
A Novel Recombinant Virus-Like Particles Displaying B and T Cell Epitopes of Japanese Encephalitis Virus Offers Protective Immunity in Mice and Guinea Pigs

 , ,
, ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells, Viruses, Mice, and Guinea Pigs
2.3. Design and Synthesis of Novel VLP Displaying B and T Cell Epitopes of JEV (VLP-JEVe)
2.4. Plasmid Construction
2.5. Expression, Purification, and Identification of Recombinant Protein VP2-JEVe
2.6. VLP Assembly
2.7. Immunization and Challenge in Mice with VLP-JEVe
2.8. Immunization and Challenge in Guinea Pigs with VLP-JEVe
2.9. Antibody Testing
2.10. Cytokine and Cytotoxic T Lymphocyte Assay
2.11. Spleen Lymphocyte Proliferation Assay
2.12. Determination of PPV in Tissue
2.13. Statistical Analyses
3. Results
3.1. Expression and Purification Assembly of VLP-JEVe
3.2. Antibody Response in Mice
3.3. Cytokine and Cytotoxic T Lymphocyte Assays in Immunized Mice
3.4. Protection against JEV Challenge in Mice
3.5. Antibody Response in Guinea Pigs
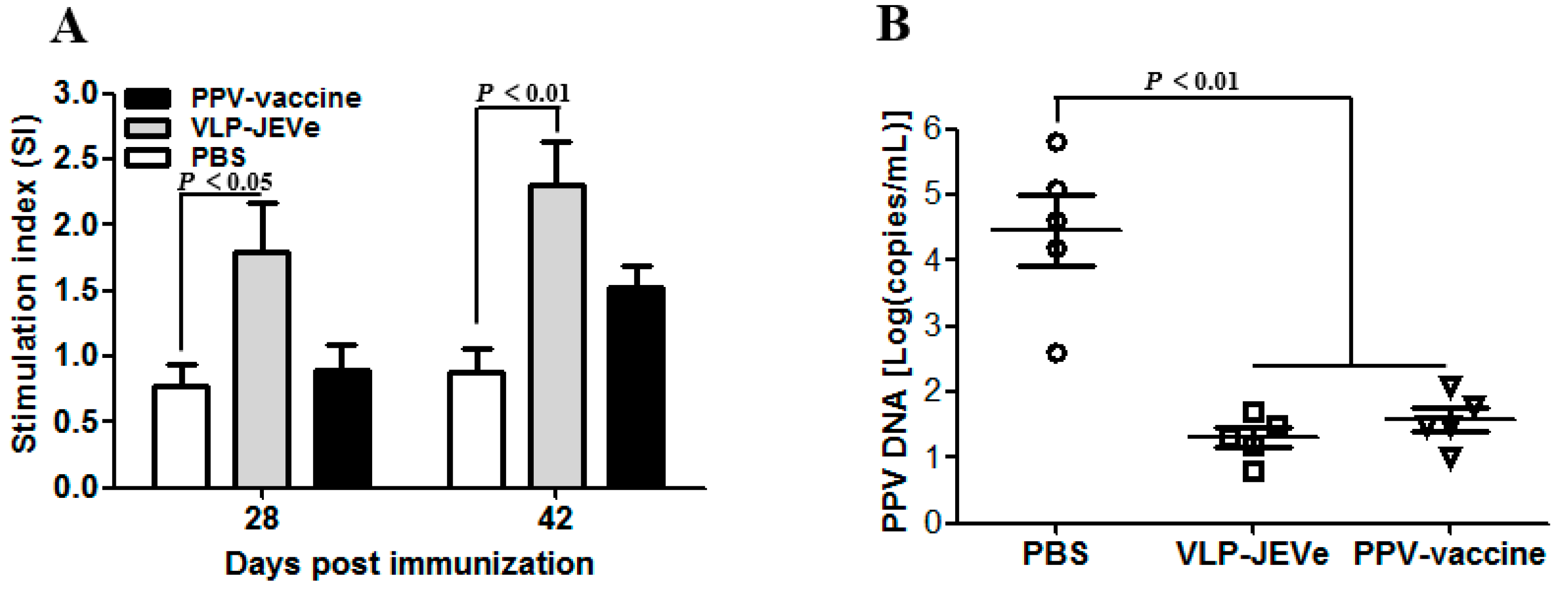
3.6. T Lymphocyte Proliferation and the Dynamic Changes in PPV Contents in the Spleen of Immunized Guinea Pigs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774, 774a–774e. [Google Scholar] [CrossRef]
- Konno, J.; Endo, K.; Agatsuma, H.; Ishida, N. Cyclic outbreaks of Japanese encephalitis among pigs and humans. Am. J. Epidemiol. 1966, 84, 292–300. [Google Scholar] [CrossRef]
- Unni, S.K.; Růžek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese encephalitis virus: From genome to infectome. Microbes Infect. 2011, 13, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef]
- Hollidge, B.S.; González-Scarano, F.; Soldan, S.S. Arboviral encephalitides: Transmission, emergence, and pathogenesis. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2010, 5, 428–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Liu, Z.J.; Jing, J.; Ren, J.Q.; Liu, Y.Y.; Guo, H.H.; Fan, M.; Lu, H.J.; Jin, N.Y. Reverse transcription loop-mediated isothermal amplification for rapid detection of Japanese encephalitis virus in swine and mosquitoes. Vector Borne Zoonotic Dis. 2012, 12, 1042–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccin Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Li, X.F.; Ye, Q.; Wang, H.J.; Deng, Y.Q.; Zhu, S.Y.; Zhang, Y.; Li, S.H.; Qin, C.F. Characterization of live-attenuated Japanese encephalitis vaccine virus SA14-14-2. Vaccine 2014, 32, 2675–2681. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Wang, X.; Zhang, J.; Guo, S.; Pang, L.; Shi, K.; Liu, K.; Shao, D.; Qiu, Y.; Liu, L.; et al. Partial cross-protection between Japanese encephalitis virus genotype I and III in mice. PLoS Negl. Trop. Dis. 2019, 13, e0007601. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Li, C.; Di, D.; Cappelle, J.; Liu, L.; Wang, X.; Pang, L.; Xu, J.; Liu, K.; Li, B.; et al. Differential replication efficiencies between Japanese encephalitis virus genotype I and III in avian cultured cells and young domestic ducklings. PLoS Negl. Trop. Dis. 2018, 12, e0007046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hills, S.L.; Walter, E.B.; Atmar, R.L.; Fischer, M.; Group, A.J.E.V.W.; Group, A.J.E.V.W.; Walter Jr, E.; Atmar, R.L.; Barnett, E.; Barrett, A. Japanese encephalitis vaccine: Recommendations of the advisory committee on immunization practices. MMWR Recomm. Rep. 2019, 68, 1. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.T.; Chang, L.C.; Lin, S.Y.; Chuang, T.Y.; Ma, C.H.; Inoue, Y.K.; Okuno, T. The effect of vaccination with a live attenuated strain of Japanese encephalitis virus on stillbirths in swine in Taiwan. Bull. World Health Organ. 1972, 46, 465–471. [Google Scholar] [PubMed]
- Arai, S.; Matsunaga, Y.; Takasaki, T.; Tanaka-Taya, K.; Taniguchi, K.; Okabe, N.; Kurane, I. Japanese encephalitis: Surveillance and elimination effort in Japan from 1982 to 2004. Jpn. J. Infect. Dis. 2008, 61, 333–338. [Google Scholar] [PubMed]
- Sedlik, C.; Saron, M.-F.; Sarraseca, J.; Casal, I.; Leclerc, C. Recombinant parvovirus-like particles as an antigen carrier: A novel nonreplicative exogenous antigen to elicit protective antiviral cytotoxic T cells. Proc. Natl. Acad. Sci. USA 1997, 94, 7503–7508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; He, K.; Huang, K. Development of recombinant porcine parvovirus-like particles as an antigen carrier formed by the hybrid VP2 protein carrying immunoreactive epitope of porcine circovirus type 2. Vaccine 2008, 26, 2119–2126. [Google Scholar] [CrossRef]
- Rueda, P.; Morón, G.; Sarraseca, J.; Leclerc, C.; Casal, J.I. Influence of flanking sequences on presentation efficiency of a CD8+ cytotoxic T-cell epitope delivered by parvovirus-like particles. J. Gen. Virol. 2004, 85, 563–572. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, J.M.; Song, B.H.; Yun, S.I.; Yun, G.N.; Byun, S.J.; Lee, Y.M. Profiling of viral proteins expressed from the genomic RNA of Japanese encephalitis virus using a panel of 15 region-specific polyclonal rabbit antisera: Implications for viral gene expression. PLoS ONE 2015, 10, e0124318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.L.; Chen, L.K.; Liao, C.L.; Yeh, C.T.; Ma, S.H.; Chen, J.L.; Huang, Y.L.; Chen, S.S.; Chiang, H.Y. DNA immunization with Japanese encephalitis virus nonstructural protein NS1 elicits protective immunity in mice. J. Virol. 1998, 72, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, F.F.; Qiu, Y.F.; Yang, Y.W.; Chen, P.Y. An hsp70 fusion protein vaccine potentiates the immune response against Japanese encephalitis virus. Arch. Virol. 2007, 152, 125–135. [Google Scholar] [CrossRef]
- Hueffer, K.; Parrish, C.R. Parvovirus host range, cell tropism and evolution. Curr. Opin. Microbiol. 2003, 6, 392–398. [Google Scholar] [CrossRef]
- Mengeling, W.L.; Lager, K.M.; Vorwald, A.C. The effect of porcine parvovirus and porcine reproductive and respiratory syndrome virus on porcine reproductive performance. Anim. Reprod. Sci. 2000, 60–61, 199–210. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef]
- Livingston, B.; Crimi, C.; Newman, M.; Higashimoto, Y.; Appella, E.; Sidney, J.; Sette, A. A rational strategy to design multiepitope immunogens based on multiple Th lymphocyte epitopes. J. Immunol. 2002, 168, 5499–5506. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Lu, Y.; Chen, Y. Candidate multi-epitope vaccines in aluminium adjuvant induce high levels of antibodies with predefined multi-epitope specificity against HIV-1. FEMS Immunol. Med. Microbiol. 2000, 29, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, S.; Zou, Y.; Yu, W.; Jiang, Y.; Zhan, Y.; Wang, N.; Dong, Y.; Yang, Y. Structure-Based Design of Porcine Circovirus Type 2 Chimeric VLPs (cVLPs) Displays Foreign Peptides on the Capsid Surface. Front. Cell. Infect. Microbiol. 2018, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Hameed, M.; Wang, X.; Zhang, J.; Guo, S.; Anwar, M.N.; Pang, L.; Liu, K.; Li, B.; Shao, D.; et al. Antiviral activity of phage display-selected peptides against Japanese encephalitis virus infection in vitro and in vivo. Antivir. Res. 2020, 174, 104673. [Google Scholar] [CrossRef]
- Fan, Y.C.; Chen, J.M.; Chiu, H.C.; Chen, Y.Y.; Lin, J.W.; Shih, C.C.; Chen, C.M.; Chang, C.C.; Chang, G.J.; Chiou, S.S. Partially neutralizing potency against emerging genotype I virus among children received formalin-inactivated Japanese encephalitis virus vaccine. PLoS Negl. Trop. Dis. 2012, 6, e1834. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.C.; Huang, Y.Z.; Zhong, D.K.; Kang, L.; Ishag, H.; Mao, X.; Cao, R.B.; Zhou, B.; Chen, P.Y. Design and evaluation of a multi-epitope peptide against Japanese encephalitis virus infection in BALB/c mice. Biochem. Biophys. Res. Commun. 2010, 396, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Liu, Y.; Chen, Y.; Wang, A.; Jiang, D.; Zhao, B.; Wang, J.; Chai, S.; Zhou, E.; Zhang, G. Porcine parvovirus capsid protein expressed in Escherichia coli self-assembles into virus-like particles with high immunogenicity in mice and guinea pigs. Antivir. Res. 2017, 139, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Donaldson-Wood, C.R.; Johnson, R.H. A standardised haemagglutination inhibition test for porcine parvovirus antibody. Aust. Vet. J. 1976, 52, 422–424. [Google Scholar] [CrossRef]
- Fischer, M.; Lindsey, N.; Staples, J.E.; Hills, S. Japanese encephalitis vaccines: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR. Recomm. Rep. Morb. Mortal. Wkly. Rep. Recomm. Rep. 2010, 59, 1–27. [Google Scholar]
- Liu, W.J.; Zhu, M.; Pei, J.J.; Dong, X.Y.; Liu, W.; Zhao, M.Q.; Wang, J.Y.; Gou, H.C.; Luo, Y.W.; Chen, J.D. Molecular phylogenetic and positive selection analysis of Japanese encephalitis virus strains isolated from pigs in China. Virus Res. 2013, 178, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Chiu, H.C.; Chen, L.K.; Chang, G.J.; Chiou, S.S. Formalin Inactivation of Japanese Encephalitis Virus Vaccine Alters the Antigenicity and Immunogenicity of a Neutralization Epitope in Envelope Protein Domain III. PLoS Negl. Trop. Dis. 2015, 9, e0004167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endy, T.P.; Nisalak, A. Japanese encephalitis virus: Ecology and epidemiology. Curr. Top. Microbiol. Immunol. 2002, 267, 11–48. [Google Scholar] [CrossRef] [PubMed]
- Chackerian, B. Virus-like particles: Flexible platforms for vaccine development. Expert Rev. Vaccines 2007, 6, 381–390. [Google Scholar] [CrossRef]
- Fiedler, J.D.; Higginson, C.; Hovlid, M.L.; Kislukhin, A.A.; Castillejos, A.; Manzenrieder, F.; Campbell, M.G.; Voss, N.R.; Potter, C.S.; Carragher, B.; et al. Engineered mutations change the structure and stability of a virus-like particle. Biomacromolecules 2012, 13, 2339–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsky, L.A.; Ault, K.A.; Wheeler, C.M.; Brown, D.R.; Barr, E.; Alvarez, F.B.; Chiacchierini, L.M.; Jansen, K.U. A controlled trial of a human papillomavirus type 16 vaccine. N. Engl. J. Med. 2002, 347, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Scolnick, E.M.; McLean, A.A.; West, D.J.; McAleer, W.J.; Miller, W.J.; Buynak, E.B. Clinical evaluation in healthy adults of a hepatitis B vaccine made by recombinant DNA. JAMA 1984, 251, 2812–2815. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Williams, J.N.; Rice, J.; Stevenson, F.K.; Heckels, J.E.; Christodoulides, M. A DNA fusion vaccine induces bactericidal antibodies to a peptide epitope from the PorA porin of Neisseria meningitidis. Infect. Immun. 2008, 76, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Vrati, S. Development of a recombinant vaccine against Japanese encephalitis. J. Neurovirol. 2003, 9, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Dent, M.; Lai, H.; Sun, H.; Chen, Q. Immunization of Zika virus envelope protein domain III induces specific and neutralizing immune responses against Zika virus. Vaccine 2017, 35, 4287–4294. [Google Scholar] [CrossRef] [PubMed]
- Young, C.L.; Lyons, A.C.; Hsu, W.W.; Vanlandingham, D.L.; Park, S.L.; Bilyeu, A.N.; Ayers, V.B.; Hettenbach, S.M.; Zelenka, A.M.; Cool, K.R.; et al. Protection of swine by potent neutralizing anti-Japanese encephalitis virus monoclonal antibodies derived from vaccination. Antivir. Res. 2020, 174, 104675. [Google Scholar] [CrossRef] [PubMed]
- Bharati, K.; Rani, R.; Vrati, S. Evaluation of Japanese encephalitis virus DNA vaccine candidates in rhesus monkeys [Macaca mulatta]. Vaccine 2009, 27, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, S.; Wang, D.; Li, F.; Liang, L.; Li, X.; Zou, Y.; Zhan, Y.; Chen, G.; Yu, W.; et al. Protective humoral immunity in guinea pigs induced by PCV2 virus-like particles displaying the B cell linear epitope ((228)QQITDA(233)) of PPV1. Vet. Microbiol. 2019, 235, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Wang, H.; Ouyang, W.; Wang, X.; Bi, Z.; Xia, X.; Wang, Y.; He, K. Immunogenicity of adenovirus-derived porcine parvovirus-like particles displaying B and T cell epitopes of foot-and-mouth disease. Vaccine 2016, 34, 578–585. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anwar, M.N.; Jiang, C.; Di, D.; Zhang, J.; Guo, S.; Wang, X.; Hameed, M.; Wahaab, A.; Shao, D.; Li, Z.; et al. A Novel Recombinant Virus-Like Particles Displaying B and T Cell Epitopes of Japanese Encephalitis Virus Offers Protective Immunity in Mice and Guinea Pigs. Vaccines 2021, 9, 980. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9090980
Anwar MN, Jiang C, Di D, Zhang J, Guo S, Wang X, Hameed M, Wahaab A, Shao D, Li Z, et al. A Novel Recombinant Virus-Like Particles Displaying B and T Cell Epitopes of Japanese Encephalitis Virus Offers Protective Immunity in Mice and Guinea Pigs. Vaccines. 2021; 9(9):980. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9090980
Chicago/Turabian StyleAnwar, Muhammad Naveed, Chunying Jiang, Di Di, Junjie Zhang, Shuang Guo, Xin Wang, Muddassar Hameed, Abdul Wahaab, Donghua Shao, Zongjie Li, and et al. 2021. "A Novel Recombinant Virus-Like Particles Displaying B and T Cell Epitopes of Japanese Encephalitis Virus Offers Protective Immunity in Mice and Guinea Pigs" Vaccines 9, no. 9: 980. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9090980