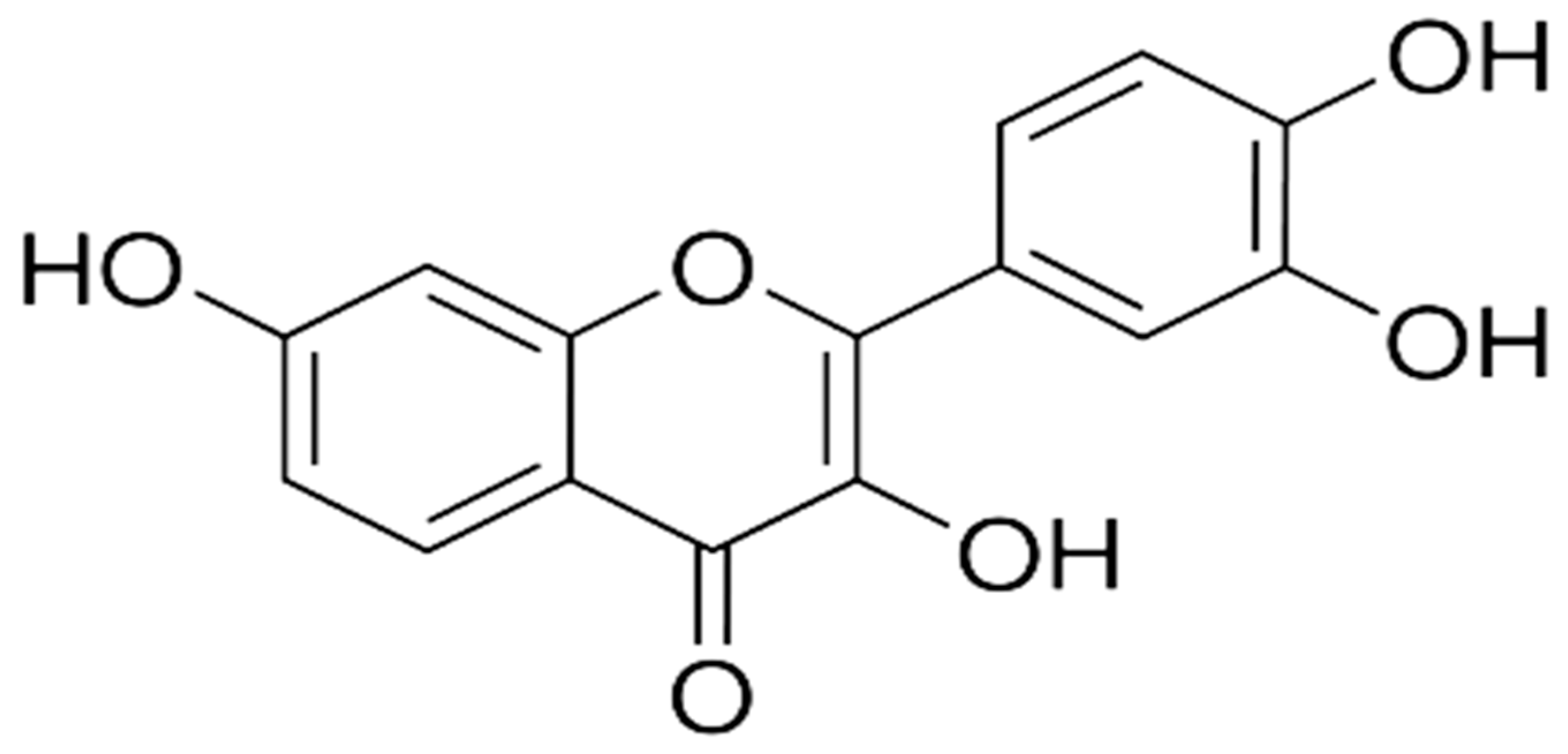
Phytomedicine-Based Potent Antioxidant, Fisetin Protects CNS-Insult LPS-Induced Oxidative Stress-Mediated Neurodegeneration and Memory Impairment


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals Used in the Experiment
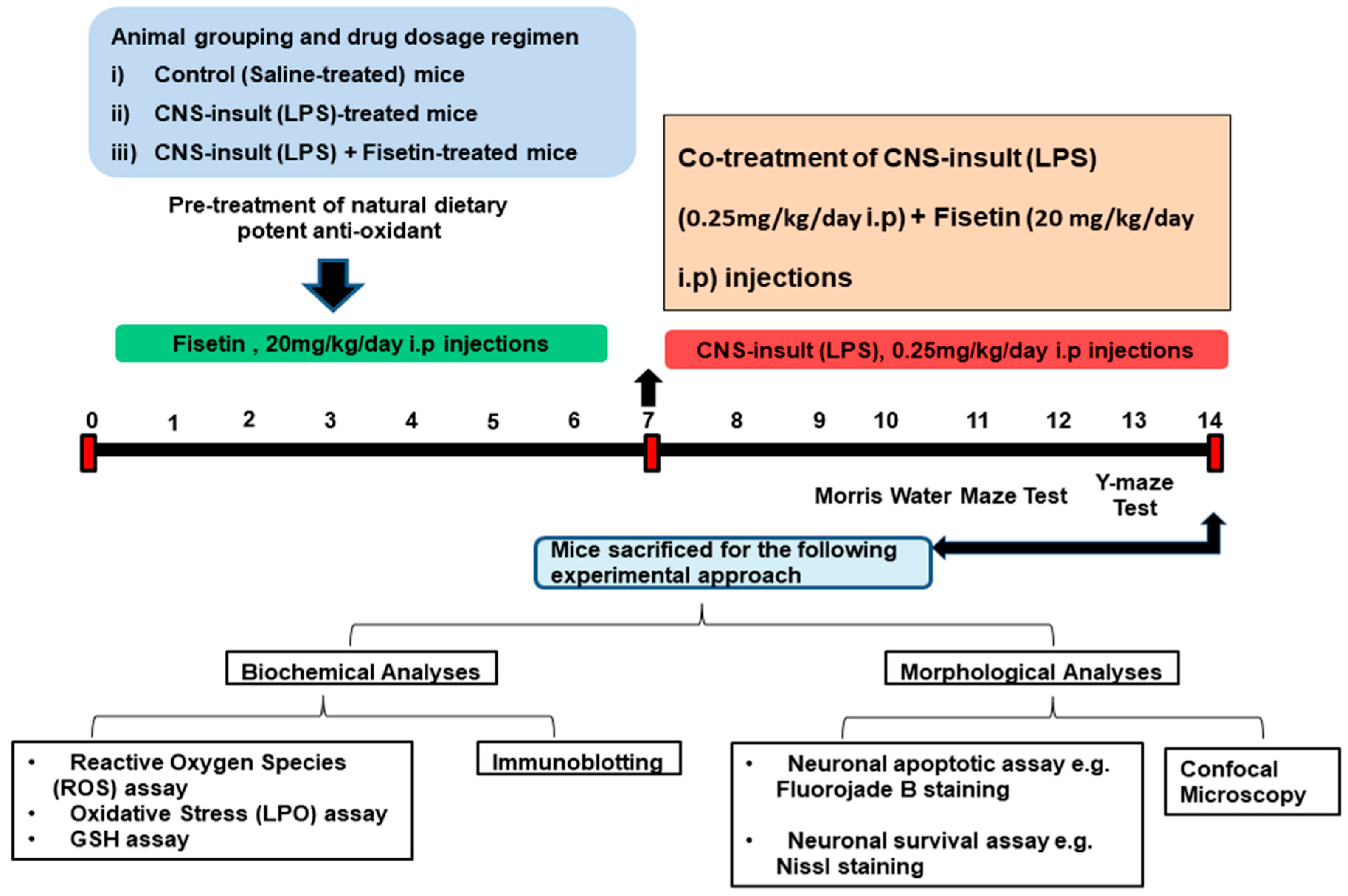
2.3. Study Designing, Animals Grouping, and Experimental Approach
2.4. Behavioral Studies
2.5. Protein Extraction from Mice Brains for Biochemical Analyses
2.6. Western Blot Analysis
2.7. Antibodies Used in the Western Blotting
2.8. Tissue Sample Preparation for Morphological Analysis
2.9. Immunofluorescence Staining
2.10. Fluoro-Jade B (FJB) Staining
2.11. Cresyl Violet (Nissl) Staining
2.12. ROS Assay in Mouse Hippocampus Homogenates
2.13. Lipid Peroxidation (LPO) Analysis in Mouse Hippocampus Homogenates
2.14. Glutathione (GSH) Analysis in Mouse Hippocampus Homogenates
2.15. Statistical Analysis
3. Results
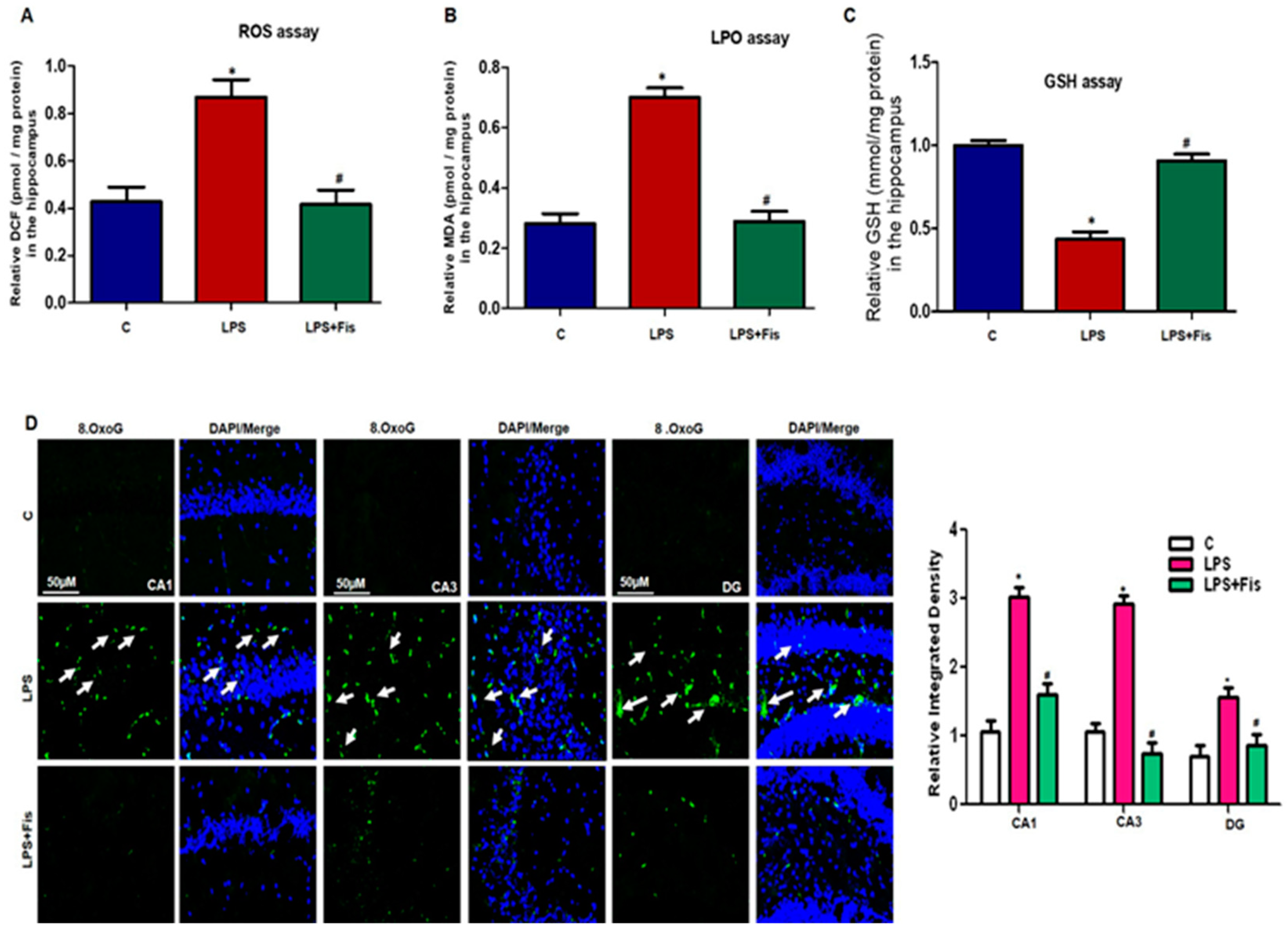
3.1. Effect of Fisetin Dosage Regimen on LPS-Induced Oxidative Stress in the Mouse Brain
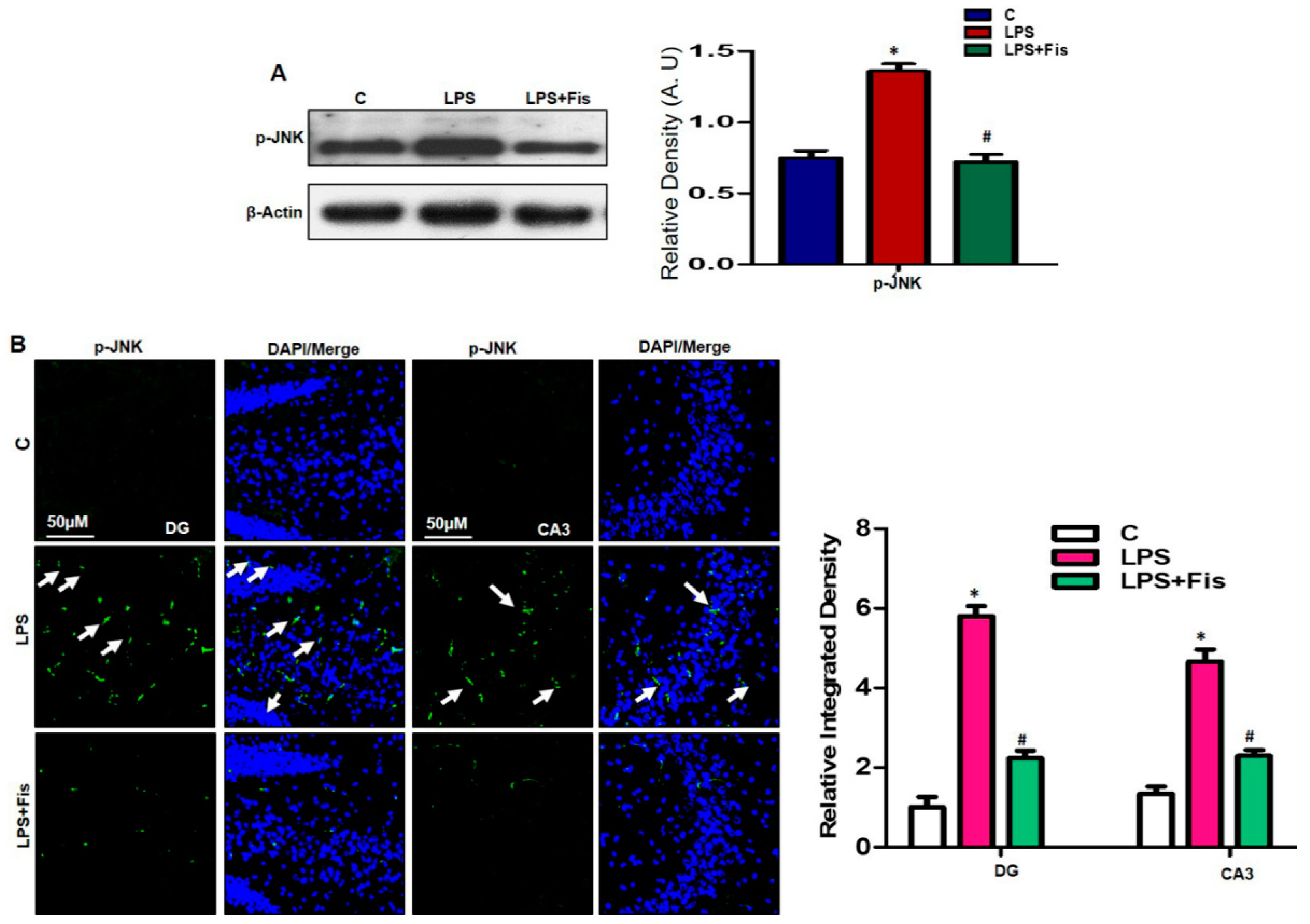
3.2. Effect of Fisetin Dosage on LPS-Induced Activation of p-JNK Expressions in the Mouse Hippocampus
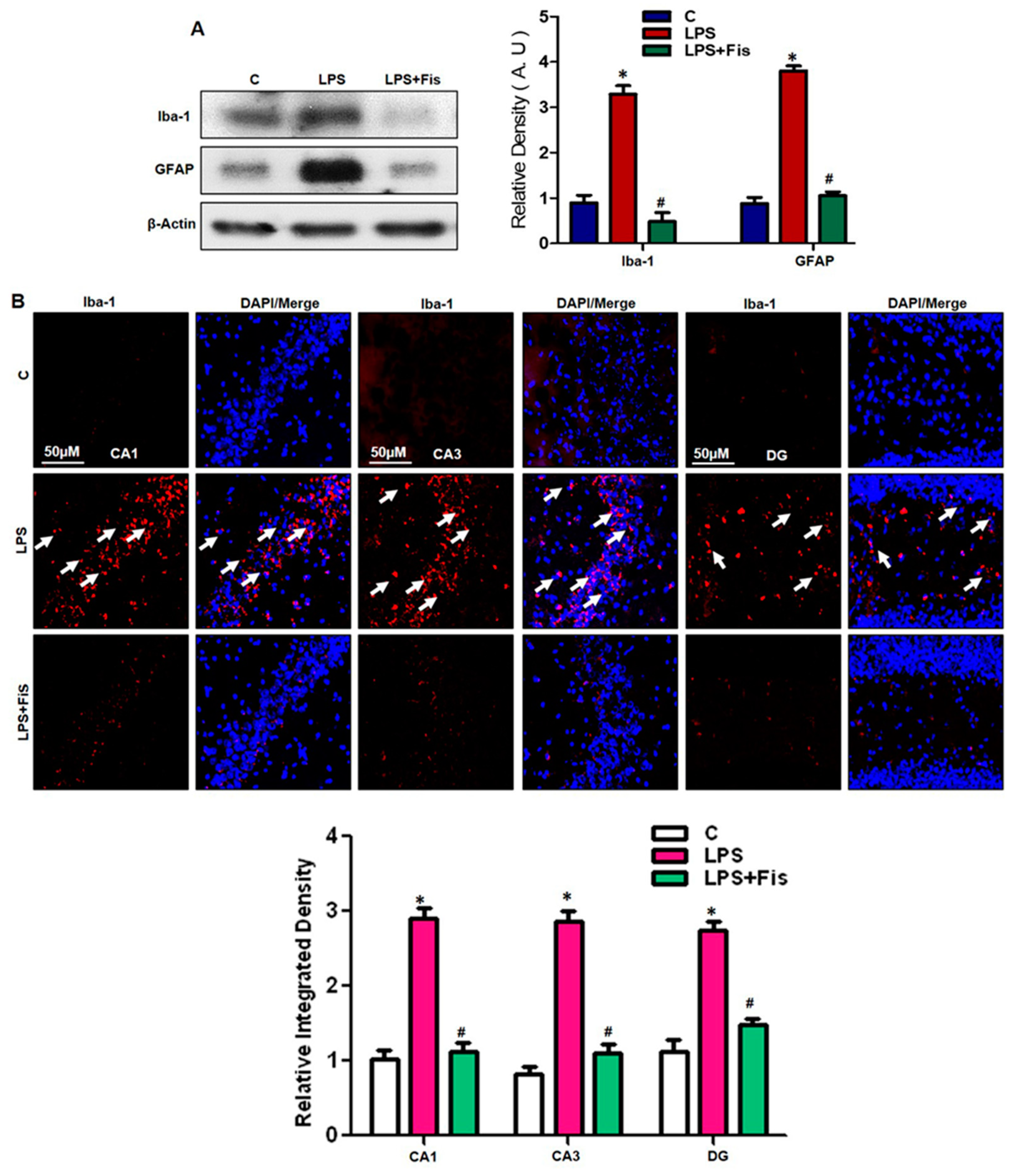
3.3. Effect of Fisetin on LPS-Induced Activation of Microglia and Astrocytes in the Adult Mouse Hippocampus
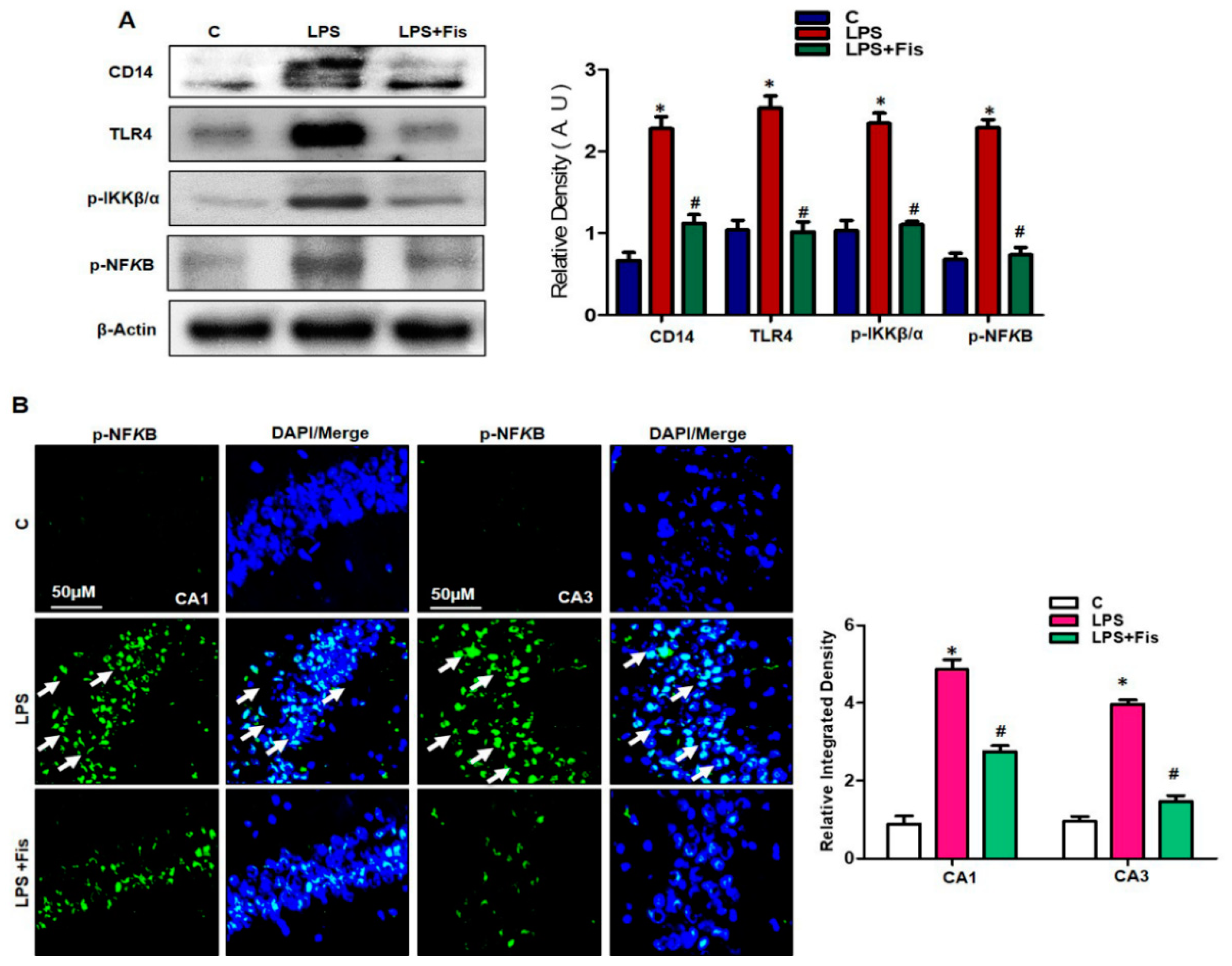
3.4. Effect of Fisetin on LPS-Induced Activation of TLR4/NFκB Signaling and Inflammatory Mediators in the Adult Mouse Hippocampus
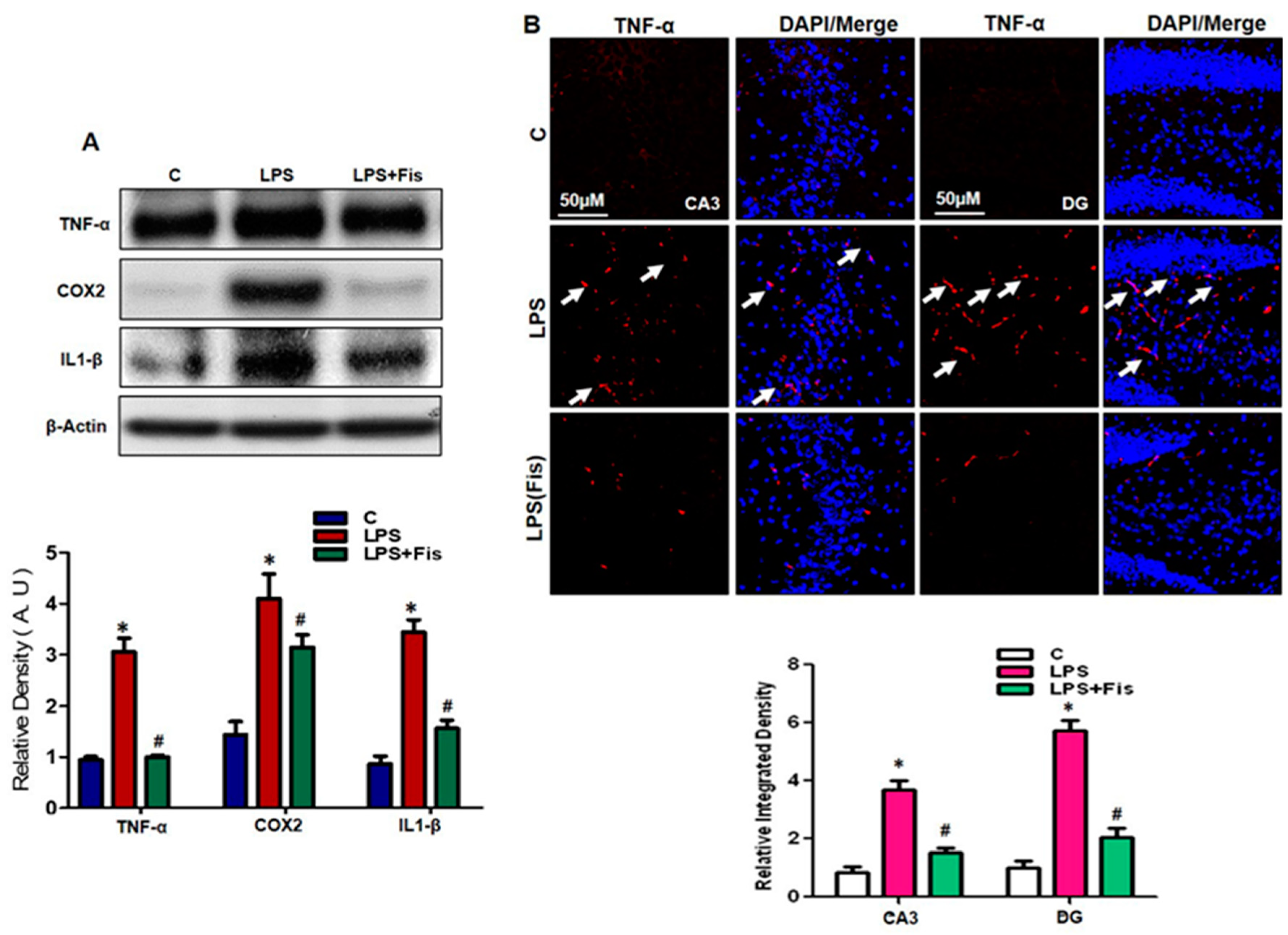
3.5. Effect of Fisetin on the LPS-Induced Upregulation of Inflammatory Mediators in the Adult Mouse Hippocampus
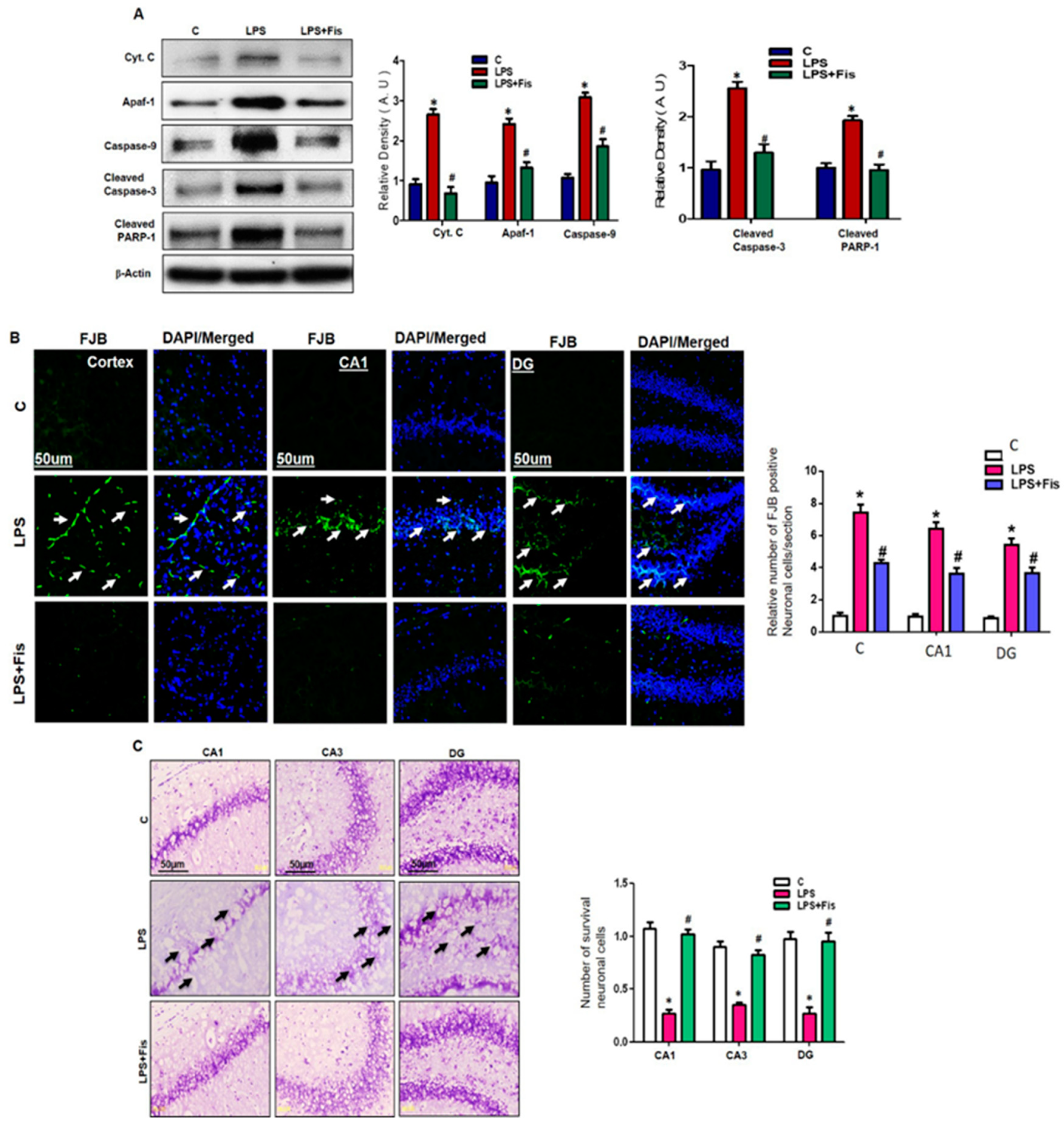
3.6. Effect of Fisetin on LPS-Induced Apoptotic Neurodegeneration in the Adult Mouse Brain
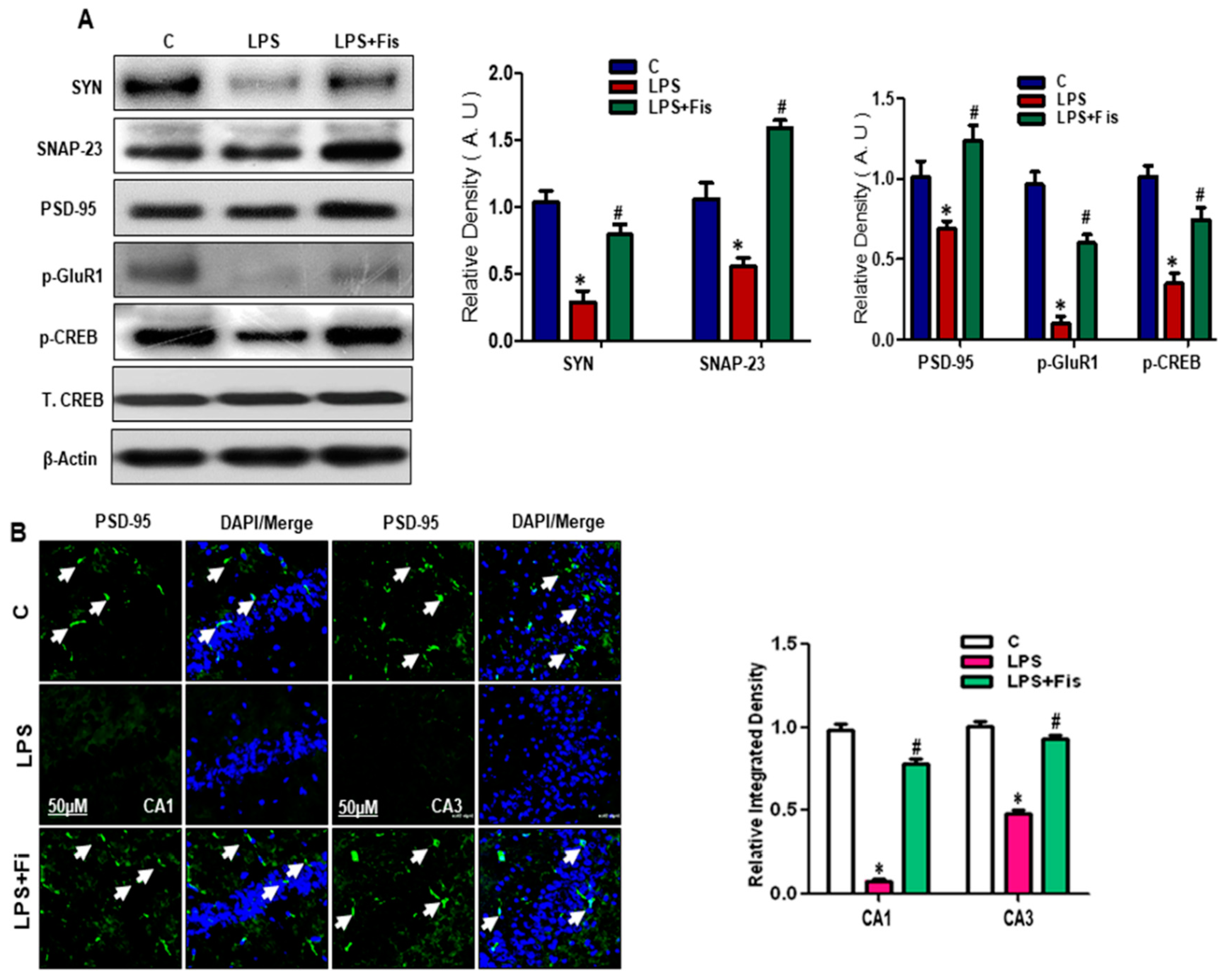
3.7. Effect of Fisetin on LPS-Induced Disruption of Pre- and Post-Synaptic and Memory Function in the Adult Mouse Hippocampus
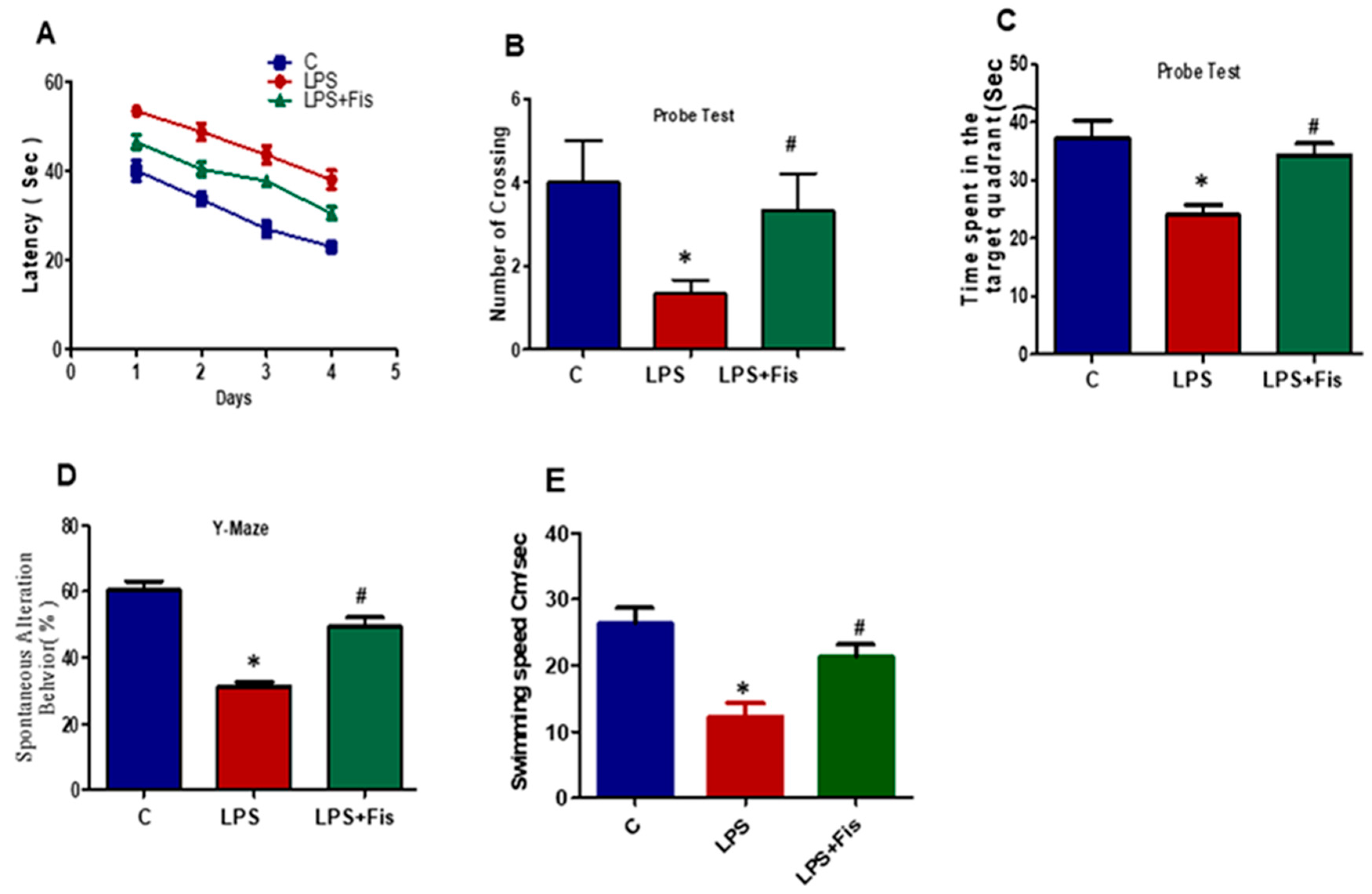
3.8. Effect of Fisetin on CNS-Insult, LPS-Induced Memory Dysfunction
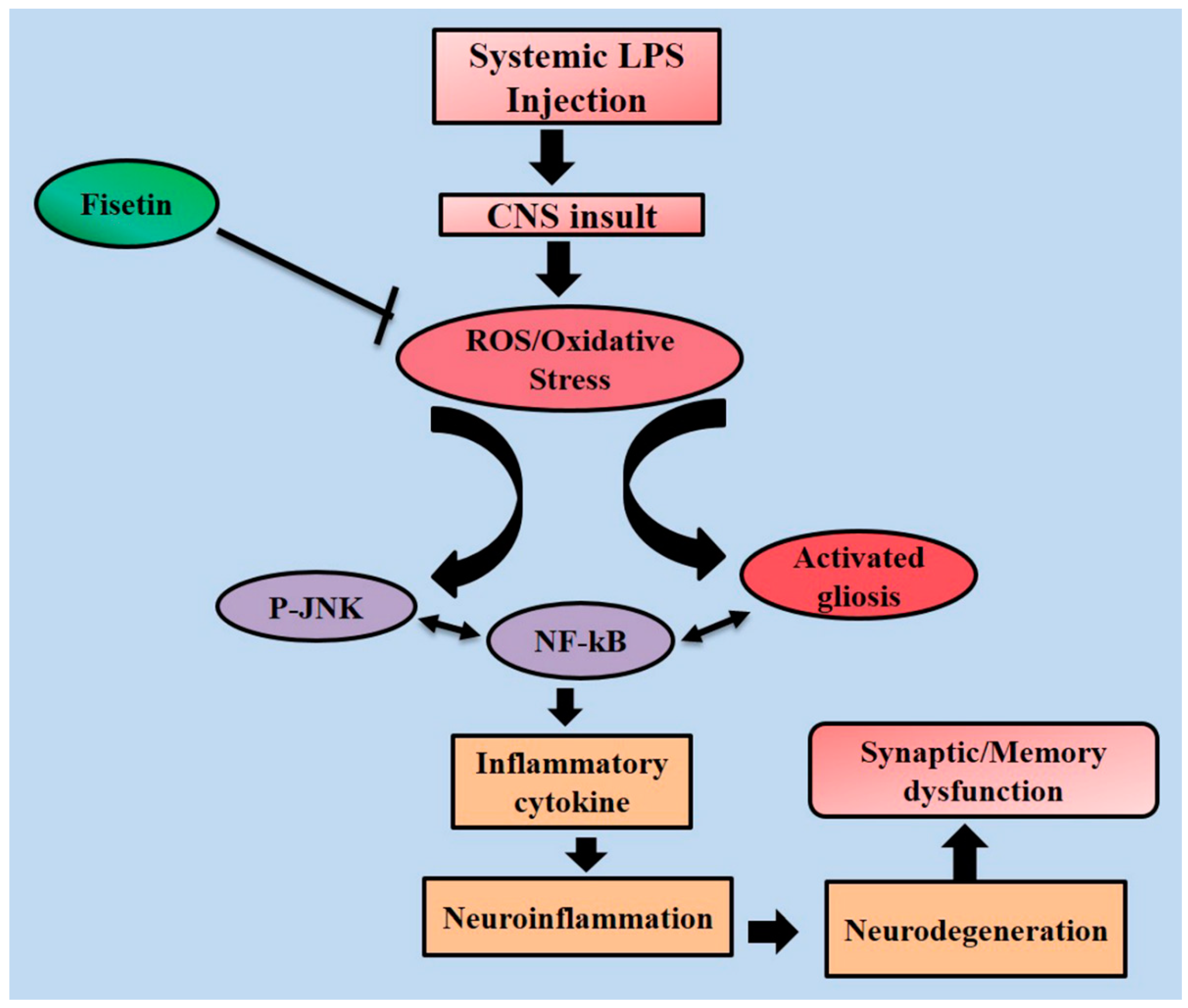
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brown, G.C.; Neher, J.J. Inflammatory neurodegeneration and mechanisms of microglial killing of neurons. Mol. Neurobiol. 2010, 41, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Von-Bernhardi, R.; Eugenín-von Bernhardi, L.; Eugenín, J. Microglial cell dysregulation in brain aging and neurodegeneration. Front. Aging Neurosci. 2015, 7, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skaper, S.D. The brain as a target for inflammatory processes and neuroprotective strategies. Ann. N. Y. Acad. Sci. 2007, 1122, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Jeong, J.W.; Lee, H.H.; Han, M.H.; Kim, G.Y.; Kim, W.J.; Choi, Y.H. Anti-inflammatory effects of genistein via suppression of the toll-like receptor 4-mediated signaling pathway in lipopolysaccharide-stimulated BV2 microglia. Chem. Biol. Interact. 2012, 214, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, C.; Kong, J. Oxidative stress in neurodegenerative diseases. Neural Regen. Res. 2012, 7, 376–385. [Google Scholar] [PubMed]
- Badshah, H.; Ali, T.; Rehman, S.; Amin, A.; Ullah, F.; Kim, T.H.; Kim, M.O. Protective effect of Lupeol against LPS-induced neuroinflammation via p38/JNK pathway in adult mice brain. J. Neuroimmune Pharmacol. 2016, 11, 48–60. [Google Scholar] [CrossRef]
- Khan, M.S.; Ali, T.; Abid, M.N.; Jo, M.H.; Khan, A.; Kim, M.W.; Yoon, G.H.; Cheon, E.W.; Rehman, S.U.; Kim, M.O. Lithium ameliorates lipopolysaccharide-induced neurotoxicity in the cortex and hippocampus of the adult rat brain. Neurochem. Int. 2017, 108, 343–354. [Google Scholar] [CrossRef]
- Khan, M.S.; Ali, T.; Kim, M.W.; Jo, M.H.; Jo, M.G.; Badshah, H.; Kim, M.O. Anthocyanins protect against LPS-induced oxidative stress-mediated neuroinflammation and neurodegeneration in the adult mouse cortex. Neurochem. Int. 2016, 100, 1–10. [Google Scholar] [CrossRef]
- Badshah, H.; Ali, T.; Kim, M.O. Osmotin attenuates LPS-induced neuroinflammation and memory impairments via the TLR4/NF-κB signalling pathway. Sci. Rep. 2016, 6, 24493. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic Neuro-inflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Ali, T.; Kim, M.W.; Jo, M.H.; Chung, J.I.; Kim, M.O. Anthocyanins improve hippocampus memory function and prevent neurodegeneration via JNK/Akt/GSK3β signaling in LPS-treated adult mice. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef]
- Noh, H.; Jeon, J.; Seo, H. Systemic injection of LPS induces region-specific neuroinflammation and mitochondrial dysfunction in normal mouse brain. Neurochem Int. 2014, 69, 35–40. [Google Scholar] [CrossRef]
- Font-Nieves, M.; Sans-Fons, M.G.; Gorina, R.; Bonfill-Teixidor, E.; Salas-Pérdomo, A.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. Induction of COX2 enzyme and downregulation of COX1 expression by lipopolysaccharide (LPS) control prostaglandin E2 production in astrocytes. J. Biol. Chem. 2012, 287, 6454–6468. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.A.; Basith, S.; Choi, S. Negative regulatory approaches to the attenuation of Toll-like receptor signaling. Exp. Mol. Med. 2013, 45, e11. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Li, M.; Ai, W.; He, L.; Lu, D.; Patrylo, P.R.; Cai, H.; Luo, X.; Li, Z.; Yan, X. Lipopolysaccharide-induced neuroinflammation is associated with Alzheimer-like amyloidogenic axonal pathology and dendritic degeneration in rats. Adv. Alzheimer Dis. 2014, 3, 78–93. [Google Scholar] [CrossRef]
- Ullah, R.; Khan, M.; Ali, S.S.; Saeed, K.; Kim, M.O. Natural antioxidant anthocyanins—A hidden therapeutic candidate in metabolic disorders with major focus in neurodegeneration. Nutrients 2019, 11, 1195. [Google Scholar] [CrossRef]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, U.R.; Kim, M.O. Hesperetin, a citrus flavonoid, attenuates LPS-induced neuroinflammation, apoptosis and memory impairments by modulating TLR4/NF-κB signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef]
- Ikram, M.; Muhammad, T.; Rehman, S.U.; Khan, A.; Jo, M.G.; Ali, T.; Kim, M.O. Hesperetin confers neuroprotection by regulating Nrf2/TLR4/NF-κB signaling in an Aβ mouse model. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef]
- Costa, S.L.; Silva, V.D.; Dos Santos Souza, C.; Santos, C.C.; Paris, I.; Muñoz, P.; Segura-Aguilar, J. Impact of plant-derived flavonoids on neurodegenerative diseases. Neurotox. Res. 2016, 30, 41–52. [Google Scholar] [CrossRef]
- Ali, T.; Kim, T.; Rehman, S.U.; Khan, M.S.; Amin, F.U.; Khan, M.; Ikram, M.; Kim, M.O. Natural dietary supplementation of anthocyanins via PI3K/Akt/Nrf2/HO-1 pathways mitigates oxidative stress, neurodegeneration, and memory impairment in a mouse model of Alzheimer’s disease. Mol. Neurobiol. 2017, 55, 6076–6093. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Shah, S.A.; Ali, T.; Chung, J.I.; Kim, M.O. Anthocyanins reversed D-galactose-induced oxidative stress and neuroinflammation mediated cognitive impairment in adult rats. Mol. Neurobiol. 2017, 54, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Badshah, H.; Ali, T.; Ahmad, A.; Kim, M.J.; Abid, N.B.; Shah, S.A.; Yoon, G.H.; Lee, H.Y.; Kim, M.O. Co-treatment with anthocyanins and vitamin C ameliorates ethanol-induced neurodegeneration via modulation of GABAB receptor signaling in the adult rat brain. CNS Neurol. Disord. Drug Targets 2015, 14, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Kim, M.J.; Rehman, S.U.; Ahmad, A.; Kim, M.O. Anthocyanin-loaded PEG-gold nanoparticles enhanced the neuroprotection of anthocyanins in an Aβ1-42 mouse model of Alzheimer’s disease. Mol. Neurobiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rengarajan, T.; Yaacob, N.S. The flavonoid fisetin as an anticancer agent targeting the growth signaling pathways. Eur. J. Pharm. 2016, 789, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Sahu, B.D.; Kumar, J.M.; Sistla, R. Fisetin a dietary flavonoid, ameliorates experimental colitis in mice: Relevance of NF-κB signaling. J. Nutr. Biochem. 2016, 28, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Leotoing, L.; Wauquier, F.; Guicheux, G.; Miot-Noirault, E.; Wittrant, Y.; Coxam, V. The polyphenol fisetin protects bone by repressing NF-κB and MKP-1-dependent signaling pathways in osteoclasts. PLoS ONE 2013, 8, e68388. [Google Scholar] [CrossRef]
- Maher, P.; Dargusch, R.; Ehren, J.L.; Okada, S.; Sharma, K.; Schubert, D. Fisetin lowers methyl glyoxal dependent protein glycation and limits the complications of diabetes. PLoS ONE 2011, 6, e21226. [Google Scholar] [CrossRef]
- Kim, S.C.; Kang, S.H.; Jeong, S.J.; Kim, S.H.; Ko, H.S. Inhibition of c-Jun N-terminal kinase and nuclear factor kappa B pathways mediates fisetin-exerted anti-inflammatory activity in lipopolysccharide-treated RAW264.7 cells. Immunopharmacol. Immunotoxicol. 2012, 34, 645–650. [Google Scholar] [CrossRef]
- Sechi, M.; Syed, D.N.; Pala, N.; Mariani, A.; Marceddu, S.; Brunetti, A.; Mukhtar, H.; Sanna, V. Nanoencapsulation of dietary flavonoid fisetin: Formulation and in vitro antioxidant and α-glucosidase inhibition activities. Mater. Sci. Eng. C. Mater. Biol. Appl. 2016, 68, 594–602. [Google Scholar] [CrossRef]
- Ahmad, A.; Ali, T.; Park, H.Y.; Badshah, H.; Rehman, S.U.; Kim, M.O. Neuroprotective effect of fisetin against amyloid-beta-induced cognitive/synaptic dysfunction, neuroinflammation, and neurodegeneration in adult mice. Mol. Neurobiol. 2017, 54, 2269–2285. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Ali, T.; Alam, S.I.; Ullah, R.; Zeb, A.; Lee, K.W.; Rutten, B.P.F.; Kim, M.O. 5-Ferulic Acid Rescues LPS-Induced Neurotoxicity via Modulation of the TLR4 Receptor in the Mouse Hippocampus. Mol. Neurobiol. 2017, 56, 2774–2790. [Google Scholar] [CrossRef]
- Lykhmus, O.; Mishra, N.; Koval, L.; Kalashnyk, O.; Gergalova, G.; Uspenska, K.; Komisarenko, S.; Soreq, H.; Skok, M. Molecular Mechanisms Regulating LPS-Induced Inflammation in the Brain. Front. Mol. Neurosci. 2016, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Liraz-Zaltsman, S.; Yaka, R.; Shabashov, D.; Shohami, E.; Biegon, A. Neuroinflammation-Induced Memory Deficits Are Amenable to Treatment with D-Cycloserine. J. Mol. Neurosci. 2016, 60, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Torika, N.; Asraf, K.; Danon, A.; Apte, R.N.; Fleisher-Berkovich, S. Telmisartan Modulates Glial Activation: In Vitro and In Vivo Studies. PLoS ONE 2016, 11, e0155823. [Google Scholar] [CrossRef]
- Moriyama, M.; Kurebayashi, R.; Kawabe, K.; Takano, K.; Nakamura, Y. Acetate attenuates lipopolysaccharide-induced nitric oxide production through an anti-oxidative mechanism in cultured primary rat astrocytes. Neurochem. Res. 2016, 41, 3138–3146. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Tong, J.; Tan, Y.; Zhu, S.; Yang, J.; Ji, M. Low molecular weight heparin prevents lipopolysaccharide induced-hippocampus-dependent cognitive impairments in mice. Int. J. Clin. Exp. Pathol. 2015, 8, 8881–8891. [Google Scholar]
- Santhanasabapathy, R.; Sudhandiran, G. Farnesol attenuates lipopolysaccharide-induced neurodegeneration in Swiss albino mice by regulating intrinsic apoptotic cascade. Brain Res. 2015, 16, 42–56. [Google Scholar] [CrossRef]
- Lee, Y.J.; Choi, D.Y.; Yun, Y.P.; Han, S.B.; Oh, K.W.; Hong, J.T. Epigallocatechin-3-gallate prevents systemic inflammation induced memory deficiency and amyloidogenesis via its anti-neuroinflammatory properties. J. Nutr. Biochem. 2013, 24, 298–310. [Google Scholar] [CrossRef]
- Sliwinska, A.; Kwiatkowski, D.; Czarny, P.; Toma, M.; Wigner, P.; Drzewoski, J.; Fabianowska-Majewska, K.; Szemraj, J.; Maes, M.; Galecki, P.; et al. The levels of 7,8-dihydrodeoxyguanosine (8-oxoG)and 8 oxoguanine DNA glycosylase 1 (OGG1)A potential diagnostic biomarkers of Alzheimer’s disease. J. Neurol. Sci. 2016, 368, 155–159. [Google Scholar] [CrossRef]
- Lovell, M.A.; Ehmann, W.D.; Butler, S.M.; Markesbery, W.R. Elevated thiobarbituric acid-reactive substances and antioxidant enzyme activity in the brain in Alzheimer’s disease. Neurology 1995, 45, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Tucsek, Z.; Radnai, B.; Racz, B.; Debreceni, B.; Priber, J.K.; Dolowschiak, T.; Palkovics, T.; Gallyas, F.J.; Sumegi, B.; Veres, B. Suppressing LPS-induced early signal transduction in macrophages by a polyphenol degradation product: A critical role of MKP-1. J. Leukoc. Biol. 2011, 89, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, P.; MacGarvey, U.; Beal, M.F. Oxidative damage to mitochondrial DNA is increased in Alzheimer’s disease. Ann. Neurol. 1994, 36, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Lin, H.; Tu, Q.; Liu, J.; Li, X. Fisetin protects DNA against oxidative damage and its possible mechanism. Adv. Pharm. Bull. 2016, 6, 267–270. [Google Scholar] [CrossRef] [PubMed]
- González-Scarano, F.; Baltuch, G. Microglia as mediators of inflammatory and degenerative diseases. Annu. Rev. Neurosci. 1999, 22, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Waetzig, V.; Czeloth, K.; Hidding, U.; Mielke, K.; Kanzow, M.; Brecht, S.; Goetz, M.; Lucius, R.; Herdegen, T.; Hanisch, U.K. c-Jun N-terminal kinases (JNKs) mediate pro-inflammatory actions of microglia. Glia 2005, 50, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, R.; Lu, F.; Xu, F.; Zheng, J.; Li, Z.; Cui, W.; Wang, C.; Zhang, J.; Xu, S.; et al. Fast Green FCF Attenuates Lipopolysaccharide-Induced Depressive-Like Behavior and Downregulates TLR4/Myd88/NF-κB Signal Pathway in the Mouse Hippocampus. Front. Pharmacol. 2019, 10, 501. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, L.; Wang, L.; Qian, S.; Zhang, D.; Fang, J.; Pan, J. Berberine protects human umbilical vein endothelial cells against LPS-induced apoptosis by blocking JNK-mediated signaling. Evid. Based Complement. Alternat. Med. 2016, 69, 839–856. [Google Scholar] [CrossRef]
- Wang, G.; Liu, L.; Zhang, Y.; Han, D.; Liu, J.; Xu, J.; Xie, X.; Wu, Y.; Zhang, D.; Ke, R.; et al. Activation of PPARγ attenuates LPS-induced acute lung injury by inhibition of HMGB1-RAGE levels. Eur. J. Pharmacol. 2014, 726, 27–32. [Google Scholar] [CrossRef]
- Go, M.; Kou, J.; Lim, J.E.; Yang, J.; Fukuchi, K.I. Microglial response to LPS increases in wild-type mice during aging but diminishes in an Alzheimer’s mouse model: Implication of TLR4 signaling in disease progression. Biochem. Biophys. Res. Commun. 2016, 479, 331–337. [Google Scholar] [CrossRef]
- Cheong, M.H.; Lee, S.R.; Yoo, H.S.; Jeong, J.W.; Kim, G.Y.; Kim, W.J.; Jung, I.C.; Choi, Y.H. Anti-inflammatory effects of Polygala tenuifolia root through inhibition of NF-κB activation inlipopolysaccharide-induced BV2 microglial cells. J. Ethnopharmacol. 2011, 137, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.T.; Bian, C.; Yuan, J.C.; Chu, W.H.; Xiang, X.; Chen, F.; Wang, C.S.; Feng, H.; Lin, J.K. Curcumin attenuates acute inflammatory injury by inhibiting the TLR4/MyD88/NF-κB signaling pathway in experimental traumatic brain injury. J. Neuroinflammation. 2014, 27, 59. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.W.; Zhao, M.B.; Ma, Z.Z.; Jiang, Y.; Tu, P.F. Protosappanin A inhibits oxidative and nitrative stress via interfering the interaction of transmembrane proteinCD14 with Toll-like receptor-4 in lipopolysaccharide-induced BV-2 microglia. Int. Immunopharmacol. 2012, 14, 558–569. [Google Scholar] [CrossRef]
- Yang, C.; Yu, L.; Kong, L.; Ma, R.; Zhang, J.; Zhu, Q.; Zhu, J.; Hao, D. Pyrroloquinoline quinone (PQQ) inhibits lipopolysaccharide induced inflammation in part via downregulated NF-κB and p38/JNK activation in microglial and attenuates microglia activation in lipopolysaccharide treatment mice. PLoS ONE 2014, 9, e109502. [Google Scholar] [CrossRef]
- Medvedev, A.E.; Lentschat, A.; Wahl, L.; Golenbock, D.T.; Vogel, S.N. Dysregulated LPS-induced TLR4-MyD88 complex formation and IRAK-1 activation in endotoxin tolerant human monocytes. FASEB J. 2002, 16, A287. [Google Scholar]
- McGeer, P.L.; McGeer, E.G. Inflammation, autotoxicity and Alzheimer disease. Neurobiol. Aging 2001, 22, 799–809. [Google Scholar] [CrossRef]
- Von Bernhardi, R.; Tichauer, J.E.; Eugenín, J. Aging-dependent changes of microglial cells and the irrelevance for neurodegenerative disorders. J. Neurochem. 2010, 112, 1099–1114. [Google Scholar] [CrossRef]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef]
- McGeer, P.L.; Itagaki, S.; Tago, H.; McGeer, E.G. Reactive microglia in patients with senile dementia of the Alzheimer type are positive for the histocompatibility glycoprotein HLA-DR. Neurosci. Lett. 1987, 79, 195–200. [Google Scholar] [CrossRef]
- Veerhuis, R.; Janssen, I.; De Groot, C.J.; Van Muiswinkel, F.L.; Hack, C.E.; Eikelenboom, P. Cytokines associated with amyloid plaques in Alzheimer’s disease brain stimulate human glial and neuronal cell cultures to secrete early complement proteins, but not C1-inhibitor. Exp. Neurol. 1999, 160, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Valero, J.; Mastrella, G.; Neiva, I.; Sánchez, S.; Malva, J.O. Long-term effects of an acute and systemic administration of LPS on adult neurogenesis and spatial memory. Front. Neurosci. 2014, 21, 83. [Google Scholar] [CrossRef] [PubMed]
- Ishige, K.; Schubert, D.; Sagara, Y. Flavonoids protect neuronal cells from oxidative stress by three distinct mechanisms. Free Radic. Biol. Med. 2011, 30, 433–446. [Google Scholar] [CrossRef]
- Maher, P.; Akaishi, T.; Abe, K. Flavonoid fisetin promotes ERK-dependent long-term potentiation and enhances memory. Proc. Natl. Acad. Sci. USA 2006, 103, 16568. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, B.; Lu, J.; Shi, H.; Gong, S.; Wang, Y.; Hamdy, R.C.; Chua, B.H.L.; Yang, L.; Xu, X. Fisetin provides antidepressant effects by activating the tropomyosin receptor kinase B signal pathway in mice. J. Neurochem. 2017, 143, 561–568. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Application | Manufacturer | Catalog Number | Concentration |
|---|---|---|---|---|---|
| Iba-1 | Rabbit | WB/IF | Santa Cruz Biotechnology, USA | SC: 98468 | 1:1000/1:100 |
| GFAP | Mouse | WB | = | SC: 33673 | 1:1000 |
| p-JNK | Mouse | WB/IF | = | SC: 6254 | 1:1000/1:100 |
| TLR-4 | Goat | WB | = | SC: 16240 | 1:1000 |
| p-NF-κB | Mouse | WB/IF | = | SC 8008 | 1:1000/1:100 |
| CD14 | Mouse | WB | = | SC: 58951 | 1:1000 |
| TNF-α | Mouse | WB/IF | = | SC: 8436 | 1:1000/1:100 |
| COX-2 | Rabbit | WB | = | SC: 7951 | 1:1000 |
| IL-1β | Mouse | IF | = | SC: 32294 | 1:100 |
| Apaf-1 | Mouse | WB | = | SC: 65891 | 1:1000 |
| Cyto. c | Mouse | WB | = | SC: 13156 | 1:1000 |
| PARP-1 | Mouse | WB | = | SC: 8007 | 1:1000 |
| Caspase-3 | Mouse | WB | = | SC: 7272 | 1:1000 |
| PSD-95 | Mouse | WB/IF | = | SC: 71933 | 1:1000/1:100 |
| Synaptophysin | Rabbit | WB | = | SC: 17750 | 1:1000 |
| SNAP-23 | Mouse | WB | = | SC: 374215 | 1:100 |
| Caspase-9 | Rabbit | WB | Cell Signaling, USA | 9508S | 1:1000 |
| p-CREB (Ser 133) | Rabbit | WB | = | 9198S | 1:1000 |
| p-GluR1 (Ser 845) | Rabbit | WB | = | 8084S | 1:1000 |
| p-IKKBβ/α | Rabbit | WB | Abcam, USA | Ab59195 | 1:1000 |
| 8-OxoG | Mouse | IF | Millipore | MAB3560 | 1:100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Ali, T.; Rehman, S.U.; Kim, M.O. Phytomedicine-Based Potent Antioxidant, Fisetin Protects CNS-Insult LPS-Induced Oxidative Stress-Mediated Neurodegeneration and Memory Impairment. J. Clin. Med. 2019, 8, 850. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8060850
Ahmad A, Ali T, Rehman SU, Kim MO. Phytomedicine-Based Potent Antioxidant, Fisetin Protects CNS-Insult LPS-Induced Oxidative Stress-Mediated Neurodegeneration and Memory Impairment. Journal of Clinical Medicine. 2019; 8(6):850. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8060850
Chicago/Turabian StyleAhmad, Ashfaq, Tahir Ali, Shafiq Ur Rehman, and Myeong Ok Kim. 2019. "Phytomedicine-Based Potent Antioxidant, Fisetin Protects CNS-Insult LPS-Induced Oxidative Stress-Mediated Neurodegeneration and Memory Impairment" Journal of Clinical Medicine 8, no. 6: 850. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8060850