
Evaluating Functional Immunity Following Encapsulated Bacterial Infection and Vaccination

 ,
, 
Abstract
:1. Introduction
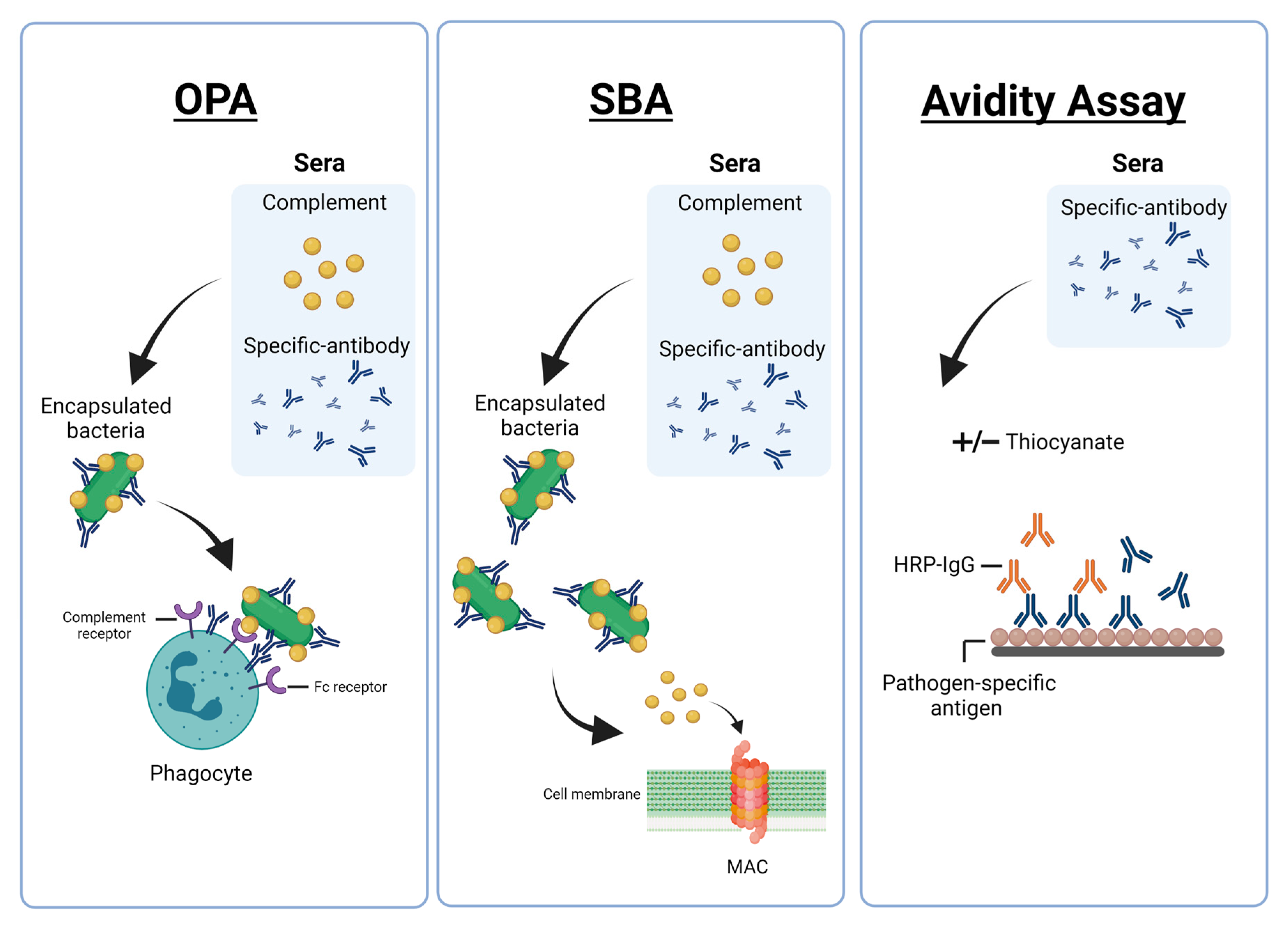
2. Functional Antibody Assays against Encapsulated Bacteria
3. Importance of Measuring Functional Antibodies Following Vaccination
4. Standardisation of Functional Antibody Assays against Encapsulated Bacteria
5. OPA for Evaluating (New) Bacterial Vaccines
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Global Burden of Disease Collaborators. Global, regional, and national burden of meningitis, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 1061–1082. [Google Scholar] [CrossRef] [Green Version]
- Wahl, B.; O’Brien, K.L.; Greenbaum, A.; Majumder, A.; Liu, L.; Chu, Y.; Luksic, I.; Nair, H.; McAllister, D.A.; Campbell, H.; et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vac-cines: Global, regional, and national estimates for 2000–2015. Lancet Glob. Health 2018, 6, e744–e757. [Google Scholar] [CrossRef] [Green Version]
- Siegel, S.J.; Weiser, J.N. Mechanisms of Bacterial Colonization of the Respiratory Tract. Annu. Rev. Microbiol. 2015, 69, 425–444. [Google Scholar] [CrossRef] [Green Version]
- Chiappini, E.; Inturrisi, F.; Orlandini, E.; De Martino, M.; De Waure, C. Hospitalization rates and outcome of invasive bacterial vaccine-preventable diseases in Tuscany: A historical cohort study of the 2000–2016 period. BMC Infect. Dis. 2018, 18, 396. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoe, E.; Anderson, J.; Nathanielsz, J.; Toh, Z.Q.; Marimla, R.; Balloch, A.; Licciardi, P.V. The contrasting roles of Th17 immunity in human health and disease. Microbiol. Immunol. 2017, 61, 49–56. [Google Scholar] [CrossRef]
- Mubarak, A.; Ahmed, M.; Upile, N.; Vaughan, C.; Xie, C.; Sharma, R.; Acar, P.; McCormick, M.; Paton, J.; Mitchell, T.; et al. A dynamic relationship between mucosal T helper type 17 and regulatory T-cell populations in nasopharynx evolves with age and associates with the clearance of pneumococcal carriage in humans. Clin. Microbiol. Infect. 2016, 22, 736.e1–736.e7. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.L.; Suscovich, T.J.; Fortune, S.M.; Alter, G. Beyond binding: Antibody effector functions in infectious diseases. Nat. Rev. Immunol. 2018, 18, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Jódar, L.; Butler, J.; Carlone, G.; Dagan, R.; Goldblatt, D.; Käyhty, H.; Klugman, K.; Plikaytis, B.; Siber, G.; Kohberger, R.; et al. Serological criteria for evaluation and licensure of new pneumococcal conjugate vaccine formulations for use in infants. Vaccine 2003, 21, 3265–3272. [Google Scholar] [CrossRef]
- Oliveira, J.B.; Fleisher, T.A. Laboratory evaluation of primary immunodeficiencies. J. Allergy Clin. Immunol. 2010, 125, S297–S305. [Google Scholar] [CrossRef] [Green Version]
- The World Health Organization. WHO Workshop on Standardization of Pneumococcal Opsonophagocytic Assay; World Health Organization: Geneva, Switzerland, 2007; Available online: https://www.who.int/publications/m/item/who-workshop-on-standardization-of-pneumococcal-opsonophagocytic-assay-geneva-switzerland-25-26-january-2007 (accessed on 22 April 2021).
- Arnold, K.B.; Chung, A.W. Prospects from systems serology research. Immunology 2018, 153, 279–289. [Google Scholar] [CrossRef]
- The World Health Organization. Guidelines on clinical evaluation of vaccines: Regulatory expectations. In WHO Technical Report Series 1004, Annex 9. 2017. Available online: https://www.who.int/publications/m/item/WHO-TRS-1004-web-annex-9 (accessed on 7 April 2021).
- Feyssaguet, M.; Bellanger, A.; Nozay, F.; Friel, D.; Merck, E.; Verlant, V.; Malevé, M.; Lallemand, S.; El Moussaoui, A.; D’Argoeuves, P.D.G.; et al. Comparison between a new multiplex electrochemiluminescence assay and the WHO reference enzyme-linked immunosorbent assay to measure serum antibodies against pneumococcal serotype-specific polysaccharides. Vaccine 2019, 37, 2208–2215. [Google Scholar] [CrossRef] [PubMed]
- Romero-Steiner, S.; Musher, D.M.; Cetron, M.S.; Pais, L.B.; Groover, J.E.; Carlone, G.M. Reduction in Functional Antibody Activity Against Streptococcus pneumoniae in Vaccinated Elderly Individuals Highly Correlates with Decreased IgG Antibody Avidity. Clin. Infect. Dis. 1999, 29, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Licciardi, P.V.; Balloch, A.; Russell, F.M.; Burton, R.L.; Lin, J.; Nahm, M.; Mulholland, E.K.; Tang, M.L.; Licciardi, P.V.; Balloch, A.; et al. Pneumococcal polysaccharide vaccine at 12 months of age produces functional immune responses. J. Allergy Clin. Immunol. 2012, 129, 794–800.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voysey, M.; Fanshawe, T.R.; Kelly, D.F.; O’Brien, K.L.; Kandasamy, R.; Shrestha, S.; Thorson, S.; Hinds, J.; Pollard, A.J. Serotype-Specific Correlates of Protection for Pneumococcal Carriage: An Analysis of Immunity in 19 Countries. Clin. Infect. Dis. 2017, 66, 913–920. [Google Scholar] [CrossRef]
- Lottenbach, K.R.; Granoff, D.M.; Barenkamp, S.J.; Powers, D.C.; Kennedy, D.; Irby-Moore, S.; Homan, S.M.; Mink, C.M. Safety and Immunogenicity of Haemophilus Influenzae Type B Polysaccharide or Conjugate Vaccines in an Elderly Adult Population. J. Am. Geriatr. Soc. 2004, 52, 1883–1887. [Google Scholar] [CrossRef] [PubMed]
- Dentinger, C.M.; Hennessy, T.W.; Bulkow, L.R.; Reasonover, A.L.; Romero-Steiner, S.; Holder, P.F.; De Leon, P.G.; Carlone, G.M.; Parks, D.J.; Parkinson, A.J.; et al. Immunogenicity and reactogenicity to Haemophilus influenzae type B (Hib) conjugate vaccine among rural Alaska adults. Hum. Vaccines 2006, 2, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Nix, E.B.; Hawdon, N.; Gravelle, S.; Biman, B.; Brigden, M.; Malik, S.; McCready, W.; Ferroni, G.; Ulanova, M. Risk of Invasive Haemophilus influenzae Type b (Hib) Disease in Adults with Secondary Immunodeficiency in the Post-Hib Vaccine Era. Clin. Vaccine Immunol. 2012, 19, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Boslego, J. Efficacy, safety, and immunogenicity of a meningococcal group B (15:P1.3) outer membrane protein vaccine in Iquique, Chile. Vaccine 1995, 13, 821–829. [Google Scholar] [CrossRef]
- Perkins, B.A.; Jonsdottir, K.; Briem, H.; Griffiths, E.; Plikaytis, B.D.; Hoiby, E.A.; Rosenqvist, E.; Holst, J.; Nokleby, H.; Sotolongo, F.; et al. Immunogenicity of two efficacious outer membrane protein-based serogroup B meningococcal vaccines among young adults in Iceland. J. Infect. Dis. 1998, 177, 683–691. [Google Scholar] [CrossRef] [Green Version]
- Vermont, C.; van den Dobbelsteen, G. Neisseria meningitidis serogroup B: Laboratory correlates of protection. FEMS Immunol. Med. Microbiol. 2002, 34, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Nahm, M.H. Older Adults Have a Low Capacity to Opsonize Pneumococci Due to Low IgM Antibody Response to Pneumococcal Vaccinations. Infect. Immun. 2011, 79, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Ulanova, M.; Hahn-Zoric, M.; Lau, Y.L.; Lucas, A.; Hanson, L.A. Expression of Haemophilus influenzae type b idiotype 1 on naturally acquired antibodies. Clin. Exp. Immunol. 1996, 105, 422–428. [Google Scholar] [CrossRef]
- Temple, B.; Toan, N.T.; Dai, V.T.T.; Bright, K.; Licciardi, P.V.; Marimla, R.A.; Nguyen, C.D.; Uyen, D.Y.; Balloch, A.; Huu, T.N.; et al. Immunogenicity and reactogenicity of ten-valent versus 13-valent pneumococcal conjugate vaccines among infants in Ho Chi Minh City, Vietnam: A randomised controlled trial. Lancet Infect. Dis. 2019, 19, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Andrews, N.J.; Waight, P.A.; Burbidge, P.; Pearce, E.; Roalfe, L.; Zancolli, M.; Slack, M.; Ladhani, S.N.; Miller, E.; Goldblatt, D. Serotype-specific effectiveness and correlates of protection for the 13-valent pneumococcal conjugate vaccine: A postlicensure indirect cohort study. Lancet Infect. Dis. 2014, 14, 839–846. [Google Scholar] [CrossRef]
- Correa, V.A.; Rodrigues, T.S.; Portilho, A.I.; De Lima, G.T.; De Gaspari, E. Modified ELISA for Antibody Avidity Evaluation: The need for standardization. Biomed. J. 2020. [Google Scholar] [CrossRef]
- Maslanka, S.E.; Gheesling, L.L.; Libutti, D.E.; Donaldson, K.B.; Harakeh, H.S.; Dykes, J.K.; Arhin, F.F.; Devi, S.J.; Frasch, C.E.; Huang, J.C.; et al. Standardization and a multilaboratory comparison of Neisseria meningitidis serogroup A and C serum bactericidal assays. The Multilaboratory Study Group. Clin. Diagn. Lab. Immunol. 1997, 4, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Borrow, R.; Aaberge, I.S.; Santos, G.F.; Eudey, T.L.; Oster, P.; Glennie, A.; Findlow, J.; Høiby, E.A.; Rosenqvist, E.; Balmer, P.; et al. Interlaboratory Standardization of the Measurement of Serum Bactericidal Activity by Using Human Complement against Meningococcal Serogroup B, Strain 44/76-SL, before and after Vaccination with the Norwegian MenBvac Outer Membrane Vesicle Vaccine. Clin. Vaccine Immunol. 2005, 12, 970–976. [Google Scholar] [CrossRef] [Green Version]
- The World Health Organization. The Immunological Basis for Immunization Series, in Meningococcal Disease. 2010. Available online: https://www.who.int/immunization/research/development/Meningococcal_disease_module15.pdf (accessed on 5 May 2021).
- Gioia, C.; Lemos, A.; Gorla, M.; Mendoza-Sassi, R.; Figueredo, B.; Ballester, T.; Von Groll, A.; Wedig, B.; Ethur, N.; Bragança, L.; et al. Seroprevalence of bactericidal antibodies against serogroup B and C Meningococci in a University Hospital. Braz. J. Med. Biol. Res. 2017, 50, 5590. [Google Scholar] [CrossRef] [Green Version]
- Feavers, I.; Walker, B. Functional Antibody Assays. Methods Mol. Biol. 2009, 626, 199–211. [Google Scholar] [CrossRef]
- Townsend, K.; Ladhani, S.N.; Findlow, H.; Borrow, R. Evaluation and validation of a serum bactericidal antibody assay for Haemophilus influenzae type b and the thresh-old of protection. Vaccine 2014, 32, 5650–5656. [Google Scholar] [CrossRef]
- Kim, H.W.; Kim, K.-H.; Kim, J.; Nahm, M.H. A high throughput serum bactericidal assay for antibodies to Haemophilus influenzae type b. BMC Infect. Dis. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findlow, J.; Balmer, P.; Borrow, R. A review of complement sources used in serum bactericidal assays for evaluating immune re-sponses to meningococcal ACWY conjugate vaccines. Hum. Vaccin Immunother. 2019, 15, 2491–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadarangani, M. Protection Against Invasive Infections in Children Caused by Encapsulated Bacteria. Front. Immunol. 2018, 9, 2674. [Google Scholar] [CrossRef] [Green Version]
- Plested, J.S.; Granoff, D.M. Vaccine-Induced Opsonophagocytic Immunity to Neisseria meningitidis Group B. Clin. Vaccine Immunol. 2008, 15, 799–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findlow, H.; Plikaytis, B.D.; Aase, A.; Bash, M.C.; Chadha, H.; Elie, C.; Laher, G.; Martinez, J.; Herstad, T.; Newton, E.; et al. Investigation of Different Group A Immunoassays following One Dose of Meningococcal Group A Conjugate Vaccine or A/C Polysaccharide Vaccine in Adults. Clin. Vaccine Immunol. 2009, 16, 969–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.; Pilishvili, T.; Barnard, S.; Caba, J.; Spear, W.; Romero-Steiner, S.; Carlone, G.M. Opsonophagocytosis of Fluorescent Polystyrene Beads Coupled to Neisseria meningitidis Serogroup A, C, Y, or W135 Polysaccharide Correlates with Serum Bactericidal Activity. Clin. Vaccine Immunol. 2002, 9, 485–488. [Google Scholar] [CrossRef] [Green Version]
- Aase, A. Comparison of functional immune responses in humans after intranasal and intramuscular immunisations with outer membrane vesicle vaccines against group B meningococcal disease. Vaccine 2003, 21, 2042–2051. [Google Scholar] [CrossRef]
- Broek, B.V.D.; Van Els, C.A.; Kuipers, B.; Van Aerde, K.; Henriet, S.S.; De Groot, R.; De Jonge, M.I.; Langereis, J.D.; Van Der Flier, M.; Els, C.A.C.M.; et al. Multi-component meningococcal serogroup B (MenB)-4C vaccine induces effective opsonophagocytic killing in children with a complement deficiency. Clin. Exp. Immunol. 2019, 198, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, S.K.; Smeesters, P.R.; Frost, H.R.C.; Licciardi, P.; Steer, A.C. Correlates of Protection for M Protein-Based Vaccines against Group A Streptococcus. J. Immunol. Res. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.J.; Noh, J.Y.; Cheong, H.J.; Kim, W.J.; Lin, S.M.; Zhi, Y.; Lim, J.H.; Lim, S.; Seo, H.S.; Song, J.Y. Evelopment of a multiplexed opsonophagocytic killing assay (MOPA) for group B Streptococcus. Hum. Vaccin Immunother. 2018, 14, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Moreland, N.; Zancolli, M.; Raynes, J.; Loh, J.M.; Smeesters, P.R.; Sriskandan, S.; Carapetis, J.R.; Fraser, J.D.; Goldblatt, D. Development of an opsonophagocytic killing assay for group a streptococcus. Vaccine 2018, 36, 3756–3763. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Assays | Advantages | Disadvantages |
|---|---|---|
| Traditional Killing OPA/MOPA pneumococcal-specific |
|
|
| Fluorescent OPA/MOPA pneumococcal-specific |
|
|
| Serum Bactericidal Assay Hib and meningococcal |
|
|
| Antibody Avidity Assay pneumococcal, Hib and meningococcal |
|
|
| Vaccines | Correlates of Protection |
|---|---|
| PCV | ELISA >0.35 µg/mL OPA ≥8 titre |
| Hib | ELISA Long term: ≥1.0 µg/mL Short term: >0.15 µg/mL SBA ≥4 titre |
| Meningococcal * | SBA rSBA (≥8 titre) or hSBA (≥4 titre) |
| Bacterial | Assay | Bacterial Strains | Complement Source | Immune Cells | Reference |
|---|---|---|---|---|---|
| Pneumococcal | OPA | Serotype-specific strains available through BEI resources | Baby Rabbit complement | HL-60 | [11] |
| Meningococcal A and C | SBA | Serogroup A strain F8238 Serogroup C strain C11 | Baby Rabbit complement | NA | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toh, Z.Q.; Higgins, R.A.; Mazarakis, N.; Abbott, E.; Nathanielsz, J.; Balloch, A.; Mulholland, K.; Licciardi, P.V. Evaluating Functional Immunity Following Encapsulated Bacterial Infection and Vaccination. Vaccines 2021, 9, 677. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060677
Toh ZQ, Higgins RA, Mazarakis N, Abbott E, Nathanielsz J, Balloch A, Mulholland K, Licciardi PV. Evaluating Functional Immunity Following Encapsulated Bacterial Infection and Vaccination. Vaccines. 2021; 9(6):677. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060677
Chicago/Turabian StyleToh, Zheng Quan, Rachel A. Higgins, Nadia Mazarakis, Elysia Abbott, Jordan Nathanielsz, Anne Balloch, Kim Mulholland, and Paul V. Licciardi. 2021. "Evaluating Functional Immunity Following Encapsulated Bacterial Infection and Vaccination" Vaccines 9, no. 6: 677. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060677