
Development and Validation of Enzyme-Linked Immunosorbent Assay for Group B Streptococcal Polysaccharide Vaccine

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Reagents
2.3. Bacteria Strains
2.4. Purification of GBS PS
2.5. PS Immobilization on 96-Well Plate
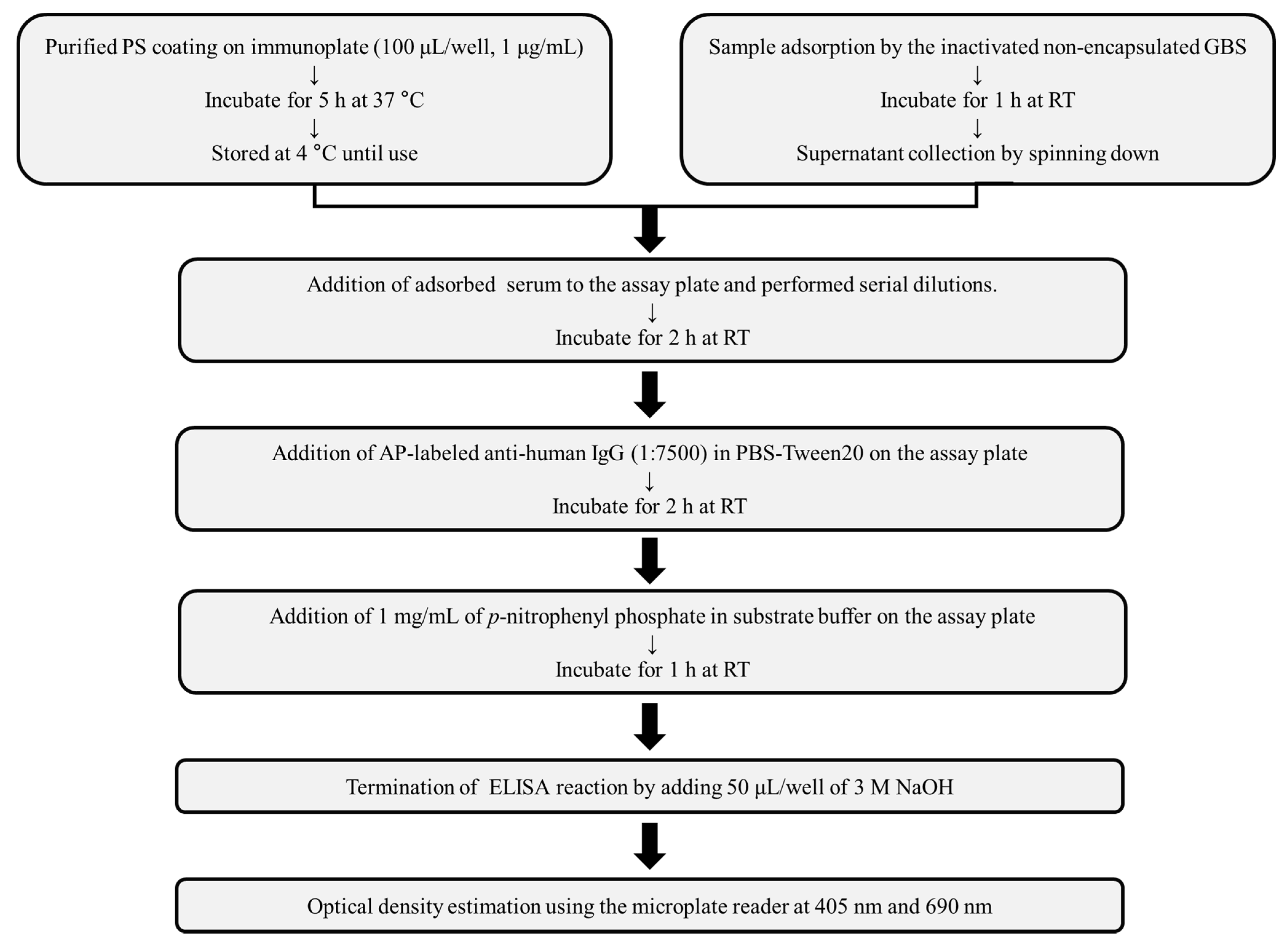
2.6. GBS-ELISA
2.7. Validation of GBS-ELISA
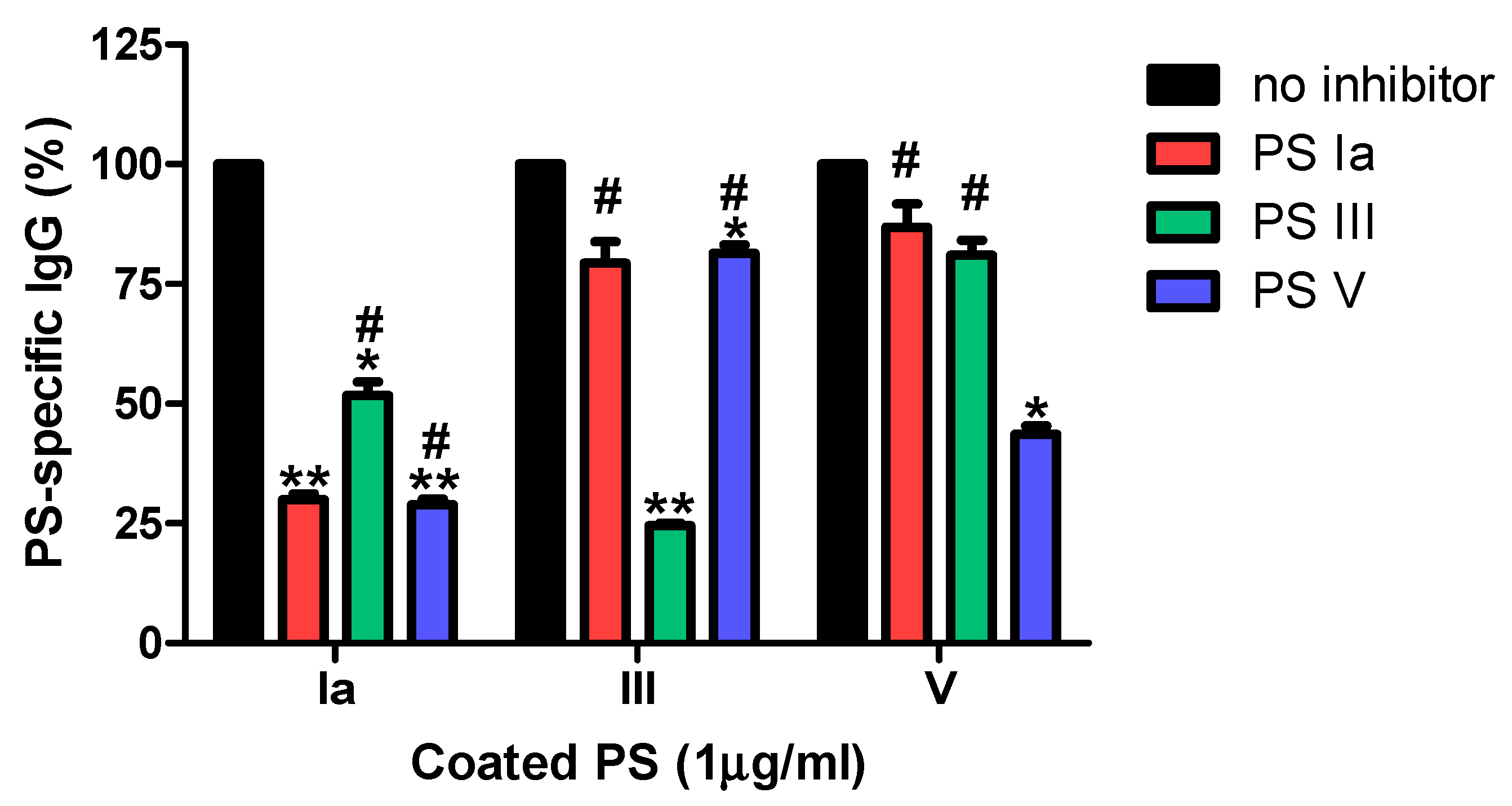
2.8. Inhibition ELISA
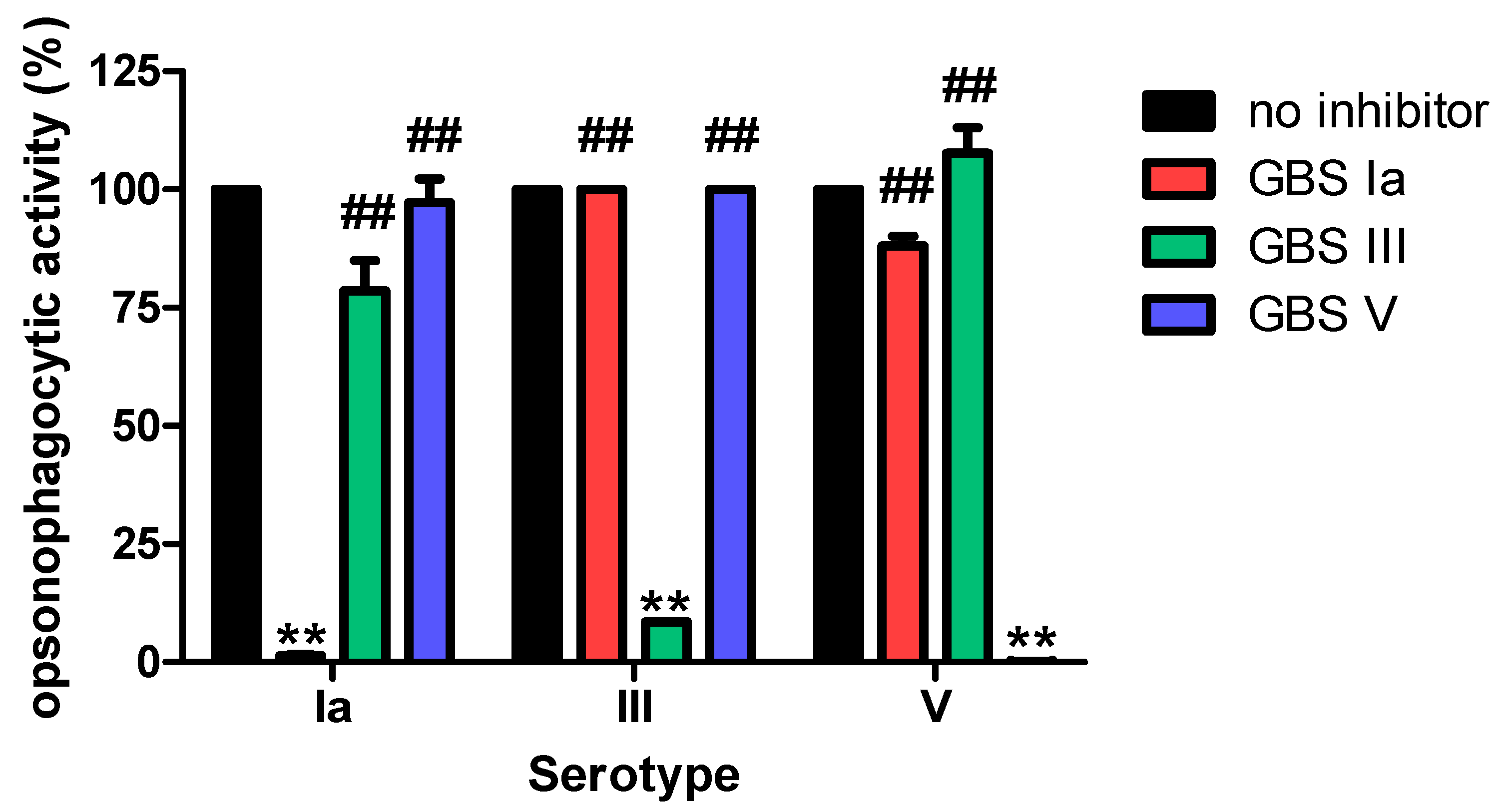
2.9. Opsonophagocytic Killing Assay
2.10. Statistical Analysis
3. Results
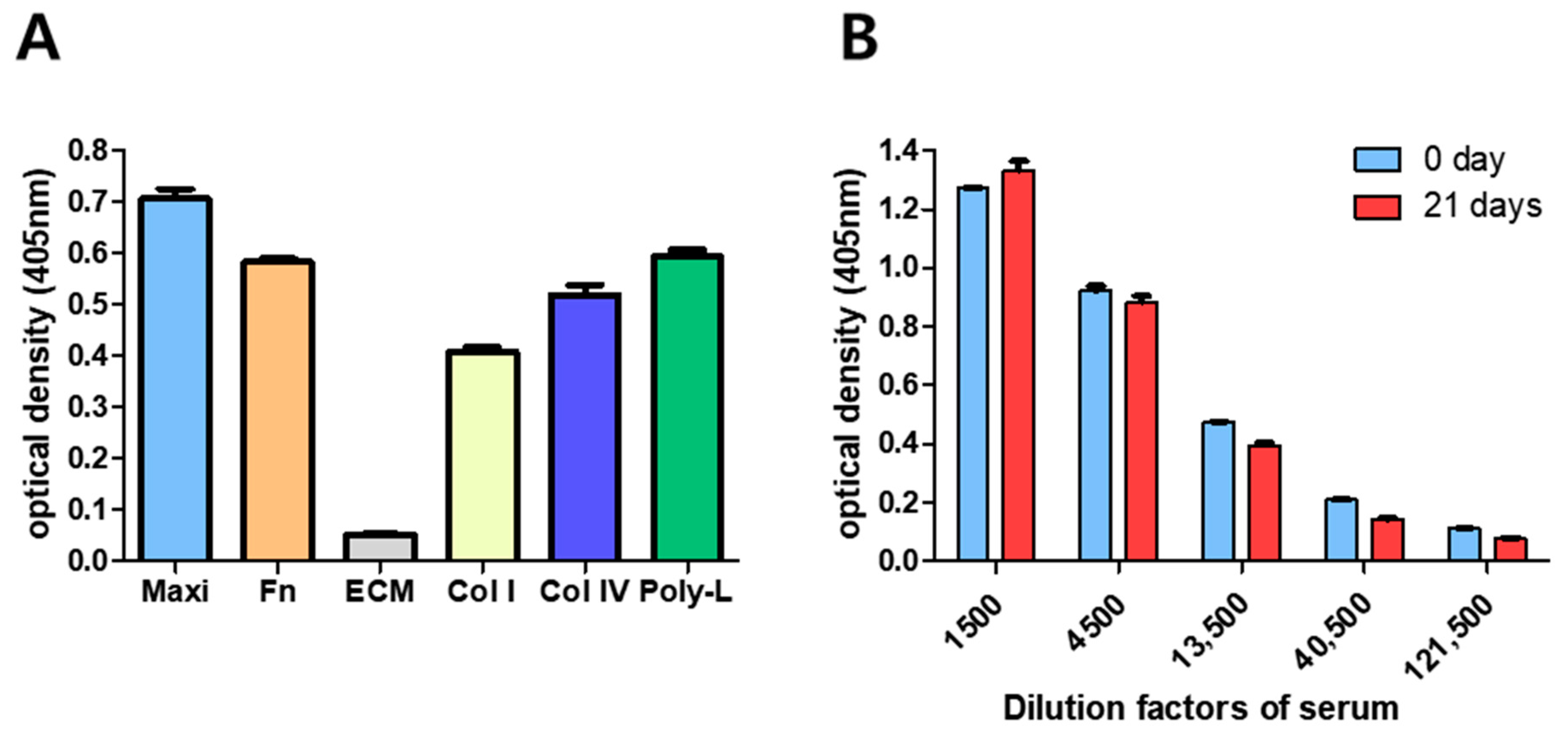
3.1. Optimization of GBS-ELISA
3.2. GBS-ELISA Validation
3.2.1. Specificity
3.2.2. Precision (Reproducibility)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berner, R. Group B Streptococcus Vaccines: One Step Further. Lancet Infect. Dis. 2021, 21, 158–160. [Google Scholar] [CrossRef]
- Song, J.Y.; Lim, J.H.; Lim, S.; Yong, Z.; Seo, H.S. Progress toward a group B streptococcal vaccine. Hum. Vaccin. Immunother. 2018, 14, 2669–2681. [Google Scholar] [CrossRef]
- Schrag, S.; Gorwitz, R.; Fultz-Butts, K.; Schuchat, A. Prevention of perinatal group B streptococcal disease. Revised guidelines from CDC. M.M.W.R. Recomm. Rep. 2002, 51, 1–22. [Google Scholar]
- Russell, N.J.; Seale, A.C.; O’Sullivan, C.; Le Doare, K.; Heath, P.T.; Lawn, J.E.; Bartlett, L.; Cutland, C.; Gravett, M.; Ip, M.; et al. Risk of early-onset neonatal group B streptococcal disease with maternal colonization worldwide: Systematic review and meta-analyses. Clin. Infect. Dis. 2017, 65, S152–S159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwatra, G.; Adrian, P.V.; Shiri, T.; Buchmann, E.J.; Cutland, C.L.; Madhi, S.A. Natural acquired humoral immunity against serotype-specific group B Streptococcus rectovaginal colonization acquisition in pregnant women. Clin. Microbiol. Infect. 2015, 21, 568.e13–568.e21. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Schrag, S.J.; Alderson, M.R.; Madhi, S.A.; Baker, C.J.; Sobanjo-Ter Meulen, A.; Kaslow, D.C.; Smith, P.G.; Moorthy, V.S.; Vekemans, J. WHO consultation on group B Streptococcus vaccine development: Report from a meeting held on 27–28 April 2016. Vaccine 2016, 37, 7307–7314. [Google Scholar] [CrossRef]
- Absalon, J.; Segall, N.; Block, S.L.; Center, K.J.; Scully, I.L.; Giardina, P.C.; Peterson, J.; Watson, W.J.; Gruber, W.C.; Jansen, K.U.; et al. Safety and immunogenicity of a novel hexavalent group B Streptococcus conjugate vaccine in healthy, non-pregnant adults: A phase 1/2, randomised, placebo-controlled, observer-blinded, dose-escalation trial. Lancet Infect. Dis. 2021, 21, 263–274. [Google Scholar] [CrossRef]
- Swamy, G.K.; Metz, T.D.; Edwards, K.M.; Soper, D.E.; Beigi, R.H.; Campbell, J.D.; Grassano, L.; Buffi, G.; Dreisbach, A.; Margarit, I.; et al. Safety and immunogenicity of an investigational maternal trivalent group B Streptococcus vaccine in pregnant women and their infants: Results from a randomized placebo-controlled phase II trial. Vaccine 2020, 38, 6930–6940. [Google Scholar] [CrossRef]
- Madhi, S.A.; Cutland, C.L.; Jose, L.; Koen, A.; Govender, N.; Wittke, F.; Olugbosi, M.; Meulen, A.S.; Baker, S.; Dull, P.M.; et al. Safety and immunogenicity of an investigational maternal trivalent group B Streptococcus vaccine in healthy women and their infants: A randomised phase 1b/2 trial. Lancet Infect. Dis. 2016, 16, 923–934. [Google Scholar] [CrossRef]
- Vekemans, J.; Crofts, J.; Baker, C.J.; Goldblatt, D.; Heath, P.T.; Madhi, S.A.; Le Doare, K.; Andrews, N.; Pollard, A.J.; Saha, S.K.; et al. The role of immune correlates of protection on the pathway to licensure, policy decision and use of group B Streptococcus vaccines for maternal immunization: Considerations from world health organization consultations. Vaccine 2019, 37, 3190–3198. [Google Scholar] [CrossRef]
- Le Doare, K.; Kampmann, B.; Vekemans, J.; Heath, P.T.; Goldblatt, D.; Nahm, M.H.; Baker, C.; Edwards, M.S.; Kwatra, G.; Andrews, N.; et al. Serocorrelates of protection against infant group B Streptococcus disease. Lancet Infect. Dis. 2019, 19, e162–e171. [Google Scholar] [CrossRef]
- Dangor, Z.; Kwatra, G.; Izu, A.; Adrian, P.; Cutland, C.L.; Velaphi, S.; Ballot, D.; Reubenson, G.; Zell, E.R.; Lala, S.G.; et al. Correlates of protection of serotype-specific capsular antibody and invasive group B Streptococcus disease in South African infants. Vaccine 2015, 33, 6793–6799. [Google Scholar] [CrossRef] [PubMed]
- Balmer, P.; Borrow, R.; Miller, E. Impact of meningococcal C conjugate vaccine in the UK. J. Med. Microbiol. 2002, 51, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.M.; Baek, C.Y.; Jung, J.H.; Kim, W.S.; Song, H.Y.; Lee, J.H.; Ji, H.J.; Zhi, Y.; Kang, B.S.; Bahn, Y.S.; et al. Antioxidant activities of an exopolysaccharide (DeinoPol) produced by the extreme radiation-resistant bacterium Deinococcus Radiodurans. Sci. Rep. 2020, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jourdian, G.W.; Dean, L.; Roseman, S. The Sialic Acids. XI. A periodate-resorcinol method for the quantitative estimation of free sialic acids and their glycosides. J. Biol. Chem. 1971, 246, 430–435. [Google Scholar] [CrossRef]
- Svennerholm, L. Quantitative estimation of sialic acids: II. A colorimetric resorcinol-hydrochloric acid method. Biochim. Biophys. Acta 1957, 24, 604–611. [Google Scholar] [CrossRef]
- World Health Organization. Training Manual for Enzyme Linked Immunosorbent Assay for the Quantitation of Streptococcus Pneumoniae Serotype Specific IgG (Pn PS ELISA). 2002. Bacterial Respiratory Pathogen Reference Laboratory Website. Available online: www.vaccine.uab.edu (accessed on 28 February 2021).
- Choi, M.J.; Noh, J.Y.; Cheong, H.J.; Kim, W.J.; Lin, S.M.; Zhi, Y.; Lim, J.H.; Lim, S.; Seo, H.S.; Song, J.Y. Development of a multiplexed opsonophagocytic killing assay (MOPA) for group B Streptococcus. Hum. Vaccin. Immunother. 2018, 14, 67–73. [Google Scholar] [CrossRef]
- Buffi, G.; Galletti, B.; Stella, M.; Proietti, D.; Balducci, E.; Romano, M.R.; Mori, E.; Fabbrini, M.; Giuliani, M.M.; Berti, F.; et al. Novel multiplex immunoassays for quantification of IgG against group B Streptococcus capsular polysaccharides in human sera. mSphere 2019, 4, e00273. [Google Scholar] [CrossRef] [Green Version]
- Edmond, K.M.; Kortsalioudaki, C.; Scott, S.; Schrag, S.J.; Zaidi, A.K.; Cousens, S.; Heath, P.T. Group B streptococcal disease in infants aged younger than 3 months: Systematic review and meta-analysis. Lancet 2012, 379, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.L.; Antonello, J.; Cooper, D.; Goldblatt, D.; Kim, K.H.; Plikaytis, B.D.; Roalfe, L.; Wauters, D.; Williams, F.; Xie, G.L.; et al. Assignment of opsonic values to pneumococcal reference serum 007sp for use in opsonophagocytic assays for 13 serotypes. Clin. Vaccin. Immunol. 2017, 24, e00457-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.J.; Noh, J.Y.; Jang, A.Y.; Cheong, H.J.; Kim, W.J.; Song, D.J.; Cho, G.J.; Oh, M.J.; Zhi, Y.; Seo, H.S.; et al. Age-stratified analysis of serotype-specific baseline immunity against group B Streptococcus. Hum. Vaccin. Immunother. 2020, 16, 1338–1344. [Google Scholar] [CrossRef]
- Song, J.Y.; Moseley, M.A.; Burton, R.L.; Nahm, M.H. Pneumococcal vaccine and opsonic pneumococcal antibody. J. Infect. Chemother. 2013, 19, 412–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbrini, M.; Rigat, F.; Rinaudo, C.D.; Passalaqua, I.; Khacheh, S.; Creti, R.; Baldassarri, L.; Carboni, F.; Anderloni, G.; Rosini, R.; et al. The protective value of maternal group B Streptococcus antibodies: Quantitative and functional analysis of naturally acquired responses to capsular polysaccharides and pilus proteins in european maternal sera. Clin. Infect. Dis. 2016, 63, 746–753. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Serotypes | Characteristics | Origin |
|---|---|---|---|
| S10118 | Ia | None | Blood |
| NSP14-212 | III | Levofloxacin-resistant | CSF |
| NSP14-168 | V | Erythromycin-resistant | Blood |
| NSP14-358 | Non-typeable | Levofloxacin -resistant | Conjunctiva |
| Serotypes | Dilution Fold | PS Concentration (μg/mL) | |||||
|---|---|---|---|---|---|---|---|
| 10 | 5 | 2.5 | 1.25 | 0.625 | 0 | ||
| Serotype Ia absorbance (405 nm) | 40 | 3.331 | 3.240 | 3.263 | 3.303 | 3.316 | 0.237 |
| 80 | 3.157 | 3.026 | 3.081 | 3.142 | 3.179 | 0.182 | |
| 160 | 2.749 | 2.550 | 2.570 | 2.764 | 2.821 | 0.126 | |
| 320 | 2.132 | 1.934 | 1.990 | 2.173 | 2.274 | 0.099 | |
| 640 | 1.399 | 1.222 | 1.325 | 1.465 | 1.436 | 0.076 | |
| r2 | 0.95 | 0.97 | 0.97 | 0.95 | 0.92 | 0.96 | |
| Serotype III absorbance (405 nm) | 40 | 2.094 | 1.933 | 1.707 | 1.393 | 0.987 | 0.194 |
| 80 | 1.564 | 1.432 | 1.249 | 1.001 | 0.696 | 0.140 | |
| 160 | 1.089 | 0.970 | 0.845 | 0.684 | 0.474 | 0.107 | |
| 320 | 0.698 | 0.635 | 0.555 | 0.457 | 0.309 | 0.088 | |
| 640 | 0.424 | 0.396 | 0.339 | 0.278 | 0.201 | 0.044 | |
| r2 | 0.99 | 0.98 | 0.98 | 0.98 | 0.97 | 0.97 | |
| Serotype V absorbance (405 nm) | 40 | 2.448 | 2.319 | 2.224 | 2.087 | 1.844 | 0.209 |
| 80 | 1.887 | 1.751 | 1.651 | 1.482 | 1.256 | 0.161 | |
| 160 | 1.295 | 1.189 | 1.104 | 0.947 | 0.782 | 0.120 | |
| 320 | 0.799 | 0.727 | 0.674 | 0.569 | 0.451 | 0.090 | |
| 640 | 0.455 | 0.409 | 0.381 | 0.312 | 0.248 | 0.073 | |
| r2 | 0.99 | 0.99 | 0.98 | 0.97 | 0.96 | 0.97 | |
| Sample | Serotype Ia | Serotype III | Serotype V | |||
|---|---|---|---|---|---|---|
| Mean ± SD | CV (%) | Mean ± SD | CV (%) | Mean ± SD | CV (%) | |
| Sample 1 | 1.792 ± 0.04 | 2.3 | 1.783 ± 0.04 | 2.0 | 1.913 ± 0.03 | 1.7 |
| Sample 2 | 1.524 ± 0.04 | 2.5 | 1.703 ± 0.03 | 1.5 | 1.903 ± 0.04 | 2.2 |
| Sample 3 | 1.614 ± 0.09 | 5.3 | 1.747 ± 0.02 | 0.9 | 1.910 ± 0.03 | 1.5 |
| Sample 4 | 1.699 ± 0.05 | 3.2 | 1.663 ± 0.05 | 3.2 | 1.905 ± 0.02 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, A.-Y.; Choi, M.-J.; Zhi, Y.; Ji, H.-J.; Noh, J.-Y.; Yoon, J.-G.; Cheong, H.-J.; Kim, W.-J.; Seo, H.-S.; Song, J.-Y. Development and Validation of Enzyme-Linked Immunosorbent Assay for Group B Streptococcal Polysaccharide Vaccine. Vaccines 2021, 9, 545. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060545
Jang A-Y, Choi M-J, Zhi Y, Ji H-J, Noh J-Y, Yoon J-G, Cheong H-J, Kim W-J, Seo H-S, Song J-Y. Development and Validation of Enzyme-Linked Immunosorbent Assay for Group B Streptococcal Polysaccharide Vaccine. Vaccines. 2021; 9(6):545. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060545
Chicago/Turabian StyleJang, A-Yeung, Min-Joo Choi, Yong Zhi, Hyun-Jung Ji, Ji-Yun Noh, Jin-Gu Yoon, Hee-Jin Cheong, Woo-Joo Kim, Ho-Seong Seo, and Joon-Young Song. 2021. "Development and Validation of Enzyme-Linked Immunosorbent Assay for Group B Streptococcal Polysaccharide Vaccine" Vaccines 9, no. 6: 545. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060545