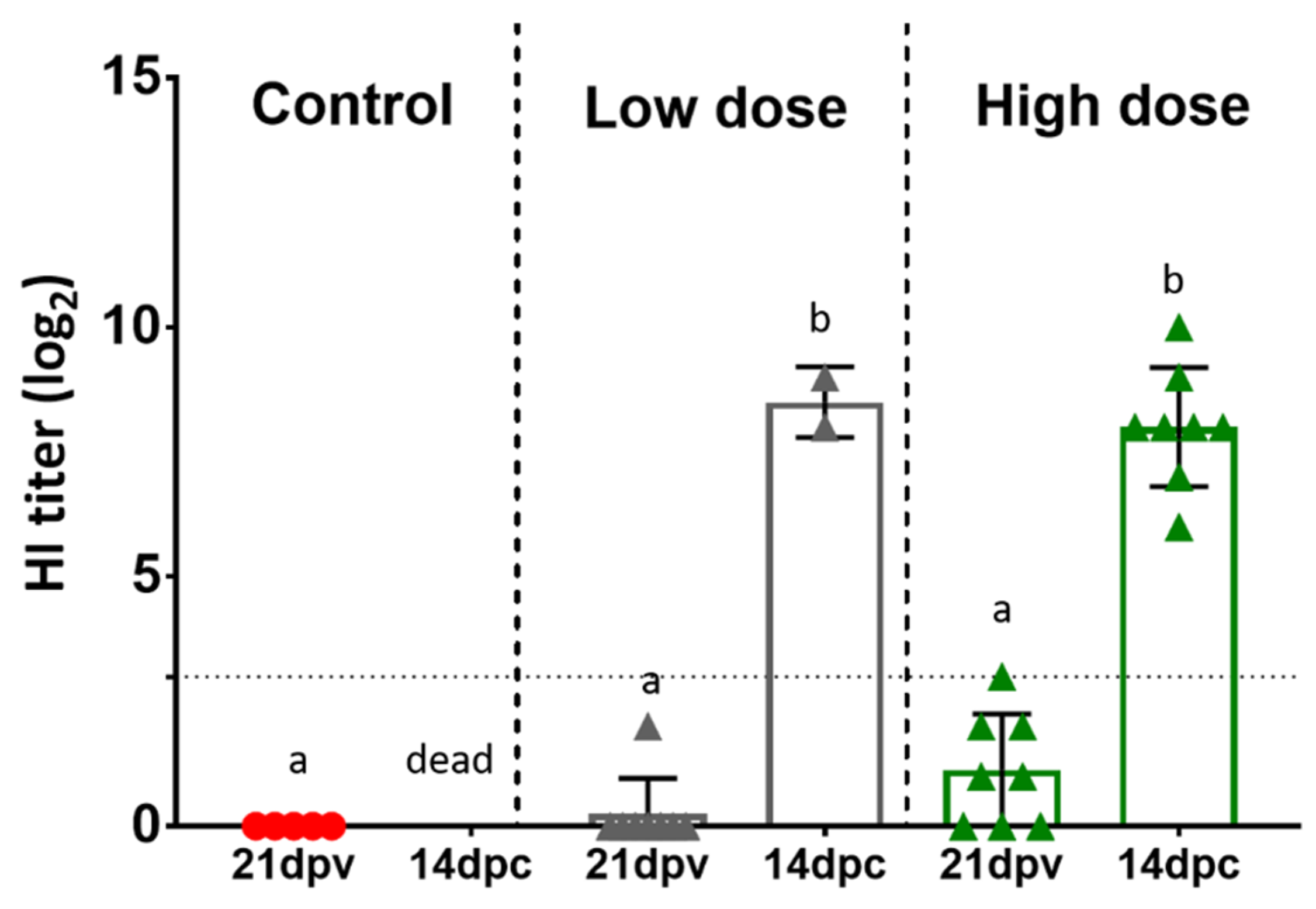
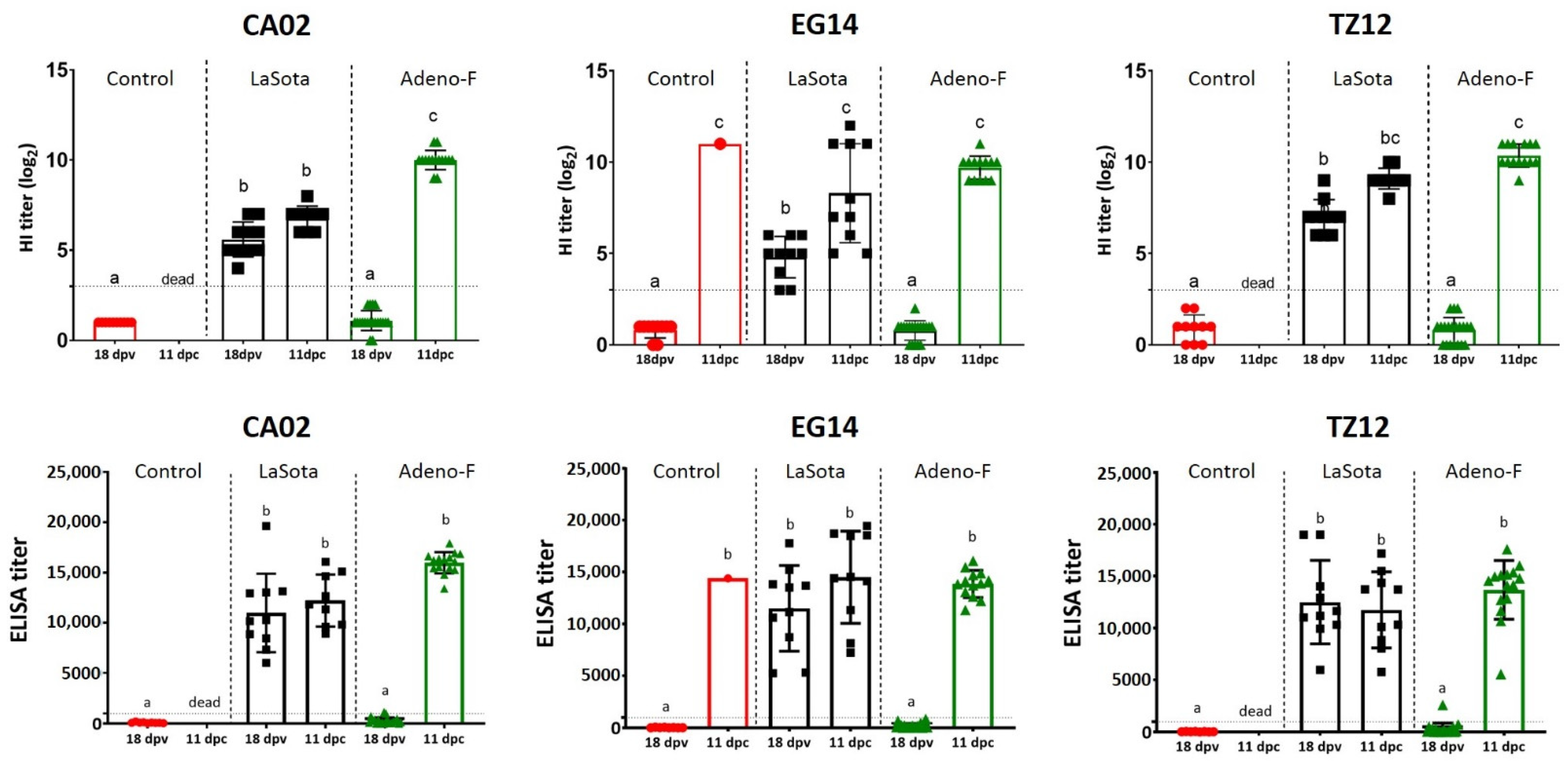
3.5. Serology
3.6. Correlation of Clinical Signs, Mortality, Virus Shedding, and Serology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, P.J.; Koch, G. Newcastle Disease. In Diseases of Poultry, 13th ed.; Swayne, D., Ed.; John Wiley & Sons, Inc.: Ames, IA, USA, 2013; pp. 89–107, 120–130. [Google Scholar]
- OIE. CHAPTER 10.9. Newcastle Disease. In Terrestrial Animal Health Code 2014, 25th ed.; OIE, Ed.; World Organisation for Animal Health: Paris, France, 2014; Volume II. [Google Scholar]
- Kuhn, J.H.; Wolf, Y.I.; Krupovic, M.; Zhang, Y.Z.; Maes, P.; Dolja, V.V.; Koonin, E.V. Classify viruses-the gain is worth the pain. Nature 2019, 566, 318–320. [Google Scholar] [CrossRef]
- Steward, M.; Vipond, I.B.; Millar, N.S.; Emmerson, P.T. RNA editing in Newcastle disease virus. J. Gen. Virol. 1993, 74, 2539–2547. [Google Scholar] [CrossRef]
- Lamb, R.A.; Parks, G. Paramyxoviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 957–995. [Google Scholar]
- Porotto, M.; Salah, Z.; DeVito, I.; Talekar, A.; Palmer, S.G.; Xu, R.; Wilson, I.A.; Moscona, A. The second receptor binding site of the globular head of the Newcastle disease virus hemagglutinin-neuraminidase activates the stalk of multiple paramyxovirus receptor binding proteins to trigger fusion. J. Virol. 2012, 86, 5730–5741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Subbiah, M.; Samuel, A.S.; Collins, P.L.; Samal, S.K. Roles of the fusion and hemagglutinin-neuraminidase proteins in replication, tropism, and pathogenicity of avian paramyxoviruses. J. Virol. 2011, 85, 8582–8596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, S.; Song, A.S.; Jardetzky, T.S.; Lamb, R.A. Fusion activation through attachment protein stalk domains indicates a conserved core mechanism of paramyxovirus entry into cells. J. Virol. 2014, 88, 3925–3941. [Google Scholar] [CrossRef] [Green Version]
- Melanson, V.R.; Iorio, R.M. Addition of N-glycans in the stalk of the Newcastle disease virus HN protein blocks its interaction with the F protein and prevents fusion. J. Virol. 2006, 80, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Panda, A.; Elankumaran, S.; Govindarajan, D.; Rockemann, D.D.; Samal, S.K. The hemagglutinin-neuraminidase protein of Newcastle disease virus determines tropism and virulence. J. Virol. 2004, 78, 4176–4184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Zhao, J.; Ren, Y.; Zhong, Q.; Zhang, G. Contribution of HN protein length diversity to Newcastle disease virus virulence, replication and biological activities. Sci. Rep. 2016, 6, 36890. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.H.; Cheng, J.L.; He, Z.R.; Ren, Y.C.; Yu, X.H.; Song, Y.; Yang, H.M.; Yang, Y.L.; Liu, T.; Zhang, G.Z. Different Origins of Newcastle Disease Virus Hemagglutinin-Neuraminidase Protein Modulate the Replication Efficiency and Pathogenicity of the Virus. Front. Microbiol. 2017, 8, 1607. [Google Scholar] [CrossRef] [Green Version]
- Dortmans, J.C.F.M.; Rottier, P.J.M.; Koch, G.; Peeters, B.P.H. The Viral Replication Complex Is Associated with the Virulence of Newcastle Disease Virus. J. Virol. 2010, 84, 10113–10120. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef]
- Arora, P.; Lakhchaura, B.D.; Garg, S.K. Evaluation of immunogenic potential of 75kDa and 56kDa proteins of newcastle disease virus (NDV). Indian J. Exp. Biol. 2010, 48, 889–895. [Google Scholar]
- Ge, J.; Liu, Y.; Jin, L.; Gao, D.; Bai, C.; Ping, W. Construction of recombinant baculovirus vaccines for Newcastle disease virus and an assessment of their immunogenicity. J. Biotechnol. 2016, 231, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Loke, C.F.; Omar, A.R.; Raha, A.R.; Yusoff, K. Improved protection from velogenic Newcastle disease virus challenge following multiple immunizations with plasmid DNA encoding for F and HN genes. Vet. Immunol. Immunopathol. 2005, 106, 259–267. [Google Scholar] [CrossRef]
- Morgan, R.W.; Gelb, J., Jr.; Schreurs, C.S.; Lutticken, D.; Rosenberger, J.K.; Sondermeijer, P.J. Protection of chickens from Newcastle and Marek’s diseases with a recombinant herpesvirus of turkeys vaccine expressing the Newcastle disease virus fusion protein. Avian Dis. 1992, 36, 858–870. [Google Scholar] [CrossRef]
- Palya, V.; Kiss, I.; Tatar-Kis, T.; Mato, T.; Felfoldi, B.; Gardin, Y. Advancement in vaccination against Newcastle disease: Recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions. Avian Dis. 2012, 56, 282–287. [Google Scholar] [CrossRef]
- Sun, H.L.; Wang, Y.F.; Tong, G.Z.; Zhang, P.J.; Miao, D.Y.; Zhi, H.D.; Wang, M.; Wang, M. Protection of chickens from Newcastle disease and infectious laryngotracheitis with a recombinant fowlpox virus co-expressing the F, HN genes of Newcastle disease virus and gB gene of infectious laryngotracheitis virus. Avian Dis. 2008, 52, 111–117. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Morgan, R.W.; Gelb, J., Jr.; Pope, C.R.; Sondermeijer, P.J. Efficacy in chickens of a herpesvirus of turkeys recombinant vaccine containing the fusion gene of Newcastle disease virus: Onset of protection and effect of maternal antibodies. Avian Dis. 1993, 37, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae: The viruses and their replication. In Fields Virology, 4th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Eds.; Lippincott Williams & Wilkins: New York, NY, USA, 2007; pp. 1450–1496. [Google Scholar]
- Miller, P.J.; Afonso, C.L.; El Attrache, J.; Dorsey, K.M.; Courtney, S.C.; Guo, Z.; Kapczynski, D.R. Effects of Newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Dev Comp. Immunol. 2013, 41, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Cornax, I.; Miller, P.J.; Afonso, C.L. Characterization of live LaSota vaccine strain-induced protection in chickens upon early challenge with a virulent Newcastle disease virus of heterologous genotype. Avian Dis. 2012, 56, 464–470. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef]
- Ferreira, H.L.; Taylor, T.L.; Dimitrov, K.M.; Sabra, M.; Afonso, C.L.; Suarez, D.L. Virulent Newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Vet. Microbiol. 2019, 235, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Ferreira, H.L.; Pantin-Jackwood, M.; Taylor, T.; Goraichuk, I.V.; Crossley, B.M.; Killian, M.L.; Bergeson, N.H.; Torchetti, M.K.; Afonso, C.L.; et al. Pathogenicity and transmission of virulent Newcastle disease virus from the 2018–2019 California outbreak and related viruses in young and adult chickens. Virology 2019, 531, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Butt, S.L.; Taylor, T.L.; Volkening, J.D.; Dimitrov, K.M.; Williams-Coplin, D.; Lahmers, K.K.; Miller, P.J.; Rana, A.M.; Suarez, D.L.; Afonso, C.L.; et al. Rapid virulence prediction and identification of Newcastle disease virus genotypes using third-generation sequencing. Virol. J. 2018, 15, 179. [Google Scholar] [CrossRef] [Green Version]
- Msoffe, P.L.M.; Chiwanga, G.H.; Cardona, C.J.; Miller, P.J.; Suarez, D.L. Isolation and Characterization of Newcastle Disease Virus from Live Bird Markets in Tanzania. Avian Dis. 2019, 63, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Senne, D. Virus propagation in embryonating eggs. In A Laboratory Manual for the Isolation, Identification and Characterization of Avian Pathogens; DofourZavala, L., Swayne, D.E., Gilsson, J.R., Pearson, J.E., Jackwood, M.W., Reed, W., Woolcock, P.R., Eds.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2008; pp. 204–208. [Google Scholar]
- Ababneh, M.; Ferreira, H.L.; Khalifeh, M.; Suarez, D.L.; Afonso, C.L. First Genome Sequence of Newcastle Disease Virus of Genotype VIIi from Jordan. Microbiol. Resour. Announc. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, K.M.; Sharma, P.; Volkening, J.D.; Goraichuk, I.V.; Wajid, A.; Rehmani, S.F.; Basharat, A.; Shittu, I.; Joannis, T.M.; Miller, P.J.; et al. A robust and cost-effective approach to sequence and analyze complete genomes of small RNA viruses. Virol. J. 2017, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Spackman, E.; Pantin-Jackwood, M.J.; Suarez, D.L. Removal of real-time reverse transcription polymerase chain reaction (RT-PCR) inhibitors associated with cloacal swab samples and tissues for improved diagnosis of Avian influenza virus by RT-PCR. J. Vet. Diagn. Invest. 2009, 21, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 8th ed.; World Organisation for Animal Health: Paris, France, 2017. [Google Scholar]
- Grimes, S.E. Newcastle disease vaccines: An overview. In A Basic Laboratory Manual for the Small-Scale Production and Testing of I-2 Newcastle Disease Vaccine; Grimes, S.E., Ed.; Food and Agriculture Organization Regional Office for Asia and the Pacific (FAO-RAP): Bangkok, Thailand, 2002. [Google Scholar]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines—A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef]
- Giambrone, J.J.; Closser, J. Effect of breeder vaccination on immunization of progeny against Newcastle disease. Avian Dis. 1990, 34, 114–119. [Google Scholar] [CrossRef]
- Westbury, H.A.; Parsons, G.; Allan, W.H. Comparison of the immunogenicity of Newcastle disease virus strains V4, Hitchner B1 and La Sota in chickens. 2. Tests in chickens with maternal antibody to the virus. Aust. Vet. J. 1984, 61, 10–13. [Google Scholar] [CrossRef]
- Martinez, J.C.S.; Chou, W.K.; Berghman, L.R.; Carey, J.B. Evaluation of the effect of live LaSota Newcastle disease virus vaccine as primary immunization on immune development in broilers. Poult. Sci. 2018, 97, 455–462. [Google Scholar] [CrossRef]
- Suarez, D.L. DIVA vaccination strategies for avian influenza virus. Avian Dis. 2012, 56, 836–844. [Google Scholar] [CrossRef]
- Boursnell, M.E.; Green, P.F.; Samson, A.C.; Campbell, J.I.; Deuter, A.; Peters, R.W.; Millar, N.S.; Emmerson, P.T.; Binns, M.M. A recombinant fowlpox virus expressing the hemagglutinin-neuraminidase gene of Newcastle disease virus (NDV) protects chickens against challenge by NDV. Virology 1990, 178, 297–300. [Google Scholar] [CrossRef]
- Bett, A.J.; Haddara, W.; Prevec, L.; Graham, F.L. An efficient and flexible system for construction of adenovirus vectors with insertions or deletions in early regions 1 and 3. Proc. Natl. Acad. Sci. USA 1994, 91, 8802–8806. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.L.; Reilley, A.M.; Goldenberg, D.; Ortiz, I.R.A.; Gallardo, R.A.; Suarez, D.L. Protection conferred by commercial NDV live attenuated and double recombinant HVT vaccines against virulent California 2018 Newcastle disease virus (NDV) in chickens. Vaccine 2020, 38, 5507–5515. [Google Scholar] [CrossRef]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector Newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.; Xu, H.; Li, J.; Hu, Z.; Hu, S.; And, X.W.; Liu, X. Effects of the HN Antigenic Difference between the Vaccine Strain and the Challenge Strain of Newcastle Disease Virus on Virus Shedding and Transmission. Viruses 2017, 9, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahar, E.; Haddas, R.; Goldenberg, D.; Lublin, A.; Bloch, I.; Bachner Hinenzon, N.; Pitcovski, J. Newcastle disease virus: Is an updated attenuated vaccine needed? Avian Pathol. 2018, 47, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Maraqa, A. Protective immunity against Newcastle disease: The role of cell-mediated immunity. Avian Dis. 2000, 44, 145–154. [Google Scholar] [CrossRef]
- Firouzamandi, M.; Moeini, H.; Hosseini, D.; Bejo, M.H.; Omar, A.R.; Mehrbod, P.; Ideris, A. Improved immunogenicity of Newcastle disease virus inactivated vaccine following DNA vaccination using Newcastle disease virus hemagglutinin-neuraminidase and fusion protein genes. J. Vet. Sci. 2016, 17, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.L.; Rauw, F.; Pirlot, J.F.; Reynard, F.; van den Berg, T.; Bublot, M.; Lambrecht, B. Comparison of single 1-day-old chick vaccination using a Newcastle disease virus vector with a prime/boost vaccination scheme against a highly pathogenic avian influenza H5N1 challenge. Avian Pathol. 2014, 43, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Li, Q.; Ertl, H.C.; Wilson, J.M. Cellular and humoral immune responses to viral antigens create barriers to lung-directed gene therapy with recombinant adenoviruses. J. Virol. 1995, 69, 2004–2015. [Google Scholar] [CrossRef] [Green Version]
- Kafri, T.; Morgan, D.; Krahl, T.; Sarvetnick, N.; Sherman, L.; Verma, I. Cellular immune response to adenoviral vector infected cells does not require de novo viral gene expression: Implications for gene therapy. Proc. Natl. Acad. Sci. USA 1998, 95, 11377–11382. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Soloff, A.C.; Lu, X.; Montecalvo, A.; Nguyen, D.C.; Matsuoka, Y.; Robbins, P.D.; Swayne, D.E.; Donis, R.O.; Katz, J.M.; et al. Protection of mice and poultry from lethal H5N1 avian influenza virus through adenovirus-based immunization. J. Virol. 2006, 80, 1959–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 18 dpv | 11 dpc | |||
|---|---|---|---|---|
| Challenge Virus | Bird Tag | Titer (1/100) | Titer (1/500) | Titer (1/500) |
| CA02 | 30 | 1283 | 104 | 16,480 |
| 31 | 779 | 0 | 15,545 | |
| 32 | 147 | 0 | 16,932 | |
| 33 | 3451 | 1128 | 15,453 | |
| 34 | 1155 | 197 | NS | |
| 35 | 809 | 394 | 16,274 | |
| 36 | 1291 | 389 | 16,214 | |
| 37 | 1674 | 482 | NT | |
| 38 | 2006 | 126 | 16,622 | |
| 39 | 2442 | 1008 | 16,377 | |
| 40 | 418 | 263 | NS | |
| 41 | 478 | 42 | 15,436 | |
| 42 | 711 | 201 | NS | |
| 43 | 3842 | 729 | 15,268 | |
| 44 | 1200 | 214 | 16,856 | |
| 45 | 1246 | 274 | 16,089 | |
| 46 | 613 | 0 | NS | |
| 47 | 297 | 0 | 14,779 | |
| 48 | 917 | 44 | 13,397 | |
| 49 | 290 | 69 | 17,900 | |
| Mean | 1252 | 283 | 15,974.8 | |
| EG14 | 70 | 147 | 11 | 15,464 |
| 71 | 0 | 27 | NS | |
| 72 | 1125 | 0 | 13,903 | |
| 73 | 1396 | 125 | 13,827 | |
| 74 | 1373 | 687 | 14,110 | |
| 75 | 1109 | 21 | 13,713 | |
| 76 | 79 | 60 | NS | |
| 77 | 3195 | 860 | 14,716 | |
| 78 | 726 | 203 | NS | |
| 79 | 470 | 186 | 12,631 | |
| 80 | 282 | 219 | NS | |
| 81 | 34 | 93 | NS | |
| 82 | 102 | 246 | NS | |
| 83 | 681 | 76 | NS | |
| 84 | 1735 | 264 | 12,220 | |
| 85 | 147 | 7 | 16,078 | |
| 86 | 1321 | 49 | 14,879 | |
| 87 | 297 | 104 | 11,329 | |
| 88 | 1629 | 186 | 14,191 | |
| 89 | 2713 | 482 | 13,074 | |
| Mean | 928 | 258 | 13,857 | |
| TZ12 | 110 | 282 | 509 | 10,668 |
| 111 | 237 | 60 | NS | |
| 112 | 64 | 88 | 11,652 | |
| 113 | 0 | 181 | NS | |
| 114 | 214 | 4 | 12,837 | |
| 115 | 2134 | 111 | 15,360 | |
| 116 | 478 | 31 | 14,132 | |
| 117 | 3533 | 729 | 15,154 | |
| 118 | 0 | 0 | 5549 | |
| 119 | 1795 | 236 | 14,955 | |
| 120 | 0 | 44 | NS | |
| 121 | 290 | 0 | 14,550 | |
| 122 | 8390 | 2613 | 14,096 | |
| 123 | 689 | 471 | 16,013 | |
| 124 | 0 | 307 | NS | |
| 125 | 0 | 0 | NS | |
| 126 | 493 | 0 | 12,704 | |
| 127 | 2585 | 323 | 15,083 | |
| 128 | 1915 | 159 | 14,787 | |
| 129 | 403 | 142 | 17,590 | |
| Mean | 1567 | 327 | 13,675 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, H.L.; Miller, P.J.; Suarez, D.L. Protection against Different Genotypes of Newcastle Disease Viruses (NDV) Afforded by an Adenovirus-Vectored Fusion Protein and Live NDV Vaccines in Chickens. Vaccines 2021, 9, 182. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020182
Ferreira HL, Miller PJ, Suarez DL. Protection against Different Genotypes of Newcastle Disease Viruses (NDV) Afforded by an Adenovirus-Vectored Fusion Protein and Live NDV Vaccines in Chickens. Vaccines. 2021; 9(2):182. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020182
Chicago/Turabian StyleFerreira, Helena L., Patti J. Miller, and David L. Suarez. 2021. "Protection against Different Genotypes of Newcastle Disease Viruses (NDV) Afforded by an Adenovirus-Vectored Fusion Protein and Live NDV Vaccines in Chickens" Vaccines 9, no. 2: 182. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020182