
CD8+ T Cell Responses during HCV Infection and HCC


1
Department of Medicine II, University Hospital Freiburg—Faculty of Medicine, University of Freiburg, Hugstetter Straße 55, 79106 Freiburg, Germany
2
Faculty of Biology, University of Freiburg, Schänzlestraße 1, 79104 Freiburg, Germany
*
Authors to whom correspondence should be addressed.
J. Clin. Med. 2021, 10(5), 991; https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050991
Submission received: 22 December 2020
/
Revised: 12 February 2021
/
Accepted: 22 February 2021
/
Published: 2 March 2021
(This article belongs to the Special Issue Viral Hepatitis and Risk of Hepatocellular Carcinoma)
Abstract
:Chronic hepatitis C virus (cHCV) infection is a major global health burden and the leading cause of hepatocellular carcinoma (HCC) in the Western world. The course and outcome of HCV infection is centrally influenced by CD8+ T cell responses. Indeed, strong virus-specific CD8+ T cell responses are associated with spontaneous viral clearance while failure of these responses, e.g., caused by viral escape and T cell exhaustion, is associated with the development of chronic infection. Recently, heterogeneity within the exhausted HCV-specific CD8+ T cells has been observed with implications for immunotherapeutic approaches also for other diseases. In HCC, the presence of tumor-infiltrating and peripheral CD8+ T cell responses correlates with a favorable prognosis. Thus, tumor-associated and tumor-specific CD8+ T cells are considered suitable targets for immunotherapeutic strategies. Here, we review the current knowledge of CD8+ T cell responses in chronic HCV infection and HCC and their respective failure with the potential consequences for T cell-associated immunotherapeutic approaches.
{kind=link}
{kind=link}
{kind=link}
1. Chronic Hepatitis C Virus Infection and Hepatocellular Carcinoma: Major Global Health Burdens Associated with the Liver
Both, chronic hepatitis C virus (cHCV) infection and hepatocellular carcinoma (HCC) affect the liver and represent major global health burdens. Worldwide, more than 71 million people are currently infected with hepatitis C virus (HCV) [1] with estimated 1.75 million new chronically infected patients per year [2]. In more than 70% of cases, acutely infected patients develop a chronic HCV infection. Acute and chronic HCV infection are mostly asymptomatic, however, chronic hepatitis is highly associated with the development of liver fibrosis which can progress to premalignant cirrhotic remodeling of the liver and ultimately to hepatocellular carcinoma [3]. HCC is the most common form of primary liver cancer in adults and is one of the main causes of cancer-related deaths worldwide [4,5,6,7]. By 2040, a further 65% increase in incidence is expected by the WHO [8].
The liver, although no lymphoid organ, has a rich and highly specified immune composition. The liver immune system is normally in a hypoimmune state, guaranteeing balance between tolerance towards harmless molecules and immunity towards pathogens. This state renders the liver susceptible towards infections and cancer [9]. Nevertheless, upon viral infection for example with HCV, the innate immune system is induced with a rapid activation of the interferon response, natural killer cells and a local increase in cytokines and chemokines [10,11]. This is subsequently followed by a delayed infiltration of CD4+ and CD8+ T cells [12] leading to necro-inflammation. Chronic liver disease associated with chronic necro-inflammation may induce an immunosuppressive, pro-tumorigenic environment [6,13,14] and therefore favors a multifactorial process in which HCC can develop. The tumor microenvironment in HCC consists of various immunosuppressive immune cell populations (e.g., regulatory T cells and myeloid-derived suppressor cells) and immunosuppressive cytokines (e.g., IL-10) [15]. An immunosuppressive tumor microenvironment modulates T cell reactivity [15] and can lead to evasion of HCC from immunosurveillance [16]. Besides chronic viral hepatitis, e.g., induced by cHCV infection, chronic alcohol abuse and non-alcoholic steatohepatosis (NASH), e.g., associated with the metabolic syndrome, frequently drive HCC development also through necro-inflammation. Yet, cHCV infection is still the leading cause of HCC in the Western world [13].
The therapeutic options of HCC are limited and curative therapies such as resection and local ablation are only available for patients with small tumor nodes and well-preserved liver function. Treatment options for patients in advanced stages are mostly restricted to transarterial chemoembolization (TACE), systemic therapy with different agents, or best supportive care due to tumor burden and poor liver function [6]. Thus, risk reduction of HCC development is an important measure in patient care. With respect to chronic viral hepatitis, this is reached by anti-viral treatment. In cHCV infection, the introduction of direct-acting antiviral (DAA) drug therapy in 2014 led to a sustained virological response rate far exceeding 90% of treated patients [17] and thus reduced the risk of HCC development with the exception of patients with undefined/non-malignant hepatic nodules [18,19,20]. Of note, this unique success story of hepatitis C research leading to the development of the highly effective DAA treatment has recently been honored with the Nobel Prize [21]. However, HCV is still far from being eradicated since high costs, limited availability of DAAs, and infrastructural restrictions problems hamper a world-wide campaign [22,23]. Additionally, the incidences of DAA-resistant cases and patient groups with poor prospects for recovery [1] have to be considered. The recent setbacks in vaccine development [24,25,26,27,28] and a lack of immunological protection from reinfection after cure [17,29,30,31] demonstrate that the success of DAA therapy will most probably not be sufficient to reach HCV eradication.
In both, HCV infection and HCC, CD8+ T cells constitute major immune effector cells that elicit cytotoxic and non-cytolytic anti-viral and -tumoral effector functions. However, in the context of chronic antigen stimulation, CD8+ T cells are often functionally impaired resulting in chronic progression of both liver-associated diseases. Therapeutic approaches that induce CD8+ T cells to release their full or at least an improved effector capacity are therefore considered promising in combating the global health burdens by cHCV infection and HCC. This strategy is nicely reflected by the recent attempts of treating HCC by the PD-1 checkpoint inhibitor nivolumab, that among others target CD8+ T cells, led to tumor reduction and to a sustained objective response in 15–20% of patients with advanced HCC [32]. A similar response was achieved with the PD-1 checkpoint inhibitor pembrolizumab [33] that also demonstrated a survival benefit after checkpoint blockade therapy in HCC. Moreover, the combination of atezolizumab (anti-PD-1) and bevacizumab (anti-VEGF) in patients with unresectable HCC (including but not stratifying viral and non-viral etiologies) showed a progression-free survival rate of over 15 months that is superior compared to multikinase inhibitor sorafenib [34] and is thus now considered first line therapy for HCC. In this review, we summarize the current knowledge of CD8+ T cell responses in cHCV infection and HCC with a special focus on their respective failure and the open questions since this sets the basis for the design of new or additive therapeutic strategies aiming at improving anti-viral and anti-tumoral CD8+ T cell responses.
2. CD8+ T Cell Responses in HCV
A robust and effective CD8+ T cell response in association with a strong support by CD4+ T helper cells is necessary for the spontaneous clearance of acute HCV infection [35,36]. Although the occurrence of virus-specific CD8+ T cells in the peripheral blood and liver is delayed (6–8 weeks after infection), it is clearly associated with significant reduction of viral load but also with the onset of liver disease [37,38]. Furthermore, CD8+ T cell depletion studies conducted in chimpanzees resulted in prolonged viremia, further highlighting the central antiviral role of CD8+ T cells [36]. The central role of CD8+ T cell responses is also supported by the observed protective effects of diverse human leukocyte antigen class I alleles, such as HLA-B*27 or –A*03 [39]. Virus-specific CD8+ T cells that are associated with HCV clearance in humans are characterized by expression of activation-associated molecules (PD-1, CD39) [38,40], high levels of T-bet [41], and a low cytokine production [37]. After successful viral elimination and subsequent cessation of antigen triggering, the phenotype of HCV-specific CD8+ T cells shifts toward a classical memory-associated phenotype (high expression of CD127), inheriting a reduced frequency and the ability to readily expand upon reinfection. This memory of CD8+ T cells contributes to an increased likelihood of viral resolution in re-infected patients [42]. In chronic HCV infection, however, the phenotype and functionality of virus-specific CD8+ T cells are tremendously altered; the frequencies of virus-specific CD8+ T cells are lower and the remaining virus-specific CD8+ T cells fail to clear the virus but still most likely contribute to ongoing liver disease [43,44,45].
Failure of Virus-Specific CD8+ T Cells in HCV
Two main mechanisms are associated with the failure of virus-specific CD8+ T cells in cHCV infection. One mechanism is the evasion of the virus from the immune system, a phenomenon called viral escape. Viral escape mutations can lead to a loss of HCV-specific CD8+ T cell epitopes and loss of de novo T cell priming, contributing to reduced recognition and thus altered anti-viral activity of virus-specific CD8+ T cells [40,46,47,48].
The other mechanism is T cell exhaustion, a phenomenon first described in chronic lymphocytic choriomeningitis virus (LCMV) infection in mice [49,50] which can be observed in many chronic viral infections and cancer also in humans. Exhausted virus-specific CD8+ T cells have an impaired survival, an altered metabolic, epigenetic, and molecular signature [48,51,52,53,54] and are characterized by a gradual loss of effector functions and proliferative capacity. In particular, exhausted CD8+ T cells exhibit an increased expression of co-inhibitory molecules such as PD-1, TIM-3, LAG3, 2B4 [55,56,57], high expression of the transcription factors Eomes [58,59] or the recently identified HMG-box transcription factor TOX [60,61,62,63]. The mechanisms leading to T cell exhaustion are manifold, with high viral load and large number of antigen presenting cells [64,65,66], as well as a loss of CD4+ T cell-mediated help and absence of IL-21 signaling playing important roles [50,67,68,69]. Noteworthy, recent studies by the groups of Georg Lauer and Carlo Ferrari showed that transcriptional and metabolic differences in HCV-specific CD8+ T cells are already present at early time points of infection and influence the outcome of infection [48,51].
Exhausted virus-specific CD8+ T cells are heterogeneous and consist of subpopulations with different functional and phenotypic properties [56,59,60,61,63,70,71,72,73,74]. Based on the expression of the inhibitory receptor PD-1 and the IL-7 receptor α-chain CD127, Wieland et al. could identify CD127+ PD-1+ memory-like subsets expressing the transcription factor TCF-1 and CD127- PD-1high Eomeshigh terminally differentiated subsets of exhausted HCV-specific CD8+ T cells in cHCV infection. CD127+ PD-1+ memory-like subsets determine the proliferative capacity of the HCV-specific CD8+ T cells population in cHCV infection that can be further appointed to the TCF-1 expression of this subset. Furthermore, only the memory-like subset is maintained independently from antigen recognition either after viral escape or after antigen withdrawal by DAA therapy [75]. These findings have two important implications: first, ongoing antigen recognition drives differentiation towards terminal exhaustion; and second, the memory-like subset maintains the virus-specific CD8+ T cell pool during and after cHCV infection. In addition, it has been shown that different exhausted virus-specific CD8+ T cell subpopulations in mice and humans are differently responsive towards checkpoint blockade with memory-like CD8+ T cells representing the best responders [55,76].
Noteworthy, the memory-like HCV-specific CD8+ T cell subset retains characteristics of exhausted T cells even after viral elimination by DAA therapy, like a molecular scar of chronicity, and remains functionally inferior compared to conventional memory HCV-specific CD8+ T cells emerging after self-limiting HCV infection [75,77]. Consequently, antigen withdrawal after long-term persisting antigen recognition does not lead to full recovery of exhausted HCV-specific CD8+ T cells and thus to an impaired CD8+ T cell memory [77]. These findings have important translational consequences as they at least partially explain the limited protective capacity of the HCV-specific CD8+ T cells after DAA cure and reinfection [75]. Thus, these results clearly implicate the need to therapeutically target molecular determinants associated with T cell exhaustion to unleash a fully functional and robust CD8+ T cell response after DAA-mediated HCV clearance. Further studies are therefore required to fully explore the imprinted molecular and epigenetic signatures in exhausted HCV-specific CD8+ T cells including the definition of master regulators associated with the differentiation program of T cell exhaustion. A first step in this direction was the recent identification of the HMG-box transcription factor TOX that regulates the epigenetic and transcriptional program in exhausted T cells in mouse models of chronic LCMV infection and cancer [60,61,62] and that is also associated with the exhausted phenotype of HCV-specific CD8+ T cells [60] and PD1+ T cells in HCC [78]. Moreover, this observation of an association of TOX with exhausted T cell characteristics in cHCV and HCC highlights shared principles of CD8+ T cell dysfunction in both liver-associated diseases.
3. CD8+ T Cell Responses in HCC
HCC patients with detectable antigen-specific CD8+ T cell responses during the natural course or induced by anti-tumoral therapy have an improved overall survival. Indeed, lymphocyte infiltrates, in particular tumor-infiltrating CD8+ T cells, have been associated with improved survival and lower relapse rates after liver resection [5,79,80,81,82,83]. However, HCC tumors are frequently only poorly infiltrated by CD8+ T cells or other immune cells [5].
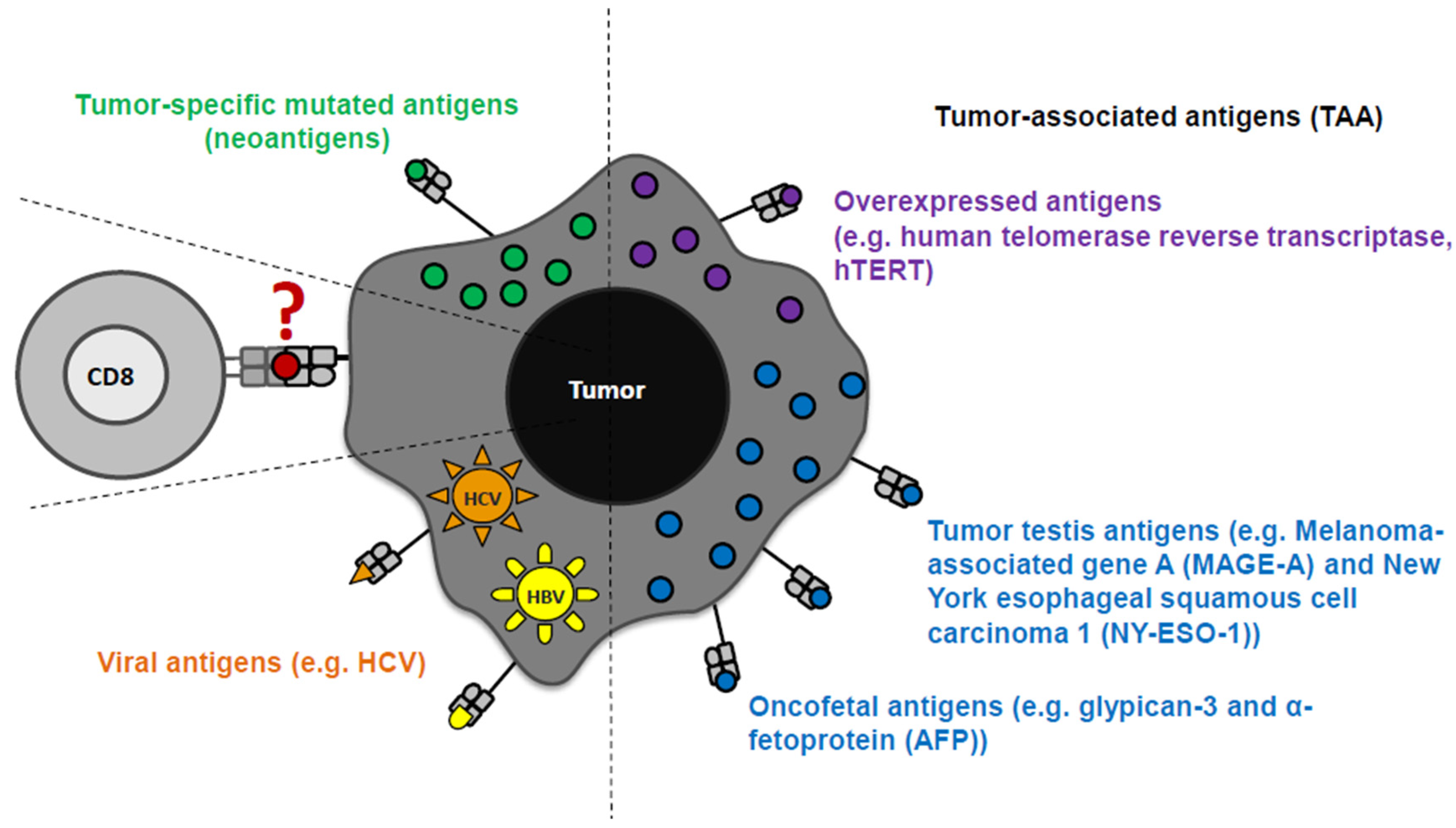
HCC is characterized by a high molecular complexity and genetic heterogeneity, leading to the identification of different tumor antigens in HCC tissue (as well as in other cancers) [84,85,86,87,88,89,90,91,92]. Tumor antigens were initially identified in other tumor types and subsequently examined for their expression and immunogenicity in HCC [93]. Tumor antigens are classified in tumor associated antigens (TAA) and tumor-specific mutant antigens (neoantigen). Neoantigens are rare and only a few have been so far identified in the context of HCC [94,95,96]. In contrast, TAA were detectable in the HLA ligandomes of HCC patients [94,95,96]. With respect to TAA, several categories are distinguished based on the expression pattern, namely tumor testis antigens, overexpressed antigens, differentiation antigens, oncofetal antigens. Another category of antigens in the context of HCC are viral antigens [16,92] (Figure 1). Although TAA may be partially subject to self-tolerance, they are considered as good targets for immunotherapeutic treatment concepts, because they are mainly expressed in tumor cells and shared between patient groups. Of note, differential TAA expression in HCC with different underlying etiologies has not been extensively addressed so far.
TAA-specific CD8+ T cells, which are able to recognize respective tumor antigens in autologous tumor tissue, have been shown to be associated with tumor control. For example, the study of Flecken et al. showed that CD8+ T cell responses specific for TAA are associated with prolonged progression-free survival in HCC patients [84]. The expression of HLA class I molecules in primary HCC [97] and several groups of TAAs that enable CD8+ T cells to recognize tumor cells have been identified [15,84,92]. However, the expression rates of TAA are different. For example, MAGE-A is expressed in up to 80% of HCC patients, while NY-ESO-1 is expressed in less than 50% of HCC patients [98,99,100]. Indeed, several studies showed a significant heterogeneity and no consistent hierarchy between different TAA-specific CD8+ T cell responses within individual cohorts of HCC patients [84,85,98]. Taken together, the relevant immunodominant epitopes targeted by CD8+ T cells are so far not well understood in HCC contrary to well defined viral CD8+ T cell epitopes in cHCV infection [86,101].
Failure of CD8+ T Cells in HCC
Although TAA-specific CD8+ T cells are associated with a better outcome [84], there is evidence of a dysfunctional state of these cells [99]. The underlying molecular mechanisms are, however, largely unknown due to the low frequency of TAA-specific CD8+ T cells in HCC patients. Thus, most studies are based on in vitro expansion protocols with cytokines such as IL-2 or IL-12 [16] prior to ELISPOT assays, limiting the analysis of the ex vivo molecular properties of TAA-specific CD8+ T cells in HCC. By applying peptide/MHCI tetramer-based enrichment, we characterized circulating TAA-specific CD8+ T cells targeting gylpican-3, NY-ESO-1, MAGE-A1, and MAGE-A3 in therapy-naïve HCC patients [102]. The frequencies of TAA-specific CD8+ T cells were comparable in HCC patients and in healthy donors (HD) or in patients with liver cirrhosis but lower compared to virus-specific CD8+ T cells present in HCC patients. Significantly more TAA-specific CD8+ T cells from HCC patients (expressing the respective TAA) displayed an antigen-experienced phenotype (% antigen-experienced MAGE-A-specific CD8+ T cells in HCC: Median: 52.9%; IQR: 60.8%) [102]. This observation indicates an at least partially inefficient TAA-specific CD8+ T cell priming and activation, which may lead to limited expansion and thus to low frequencies that are barely distinguishable from the naïve precursor frequencies. Virus-specific CD8+ T cells in the very same patients showed an antigen-experienced phenotype, which contrasts with an actively persisting general cancer-associated mechanism of improper T cell priming in HCC.
In general, as discussed above, persistent antigen recognition leads to a gradual exhaustion of CD8+ T cells in chronic viral infections and cancer [103,104]. Surprisingly, and in contrast to HCV-specific CD8+ T cells in cHCV patients, antigen-experienced MAGE-A-specific CD8+ T cells do not show a terminally exhausted phenotype (Eomeshi, PD-1hi, TCF-1lo, CD127lo) in therapy-naïve HCC patients [102], potentially reflecting different quantities and qualities of antigen recognition. In addition, only moderate expression of multiple inhibitory receptors—also characteristic marker molecules of T cell exhaustion—has been shown on TAA-specific CD8+ T cells targeting several TAAs in different cohorts of HCC patients with different underlying etiologies [98,101,105,106]. A higher expression of these inhibitory receptors was observed on TAA-specific CD8+ T cells isolated from HCC tissue compared to T cells from tumor-free liver tissue or blood, especially a significantly higher PD-1 expression was detectable on tumor-infiltrating lymphocytes compared to PBMC [98,101,105,106]. Blocking antibodies targeting these inhibitory receptors (PD-1, TIM3, and LAG3) restored T cell function and combinations of antibodies had additive effects [101]. This observation suggests that the even moderate expression of inhibitory receptors on TAA-specific CD8+ T cells dampens the T cell response in HCC. However, further investigations are required to clarify the molecular signatures and with this the exhausted state of TAA-specific CD8+ T cells also in relation to the underlying etiology. Interestingly, CD8+ T cell subsets with molecular signatures of T cell exhaustion (including PD-1 expression) were identified by single cell RNA sequencing of bulk T cells isolated from peripheral blood and HCCs [7]. Although the targeted antigens (including TAAs, neoantigens, and viral antigens) of these exhausted CD8+ T cells remain elusive this finding provides a mechanistic explanation of the durable objective response to PD-1 checkpoint blockade therapy of at least some patients with advanced HCC [32,107]. Further studies are now required to define the tumor antigens targeted by these exhausted CD8+ T cells and subsequently to more precisely analyze their specific molecular profiles in order to optimize checkpoint therapies and other immunotherapeutic approaches like vaccination strategies. Of note, CD8+ T cell responses are markedly affected by the tumor microenvironment (TME). This notion is best exemplified by (i) a study of Di Blasi et al. showing that the presence of certain TIL clusters (e.g., ICOS+ TIGIT+ CD4+ TILs) can serve as a prognostic indicator for the response to checkpoint blockade therapy [108]; and by (ii) the improved progression-free survival of HCC patients treated with Atezolizumab (blocking PD-1 checkpoint pathway) in combination with the VEGF-blocking antibody Bevacizumab (blocking VEGF) that has recently been approved as first line therapy in HCC [34]. Thus, a deep understanding of the overall immune contexture including tumor-resident and tumor-specific immune cells is crucial to answer the following important questions also in relation to the design of new or improved immunotherapeutic approaches: Which immune cells primarily respond to immunotherapy?; Which of the responding immune cells are beneficial, which are deleterious in the anti-tumor response? Or which immune cells support the anti-tumor CD8+ T cell response? Why are only some tumors accessible for immunotherapy?; and which T cell subsets mediate anti-tumoral activity in patients who respond to checkpoint blockade therapy?
4. Concluding Remarks
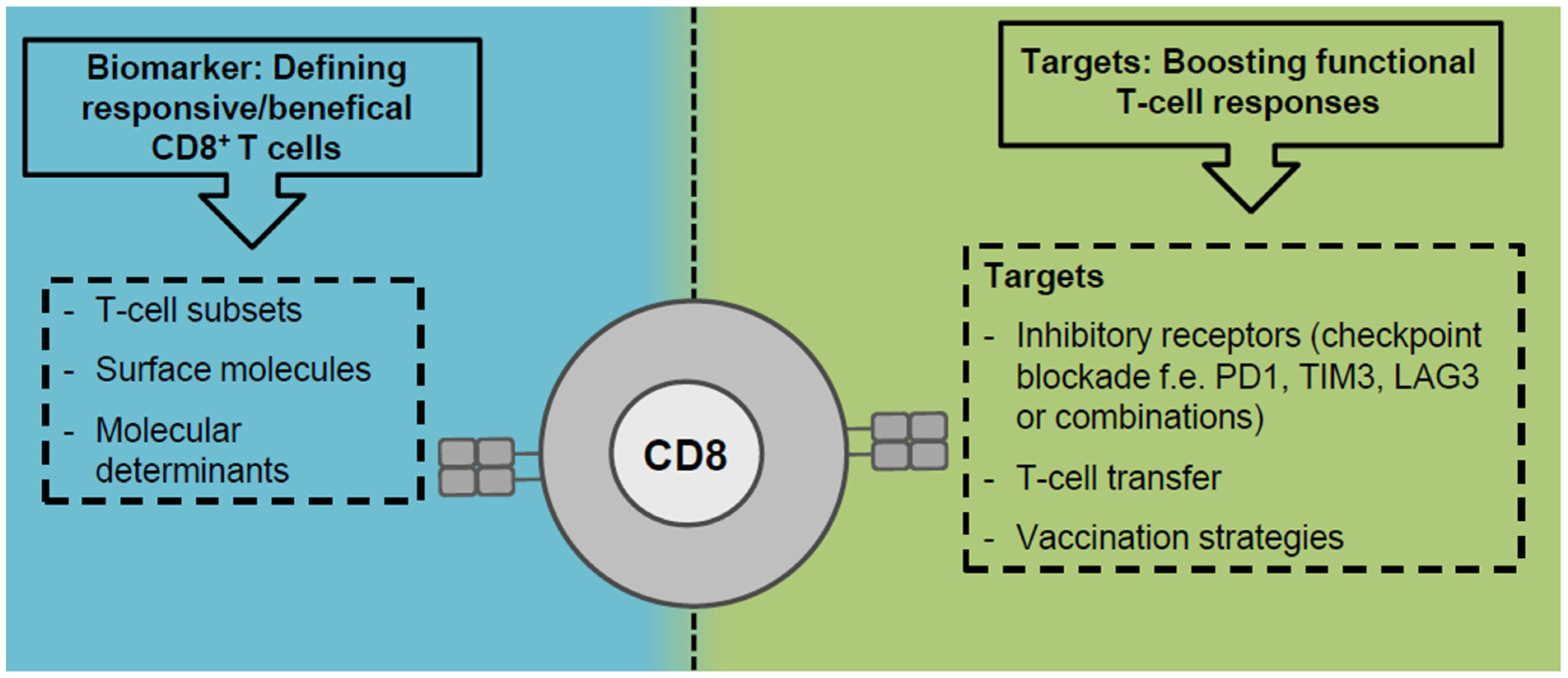
Clearly, CD8+ T cells are major effector cells in anti-viral and anti-tumoral immunity in cHCV infection and HCC and CD8+ T cell impairment is common in both liver-associated diseases. The approval of DAA therapy not only revolutionized patient care but also provides a unique chance to study CD8+ T cell impairment by T cell exhaustion in a clinically relevant setting. By taking this opportunity, recent studies [51,60,75,109,110] underlined the relevant role of ongoing antigen recognition in driving HCV-specific CD8+ T cell exhaustion and offered novel insights into phenotypic and functional heterogeneity, metabolic dysregulation and fate of exhausted CD8+ T cells during and after cHCV infection. Although there is some data available regarding CD8+ T cell exhaustion in HCC [7,101] much less is understood concerning mechanistic details due to the absence of knowledge about the targeted antigens. However, general principles of CD8+ T cell exhaustion/dysfunction seem to be conserved between virus-specific and tumor-specific CD8+ T cells [103] which is best reflected by the common master regulator TOX [60,61,62,78]. Definition of central factors driving T cell exhaustion/dysfunction that are targetable by immunotherapeutic approaches may therefore be translatable from chronic viral infections to cancer and vice versa. However, there are also considerable differences in chronic viral infections and cancer that potentially impact the CD8+ T cell response: e.g., origin of antigen (exogenous versus endogenous) associated with antigen quantity and presentation, the differential expression of cytokines and other immune mediators (pro-inflammatory versus immunosuppressive), and the composition of the other immune cells (CD4+ T cell help, CD4+ regulatory T cells, myeloid-derived suppressor cells, and tumor-associated macrophages). Thus, future studies are required to define shared and diverging determinants and molecular characteristics of CD8+ T cell dysfunction in chronic viral infection and cancer. For this, cHCV infection and HCC represent important translational settings since both affect the liver and may also occur in combination helping to dissect virus- and tumor-associated effects on CD8+ T cell dysfunction. This knowledge will provide rationales for establishing predictive biomarkers, e.g., responding/beneficial immune cells and CD8+ T cell subsets, and the design of novel or improved immunotherapeutic approaches, like combinatorial treatments lowering inhibitory signals from the microenvironment and recruiting/boosting the most functional CD8+ T cell response (Figure 2). Both are especially urgently needed for HCC treatment.
Author Contributions
R.T. and M.H. drafted and wrote the manuscript. C.T. and N.H. contributed to writing this manuscript and compiled the figures. All authors have read and agreed to the published version of the manuscript.
Funding
R.T. received funding by the CRC/TRR 179-Project 01 and M.H. by the CRC/TRR 179-Project 20 of the German Research Foundation (DFG; TRR 179 project number: 272983813). M.H. was supported by a Margarete von Wrangell fellowship (State of Baden-Wuerttemberg).
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
This review article is based on previously published work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bartenschlager, R.; Baumert, T.F.; Bukh, J.; Houghton, M.; Lemon, S.M.; Lindenbach, B.D.; Lohmann, V.; Moradpour, D.; Pietschmann, T.; Rice, C.M.; et al. Critical challenges and emerging opportunities in hepatitis c virus research in an era of potent antiviral therapy: Considerations for scientists and funding agencies. Virus Res. 2018, 248, 53–62. [Google Scholar] [CrossRef]
- Cox, A.L. Challenges and promise of a hepatitis c virus vaccine. Cold Spring Harb. Perspect. Med. 2020, 10. [Google Scholar] [CrossRef]
- Spearman, C.W.; Dusheiko, G.M.; Hellard, M.; Sonderup, M. Hepatitis c. Lancet 2019, 394, 1451–1466. [Google Scholar] [CrossRef]
- El-Serag, H.B. Hepatocellular carcinoma. N. Engl. J. Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef]
- Foerster, F.; Hess, M.; Gerhold-Ay, A.; Marquardt, J.U.; Becker, D.; Galle, P.R.; Schuppan, D.; Binder, H.; Bockamp, E. The immune contexture of hepatocellular carcinoma predicts clinical outcome. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- EASL. Easl clinical practice guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Zheng, L.; Yoo, J.K.; Guo, H.; Zhang, Y.; Guo, X.; Kang, B.; Hu, R.; Huang, J.Y.; Zhang, Q.; et al. Landscape of infiltrating t cells in liver cancer revealed by single-cell sequencing. Cell 2017, 169, 1342–1356. [Google Scholar] [CrossRef] [Green Version]
- Rawla, P.; Sunkara, T.; Muralidharan, P.; Raj, J.P. Update in global trends and aetiology of hepatocellular carcinoma. Contemp. Oncol. 2018, 22, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Kubes, P.; Jenne, C. Immune responses in the liver. Annu. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef]
- Dustin, L.B.; Rice, C.M. Flying under the radar: The immunobiology of hepatitis c. Annu. Rev. Immunol 2007, 25, 71–99. [Google Scholar] [CrossRef]
- Cheent, K.; Khakoo, S.I. Natural killer cells and hepatitis c: Action and reaction. Gut 2011, 60, 268–278. [Google Scholar] [CrossRef]
- Protzer, U.; Maini, M.K.; Knolle, P.A. Living in the liver: Hepatic infections. Nat. Rev. Immunol. 2012, 12, 201–213. [Google Scholar] [CrossRef]
- Ringelhan, M.; Pfister, D.; O’Connor, T.; Pikarsky, E.; Heikenwalder, M. The immunology of hepatocellular carcinoma. Nat. Immunol 2018, 19, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, K.; Hato, T.; Chen, Y.; Duda, D.G. Anti-vegfr therapy as a partner for immune-based therapy approaches in hcc. In Immunotherapy of Hepatocellular Carcinoma; Greten, T.F., Ed.; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Breous, E.; Thimme, R. Potential of immunotherapy for hepatocellular carcinoma. J. Hepatol. 2011, 54, 830–834. [Google Scholar] [CrossRef] [Green Version]
- Mizukoshi, E.; Kaneko, S. Antigen-specific t cell responses in hepatocellular carcinoma. In Immunotherapy of Hepatocellular Carcinoma; Greten, T.F., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 39–50. [Google Scholar]
- Falade-Nwulia, O.; Suarez-Cuervo, C.; Nelson, D.R.; Fried, M.W.; Segal, J.B.; Sulkowski, M.S. Oral direct-acting agent therapy for hepatitis c virus infection: A systematic review. Ann. Intern. Med. 2017, 166, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Calvaruso, V.; Cabibbo, G.; Cacciola, I.; Petta, S.; Madonia, S.; Bellia, A.; Tine, F.; Distefano, M.; Licata, A.; Giannitrapani, L.; et al. Incidence of hepatocellular carcinoma in patients with hcv-associated cirrhosis treated with direct-acting antiviral agents. Gastroenterology 2018, 155, 411–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannou, G.N.; Green, P.K.; Berry, K. Hcv eradication induced by direct-acting antiviral agents reduces the risk of hepatocellular carcinoma. J. Hepatol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sangiovanni, A.; Alimenti, E.; Gattai, R.; Filomia, R.; Parente, E.; Valenti, L.; Marzi, L.; Pellegatta, G.; Borgia, G.; Gambato, M.; et al. Undefined/non-malignant hepatic nodules are associated with early occurrence of hcc in daa-treated patients with hcv-related cirrhosis. J. Hepatol 2020, 73, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Prize, T.N. The Nobel Prize in Physiology or Medicine 2020 Press Release. Available online: https://www.nobelprize.org/prizes/medicine/2020/press-release/ (accessed on 5 November 2020).
- Barber, M.J.; Gotham, D.; Khwairakpam, G.; Hill, A. Price of a hepatitis C cure: Cost of production and current prices for direct-acting antivirals in 50 countries. J. Virus Erad. 2020, 6, 100001. [Google Scholar] [CrossRef]
- Iyengar, S.; Tay-Teo, K.; Vogler, S.; Beyer, P.; Wiktor, S.; de Joncheere, K.; Hill, S. Prices, costs, and affordability of new medicines for hepatitis c in 30 countries: An economic analysis. PLoS Med. 2016, 13, e1002032. [Google Scholar] [CrossRef]
- Cox, A.L.; Page, K.; Melia, M.; Veenhuis, R.; Massaccesi, G.; Osburn, W.; Wagner, K.; Giudice, L.; Stein, E.; Asher, A.K.; et al. Lb10. A randomized, double-blind, placebo-controlled efficacy trial of a vaccine to prevent chronic hepatitis c virus infection in an at-risk population. Open Forum Infect. Dis. 2019, 6, S997. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.E.; Houghton, M.; Coates, S.; Abrignani, S.; Chien, D.; Rosa, D.; Pileri, P.; Ray, R.; Di Bisceglie, A.M.; Rinella, P.; et al. Safety and immunogenicity of hcv e1e2 vaccine adjuvanted with mf59 administered to healthy adults. Vaccine 2010, 28, 6367–6373. [Google Scholar] [CrossRef] [Green Version]
- Law, J.L.; Chen, C.; Wong, J.; Hockman, D.; Santer, D.M.; Frey, S.E.; Belshe, R.B.; Wakita, T.; Bukh, J.; Jones, C.T.; et al. A hepatitis c virus (hcv) vaccine comprising envelope glycoproteins gpe1/gpe2 derived from a single isolate elicits broad cross-genotype neutralizing antibodies in humans. PLoS ONE 2013, 8, e59776. [Google Scholar] [CrossRef]
- Shoukry, N.H. Hepatitis c vaccines, antibodies, and t cells. Front. Immunol. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Swadling, L.; Capone, S.; Antrobus, R.D.; Brown, A.; Richardson, R.; Newell, E.W.; Halliday, J.; Kelly, C.; Bowen, D.; Fergusson, J.; et al. A human vaccine strategy based on chimpanzee adenoviral and mva vectors that primes, boosts, and sustains functional hcv-specific t cell memory. Sci. Transl. Med. 2014, 6, 261ra153. [Google Scholar] [CrossRef] [Green Version]
- Midgard, H.; Weir, A.; Palmateer, N.; Lo Re, V., 3rd; Pineda, J.A.; Macias, J.; Dalgard, O. Hcv epidemiology in high-risk groups and the risk of reinfection. J. Hepatol. 2016, 65, S33–S45. [Google Scholar] [CrossRef] [Green Version]
- Rossi, C.; Butt, Z.A.; Wong, S.; Buxton, J.A.; Islam, N.; Yu, A.; Darvishian, M.; Gilbert, M.; Wong, J.; Chapinal, N.; et al. Hepatitis c virus reinfection after successful treatment with direct-acting antiviral therapy in a large population-based cohort. J. Hepatol. 2018, 69, 1007–1014. [Google Scholar] [CrossRef]
- Simmons, B.; Saleem, J.; Hill, A.; Riley, R.D.; Cooke, G.S. Risk of late relapse or reinfection with hepatitis c virus after achieving a sustained virological response: A systematic review and meta-analysis. Clin. Infect. Dis 2016, 62, 683–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Khoueiry, A.B.; Sangro, B.; Yau, T.; Crocenzi, T.S.; Kudo, M.; Hsu, C.; Kim, T.Y.; Choo, S.P.; Trojan, J.; Welling, T.H.R.; et al. Nivolumab in patients with advanced hepatocellular carcinoma (checkmate 040): An open-label, non-comparative, phase 1/2 dose escalation and expansion trial. Lancet 2017, 389, 2492–2502. [Google Scholar] [CrossRef]
- Zhu, A.X.; Finn, R.S.; Edeline, J.; Cattan, S.; Ogasawara, S.; Palmer, D.; Verslype, C.; Zagonel, V.; Fartoux, L.; Vogel, A.; et al. Pembrolizumab in patients with advanced hepatocellular carcinoma previously treated with sorafenib (keynote-224): A non-randomised, open-label phase 2 trial. Lancet Oncol. 2018, 19, 940–952. [Google Scholar] [CrossRef]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.-Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus bevacizumab in unresectable hepatocellular carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef] [PubMed]
- Grakoui, A.; Shoukry, N.H.; Woollard, D.J.; Han, J.H.; Hanson, H.L.; Ghrayeb, J.; Murthy, K.K.; Rice, C.M.; Walker, C.M. Hcv persistence and immune evasion in the absence of memory t cell help. Science 2003, 302, 659–662. [Google Scholar] [CrossRef] [Green Version]
- Shoukry, N.H.; Grakoui, A.; Houghton, M.; Chien, D.Y.; Ghrayeb, J.; Reimann, K.A.; Walker, C.M. Memory cd8+ t cells are required for protection from persistent hepatitis c virus infection. J. Exp. Med. 2003, 197, 1645–1655. [Google Scholar] [CrossRef]
- Lechner, F.; Wong, D.K.; Dunbar, P.R.; Chapman, R.; Chung, R.T.; Dohrenwend, P.; Robbins, G.; Phillips, R.; Klenerman, P.; Walker, B.D. Analysis of successful immune responses in persons infected with hepatitis c virus. J. Exp. Med. 2000, 191, 1499–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thimme, R.; Oldach, D.; Chang, K.M.; Steiger, C.; Ray, S.C.; Chisari, F.V. Determinants of viral clearance and persistence during acute hepatitis c virus infection. J. Exp. Med. 2001, 194, 1395–1406. [Google Scholar] [CrossRef]
- Kemming, J.; Thimme, R.; Neumann-Haefelin, C. Adaptive immune response against hepatitis c virus. Int. J. Mol. Sci 2020, 21, 5644. [Google Scholar] [CrossRef] [PubMed]
- Kasprowicz, V.; Schulze Zur Wiesch, J.; Kuntzen, T.; Nolan, B.E.; Longworth, S.; Berical, A.; Blum, J.; McMahon, C.; Reyor, L.L.; Elias, N.; et al. High level of pd-1 expression on hepatitis c virus (hcv)-specific cd8+ and cd4+ t cells during acute hcv infection, irrespective of clinical outcome. J. Virol. 2008, 82, 3154–3160. [Google Scholar] [CrossRef] [Green Version]
- Kurktschiev, P.D.; Raziorrouh, B.; Schraut, W.; Backmund, M.; Wachtler, M.; Wendtner, C.M.; Bengsch, B.; Thimme, R.; Denk, G.; Zachoval, R.; et al. Dysfunctional cd8+ t cells in hepatitis b and c are characterized by a lack of antigen-specific t-bet induction. J. Exp. Med. 2014, 211, 2047–2059. [Google Scholar] [CrossRef]
- Osburn, W.O.; Fisher, B.E.; Dowd, K.A.; Urban, G.; Liu, L.; Ray, S.C.; Thomas, D.L.; Cox, A.L. Spontaneous control of primary hepatitis c virus infection and immunity against persistent reinfection. Gastroenterology 2010, 138, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Klenerman, P.; Thimme, R. T cell responses in hepatitis c: The good, the bad and the unconventional. Gut 2012, 61, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Shirai, M.; Arichi, T.; Nishioka, M.; Nomura, T.; Ikeda, K.; Kawanishi, K.; Engelhard, V.H.; Feinstone, S.M.; Berzofsky, J.A. Ctl responses of hla-a2.1-transgenic mice specific for hepatitis c viral peptides predict epitopes for ctl of humans carrying hla-a2.1. J. Immunol. 1995, 154, 2733–2742. [Google Scholar] [PubMed]
- Lechner, F.; Cuero, A.L.; Kantzanou, M.; Klenerman, P. Studies of human antiviral cd8+ lymphocytes using class i peptide tetramers. Rev. Med. Virol. 2001, 11, 11–22. [Google Scholar] [CrossRef]
- Cox, A.L.; Mosbruger, T.; Mao, Q.; Liu, Z.; Wang, X.H.; Yang, H.C.; Sidney, J.; Sette, A.; Pardoll, D.; Thomas, D.L.; et al. Cellular immune selection with hepatitis c virus persistence in humans. J. Exp. Med. 2005, 201, 1741–1752. [Google Scholar] [CrossRef]
- Neumann-Haefelin, C.; Frick, D.N.; Wang, J.J.; Pybus, O.G.; Salloum, S.; Narula, G.S.; Eckart, A.; Biezynski, A.; Eiermann, T.; Klenerman, P.; et al. Analysis of the evolutionary forces in an immunodominant cd8 epitope in hepatitis c virus at a population level. J. Virol. 2008, 82, 3438–3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolski, D.; Foote, P.K.; Chen, D.Y.; Lewis-Ximenez, L.L.; Fauvelle, C.; Aneja, J.; Walker, A.; Tonnerre, P.; Torres-Cornejo, A.; Kvistad, D.; et al. Early transcriptional divergence marks virus-specific primary human cd8(+) t cells in chronic versus acute infection. Immunity 2017, 47, 648–663.e8. [Google Scholar] [CrossRef] [Green Version]
- Moskophidis, D.; Lechner, F.; Pircher, H.; Zinkernagel, R.M. Virus persistence in acutely infected immunocompetent mice by exhaustion of antiviral cytotoxic effector t cells. Nature 1993, 362, 758–761. [Google Scholar] [CrossRef]
- Zajac, A.J.; Blattman, J.N.; Murali-Krishna, K.; Sourdive, D.J.; Suresh, M.; Altman, J.D.; Ahmed, R. Viral immune evasion due to persistence of activated t cells without effector function. J. Exp. Med. 1998, 188, 2205–2213. [Google Scholar] [CrossRef]
- Barili, V.; Fisicaro, P.; Montanini, B.; Acerbi, G.; Filippi, A.; Forleo, G.; Romualdi, C.; Ferracin, M.; Guerrieri, F.; Pedrazzi, G.; et al. Targeting p53 and histone methyltransferases restores exhausted cd8+ t cells in hcv infection. Nat. Commun. 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patsoukis, N.; Bardhan, K.; Chatterjee, P.; Sari, D.; Liu, B.; Bell, L.N.; Karoly, E.D.; Freeman, G.J.; Petkova, V.; Seth, P.; et al. Pd-1 alters t-cell metabolic reprogramming by inhibiting glycolysis and promoting lipolysis and fatty acid oxidation. Nat. Commun. 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pauken, K.E.; Sammons, M.A.; Odorizzi, P.M.; Manne, S.; Godec, J.; Khan, O.; Drake, A.M.; Chen, Z.; Sen, D.R.; Kurachi, M.; et al. Epigenetic stability of exhausted t cells limits durability of reinvigoration by pd-1 blockade. Science 2016, 354, 1160–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, D.R.; Kaminski, J.; Barnitz, R.A.; Kurachi, M.; Gerdemann, U.; Yates, K.B.; Tsao, H.W.; Godec, J.; LaFleur, M.W.; Brown, F.D.; et al. The epigenetic landscape of t cell exhaustion. Science 2016, 354, 1165–1169. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, S.D.; Shin, H.; Freeman, G.J.; Wherry, E.J. Selective expansion of a subset of exhausted cd8 t cells by alphapd-l1 blockade. Proc. Natl. Acad. Sci. USA 2008, 105, 15016–15021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doering, T.A.; Crawford, A.; Angelosanto, J.M.; Paley, M.A.; Ziegler, C.G.; Wherry, E.J. Network analysis reveals centrally connected genes and pathways involved in cd8+ t cell exhaustion versus memory. Immunity 2012, 37, 1130–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youngblood, B.; Oestreich, K.J.; Ha, S.J.; Duraiswamy, J.; Akondy, R.S.; West, E.E.; Wei, Z.; Lu, P.; Austin, J.W.; Riley, J.L.; et al. Chronic virus infection enforces demethylation of the locus that encodes pd-1 in antigen-specific cd8(+) t cells. Immunity 2011, 35, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Buggert, M.; Tauriainen, J.; Yamamoto, T.; Frederiksen, J.; Ivarsson, M.A.; Michaelsson, J.; Lund, O.; Hejdeman, B.; Jansson, M.; Sonnerborg, A.; et al. T-bet and eomes are differentially linked to the exhausted phenotype of cd8+ t cells in hiv infection. PLoS Pathog. 2014, 10, e1004251. [Google Scholar] [CrossRef] [Green Version]
- Paley, M.A.; Kroy, D.C.; Odorizzi, P.M.; Johnnidis, J.B.; Dolfi, D.V.; Barnett, B.E.; Bikoff, E.K.; Robertson, E.J.; Lauer, G.M.; Reiner, S.L.; et al. Progenitor and terminal subsets of cd8+ t cells cooperate to contain chronic viral infection. Science 2012, 338, 1220–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfei, F.; Kanev, K.; Hofmann, M.; Wu, M.; Ghoneim, H.E.; Roelli, P.; Utzschneider, D.T.; von Hoesslin, M.; Cullen, J.G.; Fan, Y.; et al. Tox reinforces the phenotype and longevity of exhausted t cells in chronic viral infection. Nature 2019, 571, 265–269. [Google Scholar] [CrossRef]
- Khan, O.; Giles, J.R.; McDonald, S.; Manne, S.; Ngiow, S.F.; Patel, K.P.; Werner, M.T.; Huang, A.C.; Alexander, K.A.; Wu, J.E.; et al. Tox transcriptionally and epigenetically programs cd8(+) t cell exhaustion. Nature 2019, 571, 211–218. [Google Scholar] [CrossRef]
- Scott, A.C.; Dundar, F.; Zumbo, P.; Chandran, S.S.; Klebanoff, C.A.; Shakiba, M.; Trivedi, P.; Menocal, L.; Appleby, H.; Camara, S.; et al. Tox is a critical regulator of tumour-specific t cell differentiation. Nature 2019, 571, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Sun, H.W.; Lacey, N.E.; Ji, Y.; Moseman, E.A.; Shih, H.Y.; Heuston, E.F.; Kirby, M.; Anderson, S.; Cheng, J.; et al. Single-cell rna-seq reveals tox as a key regulator of cd8(+) t cell persistence in chronic infection. Nat. Immunol. 2019, 20, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.N.; Ahmed, R. High antigen levels are the cause of t cell exhaustion during chronic viral infection. Proc. Natl. Acad. Sci. USA 2009, 106, 8623–8628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utzschneider, D.T.; Alfei, F.; Roelli, P.; Barras, D.; Chennupati, V.; Darbre, S.; Delorenzi, M.; Pinschewer, D.D.; Zehn, D. High antigen levels induce an exhausted phenotype in a chronic infection without impairing t cell expansion and survival. J. Exp. Med. 2016, 213, 1819–1834. [Google Scholar] [CrossRef]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into t cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Elsaesser, H.; Sauer, K.; Brooks, D.G. Il-21 is required to control chronic viral infection. Science 2009, 324, 1569–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohlich, A.; Kisielow, J.; Schmitz, I.; Freigang, S.; Shamshiev, A.T.; Weber, J.; Marsland, B.J.; Oxenius, A.; Kopf, M. Il-21r on t cells is critical for sustained functionality and control of chronic viral infection. Science 2009, 324, 1576–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.S.; Du, M.; Zajac, A.J. A vital role for interleukin-21 in the control of a chronic viral infection. Science 2009, 324, 1572–1576. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Hou, S.; Liu, C.; Zhang, A.; Bai, Q.; Han, M.; Yang, Y.; Wei, G.; Shen, T.; Yang, X.; et al. Follicular cxcr5- expressing cd8(+) t cells curtail chronic viral infection. Nature 2016, 537, 412–428. [Google Scholar] [CrossRef]
- Im, S.J.; Hashimoto, M.; Gerner, M.Y.; Lee, J.; Kissick, H.T.; Burger, M.C.; Shan, Q.; Hale, J.S.; Lee, J.; Nasti, T.H.; et al. Defining cd8+ t cells that provide the proliferative burst after pd-1 therapy. Nature 2016, 537, 417–421. [Google Scholar] [CrossRef]
- Leong, Y.A.; Chen, Y.; Ong, H.S.; Wu, D.; Man, K.; Deleage, C.; Minnich, M.; Meckiff, B.J.; Wei, Y.; Hou, Z.; et al. Cxcr5(+) follicular cytotoxic t cells control viral infection in b cell follicles. Nat. Immunol. 2016, 17, 1187–1196. [Google Scholar] [CrossRef]
- Man, K.; Gabriel, S.S.; Liao, Y.; Gloury, R.; Preston, S.; Henstridge, D.C.; Pellegrini, M.; Zehn, D.; Berberich-Siebelt, F.; Febbraio, M.A.; et al. Transcription factor irf4 promotes cd8(+) t cell exhaustion and limits the development of memory-like t cells during chronic infection. Immunity 2017, 47, 1129–1141.e5. [Google Scholar] [CrossRef] [Green Version]
- Utzschneider, D.T.; Charmoy, M.; Chennupati, V.; Pousse, L.; Ferreira, D.P.; Calderon-Copete, S.; Danilo, M.; Alfei, F.; Hofmann, M.; Wieland, D.; et al. T cell factor 1-expressing memory-like cd8(+) t cells sustain the immune response to chronic viral infections. Immunity 2016, 45, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieland, D.; Kemming, J.; Schuch, A.; Emmerich, F.; Knolle, P.; Neumann-Haefelin, C.; Held, W.; Zehn, D.; Hofmann, M.; Thimme, R. Tcf1(+) hepatitis c virus-specific cd8(+) t cells are maintained after cessation of chronic antigen stimulation. Nat. Commun. 2017, 8, 15050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, B.C.; Sen, D.R.; Al Abosy, R.; Bi, K.; Virkud, Y.V.; LaFleur, M.W.; Yates, K.B.; Lako, A.; Felt, K.; Naik, G.S.; et al. Subsets of exhausted cd8(+) t cells differentially mediate tumor control and respond to checkpoint blockade. Nat. Immunol. 2019, 20, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Hensel, N.; Gu, Z.; Sagar; Wieland, D.; Jechow, K.; Kemming, J.; Llewellyn-Lacey, S.; Gostick, E.; Sogukpinar, O.; Emmerich, F.; et al. Memory-like hcv-specific cd8(+) t cells retain a molecular scar after cure of chronic hcv infection. Nat. Immunol. 2021, 22, 229–239. [Google Scholar]
- Wang, X.; He, Q.; Shen, H.; Xia, A.; Tian, W.; Yu, W.; Sun, B. Tox promotes the exhaustion of antitumor cd8(+) t cells by preventing pd1 degradation in hepatocellular carcinoma. J. Hepatol. 2019, 71, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; He, J.C.; Yang, Y.; Wang, J.M.; Qian, Y.W.; Yang, T.; Ji, L. The prognostic value of tumor-infiltrating lymphocytes in hepatocellular carcinoma: A systematic review and meta-analysis. Sci. Rep. 2017, 7, 7525. [Google Scholar] [CrossRef]
- Gabrielson, A.; Wu, Y.; Wang, H.; Jiang, J.; Kallakury, B.; Gatalica, Z.; Reddy, S.; Kleiner, D.; Fishbein, T.; Johnson, L.; et al. Intratumoral cd3 and cd8 t-cell densities associated with relapse-free survival in hcc. Cancer Immunol. Res. 2016, 4, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Qiu, S.-J.; Fan, J.; Zhou, J.; Wang, X.-Y.; Xiao, Y.-S.; Xu, Y.; Li, Y.-W.; Tang, Z.-Y. Intratumoral balance of regulatory and cytotoxic t cells is associated with prognosis of hepatocellular carcinoma after resection. J. Clin. Oncol. 2007, 25, 2586–2593. [Google Scholar] [CrossRef] [Green Version]
- Unitt, E.; Marshall, A.; Gelson, W.; Rushbrook, S.M.; Davies, S.; Vowler, S.L.; Morris, L.S.; Coleman, N.; Alexander, G.J.M. Tumour lymphocytic infiltrate and recurrence of hepatocellular carcinoma following liver transplantation. J. Hepatol. 2006, 45, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Nakashima, O.; Kutami, R.; Yamamoto, O.; Kojiro, M. Clinicopathological study on hepatocellular carcinoma with lymphocytic infiltration. Hepatology 1998, 27, 407–414. [Google Scholar] [CrossRef]
- Flecken, T.; Schmidt, N.; Hild, S.; Gostick, E.; Drognitz, O.; Zeiser, R.; Schemmer, P.; Bruns, H.; Eiermann, T.; Price, D.A.; et al. Immunodominance and functional alterations of tumor-associated antigen-specific cd8+ t-cell responses in hepatocellular carcinoma. Hepatology 2014, 59, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizukoshi, E.; Nakamoto, Y.; Arai, K.; Yamashita, T.; Sakai, A.; Sakai, Y.; Kagaya, T.; Yamashita, T.; Honda, M.; Kaneko, S. Comparative analysis of various tumor-associated antigen-specific t-cell responses in patients with hepatocellular carcinoma. Hepatology 2011, 53, 1206–1216. [Google Scholar] [CrossRef] [PubMed]
- Sideras, K.; Bots, S.J.; Biermann, K.; Sprengers, D.; Polak, W.G.; JN, I.J.; de Man, R.A.; Pan, Q.; Sleijfer, S.; Bruno, M.J.; et al. Tumour antigen expression in hepatocellular carcinoma in a low-endemic western area. Br. J. Cancer 2015, 112, 1911–1920. [Google Scholar] [CrossRef] [Green Version]
- Zerbini, A.; Pilli, M.; Soliani, P.; Ziegler, S.; Pelosi, G.; Orlandini, A.; Cavallo, C.; Uggeri, J.; Scandroglio, R.; Crafa, P.; et al. Ex vivo characterization of tumor-derived melanoma antigen encoding gene-specific cd8+cells in patients with hepatocellular carcinoma. J. Hepatol. 2004, 40, 102–109. [Google Scholar] [CrossRef]
- Butterfield, L.H.; Ribas, A.; Potter, D.M.; Economou, J.S. Spontaneous and vaccine induced afp-specific t cell phenotypes in subjects with afp-positive hepatocellular cancer. Cancer Immunol. Immunother. 2007, 56, 1931–1943. [Google Scholar] [CrossRef] [PubMed]
- Komori, H.; Nakatsura, T.; Senju, S.; Yoshitake, Y.; Motomura, Y.; Ikuta, Y.; Fukuma, D.; Yokomine, K.; Harao, M.; Beppu, T.; et al. Identification of hla-a2- or hla-a24-restricted ctl epitopes possibly useful for glypican-3-specific immunotherapy of hepatocellular carcinoma. Clin. Cancer Res. 2006, 12, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Korangy, F.; Ormandy, L.A.; Bleck, J.S.; Klempnauer, J.; Wilkens, L.; Manns, M.P.; Greten, T.F. Spontaneous tumor-specific humoral and cellular immune responses to ny-eso-1 in hepatocellular carcinoma. Clin. Cancer Res. 2004, 10, 4332–4341. [Google Scholar] [CrossRef] [Green Version]
- Thimme, R.; Neagu, M.; Boettler, T.; Neumann-Haefelin, C.; Kersting, N.; Geissler, M.; Makowiec, F.; Obermaier, R.; Hopt, U.T.; Blum, H.E.; et al. Comprehensive analysis of the alpha-fetoprotein-specific cd8+ t cell responses in patients with hepatocellular carcinoma. Hepatology 2008, 48, 1821–1833. [Google Scholar] [CrossRef]
- Van den Eynde, B.J.; van der Bruggen, P. T cell defined tumor antigens. Curr. Opin. Immunol. 1997, 9, 684–693. [Google Scholar] [CrossRef]
- Schmidt, N.; Büttner, N.; Thimme, R. Perspektiven für eine immuntherapie beim hepatozellulären karzinom. Dtsch Med. Wochenschr. 2013, 138, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Jiang, J.; Zhan, M.; Zhang, H.; Wang, Q.T.; Sun, S.N.; Guo, X.K.; Yin, H.; Wei, Y.; Li, S.Y.; et al. Targeting tumor-associated antigens in hepatocellular carcinoma for immunotherapy: Past pitfalls and future strategies. Hepatology 2021, 73, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Löffler, M.W.; Mohr, C.; Bichmann, L.; Freudenmann, L.K.; Walzer, M.; Schroeder, C.M.; Trautwein, N.; Hilke, F.J.; Zinser, R.S.; Mühlenbruch, L.; et al. Multi-omics discovery of exome-derived neoantigens in hepatocellular carcinoma. Genome Med. 2019, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.Q.; Peng, L.H.; Ma, L.J.; Liu, D.B.; Zhang, S.; Luo, S.Z.; Rao, J.H.; Zhu, H.W.; Yang, S.X.; Xi, S.J.; et al. Heterogeneous immunogenomic features and distinct escape mechanisms in multifocal hepatocellular carcinoma. J. Hepatol. 2020, 72, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cai, M.-Y.; Wei, D.-P. Hla class i expression in primary hepatocellular carcinoma. World J. Gastroenterol. 2002, 8, 654–657. [Google Scholar] [CrossRef]
- Gehring, A.J.; Ho, Z.Z.; Tan, A.T.; Aung, M.O.; Lee, K.H.; Tan, K.C.; Lim, S.G.; Bertoletti, A. Profile of tumor antigen-specific cd8 t cells in patients with hepatitis b virus-related hepatocellular carcinoma. Gastroenterology 2009, 137, 682–690. [Google Scholar] [CrossRef]
- Chen, C.H.; Chen, G.J.; Lee, H.S.; Huang, G.T.; Yang, P.M.; Tsai, L.J.; Chen, D.S.; Sheu, J.C. Expressions of cancer-testis antigens in human hepatocellular carcinomas. Cancer Lett. 2001, 164, 189–195. [Google Scholar] [CrossRef]
- Tahara, K.; Mori, M.; Sadanaga, N.; Sakamoto, Y.; Kitano, S.; Makuuchi, M. Expression of the mage gene family in human hepatocellular carcinoma. Cancer 1999, 85, 1234–1240. [Google Scholar] [CrossRef]
- Zhou, G.; Sprengers, D.; Boor, P.P.C.; Doukas, M.; Schutz, H.; Mancham, S.; Pedroza-Gonzalez, A.; Polak, W.G.; de Jonge, J.; Gaspersz, M.; et al. Antibodies against immune checkpoint molecules restore functions of tumor-infiltrating t cells in hepatocellular carcinomas. Gastroenterology 2017, 153, 1107–1119. [Google Scholar] [CrossRef]
- Tauber, C.; Schultheiss, M.; Luca, R.D.; Buettner, N.; Llewellyn-Lacey, S.; Emmerich, F.; Zehe, S.; Price, D.A.; Neumann-Haefelin, C.; Schmitt-Graeff, A.; et al. Inefficient induction of circulating taa-specific cd8+ t-cell responses in hepatocellular carcinoma. Oncotarget 2019, 10, 5194–5206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, C.U.; Haining, W.N.; Held, W.; Hogan, P.G.; Kallies, A.; Lugli, E.; Lynn, R.C.; Philip, M.; Rao, A.; Restifo, N.P.; et al. Defining ‘T cell exhaustion’. Nat. Rev. Immunol. 2019, 19, 665–674. [Google Scholar] [CrossRef]
- McLane, L.M.; Abdel-Hakeem, M.S.; Wherry, E.J. Cd8 t cell exhaustion during chronic viral infection and cancer. Annu. Rev. Immunol. 2019, 37, 457–495. [Google Scholar] [CrossRef] [Green Version]
- Inada, Y.; Mizukoshi, E.; Seike, T.; Tamai, T.; Iida, N.; Kitahara, M.; Yamashita, T.; Arai, K.; Terashima, T.; Fushimi, K.; et al. Characteristics of immune response to tumor-associated antigens and immune cell profile in patients with hepatocellular carcinoma. Hepatology 2019, 69, 653–665. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.D.; Song, G.W.; Park, S.; Jung, M.K.; Kim, M.H.; Kang, H.J.; Yoo, C.; Yi, K.; Kim, K.H.; Eo, S.; et al. Association between expression level of pd1 by tumor-infiltrating cd8(+) t cells and features of hepatocellular carcinoma. Gastroenterology 2018, 155, 1936–1950. [Google Scholar] [CrossRef]
- Finn, R.S.; Ryoo, B.Y.; Merle, P.; Kudo, M.; Bouattour, M.; Lim, H.Y.; Breder, V.V.; Edeline, J.; Chao, Y.; Ogasawara, S.; et al. Results of keynote-240: Phase 3 study of pembrolizumab (pembro) vs best supportive care (bsc) for second line therapy in advanced hepatocellular carcinoma (hcc). J. Clin. Oncol. 2019, 37, 4004. [Google Scholar] [CrossRef]
- Di Blasi, D.; Boldanova, T.; Mori, L.; Terracciano, L.; Heim, M.H.; De Libero, G. Unique t cell populations define immune-inflamed hepatocellular carcinoma. Cell. Mol. Gastroenterol. Hepatol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Aregay, A.; Owusu Sekyere, S.; Deterding, K.; Port, K.; Dietz, J.; Berkowski, C.; Sarrazin, C.; Manns, M.P.; Cornberg, M.; Wedemeyer, H. Elimination of hepatitis c virus has limited impact on the functional and mitochondrial impairment of hcv-specific cd8+ t cell responses. J. Hepatol. 2019, 71, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Hennecke, N.; Lohmann, V.; Kayser, A.; Neumann-Haefelin, C.; Kukolj, G.; Bocher, W.O.; Thimme, R. Restoration of hcv-specific cd8+ t cell function by interferon-free therapy. J. Hepatol. 2014, 61, 538–543. [Google Scholar] [CrossRef]
Figure 1.
Tumor antigens in the context of hepatocellular carcinoma (HCC). Tumor-specific antigens (neoantigens), tumor-associated antigens (TAA: overexpressed antigens, tumor testis antigens, and oncofetal antigens), and viral antigens are detectable in HCC patients.
Figure 1.
Tumor antigens in the context of hepatocellular carcinoma (HCC). Tumor-specific antigens (neoantigens), tumor-associated antigens (TAA: overexpressed antigens, tumor testis antigens, and oncofetal antigens), and viral antigens are detectable in HCC patients.

Figure 2.
Perspectives of CD8+ T cells in immunotherapeutic approaches. CD8+ T cells may have potential roles as targets and biomarkers in immunotherapy.
Figure 2.
Perspectives of CD8+ T cells in immunotherapeutic approaches. CD8+ T cells may have potential roles as targets and biomarkers in immunotherapy.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hofmann, M.; Tauber, C.; Hensel, N.; Thimme, R. CD8+ T Cell Responses during HCV Infection and HCC. J. Clin. Med. 2021, 10, 991. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050991
AMA Style
Hofmann M, Tauber C, Hensel N, Thimme R. CD8+ T Cell Responses during HCV Infection and HCC. Journal of Clinical Medicine. 2021; 10(5):991. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050991
Chicago/Turabian StyleHofmann, Maike, Catrin Tauber, Nina Hensel, and Robert Thimme. 2021. "CD8+ T Cell Responses during HCV Infection and HCC" Journal of Clinical Medicine 10, no. 5: 991. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10050991
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.