Pituitary Adenylate Cyclase-Activating Polypeptide: 30 Years in Research Spotlight and 600 Million Years in Service

Abstract
:1. Introduction
2. Emerging from the Depths of Evolution: PACAP/Glucagon Family
3. PACAP Receptors and Signal Transduction Bias
4. When PACAP1-38 Hurts
4.1. Involvement in a Migraine Attack
4.2. Contribution to Post-Traumatic Stress Disorder (PTSD)
5. When PACAP1-38 Rescues
5.1. Alzheimer’s Disease and PACAP1-38
5.2. Ischemia/Stroke
6. When PACAP1-38 Appears to be Janus-Faced
6.1. Role of PACAP1-38 in Inflammation
6.2. Roles of PACAP in Stem Cell Regulation and Cancer Formation
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Miyata, A.; Jiang, L.; Dahl, R.D.; Kitada, C.; Kubo, K.; Fujino, M.; Minamino, N.; Arimura, A. Isolation of a neuropeptide corresponding to the N-terminal 27 residues of the pituitary adenylate cyclase activating polypeptide with 38 residues (PACAP38). Biochem. Biophys. Res. Commun. 1990, 170, 643–648. [Google Scholar] [CrossRef]
- Krashes, M.J.; Shah, B.P.; Madara, J.C.; Olson, D.P.; Strochlic, D.E.; Garfield, A.S.; Vong, L.; Pei, H.; Watabe-Uchida, M.; Uchida, N.; et al. An excitatory paraventricular nucleus to AgRP neuron circuit that drives hunger. Nature 2014, 507, 238–242. [Google Scholar] [CrossRef]
- Ressler, K.J.; Mercer, K.B.; Bradley, B.; Jovanovic, T.; Mahan, A.; Kerley, K.; Norrholm, S.D.; Kilaru, V.; Smith, A.K.; Myers, A.J.; et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011, 470, 492–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spengler, D.; Waeber, C.; Pantaloni, C.; Holsboer, F.; Bockaert, J.; Seeburg, P.H.; Journot, L. Differential signal transduction by five splice variants of the PACAP receptor. Nature 1993, 365, 170–175. [Google Scholar] [PubMed]
- Zhong, Y. Mediation of PACAP-like neuropeptide transmission by coactivation of Ras/Raf and cAMP signal transduction pathways in Drosophila. Nature 1995, 375, 588–592. [Google Scholar] [CrossRef]
- Harding, S.D.; Sharman, J.L.; Faccenda, E.; Southan, C.; Pawson, A.J.; Ireland, S.; Gray, A.J.G.; Bruce, L.; Alexander, S.P.H.; Anderton, S.; et al. The IUPHAR/BPS Guide to PHARMACOLOGY in 2018: Updates and expansion to encompass the new guide to IMMUNOPHARMACOLOGY. Nucleic Acids Res. 2018, 46, D1091–D1106. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Reis, D.; Ribeiro, J.A.; de Almeida, R.F.M.; Sebastiao, A.M. VPAC1 and VPAC2 receptor activation on GABA release from hippocampal nerve terminals involve several different signalling pathways. Br. J. Pharmacol. 2017, 174, 4725–4737. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Claus, T.H.; Liang, Y.; Li, Y.; Yang, L.; Zhu, J.; Dela Cruz, F.; Peng, X.; Chen, H.; Yung, S.L.; et al. A potent and highly selective VPAC2 agonist enhances glucose-induced insulin release and glucose disposal: A potential therapy for type 2 diabetes. Diabetes 2002, 51, 1453–1460. [Google Scholar] [CrossRef]
- McRory, J.; Sherwood, N.M. Two protochordate genes encode pituitary adenylate cyclase-activating polypeptide and related family members. Endocrinology 1997, 138, 2380–2390. [Google Scholar] [CrossRef]
- Blechman, J.; Levkowitz, G. Alternative Splicing of the Pituitary Adenylate Cyclase-Activating Polypeptide Receptor PAC1: Mechanisms of Fine Tuning of Brain Activity. Front. Endocrinol. (Lausanne) 2013, 4, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denes, V.; Czotter, N.; Lakk, M.; Berta, G.; Gabriel, R. PAC1-expressing structures of neural retina alter their PAC1 isoform splicing during postnatal development. Cell Tissue Res. 2014, 355, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Lakk, M.; Szabo, B.; Volgyi, B.; Gabriel, R.; Denes, V. Development-related splicing regulates pituitary adenylate cyclase-activating polypeptide (PACAP) receptors in the retina. Invest. Ophthalmol. Vis. Sci. 2012, 53, 7825–7832. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Arimura, A. PACAP: The road to discovery. Peptides 2007, 28, 1617–1619. [Google Scholar] [CrossRef] [PubMed]
- Tams, J.W.; Johnsen, A.H.; Fahrenkrug, J. Identification of pituitary adenylate cyclase-activating polypeptide1-38-binding factor in human plasma, as ceruloplasmin. Biochem. J. 1999, 341, 271–276. [Google Scholar] [CrossRef]
- Filipsson, K.; Tornoe, K.; Holst, J.; Ahren, B. Pituitary adenylate cyclase-activating polypeptide stimulates insulin and glucagon secretion in humans. J. Clin. Endocrinol. Metab. 1997, 82, 3093–3098. [Google Scholar]
- American College of Physicians, Normal Laboratory Values. Available online: www.acponline.org (accessed on 16 September 2019).
- Du, Q.; Kawabe, Y.; Schilde, C.; Chen, Z.H.; Schaap, P. The Evolution of Aggregative Multicellularity and Cell-Cell Communication in the Dictyostelia. J. Mol. Biol. 2015, 427, 3722–3733. [Google Scholar] [CrossRef]
- Nordstrom, K.J.; Sallman Almen, M.; Edstam, M.M.; Fredriksson, R.; Schioth, H.B. Independent HHsearch, Needleman—Wunsch-based, and motif analyses reveal the overall hierarchy for most of the G protein-coupled receptor families. Mol. Biol. Evol. 2011, 28, 2471–2480. [Google Scholar] [CrossRef] [PubMed]
- Foord, S.M.; Bonner, T.I.; Neubig, R.R.; Rosser, E.M.; Pin, J.P.; Davenport, A.P.; Spedding, M.; Harmar, A.J. International Union of Pharmacology. XLVI. G protein-coupled receptor list. Pharmacol. Rev. 2005, 57, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, E.V.; Roy, A.; Ciechanski, P.; Wilson, R.J. Stress peptide PACAP stimulates and stabilizes neonatal breathing through distinct mechanisms. Respir. Physiol. Neurobiol. 2013, 187, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Eiden, L.E. Two ancient neuropeptides, PACAP and AVP, modulate motivated behavior at synapses in the extrahypothalamic brain: A study in contrast. Cell Tissue Res. 2019, 375, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Jozsa, R.; Nemeth, J.; Tamas, A.; Hollosy, T.; Lubics, A.; Jakab, B.; Olah, A.; Lengvari, I.; Arimura, A.; Reglodi, D. Short-term fasting differentially alters PACAP and VIP levels in the brains of rat and chicken. Ann. N. Y. Acad. Sci. 2006, 1070, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, B.D.; Abad, C.; Chhith, S.; Cheung-Lau, G.; Hajji, O.E.; Nobuta, H.; Waschek, J.A. Impaired nerve regeneration and enhanced neuroinflammatory response in mice lacking pituitary adenylyl cyclase activating peptide. Neuroscience 2008, 151, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waschek, J.A.; Cohen, J.R.; Chi, G.C.; Proszynski, T.J.; Niewiadomski, P. PACAP Promotes Matrix-Driven Adhesion of Cultured Adult Murine Neural Progenitors. ASN Neuro 2017, 9, 1759091417708720. [Google Scholar] [CrossRef]
- Hamelink, C.; Tjurmina, O.; Damadzic, R.; Young, W.S.; Weihe, E.; Lee, H.W.; Eiden, L.E. Pituitary adenylate cyclase-activating polypeptide is a sympathoadrenal neurotransmitter involved in catecholamine regulation and glucohomeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 461–466. [Google Scholar] [CrossRef]
- Zibara, K.; Zeidan, A.; Mallah, K.; Kassem, N.; Awad, A.; Mazurier, F.; Badran, B.; El-Zein, N. Signaling pathways activated by PACAP in MCF-7 breast cancer cells. Cell Signal. 2018, 50, 37–47. [Google Scholar] [CrossRef]
- Jung, S.; Yi, L.; Jeong, D.; Kim, J.; An, S.; Oh, T.J.; Kim, C.H.; Kim, C.J.; Yang, Y.; Kim, K.I.; et al. The role of ADCYAP1, adenylate cyclase activating polypeptide 1, as a methylation biomarker for the early detection of cervical cancer. Oncol. Rep. 2011, 25, 245–252. [Google Scholar]
- Lee, J.H.; Lee, J.Y.; Rho, S.B.; Choi, J.S.; Lee, D.G.; An, S.; Oh, T.; Choi, D.C.; Lee, S.H. PACAP inhibits tumor growth and interferes with clusterin in cervical carcinomas. FEBS Lett. 2014, 588, 4730–4739. [Google Scholar] [CrossRef] [Green Version]
- Germano, P.M.; Le, S.V.; Oh, D.S.; Fan, R.; Lieu, S.; Siu, A.; Pisegna, J.R. Differential coupling of the PAC1 SV1 splice variant on human colonic tumors to the activation of intracellular cAMP but not intracellular Ca2+ does not activate tumor proliferation. J. Mol. Neurosci. 2004, 22, 83–92. [Google Scholar] [CrossRef]
- Liu, S.; Zeng, Y.; Li, Y.; Guo, W.; Liu, J.; Ouyang, N. VPAC1 overexpression is associated with poor differentiation in colon cancer. Tumour Biol. 2014, 35, 6397–6404. [Google Scholar] [CrossRef] [PubMed]
- Le, S.V.; Yamaguchi, D.J.; McArdle, C.A.; Tachiki, K.; Pisegna, J.R.; Germano, P. PAC1 and PACAP expression, signaling, and effect on the growth of HCT8, human colonic tumor cells. Regul. Pept. 2002, 109, 115–125. [Google Scholar] [CrossRef]
- Schulz, S.; Rocken, C.; Mawrin, C.; Weise, W.; Hollt, V.; Schulz, S. Immunocytochemical identification of VPAC1, VPAC2, and PAC1 receptors in normal and neoplastic human tissues with subtype-specific antibodies. Clin. Cancer Res. 2004, 10, 8235–8242. [Google Scholar] [CrossRef] [PubMed]
- Lauffer, J.M.; Tang, L.H.; Zhang, T.; Hinoue, T.; Rahbar, S.; Odo, M.; Modlin, I.M.; Kidd, M. PACAP mediates the neural proliferative pathway of Mastomys enterochromaffin-like cell transformation. Regul. Pept. 2001, 102, 157–164. [Google Scholar] [CrossRef]
- Barbarin, A.; Seite, P.; Godet, J.; Bensalma, S.; Muller, J.M.; Chadeneau, C. Atypical nuclear localization of VIP receptors in glioma cell lines and patients. Biochem. Biophys. Res. Commun. 2014, 454, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Cochaud, S.; Meunier, A.C.; Monvoisin, A.; Bensalma, S.; Muller, J.M.; Chadeneau, C. Neuropeptides of the VIP family inhibit glioblastoma cell invasion. J. Neurooncol. 2015, 122, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Moody, T.W.; Osefo, N.; Nuche-Berenguer, B.; Ridnour, L.; Wink, D.; Jensen, R.T. Pituitary adenylate cyclase-activating polypeptide causes tyrosine phosphorylation of the epidermal growth factor receptor in lung cancer cells. J. Pharmacol. Exp. Ther. 2012, 341, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Giunta, S.; Castorina, A.; Adorno, A.; Mazzone, V.; Carnazza, M.L.; D’Agata, V. PACAP and VIP affect NF1 expression in rat malignant peripheral nerve sheath tumor (MPNST) cells. Neuropeptides 2010, 44, 45–51. [Google Scholar] [CrossRef]
- Thouennon, E.; Pierre, A.; Tanguy, Y.; Guillemot, J.; Manecka, D.L.; Guerin, M.; Ouafik, L.; Muresan, M.; Klein, M.; Bertherat, J.; et al. Expression of trophic amidated peptides and their receptors in benign and malignant pheochromocytomas: High expression of adrenomedullin RDC1 receptor and implication in tumoral cell survival. Endocr. Relat. Cancer 2010, 17, 637–651. [Google Scholar] [CrossRef]
- Cohen, J.R.; Resnick, D.Z.; Niewiadomski, P.; Dong, H.; Liau, L.M.; Waschek, J.A. Pituitary adenylyl cyclase activating polypeptide inhibits gli1 gene expression and proliferation in primary medulloblastoma derived tumorsphere cultures. BMC Cancer 2010, 10, 676. [Google Scholar] [CrossRef]
- Lelievre, V.; Seksenyan, A.; Nobuta, H.; Yong, W.H.; Chhith, S.; Niewiadomski, P.; Cohen, J.R.; Dong, H.; Flores, A.; Liau, L.M.; et al. Disruption of the PACAP gene promotes medulloblastoma in ptc1 mutant mice. Dev. Biol. 2008, 313, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zia, H.; Leyton, J.; Casibang, M.; Hau, V.; Brenneman, D.; Fridkin, M.; Gozes, I.; Moody, T.W. (N-stearyl, norleucine17) VIP hybrid inhibits the growth of pancreatic cancer cell lines. Life Sci. 2000, 66, 379–387. [Google Scholar] [CrossRef]
- Gruppetta, M.; Formosa, R.; Falzon, S.; Ariff Scicluna, S.; Falzon, E.; Degeatano, J.; Vassallo, J. Expression of cell cycle regulators and biomarkers of proliferation and regrowth in human pituitary adenomas. Pituitary 2017, 20, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Martinez, A.B.; Bajo, A.M.; Sanchez-Chapado, M.; Prieto, J.C.; Carmena, M.J. Vasoactive intestinal peptide behaves as a pro-metastatic factor in human prostate cancer cells. Prostate 2009, 69, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Mammi, C.; Frajese, G.V.; Vespasiani, G.; Mariani, S.; Gnessi, L.; Farini, D.; Fabbri, A.; Frajese, G.; Moretti, C. PAC1-R null isoform expression in human prostate cancer tissue. Prostate 2006, 66, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Vacas, E.; Fernandez-Martinez, A.B.; Bajo, A.M.; Sanchez-Chapado, M.; Schally, A.V.; Prieto, J.C.; Carmena, M.J. Vasoactive intestinal peptide (VIP) inhibits human renal cell carcinoma proliferation. Biochim. Biophys. Acta 2012, 1823, 1676–1685. [Google Scholar] [CrossRef] [Green Version]
- Wojcieszak, J.; Zawilska, J.B. PACAP38 and PACAP6-38 exert cytotoxic activity against human retinoblastoma Y79 cells. J. Mol. Neurosci. 2014, 54, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Denes, V.; Hideg, O.; Nyisztor, Z.; Lakk, M.; Godri, Z.; Berta, G.; Geck, P.; Gabriel, R. The Neuroprotective Peptide PACAP1-38 Contributes to Horizontal Cell Development in Postnatal Rat Retina. Invest. Ophthalmol. Vis. Sci. 2019, 60, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Green, B.D.; Irwin, N.; Cassidy, R.S.; Gault, V.A.; Flatt, P.R. Long-term administration of PACAP receptor antagonist, PACAP(6-27), impairs glucose tolerance and insulin sensitivity in obese diabetic ob/ob mice. Peptides 2006, 27, 2343–2349. [Google Scholar] [CrossRef]
- Nakata, M.; Shintani, N.; Hashimoto, H.; Baba, A.; Yada, T. Intra-islet PACAP protects pancreatic beta-cells against glucotoxicity and lipotoxicity. J. Mol. Neurosci. 2010, 42, 404–410. [Google Scholar] [CrossRef]
- Sanlioglu, A.D.; Karacay, B.; Balci, M.K.; Griffith, T.S.; Sanlioglu, S. Therapeutic potential of VIP vs PACAP in diabetes. J. Mol. Endocrinol. 2012, 49, R157–R167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Hashimoto, H.; Tomimoto, S.; Shintani, N.; Miyazaki, J.; Tashiro, F.; Aihara, H.; Nammo, T.; Li, M.; Yamagata, K.; et al. Overexpression of PACAP in transgenic mouse pancreatic beta-cells enhances insulin secretion and ameliorates streptozotocin-induced diabetes. Diabetes 2003, 52, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Wang, J.; Li, J.; Wang, Y.; Zhang, H.; Chen, J.; Huang, L.; Liu, X. A novel cyclopeptide from the cyclization of PACAP(1-5) with potent activity towards PAC1 attenuates STZ-induced diabetes. Peptides 2010, 31, 1062–1067. [Google Scholar] [CrossRef]
- Amato, R.; Biagioni, M.; Cammalleri, M.; Dal Monte, M.; Casini, G. VEGF as a Survival Factor in Ex Vivo Models of Early Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 2016, 57, 3066–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amico, A.G.; Maugeri, G.; Rasa, D.M.; Bucolo, C.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. Modulation of IL-1beta and VEGF expression in rat diabetic retinopathy after PACAP administration. Peptides 2017, 97, 64–69. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Reitano, R.; Bucolo, C.; Saccone, S.; Drago, F.; D’Agata, V. PACAP Modulates Expression of Hypoxia-Inducible Factors in Streptozotocin-Induced Diabetic Rat Retina. J. Mol. Neurosci. 2015, 57, 501–509. [Google Scholar] [CrossRef]
- Giunta, S.; Castorina, A.; Bucolo, C.; Magro, G.; Drago, F.; D’Agata, V. Early changes in pituitary adenylate cyclase-activating peptide, vasoactive intestinal peptide and related receptors expression in retina of streptozotocin-induced diabetic rats. Peptides 2012, 37, 32–39. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP Inhibit HIF-1alpha-Mediated VEGF Expression in a Model of Diabetic Macular Edema. J. Cell Physiol. 2017, 232, 1209–1215. [Google Scholar] [CrossRef]
- Szabadfi, K.; Atlasz, T.; Kiss, P.; Reglodi, D.; Szabo, A.; Kovacs, K.; Szalontai, B.; Setalo, G., Jr.; Banki, E.; Csanaky, K.; et al. Protective effects of the neuropeptide PACAP in diabetic retinopathy. Cell Tissue Res. 2012, 348, 37–46. [Google Scholar] [CrossRef]
- Banki, E.; Degrell, P.; Kiss, P.; Kovacs, K.; Kemeny, A.; Csanaky, K.; Duh, A.; Nagy, D.; Toth, G.; Tamas, A.; et al. Effect of PACAP treatment on kidney morphology and cytokine expression in rat diabetic nephropathy. Peptides 2013, 42, 125–130. [Google Scholar] [CrossRef]
- Lauenstein, H.D.; Quarcoo, D.; Plappert, L.; Schleh, C.; Nassimi, M.; Pilzner, C.; Rochlitzer, S.; Brabet, P.; Welte, T.; Hoymann, H.G.; et al. Pituitary adenylate cyclase-activating peptide receptor 1 mediates anti-inflammatory effects in allergic airway inflammation in mice. Clin. Exp. Allergy 2011, 41, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Rasbach, E.; Splitthoff, P.; Bonaterra, G.A.; Schwarz, A.; Mey, L.; Schwarzbach, H.; Eiden, L.E.; Weihe, E.; Kinscherf, R. PACAP deficiency aggravates atherosclerosis in ApoE deficient mice. Immunobiology 2019, 224, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Vizzard, M.A. Up-regulation of pituitary adenylate cyclase-activating polypeptide in urinary bladder pathways after chronic cystitis. J. Comp. Neurol. 2000, 420, 335–348. [Google Scholar] [CrossRef]
- Girard, B.M.; Tooke, K.; Vizzard, M.A. PACAP/Receptor System in Urinary Bladder Dysfunction and Pelvic Pain Following Urinary Bladder Inflammation or Stress. Front. Syst. Neurosci. 2017, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Elekes, K.; Sandor, K.; Moricz, A.; Kereskai, L.; Kemeny, A.; Szoke, E.; Perkecz, A.; Reglodi, D.; Hashimoto, H.; Pinter, E.; et al. Pituitary adenylate cyclase-activating polypeptide plays an anti-inflammatory role in endotoxin-induced airway inflammation: In vivo study with gene-deleted mice. Peptides 2011, 32, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Escher, U.; Grunau, A.; Kuhl, A.A.; Dunay, I.R.; Tamas, A.; Reglodi, D.; Heimesaat, M.M. Pituitary Adenylate Cyclase-Activating Polypeptide-A Neuropeptide as Novel Treatment Option for Subacute Ileitis in Mice Harboring a Human Gut Microbiota. Front. Immunol. 2019, 10, 554. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Dunay, I.R.; Schulze, S.; Fischer, A.; Grundmann, U.; Alutis, M.; Kuhl, A.A.; Tamas, A.; Toth, G.; Dunay, M.P.; et al. Pituitary adenylate cyclase-activating polypeptide ameliorates experimental acute ileitis and extra-intestinal sequelae. PLoS ONE 2014, 9, e108389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Waldeck, K.; Grundemar, L.; Hakanson, R. Ocular inflammation induced by electroconvulsive treatment: Contribution of nitric oxide and neuropeptides mobilized from C-fibres. Br. J. Pharmacol. 1997, 120, 1491–1496. [Google Scholar] [CrossRef]
- Grassel, S.; Muschter, D. Do Neuroendocrine Peptides and Their Receptors Qualify as Novel Therapeutic Targets in Osteoarthritis? Int. J. Mol. Sci. 2018, 19, 367. [Google Scholar] [CrossRef]
- Giunta, S.; Castorina, A.; Marzagalli, R.; Szychlinska, M.A.; Pichler, K.; Mobasheri, A.; Musumeci, G. Ameliorative effects of PACAP against cartilage degeneration. Morphological, immunohistochemical and biochemical evidence from in vivo and in vitro models of rat osteoarthritis. Int. J. Mol. Sci. 2015, 16, 5922–5944. [Google Scholar] [CrossRef]
- Chen, Y.D.; Zhou, Z.G.; Wang, Z.; Gao, H.K.; Yan, W.W.; Wang, C.; Zhao, G.P.; Peng, X.H. Pituitary adenylate cyclase activating-peptide and its receptor antagonists in development of acute pancreatitis in rats. World J. Gastroenterol. 2005, 11, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Hamagami, K.; Sakurai, Y.; Shintani, N.; Higuchi, N.; Ikeda, K.; Hashimoto, H.; Suzuki, A.; Kiyama, H.; Baba, A. Over-expression of pancreatic pituitary adenylate cyclase-activating polypeptide (PACAP) aggravates cerulein-induced acute pancreatitis in mice. J. Pharmacol. Sci. 2009, 110, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, Y.; Shintani, N.; Arimori, A.; Hamagami, K.; Higuchi, N.; Inoue, H.; Ikeda, K.; Hayata, A.; Hashimoto, H.; Baba, A. Cerulein-induced acute pancreatitis in PACAP knockout mice. J. Mol. Neurosci. 2011, 43, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Di Nardo, A. Skin neurogenic inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Helyes, Z.; Kun, J.; Dobrosi, N.; Sandor, K.; Nemeth, J.; Perkecz, A.; Pinter, E.; Szabadfi, K.; Gaszner, B.; Tekus, V.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide Is Upregulated in Murine Skin Inflammation and Mediates Transient Receptor Potential Vanilloid-1-Induced Neurogenic Edema. J. Invest. Dermatol. 2015, 135, 2209–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, J.B.; Larkin, S.W.; Coughlan, M.; Kajekar, R.; Williams, T.J. Pituitary adenylate cyclase activating polypeptide is a potent vasodilator and oedema potentiator in rabbit skin in vivo. Br. J. Pharmacol. 1992, 106, 331–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, C.A.; Dusedau, H.P.; Steffen, J.; Gupta, N.; Dunay, M.P.; Toth, G.K.; Reglodi, D.; Heimesaat, M.M.; Dunay, I.R. Immunomodulatory Effects of the Neuropeptide Pituitary Adenylate Cyclase-Activating Polypeptide in Acute Toxoplasmosis. Front. Cell Infect. Microbiol. 2019, 9, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, T.M.; Temerozo, J.R.; Giestal-de-Araujo, E.; Bou-Habib, D.C. The effects of neurotrophins and the neuropeptides VIP and PACAP on HIV-1 infection: Histories with opposite ends. Neuroimmunomodulation 2014, 21, 268–282. [Google Scholar] [CrossRef]
- Temerozo, J.R.; de Azevedo, S.S.D.; Insuela, D.B.R.; Vieira, R.C.; Ferreira, P.L.C.; Carvalho, V.F.; Bello, G.; Bou-Habib, D.C. The Neuropeptides Vasoactive Intestinal Peptide and Pituitary Adenylate Cyclase-Activating Polypeptide Control HIV-1 Infection in Macrophages Through Activation of Protein Kinases A and C. Front. Immunol. 2018, 9, 1336. [Google Scholar] [CrossRef] [Green Version]
- Temerozo, J.R.; Joaquim, R.; Regis, E.G.; Savino, W.; Bou-Habib, D.C. Macrophage Resistance to HIV-1 Infection is Enhanced by the Neuropeptides VIP and PACAP. PLoS ONE 2013, 8, e67701. [Google Scholar] [CrossRef]
- Han, P.; Caselli, R.J.; Baxter, L.; Serrano, G.; Yin, J.; Beach, T.G.; Reiman, E.M.; Shi, J. Association of pituitary adenylate cyclase-activating polypeptide with cognitive decline in mild cognitive impairment due to Alzheimer disease. JAMA Neurol 2015, 72, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Liang, W.; Baxter, L.C.; Yin, J.; Tang, Z.; Beach, T.G.; Caselli, R.J.; Reiman, E.M.; Shi, J. Pituitary adenylate cyclase-activating polypeptide is reduced in Alzheimer disease. Neurology 2014, 82, 1724–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rat, D.; Schmitt, U.; Tippmann, F.; Dewachter, I.; Theunis, C.; Wieczerzak, E.; Postina, R.; van Leuven, F.; Fahrenholz, F.; Kojro, E. Neuropeptide pituitary adenylate cyclase-activating polypeptide (PACAP) slows down Alzheimer’s disease-like pathology in amyloid precursor protein-transgenic mice. FASEB J. 2011, 25, 3208–3218. [Google Scholar] [CrossRef] [PubMed]
- Lohoff, F.W.; Bloch, P.J.; Weller, A.E.; Ferraro, T.N.; Berrettini, W.H. Association analysis of the pituitary adenylate cyclase-activating polypeptide (PACAP/ADCYAP1) gene in bipolar disorder. Psychiatr Genet. 2008, 18, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Hattori, T.; Yamada, K.; Matsuzaki, S.; Tohyama, M. Role of the PACAP-PAC1-DISC1 and PACAP-PAC1-stathmin1 systems in schizophrenia and bipolar disorder: Novel treatment mechanisms? Pharmacogenomics 2009, 10, 1967–1978. [Google Scholar] [CrossRef] [PubMed]
- Bhandare, A.M.; Kapoor, K.; Powell, K.L.; Braine, E.; Casillas-Espinosa, P.; O’Brien, T.J.; Farnham, M.M.J.; Pilowsky, P.M. Inhibition of microglial activation with minocycline at the intrathecal level attenuates sympathoexcitatory and proarrhythmogenic changes in rats with chronic temporal lobe epilepsy. Neuroscience 2017, 350, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Reis, D.; Mde, F.A.; Ribeiro, J.A. Endogenous inhibition of hippocampal LTD and depotentiation by vasoactive intestinal peptide VPAC1 receptors. Hippocampus 2014, 24, 1353–1363. [Google Scholar] [CrossRef]
- Cabezas-Llobet, N.; Vidal-Sancho, L.; Masana, M.; Fournier, A.; Alberch, J.; Vaudry, D.; Xifro, X. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) Enhances Hippocampal Synaptic Plasticity and Improves Memory Performance in Huntington’s Disease. Mol. Neurobiol. 2018, 55, 8263–8277. [Google Scholar] [CrossRef]
- Brifault, C.; Gras, M.; Liot, D.; May, V.; Vaudry, D.; Wurtz, O. Delayed pituitary adenylate cyclase-activating polypeptide delivery after brain stroke improves functional recovery by inducing m2 microglia/macrophage polarization. Stroke 2015, 46, 520–528. [Google Scholar] [CrossRef]
- Danyadi, B.; Szabadfi, K.; Reglodi, D.; Mihalik, A.; Danyadi, T.; Kovacs, Z.; Batai, I.; Tamas, A.; Kiss, P.; Toth, G.; et al. PACAP application improves functional outcome of chronic retinal ischemic injury in rats-evidence from electroretinographic measurements. J. Mol. Neurosci. 2014, 54, 293–299. [Google Scholar] [CrossRef]
- Lazarovici, P.; Cohen, G.; Arien-Zakay, H.; Chen, J.; Zhang, C.; Chopp, M.; Jiang, H. Multimodal neuroprotection induced by PACAP38 in oxygen-glucose deprivation and middle cerebral artery occlusion stroke models. J. Mol. Neurosci. 2012, 48, 526–540. [Google Scholar] [CrossRef] [PubMed]
- Lenti, L.; Zimmermann, A.; Kis, D.; Olah, O.; Toth, G.K.; Hegyi, O.; Busija, D.W.; Bari, F.; Domoki, F. PACAP and VIP differentially preserve neurovascular reactivity after global cerebral ischemia in newborn pigs. Brain Res. 2009, 1283, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Lu, T.; Zhang, C.; Xue, Z.; Xu, J.; Busuttil, R.W.; Xia, Q.; Xu, N.; Kupiec-Weglinski, J.W.; Ji, H. Pituitary Adenylate Cyclase-activating Polypeptides Prevent Hepatocyte Damage by Promoting Yes-associated Protein in Liver Ischemia-Reperfusion Injury. Transplantation 2019, 103, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Nakamachi, T.; Watanabe, J.; Sugiyama, K.; Ohtaki, H.; Murai, N.; Sasaki, S.; Xu, Z.; Hashimoto, H.; Seki, T.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) is Involved in Adult Mouse Hippocampal Neurogenesis after Stroke. J. Mol. Neurosci. 2016, 59, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Tsuchida, M.; Kagami, N.; Yofu, S.; Wada, Y.; Hori, M.; Tsuchikawa, D.; Yoshikawa, A.; Imai, N.; Nakamura, K.; et al. IL-6 and PACAP receptor expression and localization after global brain ischemia in mice. J. Mol. Neurosci. 2012, 48, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Riek-Burchardt, M.; Kolodziej, A.; Henrich-Noack, P.; Reymann, K.G.; Hollt, V.; Stumm, R. Differential regulation of CXCL12 and PACAP mRNA expression after focal and global ischemia. Neuropharmacology 2010, 58, 199–207. [Google Scholar] [CrossRef]
- Stetler, R.A.; Zhang, F.; Liu, C.; Chen, J. Ischemic tolerance as an active and intrinsic neuroprotective mechanism. Handb. Clin. Neurol 2009, 92, 171–195. [Google Scholar] [PubMed]
- Werling, D.; Banks, W.A.; Salameh, T.S.; Kvarik, T.; Kovacs, L.A.; Vaczy, A.; Szabo, E.; Mayer, F.; Varga, R.; Tamas, A.; et al. Passage through the Ocular Barriers and Beneficial Effects in Retinal Ischemia of Topical Application of PACAP1-38 in Rodents. Int. J. Mol. Sci. 2017, 18, 675. [Google Scholar] [CrossRef]
- Akerman, S.; Goadsby, P.J. Neuronal PAC1 receptors mediate delayed activation and sensitization of trigeminocervical neurons: Relevance to migraine. Sci. Transl. Med. 2015, 7, 308ra157. [Google Scholar] [CrossRef]
- Amin, F.M.; Asghar, M.S.; Guo, S.; Hougaard, A.; Hansen, A.E.; Schytz, H.W.; van der Geest, R.J.; de Koning, P.J.; Larsson, H.B.; Olesen, J.; et al. Headache and prolonged dilatation of the middle meningeal artery by PACAP38 in healthy volunteers. Cephalalgia 2012, 32, 140–149. [Google Scholar] [CrossRef]
- Birk, S.; Sitarz, J.T.; Petersen, K.A.; Oturai, P.S.; Kruuse, C.; Fahrenkrug, J.; Olesen, J. The effect of intravenous PACAP38 on cerebral hemodynamics in healthy volunteers. Regul. Pept. 2007, 140, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Schytz, H.W.; Birk, S.; Wienecke, T.; Kruuse, C.; Olesen, J.; Ashina, M. PACAP38 induces migraine-like attacks in patients with migraine without aura. Brain 2009, 132, 16–25. [Google Scholar] [CrossRef]
- Amin, F.M.; Hougaard, A.; Schytz, H.W.; Asghar, M.S.; Lundholm, E.; Parvaiz, A.I.; de Koning, P.J.; Andersen, M.R.; Larsson, H.B.; Fahrenkrug, J.; et al. Investigation of the pathophysiological mechanisms of migraine attacks induced by pituitary adenylate cyclase-activating polypeptide-38. Brain 2014, 137, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Csati, A.; Tajti, J.; Kuris, A.; Tuka, B.; Edvinsson, L.; Warfvinge, K. Distribution of Vasoactive Intestinal Peptide, Pituitary Adenylate Cyclase-Activating Peptide, Nitric Oxide Synthase, and Their Receptors in Human and Rat Sphenopalatine Ganglion. Neuroscience 2012, 202, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Joo, K.M.; Chung, Y.H.; Kim, M.K.; Nam, R.H.; Lee, B.L.; Lee, K.H.; Cha, C.I. Distribution of vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide receptors (VPAC(1), VPAC(2), and PAC(1) receptor) in the rat brain. J. Compar. Neurol. 2004, 476, 388–413. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, D.H.; Fordyce, A.; Walker, M.T.; St Clair, D.M.; Porteous, D.J.; Muir, W.J. Schizophrenia and affective disorders--cosegregation with a translocation at chromosome 1q42 that directly disrupts brain-expressed genes: Clinical and P300 findings in a family. Am. J. Hum. Genet. 2001, 69, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Hashimoto, H.; Shintani, N.; Ohi, K.; Hori, H.; Saitoh, O.; Kosuga, A.; Tatsumi, M.; Iwata, N.; Ozaki, N.; et al. Possible association between the pituitary adenylate cyclase-activating polypeptide (PACAP) gene and major depressive disorder. Neurosci. Lett. 2010, 468, 300–302. [Google Scholar] [CrossRef]
- Pinhasov, A.; Nesher, E.; Gross, M.; Turgeman, G.; Kreinin, A.; Yadid, G. The role of the PACAP signaling system in depression. Curr. Pharm. Des. 2011, 17, 990–1001. [Google Scholar] [CrossRef]
- Tohyama, M.; Miyata, S.; Hattori, T.; Shimizu, S.; Matsuzaki, S. Molecular basis of major psychiatric diseases such as schizophrenia and depression. Anat. Sci. Int. 2015, 90, 137–143. [Google Scholar] [CrossRef]
- Abad, C.; Jayaram, B.; Becquet, L.; Wang, Y.; O’Dorisio, M.S.; Waschek, J.A.; Tan, Y.V. VPAC1 receptor (Vipr1)-deficient mice exhibit ameliorated experimental autoimmune encephalomyelitis, with specific deficits in the effector stage. J. Neuroinflamm. 2016, 13, 169. [Google Scholar] [CrossRef] [Green Version]
- Abad, C.; Waschek, J.A. Immunomodulatory roles of VIP and PACAP in models of multiple sclerosis. Curr. Pharm. Des. 2011, 17, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Baranowska-Bik, A.; Kochanowski, J.; Uchman, D.; Wolinska-Witort, E.; Kalisz, M.; Martynska, L.; Baranowska, B.; Bik, W. Vasoactive intestinal peptide (VIP) and pituitary adenylate cyclase activating polypeptide (PACAP) in humans with multiple sclerosis. J. Neuroimmunol. 2013, 263, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, G.; Iemmolo, R.; D’Amico, A.G.; La Cognata, V.; Costanzo, E.; Zappia, M.; D’Agata, V.; Conforti, F.L.; Aronica, E.; Cavallaro, S. PACAP and PAC1R are differentially expressed in motor cortex of amyotrophic lateral sclerosis patients and support survival of iPSC-derived motor neurons. J. Cell Physiol. 2018, 233, 3343–3351. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Ito, A.; Kawanokuchi, J.; Jin, S.; Mizuno, T.; Ojika, K.; Ueda, R.; Suzumura, A. Pituitary adenylate cyclase-activating polypeptide (PACAP) ameliorates experimental autoimmune encephalomyelitis by suppressing the functions of antigen presenting cells. Mult. Scler 2004, 10, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Rasa, D.M.; Federico, C.; Saccone, S.; Morello, G.; La Cognata, V.; Cavallaro, S.; D’Agata, V. Molecular mechanisms involved in the protective effect of pituitary adenylate cyclase-activating polypeptide in an in vitro model of amyotrophic lateral sclerosis. J. Cell Physiol. 2019, 234, 5203–5214. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.V.; Abad, C.; Lopez, R.; Dong, H.; Liu, S.; Lee, A.; Gomariz, R.P.; Leceta, J.; Waschek, J.A. Pituitary adenylyl cyclase-activating polypeptide is an intrinsic regulator of Treg abundance and protects against experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2009, 106, 2012–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.V.; Waschek, J.A. Targeting VIP and PACAP receptor signalling: New therapeutic strategies in multiple sclerosis. ASN Neuro 2011, 3, AN20110024. [Google Scholar] [CrossRef] [PubMed]
- Van, C.; Condro, M.C.; Lov, K.; Zhu, R.; Ricaflanca, P.T.; Ko, H.H.; Diep, A.L.; Hoang, A.Q.; Pisegna, J.; Rohrer, H.; et al. Correction to: PACAP/PAC1 Regulation of Inflammation via Catecholaminergic Neurons in a Model of Multiple Sclerosis. J. Mol. Neurosci. 2019, 68, 452. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Tamas, A.; Reglodi, D.; Tizabi, Y. PACAP protects against inflammatory-mediated toxicity in dopaminergic SH-SY5Y cells: Implication for Parkinson’s disease. Neurotox Res. 2014, 26, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Lamine, A.; Letourneau, M.; Doan, N.D.; Maucotel, J.; Couvineau, A.; Vaudry, H.; Chatenet, D.; Vaudry, D.; Fournier, A. Characterizations of a synthetic pituitary adenylate cyclase-activating polypeptide analog displaying potent neuroprotective activity and reduced in vivo cardiovascular side effects in a Parkinson’s disease model. Neuropharmacology 2016, 108, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Shivers, K.Y.; Nikolopoulou, A.; Machlovi, S.I.; Vallabhajosula, S.; Figueiredo-Pereira, M.E. PACAP27 prevents Parkinson-like neuronal loss and motor deficits but not microglia activation induced by prostaglandin J2. Biochim. Biophys. Acta 2014, 1842, 1707–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Pan, J.; Tan, Y.Y.; Sun, X.K.; Zhang, Y.F.; Zhou, H.Y.; Ren, R.J.; Wang, X.J.; Chen, S.D. Neuroprotective effects of PACAP27 in mice model of Parkinson’s disease involved in the modulation of K(ATP) subunits and D2 receptors in the striatum. Neuropeptides 2008, 42, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Xie, P.; Anton, R.F.; De Vivo, I.; Farrer, L.A.; Kranzler, H.R.; Oslin, D.; Purcell, S.M.; Roberts, A.L.; Smoller, J.W.; et al. No association between ADCYAP1R1 and post-traumatic stress disorder in two independent samples. Mol. Psychiatry 2012, 17, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Dias, B.G.; Ressler, K.J. PACAP and the PAC1 receptor in post-traumatic stress disorder. Neuropsychopharmacology 2013, 38, 245–246. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Hashimoto, H.; Shintani, N.; Chiba, S.; Hattori, S.; Okada, T.; Nakajima, M.; Tanaka, K.; Kawagishi, N.; Nemoto, K.; et al. Pituitary adenylate cyclase-activating polypeptide is associated with schizophrenia. Mol. Psychiatry 2007, 12, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, M.; Ishiguro, H.; Horiuchi, Y.; Inada, T.; Ujike, H.; Itokawa, M.; Otowa, T.; Watanabe, Y.; Someya, T.; Arinami, T. Replication study of association between ADCYAP1 gene polymorphisms and schizophrenia. Psychiatr. Genet. 2010, 20, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Hua, R.; Mao, S.S.; Zhang, Y.M.; Chen, F.X.; Zhou, Z.H.; Liu, J.Q. Effects of pituitary adenylate cyclase activating polypeptide on CD4(+)/CD8(+) T cell levels after traumatic brain injury in a rat model. World J. Emerg. Med. 2012, 3, 294–298. [Google Scholar] [CrossRef]
- Miyamoto, K.; Tsumuraya, T.; Ohtaki, H.; Dohi, K.; Satoh, K.; Xu, Z.; Tanaka, S.; Murai, N.; Watanabe, J.; Sugiyama, K.; et al. PACAP38 suppresses cortical damage in mice with traumatic brain injury by enhancing antioxidant activity. J. Mol. Neurosci. 2014, 54, 370–379. [Google Scholar] [CrossRef]
- Fang, K.M.; Chen, J.K.; Hung, S.C.; Chen, M.C.; Wu, Y.T.; Wu, T.J.; Lin, H.I.; Chen, C.H.; Cheng, H.; Yang, C.S.; et al. Effects of combinatorial treatment with pituitary adenylate cyclase activating peptide and human mesenchymal stem cells on spinal cord tissue repair. PLoS ONE 2010, 5, e15299. [Google Scholar] [CrossRef]
- Tsuchida, M.; Nakamachi, T.; Sugiyama, K.; Tsuchikawa, D.; Watanabe, J.; Hori, M.; Yoshikawa, A.; Imai, N.; Kagami, N.; Matkovits, A.; et al. PACAP stimulates functional recovery after spinal cord injury through axonal regeneration. J. Mol. Neurosci. 2014, 54, 380–387. [Google Scholar] [CrossRef]
- Lemaire, P. Evolutionary crossroads in developmental biology: The tunicates. Development 2011, 138, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, W.R. Chordate ancestry of the neural crest: New insights from ascidians. Semin Cell Dev. Biol. 2007, 18, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Ohtaki, H.; Seki, T.; Yofu, S.; Kagami, N.; Hashimoto, H.; Shintani, N.; Baba, A.; Mark, L.; Lanekoff, I.; et al. PACAP suppresses dry eye signs by stimulating tear secretion. Nat. Commun. 2016, 7, 12034. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.C.; Felix, R.C.; Martins, R.S.; Trindade, M.; Fonseca, V.G.; Fuentes, J.; Power, D.M. PACAP system evolution and its role in melanophore function in teleost fish skin. Mol. Cell Endocrinol. 2015, 411, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Zhou, R.; DiCicco-Bloom, E. Opposing mitogenic regulation by PACAP in sympathetic and cerebral cortical precursors correlates with differential expression of PACAP receptor (PAC1-R) isoforms. J. Neurosci. Res. 1998, 53, 651–662. [Google Scholar] [CrossRef]
- Robberecht, P.; Woussen-Colle, M.C.; De Neef, P.; Gourlet, P.; Buscail, L.; Vandermeers, A.; Vandermeers-Piret, M.C.; Christophe, J. The two forms of the pituitary adenylate cyclase activating polypeptide (PACAP (1-27) and PACAP (1-38)) interact with distinct receptors on rat pancreatic AR 4-2J cell membranes. FEBS Lett. 1991, 286, 133–136. [Google Scholar] [CrossRef] [Green Version]
- IUPHAR/BPS Guide to Pharmacology. Available online: https://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=67 (accessed on 16 September 2019).
- Hirabayashi, T.; Nakamachi, T.; Shioda, S. Discovery of PACAP and its receptors in the brain. J. Headache Pain 2018, 19, 28. [Google Scholar] [CrossRef]
- Felley, C.P.; Qian, J.M.; Mantey, S.; Pradhan, T.; Jensen, R.T. Chief cells possess a receptor with high affinity for PACAP and VIP that stimulates pepsinogen release. Am. J. Physiol. 1992, 263, G901–G907. [Google Scholar] [CrossRef] [PubMed]
- Harmar, A.J.; Fahrenkrug, J.; Gozes, I.; Laburthe, M.; May, V.; Pisegna, J.R.; Vaudry, D.; Vaudry, H.; Waschek, J.A.; Said, S.I. Pharmacology and functions of receptors for vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide: IUPHAR review 1. Br. J. Pharmacol. 2012, 166, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Pisegna, J.R.; Wank, S.A. Cloning and characterization of the signal transduction of four splice variants of the human pituitary adenylate cyclase activating polypeptide receptor. Evidence for dual coupling to adenylate cyclase and phospholipase C. J. Biol. Chem. 1996, 271, 17267–17274. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, T.K.; Sharma, R.V.; Fisher, R.A. Molecular cloning of a novel variant of the pituitary adenylate cyclase-activating polypeptide (PACAP) receptor that stimulates calcium influx by activation of L-type calcium channels. J. Biol. Chem. 1996, 271, 32226–32232. [Google Scholar] [CrossRef] [PubMed]
- Merriam, L.A.; Baran, C.N.; Girard, B.M.; Hardwick, J.C.; May, V.; Parsons, R.L. Pituitary adenylate cyclase 1 receptor internalization and endosomal signaling mediate the pituitary adenylate cyclase activating polypeptide-induced increase in guinea pig cardiac neuron excitability. J. Neurosci. 2013, 33, 4614–4622. [Google Scholar] [CrossRef] [PubMed]
- May, V.; Buttolph, T.R.; Girard, B.M.; Clason, T.A.; Parsons, R.L. PACAP-induced ERK activation in HEK cells expressing PAC1 receptors involves both receptor internalization and PKC signaling. Am. J. Physiol. Cell Physiol. 2014, 306, C1068–C1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopoulos, A.; Christopoulos, G.; Morfis, M.; Udawela, M.; Laburthe, M.; Couvineau, A.; Kuwasako, K.; Tilakaratne, N.; Sexton, P.M. Novel receptor partners and function of receptor activity-modifying proteins. J. Biol. Chem. 2003, 278, 3293–3297. [Google Scholar] [CrossRef] [PubMed]
- Dickson, L.; Finlayson, K. VPAC and PAC receptors: From ligands to function. Pharmacol. Ther. 2009, 121, 294–316. [Google Scholar] [CrossRef] [PubMed]
- Moody, T.W.; Ito, T.; Osefo, N.; Jensen, R.T. VIP and PACAP: Recent insights into their functions/roles in physiology and disease from molecular and genetic studies. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Puledda, F.; Messina, R.; Goadsby, P.J. An update on migraine: Current understanding and future directions. J. Neurol 2017, 264, 2031–2039. [Google Scholar] [CrossRef]
- Baun, M.; Pedersen, M.H.; Olesen, J.; Jansen-Olesen, I. Dural mast cell degranulation is a putative mechanism for headache induced by PACAP-38. Cephalalgia 2012, 32, 337–345. [Google Scholar] [CrossRef]
- Carlton, S.M. Nociceptive primary afferents: They have a mind of their own. J. Physiol. 2014, 592, 3403–3411. [Google Scholar] [CrossRef]
- Luger, T.A. Neuromediators—A crucial component of the skin immune system. J. Dermatol. Sci. 2002, 30, 87–93. [Google Scholar] [CrossRef]
- Jansen-Olesen, I.; Pedersen, S.H. PACAP and its receptors in cranial arteries and mast cells. J. Headache Pain 2018, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Minkes, R.K.; McMahon, T.J.; Hood, J.S.; Murphy, W.A.; Coy, D.H.; McNamara, D.B.; Kadowitz, P.J. Differential effects of PACAP and VIP on the pulmonary and hindquarters vascular beds of the cat. J. Appl. Physiol. 1992, 72, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Parfenova, H.; Shibata, M.; Zuckerman, S.; Armstead, W.M.; Leffler, C.W. Pituitary adenylate cyclase-activating polypeptide dilates cerebral arterioles of newborn pigs. Proc. Soc. Exp. Biol. Med. 1993, 203, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Ascuitto, R.J.; RossAscuitto, N.T.; Waddell, A.E.; Kadowitz, P.J. Contractile and coronary vascular effects of pituitary adenylate cyclase activating polypeptide in neonatal pig hearts. Cardiovascular Res. 1996, 31, E153–E159. [Google Scholar] [CrossRef]
- Edvinsson, L.; Elsas, T.; Suzuki, N.; Shimizu, T.; Lee, T.J. Origin and Co-localization of nitric oxide synthase, CGRP, PACAP, and VIP in the cerebral circulation of the rat. Microsc Res. Tech. 2001, 53, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Knutsson, M.; Edvinsson, L. Distribution of mRNA for VIP and PACAP receptors in human cerebral arteries and cranial ganglia. Neuroreport 2002, 13, 507–509. [Google Scholar] [CrossRef]
- Boni, L.J.; Ploug, K.B.; Olesen, J.; Jansen-Olesen, I.; Gupta, S. The in vivo effect of VIP, PACAP-38 and PACAP-27 and mRNA expression of their receptors in rat middle meningeal artery. Cephalalgia 2009, 29, 837–847. [Google Scholar] [CrossRef]
- Amin, F.M.; Schytz, H.W. Transport of the pituitary adenylate cyclase-activating polypeptide across the blood-brain barrier: Implications for migraine. J. Headache Pain 2018, 19, 35. [Google Scholar] [CrossRef]
- Chaudhary, P.; Baumann, T.K. Expression of VPAC2 receptor and PAC1 receptor splice variants in the trigeminal ganglion of the adult rat. Brain Res. Mol. Brain Res. 2002, 104, 137–142. [Google Scholar] [CrossRef]
- Hensley, K.; Pretorius, J.; Chan, B.; Page, K.; Liu, H.; Choi, C.; Shi, D.; Xu, C.; Edvinsson, L.; Miller, S. PAC1 receptor mRNA and protein distribution in rat and human trigeminal and sphenopalatine ganglia, spinal trigeminal nucleus and in dura mater. Cephalalgia 2019, 39, 827–840. [Google Scholar] [CrossRef]
- Okragly, A.J.; Morin, S.M.; DeRosa, D.; Martin, A.P.; Johnson, K.W.; Johnson, M.P.; Benschop, R.J. Human mast cells release the migraine-inducing factor pituitary adenylate cyclase-activating polypeptide (PACAP). Cephalalgia 2018, 38, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; D’Ovidio, C.; Conti, C.; Gallenga, C.E.; Lauritano, D.; Caraffa, A.; Kritas, S.K.; Ronconi, G. Progression in migraine: Role of mast cells and pro-inflammatory and anti-inflammatory cytokines. Eur. J. Pharmacol. 2019, 844, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Pozo, D.; Martinez, C.; Garrido, E.; Leceta, J.; Calvo, J.R.; Gomariz, R.P. Characterization of gene expression of VIP and VIP1-receptor in rat peritoneal lymphocytes and macrophages. Regul. Pept. 1996, 62, 161–166. [Google Scholar] [CrossRef]
- Schmidt-Choudhury, A.; Furuta, G.T.; Galli, S.J.; Schmidt, W.E.; Wershil, B.K. Mast cells contribute to PACAP-induced dermal oedema in mice. Regul. Pept. 1999, 82, 65–69. [Google Scholar] [CrossRef]
- Bhatt, D.K.; Gupta, S.; Olesen, J.; Jansen-Olesen, I. PACAP-38 infusion causes sustained vasodilation of the middle meningeal artery in the rat: Possible involvement of mast cells. Cephalalgia 2014, 34, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Robberecht, P.; Gourlet, P.; De Neef, P.; Woussen-Colle, M.C.; Vandermeers-Piret, M.C.; Vandermeers, A.; Christophe, J. Structural requirements for the occupancy of pituitary adenylate-cyclase-activating-peptide (PACAP) receptors and adenylate cyclase activation in human neuroblastoma NB-OK-1 cell membranes. Discovery of PACAP(6-38) as a potent antagonist. Eur. J. Biochem. 1992, 207, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R. Neurogenic inflammation and its role in migraine. Semin Immunopathol. 2018, 40, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, S.; Saito, K.; Moskowitz, M.A. Neurogenically mediated leakage of plasma protein occurs from blood vessels in dura mater but not brain. J. Neurosci. 1987, 7, 4129–4136. [Google Scholar] [CrossRef] [Green Version]
- Markowitz, S.; Saito, K.; Moskowitz, M.A. Neurogenically mediated plasma extravasation in dura mater: Effect of ergot alkaloids. A possible mechanism of action in vascular headache. Cephalalgia 1988, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- May, A.; Goadsby, P.J. Substance P receptor antagonists in the therapy of migraine. Expert Opin. Investig. Drugs 2001, 10, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.Y.; Baun, M.; de Vries, R.; van den Bogaerdt, A.J.; Dirven, C.M.; Danser, A.H.; Jansen-Olesen, I.; Olesen, J.; Villalon, C.M.; MaassenVanDenBrink, A.; et al. Pharmacological characterization of VIP and PACAP receptors in the human meningeal and coronary artery. Cephalalgia 2011, 31, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Hautmann, M.; Friis, U.G.; Desch, M.; Todorov, V.; Castrop, H.; Segerer, F.; Otto, C.; Schutz, G.; Schweda, F. Pituitary adenylate cyclase-activating polypeptide stimulates renin secretion via activation of PAC1 receptors. J. Am. Soc. Nephrol. 2007, 18, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, S.; Buddenkotte, J.; Schmidt-Choudhury, A.; Rosignoli, C.; Shpacovitch, V.; von Arnim, U.; Metze, D.; Rukwied, R.; Schmelz, M.; Paus, R.; et al. Pituitary adenylate cyclase activating polypeptide: An important vascular regulator in human skin in vivo. Am. J. Pathol. 2010, 177, 2563–2575. [Google Scholar] [CrossRef] [PubMed]
- Clinical trials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03238781 (accessed on 16 September 2019).
- Eftekhari, S.; Salvatore, C.A.; Johansson, S.; Chen, T.B.; Zeng, Z.; Edvinsson, L. Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood-brain barrier. Brain Res. 2015, 1600, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Shintani, N.; Tanida, M.; Hayata, A.; Hashimoto, R.; Baba, A. PACAP is implicated in the stress axes. Curr. Pharm. Des. 2011, 17, 985–989. [Google Scholar] [CrossRef]
- Przywara, D.A.; Guo, X.; Angelilli, M.L.; Wakade, T.D.; Wakade, A.R. A non-cholinergic transmitter, pituitary adenylate cyclase-activating polypeptide, utilizes a novel mechanism to evoke catecholamine secretion in rat adrenal chromaffin cells. J. Biol. Chem. 1996, 271, 10545–10550. [Google Scholar] [CrossRef]
- Koves, K.; Arimura, A.; Gorcs, T.G.; Somogyvarivigh, A. Comparative Distribution of Immunoreactive Pituitary Adenylate-Cyclase Activating Polypeptide and Vasoactive Intestinal Polypeptide in Rat Forebrain. Neuroendocrinology 1991, 54, 159–169. [Google Scholar] [CrossRef]
- Kozicz, T.; Vigh, S.; Arimura, A. The source of origin of PACAP- and VIP-immunoreactive fibers in the laterodorsal division of the bed nucleus of the stria terminalis in the rat. Brain Res. 1998, 810, 211–219. [Google Scholar] [CrossRef]
- Otto, C.; Zuschratter, W.; Gass, P.; Schutz, G. Presynaptic localization of the PACAP-typeI-receptor in hippocampal and cerebellar mossy fibres. Mol. Brain Res. 1999, 66, 163–174. [Google Scholar] [CrossRef]
- Piggins, H.D.; Stamp, J.A.; Burns, J.; Rusak, B.; Semba, K. Distribution of pituitary adenylate cyclase activating polypeptide (PACAP) immunoreactivity in the hypothalamus and extended amygdala of the rat. J. Comp. Neurol. 1996, 376, 278–294. [Google Scholar] [CrossRef]
- Hammack, S.E.; Cheung, J.; Rhodes, K.M.; Schutz, K.C.; Falls, W.A.; Braas, K.M.; May, V. Chronic stress increases pituitary adenylate cyclase-activating peptide (PACAP) and brain-derived neurotrophic factor (BDNF) mRNA expression in the bed nucleus of the stria terminalis (BNST): Roles for PACAP in anxiety-like behavior. Psychoneuroendocrinology 2009, 34, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammack, S.E.; Roman, C.W.; Lezak, K.R.; Kocho-Shellenberg, M.; Grimmig, B.; Falls, W.A.; Braas, K.; May, V. Roles for pituitary adenylate cyclase-activating peptide (PACAP) expression and signaling in the bed nucleus of the stria terminalis (BNST) in mediating the behavioral consequences of chronic stress. J. Mol. Neurosci. 2010, 42, 327–340. [Google Scholar] [CrossRef]
- Agarwal, A.; Halvorson, L.M.; Legradi, G. Pituitary adenylate cyclase-activating polypeptide (PACAP) mimics neuroendocrine and behavioral manifestations of stress: Evidence for PKA-mediated expression of the corticotropin-releasing hormone (CRH) gene. Brain Res. Mol. Brain Res. 2005, 138, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinevich, V.; Fournier, A.; Pelletier, G. Effects of pituitary adenylate cyclase-activating polypeptide (PACAP) on corticotropin-releasing hormone (CRH) gene expression in the rat hypothalamic paraventricular nucleus. Brain Res. 1997, 773, 190–196. [Google Scholar] [CrossRef]
- Stroth, N.; Eiden, L.E. Stress hormone synthesis in mouse hypothalamus and adrenal gland triggered by restraint is dependent on pituitary adenylate cyclase-activating polypeptide signaling. Neuroscience 2010, 165, 1025–1030. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, H.; Shintani, N.; Tanaka, K.; Mori, W.; Hirose, M.; Matsuda, T.; Sakaue, M.; Miyazaki, J.; Niwa, H.; Tashiro, F.; et al. Altered psychomotor behaviors in mice lacking pituitary adenylate cyclase-activating polypeptide (PACAP). Proc. Natl. Acad. Sci. USA 2001, 98, 13355–13360. [Google Scholar] [CrossRef] [Green Version]
- Gaszner, B.; Kormos, V.; Kozicz, T.; Hashimoto, H.; Reglodi, D.; Helyes, Z. The behavioral phenotype of pituitary adenylate-cyclase activating polypeptide-deficient mice in anxiety and depression tests is accompanied by blunted c-Fos expression in the bed nucleus of the stria terminalis, central projecting Edinger-Westphal nucleus, ventral lateral septum, and dorsal raphe nucleus. Neuroscience 2012, 202, 283–299. [Google Scholar]
- Norrholm, S.D.; Das, M.; Legradi, G. Behavioral effects of local microinfusion of pituitary adenylate cyclase activating polypeptide (PACAP) into the paraventricular nucleus of the hypothalamus (PVN). Regul. Pept. 2005, 128, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.L.; Davis, M. Double dissociation between the involvement of the bed nucleus of the stria terminalis and the central nucleus of the amygdala in startle increases produced by conditioned versus unconditioned fear. J. Neurosci. 1997, 17, 9375–9383. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.L.; Toufexis, D.J.; Davis, M. Role of the bed nucleus of the stria terminalis versus the amygdala in fear, stress, and anxiety. Eur. J. Pharmacol. 2003, 463, 199–216. [Google Scholar] [CrossRef]
- King, S.B.; Toufexis, D.J.; Hammack, S.E. Pituitary adenylate cyclase activating polypeptide (PACAP), stress, and sex hormones. Stress 2017, 20, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Miles, O.W.; Maren, S. Role of the Bed Nucleus of the Stria Terminalis in PTSD: Insights From Preclinical Models. Front. Behav. Neurosci. 2019, 13, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starr, E.R.; Margiotta, J.F. Pituitary adenylate cyclase activating polypeptide induces long-term, transcription-dependent plasticity and remodeling at autonomic synapses. Mol. Cell Neurosci. 2017, 85, 170–182. [Google Scholar] [CrossRef]
- Meloni, E.G.; Kaye, K.T.; Venkataraman, A.; Carlezon, W.A., Jr. PACAP increases Arc/Arg 3.1 expression within the extended amygdala after fear conditioning in rats. Neurobiol. Learn. Mem. 2019, 157, 24–34. [Google Scholar] [CrossRef]
- Kirry, A.J.; Herbst, M.R.; Poirier, S.E.; Maskeri, M.M.; Rothwell, A.C.; Twining, R.C.; Gilmartin, M.R. Pituitary adenylate cyclase-activating polypeptide (PACAP) signaling in the prefrontal cortex modulates cued fear learning, but not spatial working memory, in female rats. Neuropharmacology 2018, 133, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meloni, E.G.; Venkataraman, A.; Donahue, R.J.; Carlezon, W.A., Jr. Bi-directional effects of pituitary adenylate cyclase-activating polypeptide (PACAP) on fear-related behavior and c-Fos expression after fear conditioning in rats. Psychoneuroendocrinology 2016, 64, 12–21. [Google Scholar] [CrossRef]
- Roman, C.W.; Lezak, K.R.; Hartsock, M.J.; Falls, W.A.; Braas, K.M.; Howard, A.B.; Hammack, S.E.; May, V. PAC1 receptor antagonism in the bed nucleus of the stria terminalis (BNST) attenuates the endocrine and behavioral consequences of chronic stress. Psychoneuroendocrinology 2014, 47, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Ago, Y.; Hayata, A.; Hashimoto, H. [Pathophysiological implication of the VPAC2 receptor in psychiatric disorders]. Nihon Yakurigaku Zasshi 2018, 151, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.S.; Almli, L.M.; Fani, N.; Gutman, D.A.; Bradley, B.; Norrholm, S.D.; Reiser, E.; Ely, T.D.; Dhanani, R.; Glover, E.M.; et al. PACAP receptor gene polymorphism impacts fear responses in the amygdala and hippocampus. Proc. Natl. Acad. Sci. USA 2014, 111, 3158–3163. [Google Scholar] [CrossRef] [Green Version]
- Lindert, J.; von Ehrenstein, O.S.; Wehrwein, A.; Brahler, E.; Schafer, I. [Anxiety, Depression and Posttraumatic Stress Disorder in Refugees—A Systematic Review]. Psychother. Psychosom. Med. Psychol. 2018, 68, 22–29. [Google Scholar]
- Lao, K.; Ji, N.; Zhang, X.; Qiao, W.; Tang, Z.; Gou, X. Drug development for Alzheimer’s disease: Review. J. Drug Target. 2019, 27, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Onoue, S.; Endo, K.; Ohshima, K.; Yajima, T.; Kashimoto, K. The neuropeptide PACAP attenuates beta-amyloid (1-42)-induced toxicity in PC12 cells. Peptides 2002, 23, 1471–1478. [Google Scholar] [CrossRef]
- Kojro, E.; Postina, R.; Buro, C.; Meiringer, C.; Gehrig-Burger, K.; Fahrenholz, F. The neuropeptide PACAP promotes the alpha-secretase pathway for processing the Alzheimer amyloid precursor protein. FASEB J. 2006, 20, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, W.D.; Wang, Y.D. beta-Amyloid: The key peptide in the pathogenesis of Alzheimer’s disease. Front. Pharmacol. 2015, 6, 221. [Google Scholar] [CrossRef]
- Dogrukol-Ak, D.; Kumar, V.B.; Ryerse, J.S.; Farr, S.A.; Verma, S.; Nonaka, N.; Nakamachi, T.; Ohtaki, H.; Niehoff, M.L.; Edwards, J.C.; et al. Isolation of peptide transport system-6 from brain endothelial cells: Therapeutic effects with antisense inhibition in Alzheimer and stroke models. J. Cereb. Blood Flow Metab. 2009, 29, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Ciallella, J.R.; Flood, D.G.; O’Kane, T.M.; Bozyczko-Coyne, D.; Savage, M.J. Comparative analysis of cortical gene expression in mouse models of Alzheimer’s disease. Neurobiol. Aging 2006, 27, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.; Ceconi, C.; Curello, S.; Cargnoni, A.; De Giuli, F.; Visioli, O. Occurrence of oxidative stress during myocardial reperfusion. Mol. Cell Biochem. 1992, 111, 61–69. [Google Scholar] [CrossRef]
- Ross-Ascuitto, N.T.; Ascuitto, R.J.; Ramage, D.; Kydon, D.W.; Coy, D.H.; Kadowitz, P.J. Pituitary adenylate cyclase activating polypeptide: A neuropeptide with potent inotropic and coronary vasodilatory effects in neonatal pig hearts. Pediatr. Res. 1993, 34, 323–328. [Google Scholar] [CrossRef]
- Sano, H.; Miyata, A.; Horio, T.; Nishikimi, T.; Matsuo, H.; Kangawa, K. The effect of pituitary adenylate cyclase activating polypeptide on cultured rat cardiocytes as a cardioprotective factor. Regul. Pept. 2002, 109, 107–113. [Google Scholar] [CrossRef]
- Racz, B.; Reglodi, D.; Horvath, G.; Szigeti, A.; Balatonyi, B.; Roth, E.; Weber, G.; Alotti, N.; Toth, G.; Gasz, B. Protective effect of PACAP against doxorubicin-induced cell death in cardiomyocyte culture. J. Mol. Neurosci. 2010, 42, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.; Weber, G.; Kiss, P.; Horvath, G.; Toth, G.; Gasz, B.; Ferencz, A.; Gallyas, F., Jr.; Reglodi, D.; Racz, B. Effects of PACAP and preconditioning against ischemia/reperfusion-induced cardiomyocyte apoptosis in vitro. Ann. N. Y. Acad. Sci. 2009, 1163, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Gasz, B.; Racz, B.; Roth, E.; Borsiczky, B.; Ferencz, A.; Tamas, A.; Cserepes, B.; Lubics, A.; Gallyas, F., Jr.; Toth, G.; et al. Pituitary adenylate cyclase activating polypeptide protects cardiomyocytes against oxidative stress-induced apoptosis. Peptides 2006, 27, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Zhang, Y.; Shen, X.D.; Gao, F.; Huang, C.Y.; Abad, C.; Busuttil, R.W.; Waschek, J.A.; Kupiec-Weglinski, J.W. Neuropeptide PACAP in mouse liver ischemia and reperfusion injury: Immunomodulation by the cAMP-PKA pathway. Hepatology 2013, 57, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Alston, E.N.; Parrish, D.C.; Hasan, W.; Tharp, K.; Pahlmeyer, L.; Habecker, B.A. Cardiac ischemia-reperfusion regulates sympathetic neuropeptide expression through gp130-dependent and independent mechanisms. Neuropeptides 2011, 45, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Nakamachi, T.; Ohtaki, H.; Yofu, S.; Sato, A.; Endo, K.; Iso, Y.; Suzuki, H.; Takeyama, Y.; Shintani, N.; et al. Cardioprotective effect of endogenous pituitary adenylate cyclase-activating polypeptide on Doxorubicin-induced cardiomyopathy in mice. Circ. J. 2010, 74, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Szakaly, P.; Laszlo, E.; Kovacs, K.; Racz, B.; Horvath, G.; Ferencz, A.; Lubics, A.; Kiss, P.; Tamas, A.; Brubel, R.; et al. Mice deficient in pituitary adenylate cyclase activating polypeptide (PACAP) show increased susceptibility to in vivo renal ischemia/reperfusion injury. Neuropeptides 2011, 45, 113–121. [Google Scholar] [CrossRef]
- Brar, B.K.; Stephanou, A.; Liao, Z.; O’Leary, R.M.; Pennica, D.; Yellon, D.M.; Latchman, D.S. Cardiotrophin-1 can protect cardiac myocytes from injury when added both prior to simulated ischaemia and at reoxygenation. Cardiovasc. Res. 2001, 51, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Taga, T.; Saito, M.; Suematsu, S.; Kumanogoh, A.; Tanaka, T.; Fujiwara, H.; Hirata, M.; Yamagami, T.; Nakahata, T.; et al. Targeted disruption of gp130, a common signal transducer for the interleukin 6 family of cytokines, leads to myocardial and hematological disorders. Proc. Natl. Acad. Sci. USA 1996, 93, 407–411. [Google Scholar] [CrossRef]
- Khan, A.M.; Li, M.; Abdulnour-Nakhoul, S.; Maderdrut, J.L.; Simon, E.E.; Batuman, V. Delayed administration of pituitary adenylate cyclase-activating polypeptide 38 ameliorates renal ischemia/reperfusion injury in mice by modulating Toll-like receptors. Peptides 2012, 38, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Balamuthusamy, S.; Khan, A.M.; Maderdrut, J.L.; Simon, E.E.; Batuman, V. Pituitary adenylate cyclase-activating polypeptide ameliorates cisplatin-induced acute kidney injury. Peptides 2010, 31, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Peuskens, J.; Devoitille, J.M.; Kusters, J.; Vandenhoven, G. An open multicentre pilot study examining the safety, efficacy and tolerability of fast titrated (800 mg/day by day 4) quetiapine in the treatment of schizophrenia/schizoaffective disorder. Int. J. Psychiatry Clin. Pract. 2008, 12, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Abylkassov, R.; Xie, Y. Role of Yes-associated protein in cancer: An update. Oncol. Lett. 2016, 12, 2277–2282. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W. Methods for antagonizing glutamate neurotoxicity. Cerebrovasc. Brain Metab. Rev. 1990, 2, 105–147. [Google Scholar] [PubMed]
- Won, S.J.; Kim, D.Y.; Gwag, B.J. Cellular and molecular pathways of ischemic neuronal death. J. Biochem. Mol. Biol. 2002, 35, 67–86. [Google Scholar] [CrossRef] [PubMed]
- Dijk, F.; Kamphuis, W. Ischemia-induced alterations of AMPA-type glutamate receptor subunit. Expression patterns in the rat retina—An immunocytochemical study. Brain Res. 2004, 997, 207–221. [Google Scholar] [CrossRef]
- Sucher, N.J.; Lipton, S.A.; Dreyer, E.B. Molecular basis of glutamate toxicity in retinal ganglion cells. Vis. Res. 1997, 37, 3483–3493. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Kern, T.S.; Song, B.; Stuebe, C. Mechanistic Insights into Pathological Changes in the Diabetic Retina: Implications for Targeting Diabetic Retinopathy. Am. J. Pathol. 2017, 187, 9–19. [Google Scholar] [CrossRef]
- Atlasz, T.; Babai, N.; Reglodi, D.; Kiss, P.; Tamas, A.; Bari, F.; Domoki, F.; Gabriel, R. Diazoxide is protective in the rat retina against ischemic injury induced by bilateral carotid occlusion and glutamate-induced degeneration. Neurotox Res. 2007, 12, 105–111. [Google Scholar] [CrossRef]
- Seki, S.; Aoki, M.; Hosokawa, T.; Saito, T.; Masuma, R.; Komori, M.; Kurasaki, M. Bisphenol-A suppresses neurite extension due to inhibition of phosphorylation of mitogen-activated protein kinase in PC12 cells. Chem. Biol. Interact. 2011, 194, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Vaczy, A.; Reglodi, D.; Somoskeoy, T.; Kovacs, K.; Lokos, E.; Szabo, E.; Tamas, A.; Atlasz, T. The Protective Role of PAC1-Receptor Agonist Maxadilan in BCCAO-Induced Retinal Degeneration. J. Mol. Neurosci. 2016, 60, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Danyadi, B.; Bognar, E.; Szabadfi, K.; Fabian, E.; Kiss, P.; Mester, L.; Manavalan, S.; Atlasz, T.; Gabriel, R.; et al. Effect of PACAP on MAP kinases, Akt and cytokine expressions in rat retinal hypoperfusion. Neurosci. Lett. 2012, 523, 93–98. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Cervia, D.; Catalani, E.; Gevi, F.; Zolla, L.; Casini, G. Protective effects of the neuropeptides PACAP, substance P and the somatostatin analogue octreotide in retinal ischemia: A metabolomic analysis. Mol. Biosyst. 2014, 10, 1290–1304. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Pink, M.; Boland, S.; Rettenmeier, A.W.; Schmitz-Spanke, S. Benzo[a]pyrene-induced metabolic shift from glycolysis to pentose phosphate pathway in the human bladder cancer cell line RT4. Sci. Rep. 2017, 7, 9773. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Nakamachi, T.; Endo, K.; Seki, T.; Ohtaki, H.; Tsuchikawa, D.; Hori, M.; Tsuchida, M.; Yoshikawa, A.; Matkovits, A.; et al. PACAP attenuates NMDA-induced retinal damage in association with modulation of the microglia/macrophage status into an acquired deactivation subtype. J. Mol. Neurosci. 2013, 51, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zeltzer, S.; Zhang, Y.; Wang, F.; Chen, G.H.; Dayrit, J.; Murdock, B.J.; Bhan, U.; Toews, G.B.; Osterholzer, J.J.; et al. Early induction of CCL7 downstream of TLR9 signaling promotes the development of robust immunity to cryptococcal infection. J. Immunol. 2012, 188, 3940–3948. [Google Scholar] [CrossRef]
- Suk, K.; Park, J.H.; Lee, W.H. Neuropeptide PACAP inhibits hypoxic activation of brain microglia: A protective mechanism against microglial neurotoxicity in ischemia. Brain Res. 2004, 1026, 151–156. [Google Scholar] [CrossRef]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef]
- Stetler, R.A.; Gao, Y.; Zukin, R.S.; Vosler, P.S.; Zhang, L.; Zhang, F.; Cao, G.; Bennett, M.V.; Chen, J. Apurinic/apyrimidinic endonuclease APE1 is required for PACAP-induced neuroprotection against global cerebral ischemia. Proc. Natl. Acad. Sci. USA 2010, 107, 3204–3209. [Google Scholar] [CrossRef] [Green Version]
- Mercer, A.; Ronnholm, H.; Holmberg, J.; Lundh, H.; Heidrich, J.; Zachrisson, O.; Ossoinak, A.; Frisen, J.; Patrone, C. PACAP promotes neural stem cell proliferation in adult mouse brain. J. Neurosci. Res. 2004, 76, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shi, Q.D.; Song, T.B.; Feng, G.F.; Zang, W.J.; Zong, C.H.; Chang, L. Vasoactive intestinal peptide increases VEGF expression to promote proliferation of brain vascular endothelial cells via the cAMP/PKA pathway after ischemic insult in vitro. Peptides 2013, 42, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Darsalia, V.; Mansouri, S.; Wolbert, P.; Barde, S.; Sjoholm, A.; Patrone, C. The specific VPAC2 agonist Bay 55-9837 increases neuronal damage and hemorrhagic transformation after stroke in type 2 diabetic rats. Neuropeptides 2013, 47, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Vanhoutte, P.M. Macro- and microvascular endothelial dysfunction in diabetes. J. Diabetes 2017, 9, 434–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, Y.; Tuazon, J.P.; Ji, X.; Borlongan, C.V. Pituitary Adenylate Cyclase Activating Polypeptide Elicits Neuroprotection Against Acute Ischemic Neuronal Cell Death Associated with NMDA Receptors. Cell Physiol. Biochem. 2018, 51, 1982–1995. [Google Scholar] [CrossRef] [PubMed]
- Gomariz, R.P.; Juarranz, Y.; Abad, C.; Arranz, A.; Leceta, J.; Martinez, C. VIP-PACAP system in immunity: New insights for multitarget therapy. Ann. N. Y. Acad. Sci. 2006, 1070, 51–74. [Google Scholar] [CrossRef] [PubMed]
- Pozo, D.; Delgado, M.; Martinez, C.; Gomariz, R.P.; Guerrero, J.M.; Calvo, J.R. Functional characterization and mRNA expression of pituitary adenylate cyclase activating polypeptide (PACAP) type I receptors in rat peritoneal macrophages. Biochim. Biophys. Acta 1997, 1359, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Ottaway, C.A. In vitro alteration of receptors for vasoactive intestinal peptide changes the in vivo localization of mouse T cells. J. Exp. Med. 1984, 160, 1054–1069. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Shima, K.; Said, S.I. Characterization of vasoactive intestinal peptide receptors on rat alveolar macrophages. Am. J. Physiol. 1994, 267, L256–L262. [Google Scholar] [CrossRef] [PubMed]
- Wiik, P.; Opstad, P.K.; Boyum, A. Binding of vasoactive intestinal polypeptide (VIP) by human blood monocytes: Demonstration of specific binding sites. Regul. Pept. 1985, 12, 145–153. [Google Scholar] [CrossRef]
- Lara-Marquez, M.L.; O’Dorisio, M.S.; Karacay, B. Vasoactive intestinal peptide (VIP) receptor type 2 (VPAC2) is the predominant receptor expressed in human thymocytes. Ann. N. Y. Acad. Sci. 2000, 921, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.; Juarranz, Y.; Abad, C.; Arranz, A.; Miguel, B.G.; Rosignoli, F.; Leceta, J.; Gomariz, R.P. Analysis of the role of the PAC1 receptor in neutrophil recruitment, acute-phase response, and nitric oxide production in septic shock. J. Leukoc Biol. 2005, 77, 729–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, M.; Ganea, D. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibit interleukin-12 transcription by regulating nuclear factor kappaB and Ets activation. J. Biol. Chem. 1999, 274, 31930–31940. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, K.; Jiang, H.; Song, F.; Gao, H.; Pan, X.; Shi, B.; Bi, Y.; Wang, H.; Wang, H.; et al. Development of T cells carrying two complementary chimeric antigen receptors against glypican-3 and asialoglycoprotein receptor 1 for the treatment of hepatocellular carcinoma. Cancer Immunol. Immunother. 2017, 66, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Abad, C.; Tan, Y.V. Immunomodulatory Roles of PACAP and VIP: Lessons from Knockout Mice. J. Mol. Neurosci. 2018, 66, 102–113. [Google Scholar] [CrossRef]
- Martinez, C.; Abad, C.; Delgado, M.; Arranz, A.; Juarranz, M.G.; Rodriguez-Henche, N.; Brabet, P.; Leceta, J.; Gomariz, R.P. Anti-inflammatory role in septic shock of pituitary adenylate cyclase-activating polypeptide receptor. Proc. Natl. Acad. Sci. USA 2002, 99, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Goetzl, E.J.; Voice, J.K.; Shen, S.; Dorsam, G.; Kong, Y.; West, K.M.; Morrison, C.F.; Harmar, A.J. Enhanced delayed-type hypersensitivity and diminished immediate-type hypersensitivity in mice lacking the inducible VPAC(2) receptor for vasoactive intestinal peptide. Proc. Natl. Acad. Sci. USA 2001, 98, 13854–13859. [Google Scholar] [CrossRef]
- Jongsma, H.; Pettersson, L.M.; Zhang, Y.; Reimer, M.K.; Kanje, M.; Waldenstrom, A.; Sundler, F.; Danielsen, N. Markedly reduced chronic nociceptive response in mice lacking the PAC1 receptor. Neuroreport 2001, 12, 2215–2219. [Google Scholar] [CrossRef]
- Jana, B.; Palus, K.; Meller, K.; Calka, J. Porcine dorsal root ganglia ovarian neurons are affected by long lasting testosterone treatment. Physiol. Res. 2016, 65, 1019–1030. [Google Scholar]
- Kozlowska, A.; Mikolajczyk, A.; Adamiak, Z.; Majewski, M. Distribution and chemical coding of sensory neurons innervating the skin of the porcine hindlimb. Neuropeptides 2017, 61, 1–14. [Google Scholar] [CrossRef]
- Moller, K.; Zhang, Y.Z.; Hakanson, R.; Luts, A.; Sjolund, B.; Uddman, R.; Sundler, F. Pituitary adenylate cyclase activating peptide is a sensory neuropeptide: Immunocytochemical and immunochemical evidence. Neuroscience 1993, 57, 725–732. [Google Scholar] [CrossRef]
- Dun, E.C.; Huang, R.L.; Dun, S.L.; Dun, N.J. Pituitary adenylate cyclase activating polypeptide-immunoreactivity in human spinal cord and dorsal root ganglia. Brain Res. 1996, 721, 233–237. [Google Scholar] [CrossRef]
- Zhang, Y.; Danielsen, N.; Sundler, F.; Mulder, H. Pituitary adenylate cyclase-activating peptide is upregulated in sensory neurons by inflammation. Neuroreport 1998, 9, 2833–2836. [Google Scholar] [CrossRef] [PubMed]
- Mulder, H.; Jongsma, H.; Zhang, Y.; Gebre-Medhin, S.; Sundler, F.; Danielsen, N. Pituitary adenylate cyclase-activating polypeptide and islet amyloid polypeptide in primary sensory neurons: Functional implications from plasticity in expression on nerve injury and inflammation. Mol. Neurobiol. 1999, 19, 229–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Alm, P.; Hakanson, R. The contribution of nitric oxide to endotoxin-induced ocular inflammation: Interaction with sensory nerve fibres. Br. J. Pharmacol 1996, 118, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Shioda, S.; Takenoya, F.; Hirabayashi, T.; Wada, N.; Seki, T.; Nonaka, N.; Nakamachi, T. Effects of PACAP on Dry Eye Symptoms, and Possible Use for Therapeutic Application. J. Mol. Neurosci 2019, 68, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Kazanis, I. The subependymal zone neurogenic niche: A beating heart in the centre of the brain: How plastic is adult neurogenesis? Opportunities for therapy and questions to be addressed. Brain 2009, 132, 2909–2921. [Google Scholar] [CrossRef] [PubMed]
- Mercier, F. Fractones: Extracellular matrix niche controlling stem cell fate and growth factor activity in the brain in health and disease. Cell Mol. Life Sci. 2016, 73, 4661–4674. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, R.; Ohtaki, H.; Nakamachi, T.; Watanabe, J.; Yofu, S.; Hayashi, D.; Takeda, T.; Nonaka, N.; Seki, M.; Nakamura, M.; et al. Distribution and localization of pituitary adenylate cyclase-activating polypeptide-specific receptor (PAC1R) in the rostral migratory stream of the infant mouse brain. Regul. Pept. 2008, 145, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Chiu, L.; Lee, H.T.; Chiang, C.W.; Liu, S.P.; Hsu, Y.H.; Lin, S.Z.; Hsu, C.Y.; Hsieh, C.H.; Shyu, W.C. PACAP38/PAC1 signaling induces bone marrow-derived cells homing to ischemic brain. Stem Cells 2015, 33, 1153–1172. [Google Scholar] [CrossRef]
- Suh, J.; Lu, N.; Nicot, A.; Tatsuno, I.; DiCicco-Bloom, E. PACAP is an anti-mitogenic signal in developing cerebral cortex. Nat. Neurosci. 2001, 4, 123–124. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, S.; Agartz, I.; Ogren, S.O.; Patrone, C.; Lundberg, M. PACAP Protects Adult Neural Stem Cells from the Neurotoxic Effect of Ketamine Associated with Decreased Apoptosis, ER Stress and mTOR Pathway Activation. PLoS ONE 2017, 12, e0170496. [Google Scholar] [CrossRef] [PubMed]
- Nyisztor, Z.; Denes, V.; Kovacs-Valasek, A.; Hideg, O.; Berta, G.; Gabriel, R. Pituitary Adenylate Cyclase Activating Polypeptide (PACAP1-38) Exerts Both Pro and Anti-Apoptotic Effects on Postnatal Retinal Development in Rat. Neuroscience 2018, 385, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, M.; Furuta, A.; Aoki, S.; Kudo, Y.; Miyakawa, H.; Wada, K. PACAP/PAC1 autocrine system promotes proliferation and astrogenesis in neural progenitor cells. Glia 2007, 55, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Njaine, B.; Rocha-Martins, M.; Vieira-Vieira, C.H.; De-Melo, L.D.; Linden, R.; Braas, K.; May, V.; Martins, R.A.; Silveira, M.S. Pleiotropic functions of pituitary adenylyl cyclase-activating polypeptide on retinal ontogenesis: Involvement of KLF4 in the control of progenitor cell proliferation. J. Mol. Neurosci. 2014, 54, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Hansel, D.E.; Eipper, B.A.; Ronnett, G.V. Regulation of olfactory neurogenesis by amidated neuropeptides. J. Neurosci. Res. 2001, 66, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ohtaki, H.; Watanabe, J.; Miyamoto, K.; Murai, N.; Sasaki, S.; Matsumoto, M.; Hashimoto, H.; Hiraizumi, Y.; Numazawa, S.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) contributes to the proliferation of hematopoietic progenitor cells in murine bone marrow via PACAP-specific receptor. Sci. Rep. 2016, 6, 22373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce, M.; Canipari, R.; Ferri, G.L.; Siracusa, G.; De Felici, M. Pituitary adenylate cyclase-activating polypeptide (PACAP) stimulates adenylate cyclase and promotes proliferation of mouse primordial germ cells. Development 1996, 122, 215–221. [Google Scholar] [PubMed]
- McManus, M.F.; Chen, L.C.; Vallejo, I.; Vallejo, M. Astroglial differentiation of cortical precursor cells triggered by activation of the cAMP-dependent signaling pathway. J. Neurosci. 1999, 19, 9004–9015. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Lelievre, V.; Zhao, P.; Torres, M.; Rodriguez, W.; Byun, J.Y.; Doshi, S.; Ioffe, Y.; Gupta, G.; de los Monteros, A.E.; et al. Pituitary adenylyl cyclase-activating polypeptide stimulates DNA synthesis but delays maturation of oligodendrocyte progenitors. J. Neurosci. 2001, 21, 3849–3859. [Google Scholar] [CrossRef] [PubMed]
- Masmoudi-Kouki, O.; Gandolfo, P.; Castel, H.; Leprince, J.; Fournier, A.; Dejda, A.; Vaudry, H.; Tonon, M.C. Role of PACAP and VIP in astroglial functions. Peptides 2007, 28, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Falluel-Morel, A.; Chafai, M.; Vaudry, D.; Basille, M.; Cazillis, M.; Aubert, N.; Louiset, E.; de Jouffrey, S.; Le Bigot, J.F.; Fournier, A.; et al. The neuropeptide pituitary adenylate cyclase-activating polypeptide exerts anti-apoptotic and differentiating effects during neurogenesis: Focus on cerebellar granule neurones and embryonic stem cells. J. Neuroendocrinol. 2007, 19, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Fila, T.; Trazzi, S.; Crochemore, C.; Bartesaghi, R.; Ciani, E. Lot1 is a key element of the pituitary adenylate cyclase-activating polypeptide (PACAP)/cyclic AMP pathway that negatively regulates neuronal precursor proliferation. J. Biol. Chem. 2009, 284, 15325–15338. [Google Scholar] [CrossRef] [PubMed]
- Allais, A.; Burel, D.; Isaac, E.R.; Gray, S.L.; Basille, M.; Ravni, A.; Sherwood, N.M.; Vaudry, H.; Gonzalez, B.J. Altered cerebellar development in mice lacking pituitary adenylate cyclase-activating polypeptide. Eur. J. Neurosci. 2007, 25, 2604–2618. [Google Scholar] [CrossRef] [PubMed]
- Hirose, M.; Niewiadomski, P.; Tse, G.; Chi, G.C.; Dong, H.; Lee, A.; Carpenter, E.M.; Waschek, J.A. Pituitary adenylyl cyclase-activating peptide counteracts hedgehog-dependent motor neuron production in mouse embryonic stem cell cultures. J. Neurosci. Res. 2011, 89, 1363–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Njaine, B.; Martins, R.A.; Santiago, M.F.; Linden, R.; Silveira, M.S. Pituitary adenylyl cyclase-activating polypeptide controls the proliferation of retinal progenitor cells through downregulation of cyclin D1. Eur. J. Neurosci. 2010, 32, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Freson, K. From patient to mouse to therapy: Role of the neuropeptide pacap in platelet function and formation. Verh K Acad. Geneeskd Belg. 2010, 72, 239–251. [Google Scholar] [PubMed]
- Merlo, L.M.; Pepper, J.W.; Reid, B.J.; Maley, C.C. Cancer as an evolutionary and ecological process. Nat. Rev. Cancer 2006, 6, 924–935. [Google Scholar] [CrossRef]
- Capp, J.P. Cancer Stem Cells: From Historical Roots to a New Perspective. J. Oncol. 2019, 2019, 5189232. [Google Scholar] [CrossRef]
- Wang, X. Stem cells in tissues, organoids, and cancers. Cell Mol. Life Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Cauvin, A.; Buscail, L.; Gourlet, P.; De Neef, P.; Gossen, D.; Arimura, A.; Miyata, A.; Coy, D.H.; Robberecht, P.; Christophe, J. The novel VIP-like hypothalamic polypeptide PACAP interacts with high affinity receptors in the human neuroblastoma cell line NB-OK. Peptides 1990, 11, 773–777. [Google Scholar] [CrossRef]
- Vertongen, P.; De Clerck, P.; Fournet, J.C.; Martelli, H.; Helardot, P.; Devalck, C.; Peeters, T.; Sariban, E.; Robberecht, P. Comparison between vasoactive intestinal polypeptide and pituitary adenylate cyclase activating polypeptide levels in neuroblastoma tumour tissues. Neuropeptides 1997, 31, 409–413. [Google Scholar] [CrossRef]
- Jaworski, D.M. Expression of pituitary adenylate cyclase-activating polypeptide (PACAP) and the PACAP-selective receptor in cultured rat astrocytes, human brain tumors, and in response to acute intracranial injury. Cell Tissue Res. 2000, 300, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Lieu, S.N.; Oh, D.S.; Pisegna, J.R.; Germano, P.M. Neuroendocrine tumors express PAC1 receptors. Ann. N. Y. Acad. Sci. 2006, 1070, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Vertongen, P.; Ciccarelli, E.; Woussen-Colle, M.C.; De Neef, P.; Robberecht, P.; Cauvin, A. Pituitary adenylate cyclase-activating polypeptide receptors of types I and II and glucagon-like peptide-I receptors are expressed in the rat medullary carcinoma of the thyroid cell line 6/23. Endocrinology 1994, 135, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Buscail, L.; Gourlet, P.; Cauvin, A.; De Neef, P.; Gossen, D.; Arimura, A.; Miyata, A.; Coy, D.H.; Robberecht, P.; Christophe, J. Presence of highly selective receptors for PACAP (pituitary adenylate cyclase activating peptide) in membranes from the rat pancreatic acinar cell line AR 4-2J. FEBS Lett. 1990, 262, 77–81. [Google Scholar] [CrossRef]
- Olcese, J.; McArdle, C.A.; Middendorff, R.; Greenland, K. Pituitary adenylate cyclase-activating peptide and vasoactive intestinal peptide receptor expression in immortalized LHRH neurons. J. Neuroendocrinol. 1997, 9, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Jamen, F.; Laden, J.C.; Bouschet, T.; Rodriguez-Henche, N.; Bockaert, J.; Brabet, P. Nerve growth factor upregulates the PAC1 promoter by activating the MAP kinase pathway in rat PC12 cells. Ann. N. Y. Acad. Sci. 2000, 921, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Haidan, A.; Hilbers, U.; Bornstein, S.R.; Ehrhart-Bornstein, M. Human adrenocortical NCI-H295 cells express VIP receptors. Steroidogenic effect of vasoactive intestinal peptide (VIP). Peptides 1998, 19, 1511–1517. [Google Scholar] [CrossRef]
- Dagar, S.; Sekosan, M.; Rubinstein, I.; Onyuksel, H. Detection of VIP receptors in MNU-induced breast cancer in rats: Implications for breast cancer targeting. Breast Cancer Res. Treat. 2001, 65, 49–54. [Google Scholar] [CrossRef]
- Madsen, B.; Georg, B.; Madsen, M.W.; Fahrenkrug, J. Estradiol down regulates expression of vasoactive intestinal polypeptide receptor type-1 in breast cancer cell lines. Mol. Cell Endocrinol. 2001, 172, 203–211. [Google Scholar] [CrossRef]
- Waschek, J.A.; Richards, M.L.; Bravo, D.T. Differential expression of VIP/PACAP receptor genes in breast, intestinal, and pancreatic cell lines. Cancer Lett. 1995, 92, 143–149. [Google Scholar] [CrossRef]
- Busto, R.; Carrero, I.; Guijarro, L.G.; Solano, R.M.; Zapatero, J.; Noguerales, F.; Prieto, J.C. Expression, pharmacological, and functional evidence for PACAP/VIP receptors in human lung. Am. J. Physiol. 1999, 277, L42–L48. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernandez, M.O.; Collado, B.; Bodega, G.; Cortes, J.; Ruiz-Villaespesa, A.; Carmena, M.J.; Prieto, J.C. Pituitary adenylate cyclase-activating peptide/vasoactive intestinal peptide receptors in human normal mammary gland and breast cancer tissue. Gynecol. Endocrinol. 2005, 20, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Moretti, C.; Mammi, C.; Frajese, G.V.; Mariani, S.; Gnessi, L.; Arizzi, M.; Wannenes, F.; Frajese, G. PACAP and type I PACAP receptors in human prostate cancer tissue. Ann. N. Y. Acad. Sci. 2006, 1070, 440–449. [Google Scholar] [CrossRef]
- Robberecht, P.; Vertongen, P.; Velkeniers, B.; de Neef, P.; Vergani, P.; Raftopoulos, C.; Brotchi, J.; Hooghe-Peters, E.L.; Christophe, J. Receptors for pituitary adenylate cyclase activating peptides in human pituitary adenomas. J. Clin. Endocrinol. Metab. 1993, 77, 1235–1239. [Google Scholar] [PubMed]
- Vertongen, P.; d’Haens, J.; Michotte, A.; Velkeniers, B.; van Rampelbergh, J.; Svoboda, M.; Robberecht, P. Expression of pituitary adenylate cyclase activating polypeptide and receptors in human brain tumors. Peptides 1995, 16, 713–719. [Google Scholar] [CrossRef]
- Reubi, J.C.; Laderach, U.; Waser, B.; Gebbers, J.O.; Robberecht, P.; Laissue, J.A. Vasoactive intestinal peptide/pituitary adenylate cyclase-activating peptide receptor subtypes in human tumors and their tissues of origin. Cancer Res. 2000, 60, 3105–3112. [Google Scholar] [PubMed]
- Isobe, K.; Kaneko, M.; Kaneko, S.; Nissato, S.; Nanmoku, T.; Takekoshi, K.; Okuda, Y.; Kawakami, Y. Expression of mRNAs for PACAP and its receptor in human neuroblastomas and their relationship to catecholamine synthesis. Regul. Pept. 2004, 123, 29–32. [Google Scholar] [CrossRef]
- Waschek, J.A.; Lelievre, V.; Bravo, D.T.; Nguyen, T.; Muller, J.M. Retinoic acid regulation of the VIP and PACAP autocrine ligand and receptor system in human neuroblastoma cell lines. Peptides 1997, 18, 835–841. [Google Scholar] [CrossRef]
- Takahashi, K.; Totsune, K.; Murakami, O.; Sone, M.; Itoi, K.; Hayashi, Y.; Ohi, R.; Mouri, T. Pituitary adenylate cyclase activating polypeptide (PACAP)-like immunoreactivity in ganglioneuroblastoma and neuroblastoma. Regul. Pept. 1993, 49, 19–24. [Google Scholar] [CrossRef]
- Spada, A.; Lania, A.; Mantovani, S. Cellular abnormalities in pituitary tumors. Metabolism 1996, 45 (Suppl. 1), 46–48. [Google Scholar] [CrossRef]
- Garcia-Fernandez, M.O.; Bodega, G.; Ruiz-Villaespesa, A.; Cortes, J.; Prieto, J.C.; Carmena, M.J. PACAP expression and distribution in human breast cancer and healthy tissue. Cancer Lett. 2004, 205, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Fahrenkrug, J.; Buhl, T.; Hannibal, J. PreproPACAP-derived peptides occur in VIP-producing tumours and co-exist with VIP. Regul. Pept. 1995, 58, 89–98. [Google Scholar] [CrossRef]
- Odum, L.; Fahrenkrug, J. Pituitary adenylate cyclase activating polypeptide (PACAP) in human ovarian cancers. Cancer Lett. 1998, 125, 185–189. [Google Scholar] [CrossRef]
- Gutierrez-Canas, I.; Rodriguez-Henche, N.; Bolanos, O.; Carmena, M.J.; Prieto, J.C.; Juarranz, M.G. VIP and PACAP are autocrine factors that protect the androgen-independent prostate cancer cell line PC-3 from apoptosis induced by serum withdrawal. Br. J. Pharmacol. 2003, 139, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godlewski, J.; Kowalczyk, A.; Kozielec, Z.; Pidsudko, Z.; Kmiec, A.; Siedlecka-Kroplewska, K. Plasticity of neuropeptidergic neoplasm cells in the primary and metastatic Merkel cell carcinoma. Folia Histochem. Cytobiol. 2013, 51, 168–173. [Google Scholar] [CrossRef]
- Nakamachi, T.; Sugiyama, K.; Watanabe, J.; Imai, N.; Kagami, N.; Hori, M.; Arata, S.; Shioda, S. Comparison of expression and proliferative effect of pituitary adenylate cyclase-activating polypeptide (PACAP) and its receptors on human astrocytoma cell lines. J. Mol. Neurosci. 2014, 54, 388–394. [Google Scholar] [CrossRef]
- Zia, F.; Fagarasan, M.; Bitar, K.; Coy, D.H.; Pisegna, J.R.; Wank, S.A.; Moody, T.W. Pituitary adenylate cyclase activating peptide receptors regulate the growth of non-small cell lung cancer cells. Cancer Res. 1995, 55, 4886–4891. [Google Scholar]
- Castorina, A.; Giunta, S.; Scuderi, S.; D’Agata, V. Involvement of PACAP/ADNP signaling in the resistance to cell death in malignant peripheral nerve sheath tumor (MPNST) cells. J. Mol. Neurosci. 2012, 48, 674–683. [Google Scholar] [CrossRef]
- Vertongen, P.; Camby, I.; Darro, F.; Kiss, R.; Robberecht, P. VIP and pituitary adenylate cyclase activating polypeptide (PACAP) have an antiproliferative effect on the T98G human glioblastoma cell line through interaction with VIP2 receptor. Neuropeptides 1996, 30, 491–496. [Google Scholar] [CrossRef]
- Li, M.; Cortez, S.; Nakamachi, T.; Batuman, V.; Arimura, A. Pituitary adenylate cyclase-activating polypeptide is a potent inhibitor of the growth of light chain-secreting human multiple myeloma cells. Cancer Res. 2006, 66, 8796–8803. [Google Scholar] [CrossRef] [PubMed]
- Ajioka, I.; Martins, R.A.; Bayazitov, I.T.; Donovan, S.; Johnson, D.A.; Frase, S.; Cicero, S.A.; Boyd, K.; Zakharenko, S.S.; Dyer, M.A. Differentiated horizontal interneurons clonally expand to form metastatic retinoblastoma in mice. Cell 2007, 131, 378–390. [Google Scholar] [CrossRef]
- Denes, V.; Hideg, O.; Nyisztor, Z.; Lakk, M.; Berta, G.; Toth, K.; Gabriel, R. The neuroprotective peptide, PACAP signaling could be critical in retinoblastoma through induction of horizontal cell proliferation. Presented at European Retina Meeting, Helsinki, Finland, 12–14 September 2019. [Google Scholar]
- Rubio, G.; Marin, M.; Arias, F.; Lopez-Trabada, J.R.; Iribarren, M.; Alfonso, S.; Prieto, R.; Blanco, A.; Urosa, B.; Montes, V.; et al. Inclusion of Alcoholic Associations Into a Public Treatment Programme for Alcoholism Improves Outcomes During the Treatment and Continuing Care Period: A 6-Year Experience. Alcohol. Alcohol. 2018, 53, 78–88. [Google Scholar] [CrossRef] [PubMed]



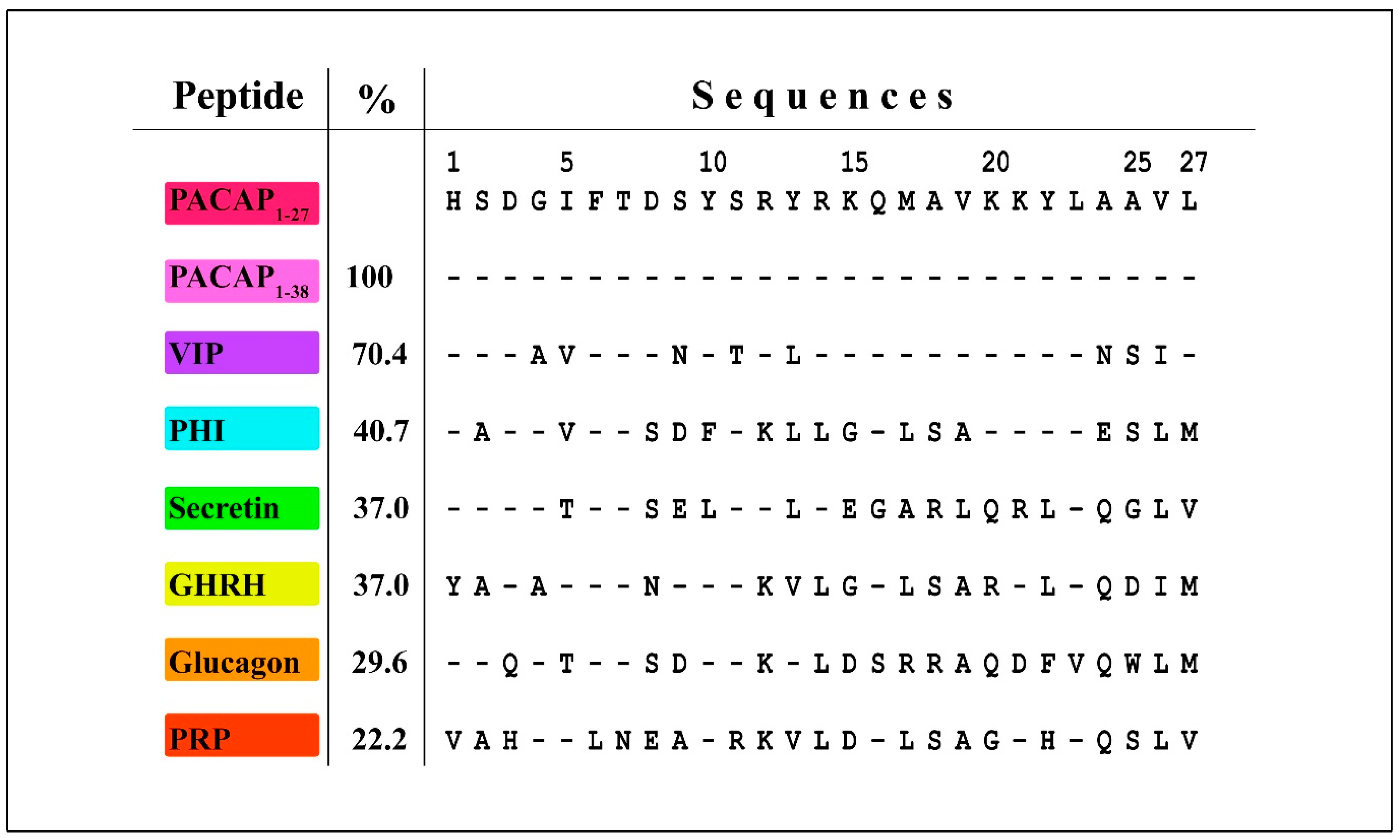{kind=link}
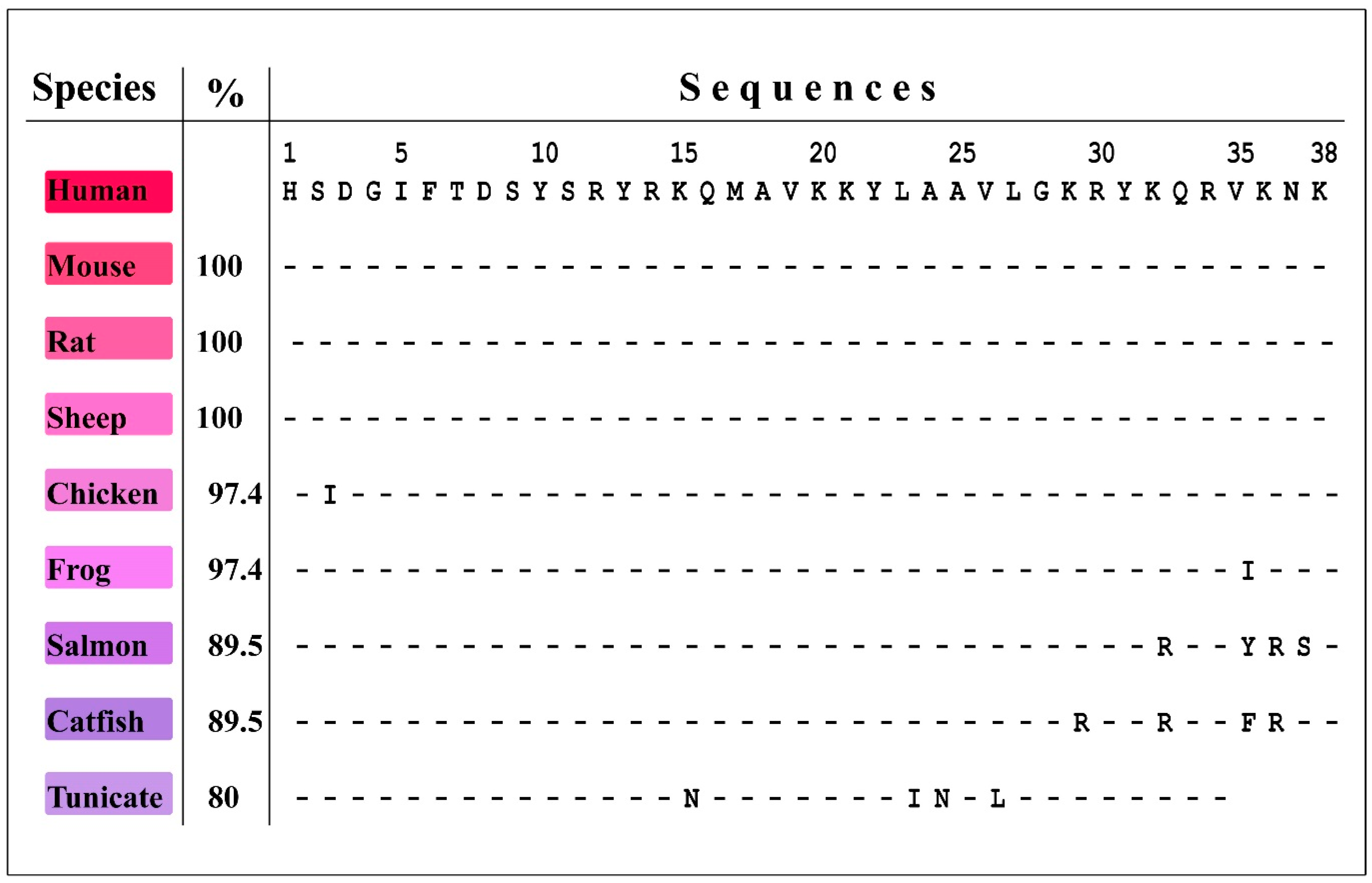{kind=link}
| Disorder | Implication | PACAP/VIP Receptor | Outcome | Literature |
|---|---|---|---|---|
| Cancer | pathogenesis/therapeutic | for details see Section 6.2 | ||
| Breast cancer | pathogenesis | PAC1, VPAC1, VPAC2 | [28] | |
| Cervical cancer | therapeutic | PAC1 | [29,30] | |
| Colon cancer | pathogenesis/therapeutic | PAC1 (Hip), VPAC1 | [31,32,33] | |
| Gastric cancer | pathogenesis | PAC1, VPAC1 | [34,35] | |
| Gliomas | pathogenesis/therapeutic | PAC1, VPAC1, VPAC2 | [36,37] | |
| Lung caner | pathogenesis | PAC1, VPAC1, VPAC2 | [38] | |
| Malignant peripheral nerve sheath tumor (MPNST) | pathogenesis | not determined | [39] | |
| Malignant pheochromocytomas | pathogenesis | PAC1, VPAC1 | [40] | |
| Medulloblastoma | therapeutic | PAC1 | [41,42] | |
| Pancreatic cancer | pathogenesis | PAC1, VPAC1 | [43] | |
| Pituitary adenomas | [44] | |||
| Prostate | pathogenesis | PAC1 (null), VPAC1 | [45,46] | |
| Renal cell carcinoma | therapeutic | VPAC1 | [47] | |
| Retinoblastoma | unclear | PAC1 | [48,49] | |
| Diabetes (Type II, insulin resistance) | therapeutic | PAC1, VPAC1, VPAC2 | - induces insulin secretion, - PAC1 antagonist impairs glucose tolerance - PACAP overexpression induces beta cell proliferation | [50,51,52,53,54] |
| Diabetic retinopathy | therapeutic | PAC1, VPAC1, VPAC2 | - increases Akt and ERK1/2 phosphorylation - reduces the activation of p38 mitogen-activated protein kinase - reduces the expression of IL-1β in diabetic animals - downregulates VEGF and VEGF receptors, inversely regulates HIFs: downregulating HIF-1α and HIF-2α while upregulating HIF-3α - restores both Bcl-2 and p53 mRNA and protein expression - maintains synapse integrity | [55,56,57,58,59,60] |
| Diabetic nephropathy | therapeutic | not determined | - downregulates of several cytokines including CINC-1, TIMP-1, LIX, MIG, s-ICAM | [61] |
| Inflammation | Therapeutic/pathogenesis | for details see Section 6.1 | ||
| Allergic airway inflammation | therapeutic | PAC1, VPAC1, VPAC2 | [62] | |
| Atherosclerosis | therapeutic | not determined | [63] | |
| Chronic cystitis, urinary bladder inflammation | pathogenesis | PAC1, VPAC1, VPAC2 | [64,65] | |
| Endotoxin induced airway inflammation | therapeutic | PAC1, VPAC1, VPAC2 | [66] | |
| Ileitis | therapeutic | not determined | [67,68] | |
| Ocular inflammation | pathogenesis | not determined | [69] | |
| Osteoarthritis | therapeutic | not determined | [70,71] | |
| Pancreatitis | pathogenesis | not determined | - enhances caerulein-induced pancreatitis via downregulation of RegIII-β - lack of endogenous PACAP ameliorates symptoms | [72,73,74] |
| Skin neurogenic inflammation | pathogenesis | PAC1 | [75,76,77] | |
| Toxoplasmosis | therapeutic | VPAC1, VPAC2 | [78] | |
| Human immunodeficiency virus (HIV) infection | therapeutic | PAC1, VPAC1, VPAC2 | - increases macrophage resistance to HIV-1 replication - reduces macrophage production of HIV-1 - induces the synthesis of β-chemokines and IL-10 - inhibits NF-kB, and reduced Cyclin D1 levels | [79,80,81] |
| Neurological/neurodegenerative disorders | Therapeutic/pathogenesis | |||
| Alzheimer’s diseases | therapeutic | PAC1 | for details, see Section 5.1 | [82,83,84] |
| Bipolar disorder | therapeutic | PAC1 | - no association with SNPs of PACAP gene - regulates binding between DISC1 and DISC1-binding zinc-finger protein responsible for neurite outgrowth | [85,86] |
| Epilepsy | therapeutic | PAC1, VPAC1 | - during seizure PACAP is secreted and exert neuroprotective effects by regulating microglial phenotype, microglial interleukin secretion - inhibits long-term depression and depotentiation | [87,88] |
| Huntington’s diseases | therapeutic | PAC1 | - improves memory performance | [89] |
| Ischemia/stroke (cardiac, renal, hepatic, retinal, brain) | therapeutic | PAC1 | for details, see Section 5.2 | [90,91,92,93,94,95,96,97,98,99] |
| Migraine | pathogenesis | PAC1, VPAC1, VPAC2 | for details, see Section 4.1 | [100,101,102,103,104,105,106] |
| Major depression (MDD) | therapeutic | not determined | PACAP regulated DISC1 mutation is linked to major MDD - SNP3 (rs1893154) of the PACAP gene is significantly correlated with MDD | [107,108,109,110]. |
| Multiple sclerosis | therapeutic | VPAC1, PAC1 | - prevents Thelper1 cell differentiation and suppression of inflammatory interleukin production | [111,112,113,114,115,116,117,118,119] |
| Parkinson’s diseases | therapeutic | - potent preventive molecule against cell loss and autophagy - facilitates dopaminergic neurotransmission - effective PACAP analog was developed with reduced cardiovascular side effects | [120,121,122,123] | |
| Post-traumatic stress disorder | pathogenesis | PAC1 (VPAC2) | for details, see Section 4.2 | [4,124,125] |
| Schizophrenia | therapeutic | PAC1 | - SNPs of PACAP or PAC1 gene are associated with schizophrenia - regulates binding between DISC1 and DBZ responsible for neurite outgrowth | [86,110,126,127] |
| Traumatic injuries | ||||
| Traumatic brain injury | therapeutic | not determined | - attenuates neural injury by increasing level of SOD-2 and GPx-1 - suppresses T-cell response - decreases inflammatory interleukin levels | [128,129] |
| Spinal cord injury | therapeutic | PAC1 | - activates axon regeneration through CRMP-2 and activation of glial elements | [130,131] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denes, V.; Geck, P.; Mester, A.; Gabriel, R. Pituitary Adenylate Cyclase-Activating Polypeptide: 30 Years in Research Spotlight and 600 Million Years in Service. J. Clin. Med. 2019, 8, 1488. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091488
Denes V, Geck P, Mester A, Gabriel R. Pituitary Adenylate Cyclase-Activating Polypeptide: 30 Years in Research Spotlight and 600 Million Years in Service. Journal of Clinical Medicine. 2019; 8(9):1488. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091488
Chicago/Turabian StyleDenes, Viktoria, Peter Geck, Adrienn Mester, and Robert Gabriel. 2019. "Pituitary Adenylate Cyclase-Activating Polypeptide: 30 Years in Research Spotlight and 600 Million Years in Service" Journal of Clinical Medicine 8, no. 9: 1488. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091488