
Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development

 , and
, and
Abstract
:1. Introduction
2. Viral Hepatitis B and C
3. Antiviral Therapies
4. Fibrosis and HCC Screening in the Clinics—State of the Art
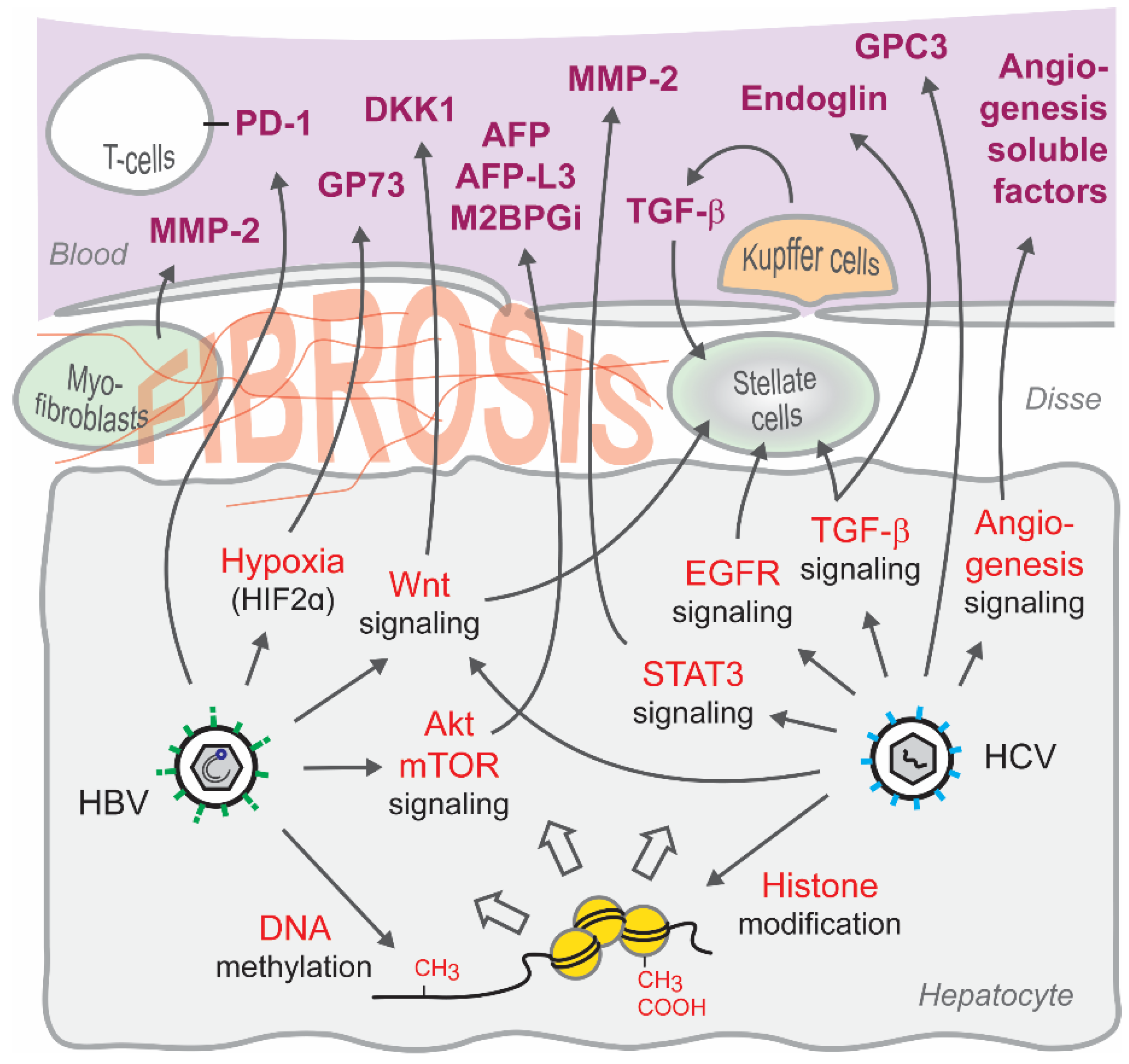
5. Signaling Pathways Associated with Candidate Serum Biomarkers
6. Virus-Induced Epigenetic Changes as Biomarkers
7. Discussion and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of Liver Diseases in the World. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Effenberger, M. From NAFLD to MAFLD: When Pathophysiology Succeeds. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 387–388. [Google Scholar] [CrossRef]
- Baumert, T.F.; Berg, T.; Lim, J.K.; Nelson, D.R. Status of Direct-Acting Antiviral Therapy for Hepatitis C Virus Infection and Remaining Challenges. Gastroenterology 2019, 156, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Chung, R.T.; Baumert, T.F. Curing Chronic Hepatitis C—The Arc of a Medical Triumph. N. Engl. J. Med. 2014, 370, 1576–1578. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD Development and Therapeutic Strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Villanueva, A. Hepatocellular Carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef] [Green Version]
- Tang, A.; Hallouch, O.; Chernyak, V.; Kamaya, A.; Sirlin, C.B. Epidemiology of Hepatocellular Carcinoma: Target Population for Surveillance and Diagnosis. Abdom. Radiol. 2018, 43, 13–25. [Google Scholar] [CrossRef]
- Desai, A.; Sandhu, S.; Lai, J.-P.; Sandhu, D.S. Hepatocellular Carcinoma in Non-cirrhotic Liver: A Comprehensive Review. World J. Hepatol. 2019, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Hepatitis Report; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Bandiera, S.; Pernot, S.; El Saghire, H.; Durand, S.C.; Thumann, C.; Crouchet, E.; Ye, T.; Fofana, I.; Oudot, M.A.; Barths, J.; et al. Hepatitis C Virus-Induced Upregulation of MicroRNA miR-146a-5p in Hepatocytes Promotes Viral Infection and Deregulates Metabolic Pathways Associated with Liver Disease Pathogenesis. J. Virol. 2016, 90, 6387–6400. [Google Scholar] [CrossRef] [Green Version]
- Diao, J.; Pantua, H.; Ngu, H.; Komuves, L.; Diehl, L.; Schaefer, G.; Kapadia, S.B. Hepatitis C Virus Induces Epidermal Growth Factor Receptor Activation via CD81 Binding for Viral Internalization and Entry. J. Virol. 2012, 86, 10935–10949. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, B.C.; Hoshida, Y.; Fujii, T.; Wei, L.; Yamada, S.; Lauwers, G.Y.; McGinn, C.M.; Deperalta, D.K.; Chen, X.; Kuroda, T.; et al. Epidermal Growth Factor Receptor Inhibition Attenuates Liver Fibrosis and Development of Hepatocellular Carcinoma. Hepatology 2014, 59, 1577–1590. [Google Scholar] [CrossRef] [Green Version]
- Igloi, Z.; Kazlauskas, A.; Saksela, K.; Macdonald, A.; Mankouri, J.; Harris, M. Hepatitis C virus NS5A Protein Blocks Epidermal Growth Factor Receptor Degradation via a Proline Motif-Dependent Interaction. J. Gen. Virol. 2015, 96, 2133–2144. [Google Scholar] [CrossRef]
- Lou, W.; Liu, J.; Ding, B.; Chen, D.; Xu, L.; Ding, J.; Jiang, D.; Zhou, L.; Zheng, S.; Fan, W. Identification of Potential miRNA–mRNA Regulatory Network Contributing to Pathogenesis of HBV-Related HCC. J. Transl. Med. 2019, 17, 1–14. [Google Scholar] [CrossRef]
- Lupberger, J.; Croonenborghs, T.; Suarez, A.A.R.; Van Renne, N.; Jühling, F.; Oudot, M.A.; Virzì, A.; Bandiera, S.; Jamey, C.; Meszaros, G.; et al. Combined Analysis of Metabolomes, Proteomes, and Transcriptomes of Hepatitis C Virus–Infected Cells and Liver to Identify Pathways Associated with Disease Development. Gastroenterology 2019, 157, 537–551.e9. [Google Scholar] [CrossRef] [Green Version]
- Van Renne, N.; Suarez, A.A.R.; Duong, F.H.T.; Gondeau, C.; Calabrese, D.; Fontaine, N.; Ababsa, A.; Bandiera, S.; Croonenborghs, T.; Pochet, N.; et al. miR-135a-5p-Mediated Downregulation of Protein Tyrosine Phosphatase Receptor Delta Is a Candidate Driver of HCV-Associated Hepatocarcinogenesis. Gut 2017, 67, 953–962. [Google Scholar] [CrossRef]
- Yavuz, B.G.; Pestana, R.C.; AbuGabal, Y.I.; Krishnan, S.; Chen, J.; Hassan, M.M.; Wolff, R.A.; Rashid, A.; Amin, H.M.; Kaseb, A.O. Origin and Role of Hepatic Myofibroblasts in Hepatocellular Carcinoma. Oncotarget 2020, 11, 1186–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshida, Y.; Villanueva, A.; Kobayashi, M.; Peix, J.; Chiang, D.Y.; Camargo, A.; Gupta, S.; Moore, J.; Wrobel, M.J.; Lerner, J.; et al. Gene Expression in Fixed Tissues and Outcome in Hepatocellular Carcinoma. N. Engl. J. Med. 2008, 359, 1995–2004. [Google Scholar] [CrossRef] [Green Version]
- Hoshida, Y.; Villanueva, A.; SanGiovanni, A.; Sole, M.; Hur, C.; Andersson, K.L.; Chung, R.T.; Gould, J.; Kojima, K.; Gupta, S.; et al. Prognostic Gene Expression Signature for Patients with Hepatitis C–Related Early-Stage Cirrhosis. Gastroenterology 2013, 144, 1024–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castera, L.; Friedrich-Rust, M.; Loomba, R. Noninvasive Assessment of Liver Disease in Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2019, 156, 1264–1281.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, J.; Reeves, H.L.; Feldstein, E.A. Liquid Biopsy for Liver Diseases. Gut 2018, 67, 2204–2212. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium Taurocholate Cotransporting Polypeptide Is a Functional Receptor for Human Hepatitis B and D Virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Saso, W.; Sugiyama, R.; Ishii, K.; Ohki, M.; Nagamori, S.; Suzuki, R.; Aizaki, H.; Ryo, A.; Yun, J.-H.; et al. Epidermal Growth Factor Receptor Is a Host-Entry Cofactor Triggering Hepatitis B Virus Internalization. Proc. Natl. Acad. Sci. USA 2019, 116, 8487–8492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukuda, S.; Watashi, K. Hepatitis B Virus Biology and Life Cycle. Antivir. Res. 2020, 182, 104925. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Arzberger, S.; Durantel, D.; Belloni, L.; Strubin, M.; Levrero, M.; Zoulim, F.; Hantz, O.; Protzer, U. Hepatitis B Virus X Protein Is Essential to Initiate and Maintain Virus Replication after Infection. J. Hepatol. 2011, 55, 996–1003. [Google Scholar] [CrossRef]
- Colpitts, C.C.; Tsai, P.-L.; Zeisel, M.B. Hepatitis C Virus Entry: An Intriguingly Complex and Highly Regulated Process. Int. J. Mol. Sci. 2020, 21, 2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabata, K.; Neufeldt, C.J.; Bartenschlager, R. Hepatitis C Virus Replication. Cold Spring Harb. Perspect. Med. 2019, 10, a037093. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Sun, F.; Wang, L.; Gao, M.; Xie, Y.; Sun, Y.; Liu, H.; Yuan, Y.; Yi, W.; Huang, Z.; et al. Virus-Induced p38 MAPK Activation Facilitates Viral Infection. Theranostics 2020, 10, 12223–12240. [Google Scholar] [CrossRef]
- Negash, A.A.; Olson, R.M.; Griffin, S.; Gale, M. Modulation of Calcium Signaling Pathway by Hepatitis C Virus Core Protein Stimulates NLRP3 Inflammasome Activation. PLoS Pathog. 2019, 15, e1007593. [Google Scholar] [CrossRef] [PubMed]
- Virzì, A.; Suarez, A.A.R.; Baumert, T.F.; Lupberger, J. Oncogenic Signaling Induced by HCV Infection. Viruses 2018, 10, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, A.; Yang, J.; Jo, E.; Park, K.H.P.; Kim, H.; Than, T.T.; Song, X.; Qi, X.; Dai, X.; Park, S.; et al. Efficient Long-Term Amplification of Hepatitis B Virus Isolates after Infection of Slow Proliferating HepG2-NTCP Cells. J. Hepatol. 2019, 71, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Fanning, G.C.; Zoulim, F.; Hou, J.; Bertoletti, A. Therapeutic Strategies for Hepatitis B Virus Infection: Towards a Cure. Nat. Rev. Drug Discov. 2019, 18, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Lok, A.S.; Zoulim, F.; Dusheiko, G.; Ghany, M.G. Hepatitis B Cure: From Discovery to Regulatory Approval. Hepatology 2017, 66, 1296–1313. [Google Scholar] [CrossRef]
- Mu, D.; Yuan, F.-C.; Chen, Y.; Jiang, X.-Y.; Yan, L.; Jiang, L.-Y.; Gong, J.-P.; Zhang, D.-Z.; Ren, H.; Liao, Y. Baseline Value of Intrahepatic HBV DNA Over cccDNA Predicts Patient’s Response to Interferon Therapy. Sci. Rep. 2017, 7, 5937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassal, M. HBV cccDNA: Viral Persistence Reservoir and Key Obstacle for a Cure of Chronic Hepatitis B. Gut 2015, 64, 1972–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Lu, W.; Zheng, Y.; Wang, W.; Bai, L.; Chen, L.; Feng, Y.; Zhang, Z.; Yuan, Z. In Situ Analysis of Intrahepatic Virological Events in Chronic Hepatitis B Virus Infection. J. Clin. Investig. 2016, 126, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. Electronic Address Eee, Clinical Practice Guideline Panel C, Panel m, Representative EGB. EASL Clinical Practice Guidelines: Drug-Induced Liver Injury. J. Hepatol. 2020, 73, 1170–1218. [Google Scholar] [CrossRef]
- Lu, C.Y.; Ross-Degnan, D.; Zhang, F.; LeCates, R.; Lupton, C.; Sherman, M.; Wagner, A. Cost Burden of Hepatitis C Virus Treatment in Commercially Insured Patients. Am J Manag Care 2019, 25, e379–e387. [Google Scholar]
- Baumert, T.F.; Jühling, F.; Ono, A.; Hoshida, Y. Hepatitis C-Related Hepatocellular Carcinoma in the Era of New Generation Antivirals. BMC Med. 2017, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.L.; Mo, F.; Johnson, P.J.; Siu, D.Y.W.; Chan, M.H.M.; Lau, W.Y.; Lai, P.B.S.; Lam, C.W.K.; Yeo, W.; Yu, S.C.H. Performance of Serum α-Fetoprotein Levels in the Diagnosis of Hepatocellular Carcinoma in Patients with a Hepatic Mass. HPB 2014, 16, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Manduca, A.; Bayly, P.J.; Ehman, R.L.; Kolipaka, A.; Royston, T.J.; Sack, I.; Sinkus, R.; Van Beers, B.E. MR Elastography: Principles, Guidelines, and Terminology. Magn. Reson. Med. 2021, 85, 2377–2390. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver; Asociación Latinoamericana para el Estudio del Hígado. EASL-ALEH Clinical Practice Guidelines: Non-Invasive Tests for Evaluation of Liver Disease Severity and Prognosis. J. Hepatol. 2015, 63, 237–264. [Google Scholar] [CrossRef] [Green Version]
- Tsoris, A.; Marlar, C.A. Use of the Child Pugh Score in Liver Disease; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Kubota, N.; Fujiwara, N.; Hoshida, Y. Clinical and Molecular Prediction of Hepatocellular Carcinoma Risk. J. Clin. Med. 2020, 9, 3843. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. Electronic Address Eee, European Association for the Study of the L. EASL Clinical Practice Guidelines: Management of Hepatocellular Carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [Green Version]
- Heimbach, J.K.; Kulik, L.M.; Finn, R.S.; Sirlin, C.B.; Abecassis, M.M.; Roberts, L.R.; Zhu, A.X.; Murad, M.H.; Marrero, J.A. AASLD Guidelines for the Treatment of Hepatocellular Carcinoma. Hepatology 2018, 67, 358–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertle, J.M.; Heider, D.; Wichert, M.; Keller, B.; Kueper, R.; Hilgard, P.; Gerken, G.; Schlaak, J.F. A Combination of A-Fetoprotein and Des-γ-Carboxy Prothrombin Is Superior in Detection of Hepatocellular Carcinoma. Digestion 2013, 87, 121–131. [Google Scholar] [CrossRef]
- Loglio, A.; Iavarone, M.; Facchetti, F.; Di Paolo, D.; Perbellini, R.; Lunghi, G.; Ceriotti, F.; Galli, C.; Sandri, M.T.; Viganò, M.; et al. The Combination of PIVKA-II and AFP Improves the Detection Accuracy for HCC in HBV Caucasian Cirrhotics on Long-Term Oral Therapy. Liver Int. 2020, 40, 1987–1996. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Wang, H.; Li, Y.; Zheng, L.; Yin, Y.; Zou, Z.; Zhou, F.; Zhou, W.; Shen, F.; Gao, C. Diagnostic Evaluation of Des-Gamma-Carboxy Prothrombin versus α-Fetoprotein for Hepatitis B Virus-Related Hepatocellular Carcinoma in China: A Large-Scale, Multicentre Study. PLoS ONE 2016, 11, e0153227. [Google Scholar] [CrossRef]
- Song, T.; Wang, L.; Su, B.; Zeng, W.; Jiang, T.; Zhang, T.; Sun, G.; Wu, H. Diagnostic Value of Alpha-Fetoprotein, Lens Culinaris Agglutinin-Reactive Alpha-Fetoprotein, and Des-Gamma-Carboxyprothrombin in Hepatitis B Virus-Related Hepatocellular Carcinoma. J. Int. Med. Res. 2020, 48, 0300060519889270. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, Y.; Yang, N.; He, H.; Tao, X.; Kou, C.; Jiang, J. Evaluation of the Combined Application of AFP, AFP-L3%, and DCP for Hepatocellular Carcinoma Diagnosis: A Meta-Analysis. BioMed Res. Int. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Ikoma, J.; Kaito, M.; Ishihara, T.; Nakagawa, N.; Kamei, A.; Fujita, N.; Iwasa, M.; Tamaki, S.; Watanabe, S.; Adachi, Y. Early Diagnosis of Hepatocellular Carcinoma Using a Sensitive Assay for Serum Des-Gamma-Carboxy Prothrombin: A Prospective Study. Hepatogastroenterology 2002, 49, 235–238. [Google Scholar]
- Fasolato, S.; Pigozzo, S.; Pontisso, P.; Angeli, P.; Ruscica, M.; Savarino, E.; De Martin, S.; Lupo, M.G.; Ferri, N. PCSK9 Levels Are Raised in Chronic HCV Patients with Hepatocellular Carcinoma. J. Clin. Med. 2020, 9, 3134. [Google Scholar] [CrossRef]
- Wiciński, M.; Żak, J.; Malinowski, B.; Popek, G.; Grześk, G. PCSK9 Signaling Pathways and Their Potential Importance in Clinical Practice. EPMA J. 2017, 8, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.-Y.; Lin, B.-D.; Li, B.-R. Evaluation of the Diagnostic Value of Alpha-L-Fucosidase, Alpha-Fetoprotein and Thymidine Kinase 1 with Roc and Logistic Regression for Hepatocellular Carcinoma. FEBS Open Bio 2015, 5, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Moudi, B.; Heidari, Z.; Mahmoudzadeh-Sagheb, H.; Alavian, S.-M.; Lankarani, K.B.; Farrokh, P.; Nyengaard, J.R. Concomitant Use of Heat-Shock Protein 70, Glutamine Synthetase and Glypican-3 Is Useful in Diagnosis of HBV-Related Hepatocellular Carcinoma with Higher Specificity and Sensitivity. Eur. J. Histochem. 2018, 62, 2859. [Google Scholar] [CrossRef] [PubMed]
- Murata, A.; Genda, T.; Ichida, T.; Amano, N.; Sato, S.; Tsuzura, H.; Sato, S.; Narita, Y.; Kanemitsu, Y.; Shimada, Y.; et al. Pretreatment AKR1B10 Expression Predicts the Risk of Hepatocellular Carcinoma Development after Hepatitis C Virus Eradication. World J. Gastroenterol. 2016, 22, 7569–7578. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Zhang, L.; Lv, S.; Zhang, C.; Jiang, S. Biomarkers for Hepatocellular Carcinoma. Biomark. Cancer 2017, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Best, J.; Bechmann, L.P.; Sowa, J.-P.; Sydor, S.; Dechêne, A.; Pflanz, K.; Bedreli, S.; Schotten, C.; Geier, A.; Berg, T.; et al. GALAD Score Detects Early Hepatocellular Carcinoma in an International Cohort of Patients with Nonalcoholic Steatohepatitis. Clin. Gastroenterol. Hepatol. 2020, 18, 728–735.e4. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.J.; Pirrie, S.J.; Cox, T.F.; Berhane, S.; Teng, M.; Palmer, D.; Morse, J.; Hull, D.; Patman, G.; Kagebayashi, C.; et al. The Detection of Hepatocellular Carcinoma Using a Prospectively Developed and Validated Model Based on Serological Biomarkers. Cancer Epidemiol. Biomark. Prev. 2014, 23, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.D.; Addissie, B.D.; Mara, K.C.; Harmsen, W.S.; Dai, J.; Zhang, N.; Wongjarupong, N.; Ali, H.M.; Ali, H.A.; Hassan, F.A.; et al. GALAD Score for Hepatocellular Carcinoma Detection in Comparison with Liver Ultrasound and Proposal of GALADUS Score. Cancer Epidemiol. Biomark. Prev. 2019, 28, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.J.; Berhane, S.; Kagebayashi, C.; Satomura, S.; Teng, M.; Reeves, H.L.; O’Beirne, J.; Fox, R.; Skowronska, A.; Palmer, D.; et al. Assessment of Liver Function in Patients With Hepatocellular Carcinoma: A New Evidence-Based Approach—The ALBI Grade. J. Clin. Oncol. 2015, 33, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Kariyama, K.; Nouso, K.; Hiraoka, A.; Wakuta, A.; Oonishi, A.; Kuzuya, T.; Toyoda, H.; Tada, T.; Tsuji, K.; Itobayashi, E.; et al. EZ-ALBI Score for Predicting Hepatocellular Carcinoma Prognosis. Liver Cancer 2020, 9, 734–743. [Google Scholar] [CrossRef]
- Dieter, P.; Altin, J.G.; Decker, K.; Bygrave, F.L. Possible Involvement of Eicosanoids in the Zymosan and Arachidonic-Acid-Induced Oxygen Uptake, Glycogenolysis and Ca2+ Mobilization in the Perfused Rat Liver. JBIC J. Biol. Inorg. Chem. 1987, 165, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, W.; Voelker, M.; Thiel, R.; Becka, M.; Burtb, A.; Schuppan, D.; Hubscher, S.; Roskams, T.; Pinzani, M.; Arthur, M.J. Serum Markers Detect the Presence of Liver Fibrosis: A Cohort Study. Gastroenterology 2004, 127, 1704–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkes, J.; Guha, I.N.; Roderick, P.; Harris, S.; Cross, R.; Manos, M.M.; Irving, W.; Zaitoun, A.; Wheatley, M.; Ryder, S.; et al. Enhanced Liver Fibrosis (ELF) Test Accurately Identifies Liver Fibrosis in Patients with Chronic Hepatitis C. J. Viral Hepat. 2010, 18, 23–31. [Google Scholar] [CrossRef]
- Vali, Y.; Lee, J.; Boursier, J.; Spijker, R.; Löffler, J.; Verheij, J.; Brosnan, M.J.; Böcskei, Z.; Anstee, Q.M.; Bossuyt, P.M.; et al. Enhanced Liver Fibrosis Test for the Non-invasive Diagnosis of Fibrosis in Patients With NAFLD: A Systematic Review and Meta-Analysis. J. Hepatol. 2020, 73, 252–262. [Google Scholar] [CrossRef]
- Lemmon, M.A.; Schlessinger, J. Cell Signaling by Receptor Tyrosine Kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [Green Version]
- Krump, N.A.; You, J. Molecular Mechanisms of Viral Oncogenesis in Humans. Nat. Rev. Genet. 2018, 16, 684–698. [Google Scholar] [CrossRef]
- Cong, M.; Iwaisako, K.; Jiang, C.; Kisseleva, T. Cell Signals Influencing Hepatic Fibrosis. Int. J. Hepatol. 2012, 2012, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Szabó, E.; Páska, C.; Novák, P.K.; Schaff, Z.; Kiss, A. Similarities and Differences in Hepatitis B and C Virus Induced Hepatocarcinogenesis. Pathol. Oncol. Res. 2004, 10, 5–11. [Google Scholar] [CrossRef]
- Faure-Dupuy, S.; Lucifora, J.; Durantel, D. Interplay between the Hepatitis B Virus and Innate Immunity: From an Understanding to the Development of Therapeutic Concepts. Viruses 2017, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suslov, A.; Boldanova, T.; Wang, X.; Wieland, S.; Heim, M.H. Hepatitis B Virus Does Not Interfere with Innate Immune Responses in the Human Liver. Gastroenterology 2018, 154, 1778–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Li, H.; Li, C.; Wang, S.; Jiang, W.; Liu, Z.; Zhou, S.; Liu, X.; McNutt, M.A.; Li, G. Alpha-Fetoprotein: A New Member of Intracellular Signal Molecules in Regulation of the PI3K/AKT Signaling in Human Hepatoma Cell Lines. Int. J. Cancer 2010, 128, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Guo, J.; Li, W.; Lu, Y.; Fu, S.; Xie, X.; Xia, H.; Dong, X.; Chen, Y.; Quan, M.; et al. Hepatitis B Virus X Protein Induces Expression of Alpha-Fetoprotein and Activates PI3K/MTOR Signaling Pathway in Liver Cells. Oncotarget 2015, 6, 12196–12208. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Cao, Z.; Cheng, Y.; Wang, J.; Liu, Y.; Yang, R.; Li, H.; Jiang, W.; Li, G.; Zhao, W.; et al. Acetylation of Alpha-Fetoprotein Promotes Hepatocellular Carcinoma Progression. Cancer Lett. 2020, 471, 12–26. [Google Scholar] [CrossRef]
- Golob-Schwarzl, N.; Krassnig, S.; Toeglhofer, A.M.; Park, Y.N.; Gogg-Kamerer, M.; Vierlinger, K.; Schröder, F.; Rhee, H.; Schicho, R.; Fickert, P.; et al. New Liver Cancer Biomarkers: PI3K/AKT/mTOR Pathway Members and Eukaryotic Translation Initiation Factors. Eur. J. Cancer 2017, 83, 56–70. [Google Scholar] [CrossRef]
- Zhang, R.; Lin, H.-M.; Broering, R.; Shi, X.-D.; Yu, X.-H.; Xu, L.-B.; Wu, W.-R.; Liu, C. Dickkopf-1 Contributes to Hepatocellular Carcinoma Tumorigenesis by Activating the Wnt/β-Catenin Signaling Pathway. Signal Transduct. Target. Ther. 2019, 4, 1–10. [Google Scholar] [CrossRef]
- Fezza, M.; Moussa, M.; Aoun, R.; Haber, R.; Hilal, G. DKK1 Promotes Hepatocellular Carcinoma Inflammation, Migration and Invasion: Implication of TGF-β1. PLoS ONE 2019, 14, e0223252. [Google Scholar] [CrossRef] [Green Version]
- Zekri, A.-R.N.; El Kassas, M.; El Salam, E.S.; Hassan, R.M.; Mohanad, M.; Gabr, R.M.; Lotfy, M.M.; Abdel-Zaher, R.A.T.; Bahnassy, A.A.; Ahmed, O.S. The Possible Role of Dickkopf-1, Golgi Protein-73 and Midkine as Predictors of Hepatocarcinogenesis: A Review and an Egyptian Study. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Fan, J.; Yang, X.-R.; Tan, Y.; Zhao, W.; Xu, Y.; Wang, N.; Niu, Y.; Wu, Z.; Zhou, J.; et al. Serum DKK1 as a Protein Biomarker for the Diagnosis of Hepatocellular Carcinoma: A Large-Scale, Multicentre Study. Lancet Oncol. 2012, 13, 817–826. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.; Tang, J.; Tang, R.; Shan, X.; Zhang, W.; Chen, Q.; Zhou, F.; Chen, K.; Huang, A.; et al. Hepatitis C Virus Core Protein Activates Wnt/β-Catenin Signaling through Multiple Regulation of Upstream Molecules in the SMMC-7721 Cell Line. Arch. Virol. 2011, 156, 1013–1023. [Google Scholar] [CrossRef]
- Umer, M.; Qureshi, S.A.; Hashmi, Z.Y.; Raza, A.; Ahmad, J.; Rahman, M.; Iqbal, M. Promoter Hypermethylation of Wnt Pathway Inhibitors in Hepatitis C Virus-Induced Multistep Hepatocarcinogenesis. Virol. J. 2014, 11, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldeeb, M.K.; Magour, G.M.; Bedair, R.N.; Shamseya, M.M.; Hammouda, M.A. Study of Dickkopf-1 (DKK-1) in Patients with Chronic Viral Hepatitis C-Related Liver Cirrhosis with and without Hepatocellular Carcinoma. Clin. Exp. Hepatol. 2020, 6, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Essa, E.S.; Montaser, A.B.; Badawy, M.T.; Essa, A.S.; Mokhtar, M.S. DKK1 in Relation to HCV Induced Liver Cirrhosis and HCV Induced HCC Curative Resection. Acta Gastro-Enterol. Belg. 2016, 79, 309–313. [Google Scholar]
- Qiao, Y.; Chen, J.; Li, X.; Wei, H.; Xiao, F.; Chang, L.; Zhang, R.; Hao, X.; Wei, H. Serum gp73 is Also a Biomarker for Diagnosing Cirrhosis in Population with Chronic HBV Infection. Clin. Biochem. 2014, 47, 216–222. [Google Scholar] [CrossRef]
- Wei, M.; Xu, Z.; Pan, X.; Zhang, X.; Liu, L.; Yang, B.; Chen, Y. Serum GP73-An Additional Biochemical Marker for Liver Inflammation in Chronic HBV Infected Patients with Normal or Slightly Raised ALT. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Zhang, Y.; Shen, M.; Xu, H.; Li, Z.; He, N. Golgi Protein 73 and Its Diagnostic Value in Liver Diseases. Cell Prolif. 2019, 52, e12538. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhu, J.; Yang, J.; Li, X.; Yuan, J.; Wu, J.; Liu, Z. GP73 Facilitates Hepatitis B Virus Replication by Repressing the NF-kappaB Signaling Pathway. J. Med. Virol. 2020, 92, 3327–3335. [Google Scholar] [CrossRef]
- Yang, S.; Zeng, C.; Fang, X.; He, Q.; Liu, L.; Bao, S.; Pan, X.; Xiong, Z. Hepatitis B Virus Upregulates GP73 Expression by Activating the HIF-2α Signaling Pathway. Oncol. Lett. 2018, 15, 5264–5270. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, Z. HBV-Induced Immune Imbalance in the Development of HCC. Front. Immunol. 2019, 10, 2048. [Google Scholar] [CrossRef] [Green Version]
- Rivino, L.; Le Bert, N.; Gill, U.S.; Kunasegaran, K.; Cheng, Y.; Tan, D.Z.; Becht, E.; Hansi, N.K.; Foster, G.R.; Su, T.-H.; et al. Hepatitis B Virus–Specific T Cells Associate with Viral Control upon Nucleos(t)ide-Analogue Therapy Discontinuation. J. Clin. Investig. 2018, 128, 668–681. [Google Scholar] [CrossRef] [Green Version]
- Dolgormaa, G.; Harimoto, N.; Ishii, N.; Yamanaka, T.; Hagiwara, K.; Tsukagoshi, M.; Igarashi, T.; Watanabe, A.; Kubo, N.; Araki, K.; et al. Mac-2-Binding Protein Glycan Isomer Enhances the Aggressiveness of Hepatocellular Carcinoma by Activating mTOR Signaling. Br. J. Cancer 2020, 123, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Kuno, A.; Ikehara, Y.; Tanaka, Y.; Ito, K.; Matsuda, A.; Sekiya, S.; Hige, S.; Sakamoto, M.; Kage, M.; Mizokami, M.; et al. A Serum “Sweet-Doughnut” Protein Facilitates Fibrosis Evaluation and Therapy Assessment in Patients with Viral Hepatitis. Sci. Rep. 2013, 3, 1065. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, H. Development of M2BPGi: A Novel Fibrosis Serum GlyCo-biomarker for Chronic Hepatitis/Cirrhosis Diagnostics. Expert Rev. Proteom. 2015, 12, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Jekarl, D.W.; Choi, H.; Lee, S.; Kwon, J.H.; Lee, S.W.; Yu, H.; Kim, M.; Kim, Y.; Sung, P.S.; Yoon, S.K. Diagnosis of Liver Fibrosis with Wisteria Floribunda Agglutinin-Positive Mac-2 Binding Protein (WFA-M2BP) Among Chronic Hepatitis B Patients. Ann. Lab. Med. 2018, 38, 348–354. [Google Scholar] [CrossRef]
- Jun, T.; Hsu, Y.-C.; Ogawa, S.; Huang, Y.-T.; Yeh, M.-L.; Tseng, C.-H.; Huang, C.-F.; Tai, C.-M.; Dai, C.-Y.; Huang, J.-F.; et al. Mac-2 Binding Protein Glycosylation Isomer as a Hepatocellular Carcinoma Marker in Patients with Chronic Hepatitis B or C Infection. Hepatol. Commun. 2019, 3, 493–503. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Hu, H.-H.; Lee, M.-H.; Korenaga, M.; Jen, C.-L.; Batrla-Utermann, R.; Lu, S.-N.; Wang, L.-Y.; Mizokami, M.; Chen, C.-J.; et al. Serum Levels of M2BPGi as Short-Term Predictors of Hepatocellular Carcinoma in Untreated Chronic Hepatitis B Patients. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mak, L.-Y.; To, W.-P.; Wong, D.K.-H.; Fung, J.; Liu, F.; Seto, W.-K.; Lai, C.-L.; Yuen, M.-F. Serum Mac-2 Binding Protein Glycosylation Isomer Level Predicts Hepatocellular Carcinoma Development in E-negative Chronic Hepatitis B Patients. World J. Gastroenterol. 2019, 25, 1398–1408. [Google Scholar] [CrossRef]
- Hsu, Y.-C.; Jun, T.; Huang, Y.-T.; Yeh, M.-L.; Lee, C.-L.; Ogawa, S.; Cho, S.-H.; Lin, J.-T.; Yu, M.-L.; Nguyen, M.H.; et al. Serum M2BPGi Level and Risk of Hepatocellular Carcinoma after Oral Anti-viral Therapy in Patients with Chronic Hepatitis B. Aliment. Pharmacol. Ther. 2018, 48, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Tateyama, M.; Abiru, S.; Komori, A.; Nagaoka, S.; Saeki, A.; Hashimoto, S.; Sasaki, R.; Bekki, S.; Kugiyama, Y.; et al. Elevated Serum Levels of Wisteria Floribunda Agglutinin-Positive Human Mac-2 Binding Protein Predict the Development of Hepatocellular Carcinoma in Hepatitis C Patients. Hepatology 2014, 60, 1563–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirabe, K.; Bekki, Y.; Gantumur, D.; Araki, K.; Ishii, N.; Kuno, A.; Narimatsu, H.; Mizokami, M. Mac-2 Binding Protein Glycan Isomer (M2BPGI) Is a New Serum Biomarker for Assessing Liver Fibrosis: More Than a Biomarker of Liver Fibrosis. J. Gastroenterol. 2018, 53, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.S.; Salama, M.M.; Alhusseini, M.M.; A Mohamed, G. M2BPGi for Assessing Liver Fibrosis in Patients with Hepatitis C Treated with Direct-Acting Antivirals. World J. Gastroenterol. 2020, 26, 2864–2876. [Google Scholar] [CrossRef] [PubMed]
- Lupberger, J.; Zeisel, M.B.; Xiao, F.; Thumann, C.; Fofana, I.; Zona, L.; Davis, C.; Mee, C.J.; Turek, M.; Gorke, S.; et al. EGFR and EphA2 are Host Factors for Hepatitis C Virus Entry and Possible Targets for Antiviral Therapy. Nat. Med. 2011, 17, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zona, L.; Lupberger, J.; SidAhmed-Adrar, N.; Thumann, C.; Harris, H.J.; Barnes, A.; Florentin, J.; Tawar, R.G.; Xiao, F.; Turek, M.; et al. HRas Signal Transduction Promotes Hepatitis C Virus Cell Entry by Triggering Assembly of the Host Tetraspanin Receptor Complex. Cell Host Microbe 2013, 13, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Mailly, L.; Xiao, F.; Lupberger, J.; Wilson, G.K.; Aubert, P.; Duong, F.H.T.; Calabrese, D.; Leboeuf, C.; Fofana, I.; Thumann, C.; et al. Clearance of Persistent Hepatitis C Virus Infection in Humanized Mice Using a Claudin-1-Targeting Monoclonal Antibody. Nat. Biotechnol. 2015, 33, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seoane, J.; Gomis, R.R. TGF-β Family Signaling in Tumor Suppression and Cancer Progression. Cold Spring Harb. Perspect. Biol. 2017, 9, a022277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chusri, P.; Kumthip, K.; Hong, J.; Zhu, C.; Duan, X.; Jilg, N.; Fusco, D.N.; Brisac, C.; Schaefer, E.A.; Cai, D.; et al. HCV Induces Transforming Growth Factor β1 through Activation of Endoplasmic Reticulum Stress and the Unfolded Protein Response. Sci. Rep. 2016, 6, 22487. [Google Scholar] [CrossRef]
- Cheng, P.-L.; Chang, M.-H.; Chao, C.-H.; Lee, Y.-H.W. Hepatitis C Viral Proteins Interact with Smad3 and Differentially Regulate TFG-β/Smad3-Mediated Transcriptional Activation. Oncogene 2004, 23, 7821–7838. [Google Scholar] [CrossRef] [Green Version]
- Clemente, M.; Nunez, O.; Lorente, R.; Rincon, D.; Matilla, A.; Salcedo, M.; Catalina, M.V.; Ripoll, C.; Iacono, O.L.; Banares, R.; et al. Increased Intrahepatic and Circulating Levels of Endoglin, a TFG-β1 Co-receptor, in Patients with Chronic Hepatitis C Virus Infection: Relationship to Histological and Serum Markers of Hepatic Fibrosis. J. Viral Hepat. 2006, 13, 625–632. [Google Scholar] [CrossRef]
- Tardif, K.D.; Waris, G.; Siddiqui, A. Hepatitis C Virus, ER Stress, and Oxidative Stress. Trends Microbiol. 2005, 13, 159–163. [Google Scholar] [CrossRef]
- Abdalla, M.Y.; Mathahs, M.M.; Ahmad, I.M. Reduced Heme Oxygenase-1 Expression in Steatotic Livers Infected with Hepatitis C Virus. Eur. J. Intern. Med. 2012, 23, 649–655. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Karin, M. NF-κB and STAT3–Key Players in Liver Inflammation and Cancer. Cell Res. 2010, 21, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, Q.; Liu, Y.; Luo, Z.; Kang, L.; Qu, J.; Liu, W.; Xia, X.; Wu, K.; Wu, J. Hepatitis C Virus Activates Bcl-2 and MMP-2 Expression through Multiple Cellular Signaling Pathways. J. Virol. 2012, 86, 12531–12543. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Latif, M.S. Plasma Levels of Matrix Metalloproteinase (MMP)-2, MMP-9 and Tumor Necrosis Factor-α in Chronic Hepatitis C Virus Patients. Open Microbiol. J. 2015, 9, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.K.; Brimacombe, C.L.; Rowe, I.A.; Reynolds, G.M.; Fletcher, N.F.; Stamataki, Z.; Bhogal, R.H.; Simões, M.L.; Ashcroft, M.; Afford, S.C.; et al. A Dual Role for Hypoxia Inducible Factor-1α in the Hepatitis C Virus Lifecycle and Hepatoma Migration. J. Hepatol. 2012, 56, 803–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas, V.R.; Maluf, D.G.; Archer, K.J.; Yanek, K.C.; Fisher, R.A. Angiogenesis Soluble Factors as Hepatocellular Carcinoma Noninvasive Markers for Monitoring Hepatitis C Virus Cirrhotic Patients Awaiting Liver Transplantation. Transplantation 2007, 84, 1262–1271. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Bartolomé, Á; López-Rodríguez, R.; Borque, M.J.; González-Moreno, L.; Real-Martínez, Y.; García-Buey, L.; Moreno-Otero, R.; Sanz-Cameno, P. Angiopoietin-2/Angiopoietin-1 as Non-invasive Biomarker of Cirrhosis in Chronic Hepatitis C. World J. Gastroenterol. 2016, 22, 9744–9751. [Google Scholar] [CrossRef]
- Hernández-Bartolomé, Á.; López-Rodríguez, R.; García-Buey, L.; Martín-Vílchez, S.; Rodríguez-Muñoz, Y.; Borque, M.J.; González-Moreno, L.; Real-Martínez, Y.; Mendoza-Ridruejo, J.; Martín-Pérez, E.; et al. Intrahepatic Angiopoietin-2 Correlates with Chronic Hepatitis C Progression and Is Induced in Hepatitis C Virus Replicon Systems. Liver Int. 2016, 37, 1148–1156. [Google Scholar] [CrossRef]
- Benkheil, M.; Van Haele, M.; Roskams, T.; Laporte, M.; Noppen, S.; Abbasi, K.; Delang, L.; Neyts, J.; Liekens, S. CCL20, A Direct-Acting Pro-angiogenic Chemokine Induced by Hepatitis C Virus (HCV): Potential Role in HCV-Related Liver Cancer. Exp. Cell Res. 2018, 372, 168–177. [Google Scholar] [CrossRef]
- Soliman, H.H.; Nagy, H.; Kotb, N.; El-Din, M.A.A. The Role of Chemokine CC Ligand 20 in Patients with Liver Cirrhosis and Hepatocellular Carcinoma. Int. J. Biol. Markers 2012, 27, 125–131. [Google Scholar] [CrossRef]
- Oliveira, D.A.R.; Castanhole-Nunes, M.M.; Biselli-Chicote, P.M.; Pavarino, É.C.; da Silva, R.C.M.A.; Da Silva, R.F.; Goloni-Bertollo, E.M. Differential Expression of Angiogenesis-Related miRNAs and VEGFA in Cirrhosis and Hepatocellular Carcinoma. Arch. Med. Sci. 2020, 16, 1150–1157. [Google Scholar] [CrossRef]
- Salum, G.M.; El Din, N.G.B.; Ibrahim, M.K.; Anany, M.A.; Dawood, R.M.; Khairy, A.; El Awady, M.K. Vascular Endothelial Growth Factor Expression in Hepatitis C Virus-Induced Liver Fibrosis: A Potential Biomarker. J. Interf. Cytokine Res. 2017, 37, 310–316. [Google Scholar] [CrossRef]
- Zhu, C.L.; Li, W.T.; Li, Y.; Gao, R.T. Serum Levels of Tissue Inhibitor of Metalloproteinase-1 Are Correlated with Liver Fibrosis in Patients with Chronic Hepatitis B. J. Dig. Dis. 2012, 13, 558–563. [Google Scholar] [CrossRef]
- Seven, G.; Karatayli, S.C.; Kose, S.K.; Yakut, M.; Kabacam, G.; Toruner, M.; Heper, A.O.; Voelker, M.; Erden, E.; Bozdayi, A.M.; et al. Serum Connective Tissue Markers as Predictors of Advanced Fibrosis in Patients with Chronic Hepatitis B and D. Turk. J. Gastroenterol. 2011, 22, 305–314. [Google Scholar] [CrossRef]
- Xue, Y.; Mars, W.M.; Bowen, W.; Singhi, A.D.; Stoops, J.; Michalopoulos, G.K. Hepatitis C Virus Mimics Effects of Glypican-3 on CD81 and Promotes Development of Hepatocellular Carcinomas via Activation of Hippo Pathway in Hepatocytes. Am. J. Pathol. 2018, 188, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Mizuno, S.; Fujinami, N.; Suzuki, T.; Saito, K.; Konishi, M.; Takahashi, S.; Gotohda, N.; Tada, T.; Toyoda, H.; et al. Plasma and Tumoral Glypican-3 Levels Are Correlated in Patients with Hepatitis C Virus-Related Hepatocellular Carcinoma. Cancer Sci. 2019, 111, 334–342. [Google Scholar] [CrossRef]
- Jaenisch, R.; Bird, A. Epigenetic Regulation of Gene Expression: How the Genome Integrates Intrinsic and Environmental Signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Hardy, T.; Mann, A.D. Epigenetics in Liver Disease: From Biology to Therapeutics. Gut 2016, 65, 1895–1905. [Google Scholar] [CrossRef] [PubMed]
- Moran-Salvador, E.; Mann, J. Epigenetics and Liver Fibrosis. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.L.; Mann, D.A.; Borthwick, L.A. Epigenetic Reprogramming in Liver Fibrosis and Cancer. Adv. Drug Deliv. Rev. 2017, 121, 124–132. [Google Scholar] [CrossRef]
- Zhang, Y.; Petropoulos, S.; Liu, J.; Cheishvili, D.; Zhou, R.; Dymov, S.; Li, K.; Li, N.; Szyf, M. The Signature of Liver Cancer in Immune Cells DNA Methylation. Clin. Epigenetics 2018, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Qin, L.; Jiang, S.; Zhang, C.; Liu, G.; Sun, J.; Sun, H.; Zhao, Y.; Li, N.; Zhang, Y.; et al. The Signature of HBV-Related Liver Disease in Peripheral Blood Mononuclear Cell DNA Methylation. Clin. Epigenetics 2020, 12, 1–13. [Google Scholar] [CrossRef]
- Vivekanandan, P.; Daniel, H.D.-J.; Kannangai, R.; Martinez-Murillo, F.; Torbenson, M. Hepatitis B Virus Replication Induces Methylation of both Host and Viral DNA. J. Virol. 2010, 84, 4321–4329. [Google Scholar] [CrossRef] [Green Version]
- Ancey, P.-B.; Testoni, B.; Gruffaz, M.; Cros, M.-P.; Durand, G.; Le Calvez-Kelm, F.; Durantel, D.; Herceg, Z.; Hernandez-Vargas, H. Genomic Responses to Hepatitis B Virus (HBV) Infection in Primary Human Hepatocytes. Oncotarget 2015, 6, 44877–44891. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Hou, Q.; Chen, Y.; Wang, X. Diagnostic Value of the Methylation of Multiple Gene Promoters in Serum in Hepatitis B Virus-Related Hepatocellular Carcinoma. Dis. Markers 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Huang, Y.; Zhao, R.; Zhang, J.; Liu, Q.; Liang, W.; Ding, X.; Gao, B.; Li, B.; Sun, C.; et al. Detection of Promoter Methylation Status of Suppressor of Cytokine Signaling 3 (SOCS3) in Tissue and Plasma from Chinese Patients with Different Hepatic Diseases. Clin. Exp. Med. 2018, 18, 79–87. [Google Scholar] [CrossRef]
- Wu, H.-C.; Yang, H.-I.; Wang, Q.; Chen, C.-J.; Santella, R.M. Plasma DNA Methylation Marker and Hepatocellular Carcinoma Risk Prediction Model for the General Population. Carcinogenesis 2017, 38, 1021–1028. [Google Scholar] [CrossRef] [Green Version]
- Hamdane, N.; Jühling, F.; Crouchet, E.; El Saghire, H.; Thumann, C.; Oudot, M.A.; Bandiera, S.; Saviano, A.; Ponsolles, C.; Suarez, A.A.R.; et al. HCV-Induced Epigenetic Changes Associated with Liver Cancer Risk Persist After Sustained Virologic Response. Gastroenterology 2019, 156, 2313–2329.e7. [Google Scholar] [CrossRef] [Green Version]
- Juhling, F.; Hamdane, N.; Crouchet, E.; Li, S.; El Saghire, H.; Mukherji, A.; Fujiwara, N.; Oudot, M.A.; Thumann, C.; Saviano, A.; et al. Targeting Clinical Epigenetic Reprogramming for Chemoprevention of Metabolic and Viral Hepatocellular Carcinoma. Gut 2020, 70, 157–169. [Google Scholar] [CrossRef]
- Perez, S.; Kaspi, A.; Domovitz, T.; Davidovich, A.; Lavi-Itzkovitz, A.; Meirson, T.; Holmes, J.A.; Dai, C.-Y.; Huang, C.-F.; Chung, R.T.; et al. Hepatitis C Virus Leaves an Epigenetic Signature Post Cure of Infection by Direct-Acting Antivirals. PLoS Genet. 2019, 15, e1008181. [Google Scholar] [CrossRef] [PubMed]
- Shaker, O.G.; Senousy, M.A. Serum microRNAs as Predictors for Liver Fibrosis Staging in Hepatitis C Virus-Associated Chronic Liver Disease Patients. J. Viral Hepat. 2017, 24, 636–644. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Mackowiak, B.; Gao, B. MicroRNAs as Regulators, Biomarkers and Therapeutic Targets in Liver Diseases. Gut 2020, 10, 1136. [Google Scholar] [CrossRef]
- Thietart, S.; Rautou, P.-E. Extracellular Vesicles as Biomarkers in Liver Diseases: A Clinician’s Point of View. J. Hepatol. 2020, 73, 1507–1525. [Google Scholar] [CrossRef]
- Matsuura, K.; De Giorgi, V.; Schechterly, C.; Wang, R.Y.; Farci, P.; Tanaka, Y.; Alter, H.J. Circulating Let-7 Levels in Plasma and Extracellular Vesicles Correlate with Hepatic Fibrosis Progression in Chronic Hepatitis C. Hepatology 2016, 64, 732–745. [Google Scholar] [CrossRef]
- Rattan, P.; Minacapelli, C.D.; Rustgi, V. The Microbiome and Hepatocellular Carcinoma. Liver Transplant. 2020, 26, 1316–1327. [Google Scholar] [CrossRef]
- Yu, L.-X.; Schwabe, L.-X.Y.R.F. The Gut Microbiome and Liver Cancer: Mechanisms and Clinical Translation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 527–539. [Google Scholar] [CrossRef]
- Hoshida, Y.; Nijman, S.M.; Kobayashi, M.; Chan, J.A.; Brunet, J.-P.; Chiang, D.Y.; Villanueva, A.; Newell, P.; Ikeda, K.; Hashimoto, M.; et al. Integrative Transcriptome Analysis Reveals Common Molecular Subclasses of Human Hepatocellular Carcinoma. Cancer Res. 2009, 69, 7385–7392. [Google Scholar] [CrossRef] [Green Version]
- Guichard, C.; Amaddeo, G.; Imbeaud, S.; Ladeiro, Y.; Pelletier, L.; Ben Maad, I.; Calderaro, J.; Bioulac-Sage, P.; Letexier, M.; Degos, F.; et al. Integrated Analysis of Somatic Mutations and Focal Copy-Number Changes Identifies Key Genes and Pathways in Hepatocellular Carcinoma. Nat. Genet. 2012, 44, 694–698. [Google Scholar] [CrossRef]
- Schulze, K.; Imbeaud, S.; Letouzé, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome Sequencing of Hepatocellular Carcinomas Identifies New Mutational Signatures and Potential Therapeutic Targets. Nat. Genet. 2015, 47, 505–511. [Google Scholar] [CrossRef]
- Sengupta, S.; Parikh, N.D. Biomarker Development for Hepatocellular Carcinoma Early Detection: Current and Future Perspectives. Hepatic Oncol. 2017, 4, 111–122. [Google Scholar] [CrossRef]
- Saad, Y.; El-Serafy, M.; Eldin, M.S.; Abdellatif, Z.; Khatab, H.; Elbaz, T.; Elgarem, H. New Genetic Markers for Diagnosis of Hepatitis C Related Hepatocellular Carcinoma in Egyptian Patients. J. Gastrointest. Liver Dis. 2013, 22, 419–425. [Google Scholar]
- Hodeib, H.; Elshora, O.; Selim, A.; Sabry, N.M.; El-Ashry, H.M. Serum Midkine and Osteopontin Levels as Diagnostic Biomarkers of Hepatocellular Carcinoma. Electron. Physician 2017, 9, 3492–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobhy, A.M.M.F.; Azeem, A.H.; Ashmawy, A.M.; Khalifa, H.O. Significance of Biglycan and Osteopontin as Non-invasive Markers of Liver Fibrosis in Patients with Chronic Hepatitis B Virus and Chronic Hepatitis C Virus. J. Investig. Med. 2018, 67, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zheng, J.; Wu, F.; Kang, B.; Liang, J.; Heskia, F.; Zhang, X.; Shan, Y. OPN is a Promising Serological Biomarker for Hepatocellular Carcinoma Diagnosis. J. Med. Virol. 2020, 92, 3596–3603. [Google Scholar] [CrossRef] [PubMed]
- Kolgelier, S.; Demir, N.A.; Inkaya, A.C.; Sumer, S.; Ozcimen, S.; Demir, L.S.; Pehlivan, F.S.; Arslan, M.; Arpaci, A. Serum Levels of Annexin A2 as a Candidate Biomarker for Hepatic Fibrosis in Patients with Chronic Hepatitis B. Hepat. Mon. 2015, 15, e30655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Liu, J.-X.; Zhang, Z.-J.; Xu, C.-Z.; Zhang, X.-N.; Huang, W.-R.; Zhou, D.-H.; Wang, R.-R.; Chen, X.-D.; Xiao, M.-B.; et al. High Expression of Anxa2 and Stat3 Promote Progression of Hepatocellular Carcinoma and Predict Poor Prognosis. Pathol. Res. Pr. 2019, 215, 152386. [Google Scholar] [CrossRef] [PubMed]
- Ali, L.H.; Higazi, A.M.; Moness, H.M.; Farag, N.M.; Saad, Z.M.; Moukareb, A.H.; Soliman, W.; El Sagheer, G.; El Hamid, S.R.A.; Hamid, H.A. Clinical Significances and Diagnostic Utilities of Both miR-215 and Squamous Cell Carcinoma Antigen-IgM versus Alpha-Fetoprotein in Egyptian Patients with Hepatitis C Virus-Induced Hepatocellular Carcinoma. Clin. Exp. Gastroenterol. 2019, 12, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Gallotta, A.; Paneghetti, L.; Mrázová, V.; Bednárová, A.; Kružlicová, D.; Frecer, V.; Miertus, S.; Biasiolo, A.; Martini, A.; Pontisso, P.; et al. Development of a Novel Diagnostic Algorithm to Predict NASH in HCV-Positive Patients. Int. J. Biol. Markers 2018, 33, 231–236. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Biomarker | Specificity | Viral Etiology | Development | Reference |
|---|---|---|---|---|
| Status | ||||
| AFP | HCC | HBV, HCV | Phase V | [41,46,47,51,52,77,101] |
| DCP | HCC | HBV, HCV | Phase IV | [48,49,50,51,52,53] |
| M2BPGi | HCC | HBV, HCV | Phase IV | [95,96,97,98,99,100,101,102,103,104] |
| MDK | HCC | HCV | Phase III | [81,154] |
| OPN | HCC, fibrosis | HBV, HCV | Phase III | [155,156,157] |
| Annexin A2 | HCC, fibrosis | HBV, HCV | Phase II | [158,159] |
| DKK1 | HCC | HBV, HCV | Phase II | [81,82,85,86] |
| GPC3 | HCC | HCV | Phase II | [128,129] |
| HSP-70 | HCC | HBV, HCV | Phase II | [57] |
| PCSK9 | HCC | HCV | Phase II | [54,55] |
| SCCA | HCC, fibrosis | HCV | Phase II | [160,161] |
| TIMP-1 | Fibrosis | HBV, HCV | Phase II | [119,126] |
| Angiopoietin-2 | HCC, fibrosis | HCV | Phase I | [119,120,121] |
| CCL20 | HCC | HCV | Phase I | [122,123] |
| Endoglin | Fibrosis | HCV | Phase I | [112] |
| VEGF | HCC, fibrosis | HCV | Phase I | [119,123,124,125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Virzì, A.; Gonzalez-Motos, V.; Tripon, S.; Baumert, T.F.; Lupberger, J. Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development. J. Clin. Med. 2021, 10, 977. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050977
Virzì A, Gonzalez-Motos V, Tripon S, Baumert TF, Lupberger J. Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development. Journal of Clinical Medicine. 2021; 10(5):977. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050977
Chicago/Turabian StyleVirzì, Alessia, Victor Gonzalez-Motos, Simona Tripon, Thomas F. Baumert, and Joachim Lupberger. 2021. "Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development" Journal of Clinical Medicine 10, no. 5: 977. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050977