Grain Quality of Maize Cultivars as a Function of Planting Dates, Irrigation and Nitrogen Stress: A Case Study from Semiarid Conditions of Iran


Abstract
:
1. Introduction
2. Materials and Methods
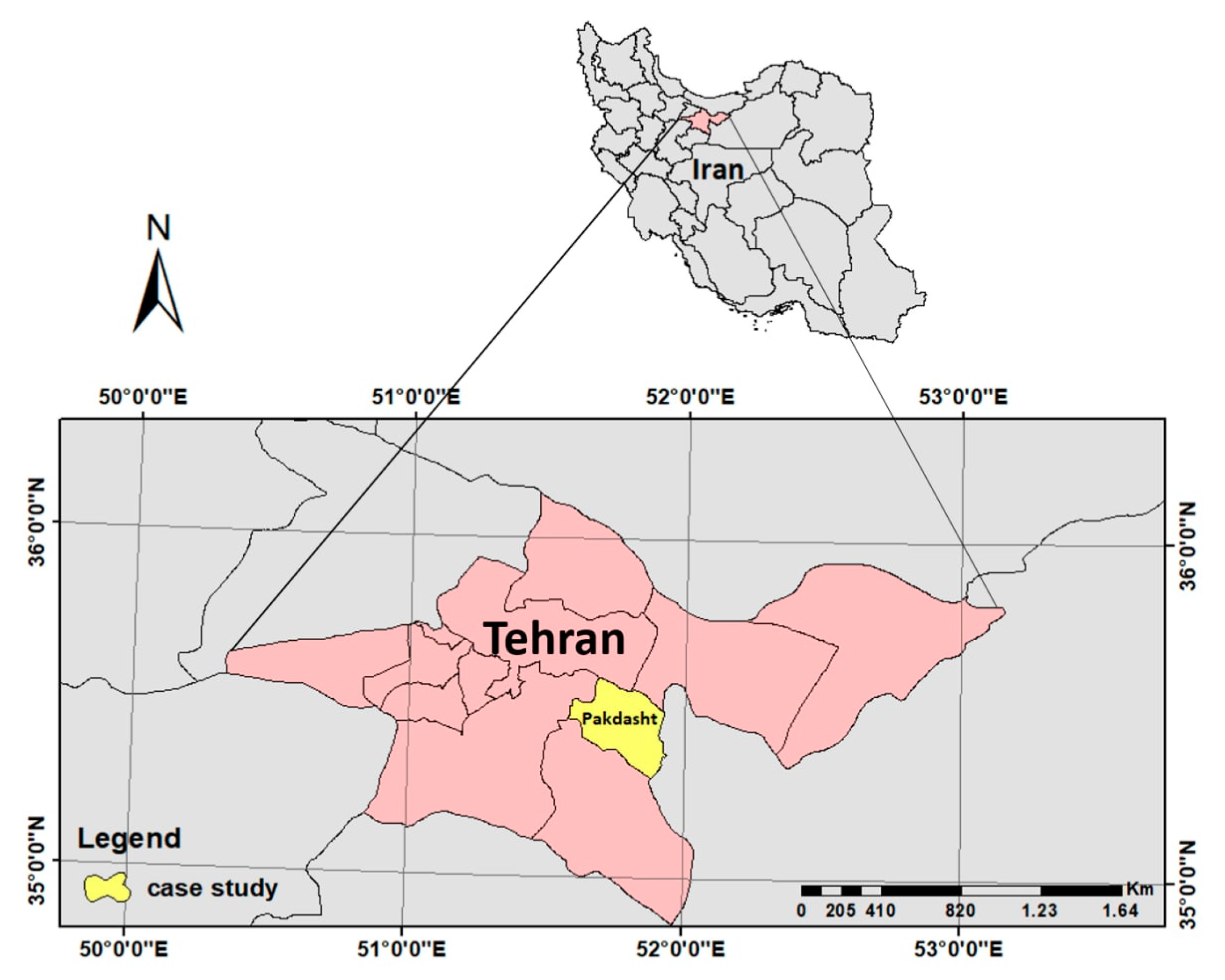
2.1. Experiment Site
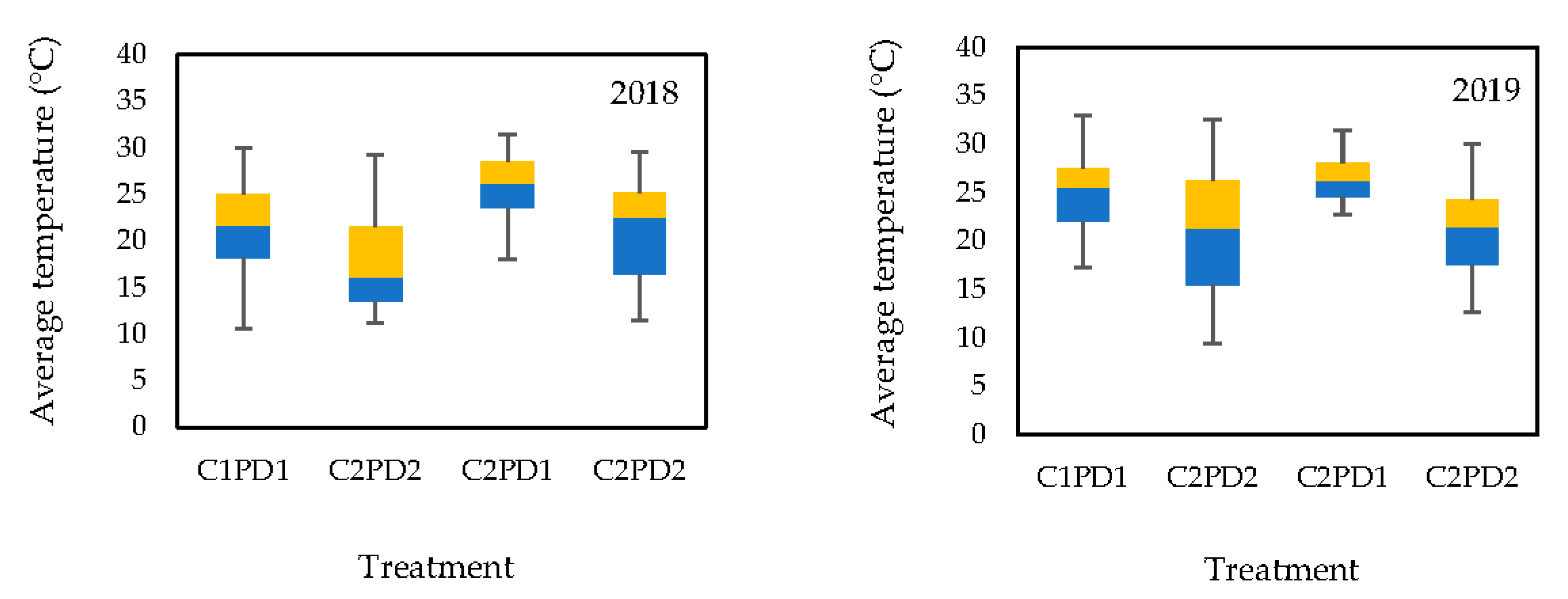
2.2. Experimental Design
2.3. Procedures of Sampling and Laboratory
2.3.1. Sample Preparation
2.3.2. Analysis of Starch Content and Composition
2.3.3. Analysis of Protein Content and Composition
2.3.4. Analysis of Oil Concentration and Fatty Acid Composition
2.4. Statistical Analysis
3. Results
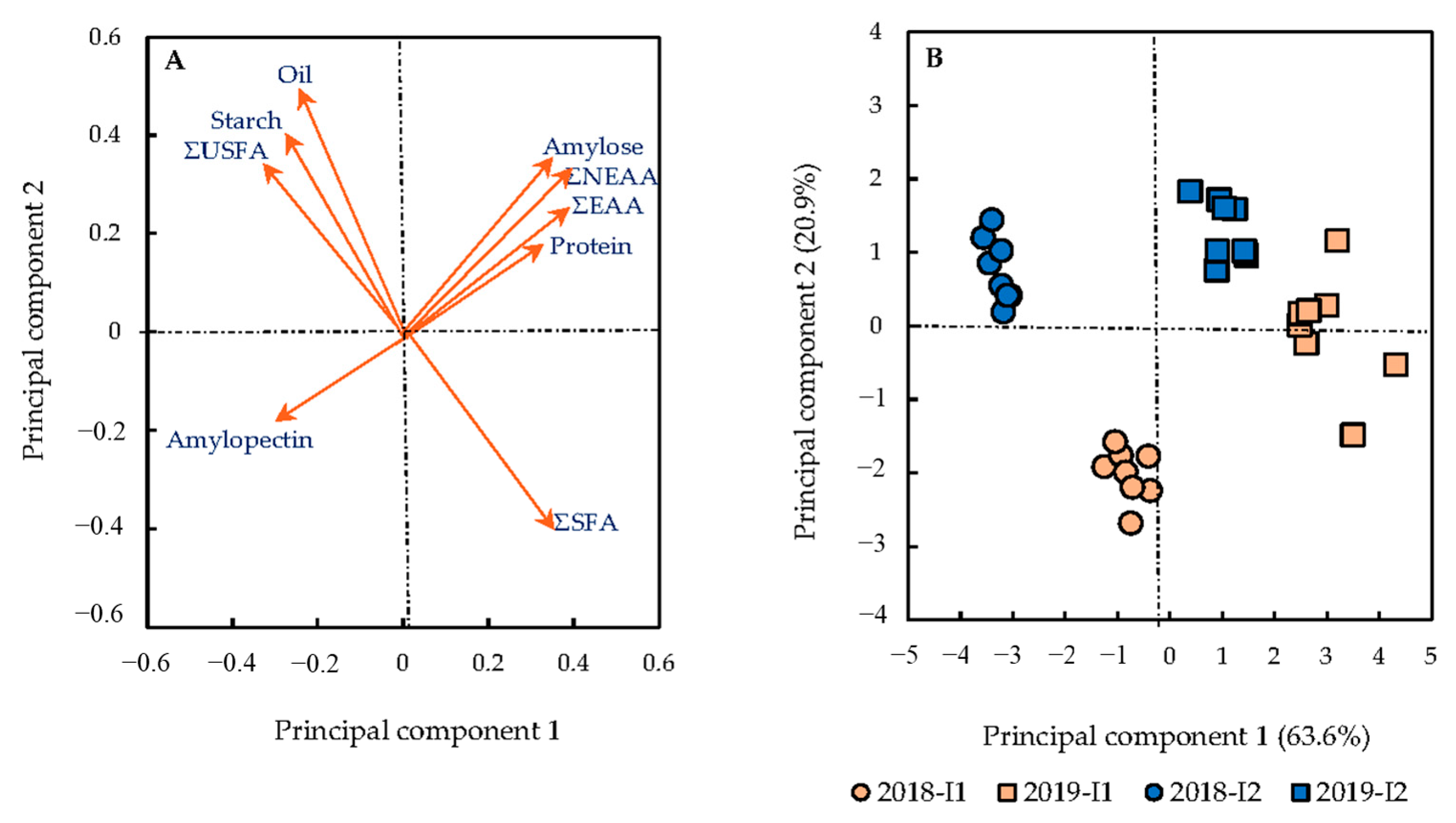
3.1. Correlations between Variables and Treatments
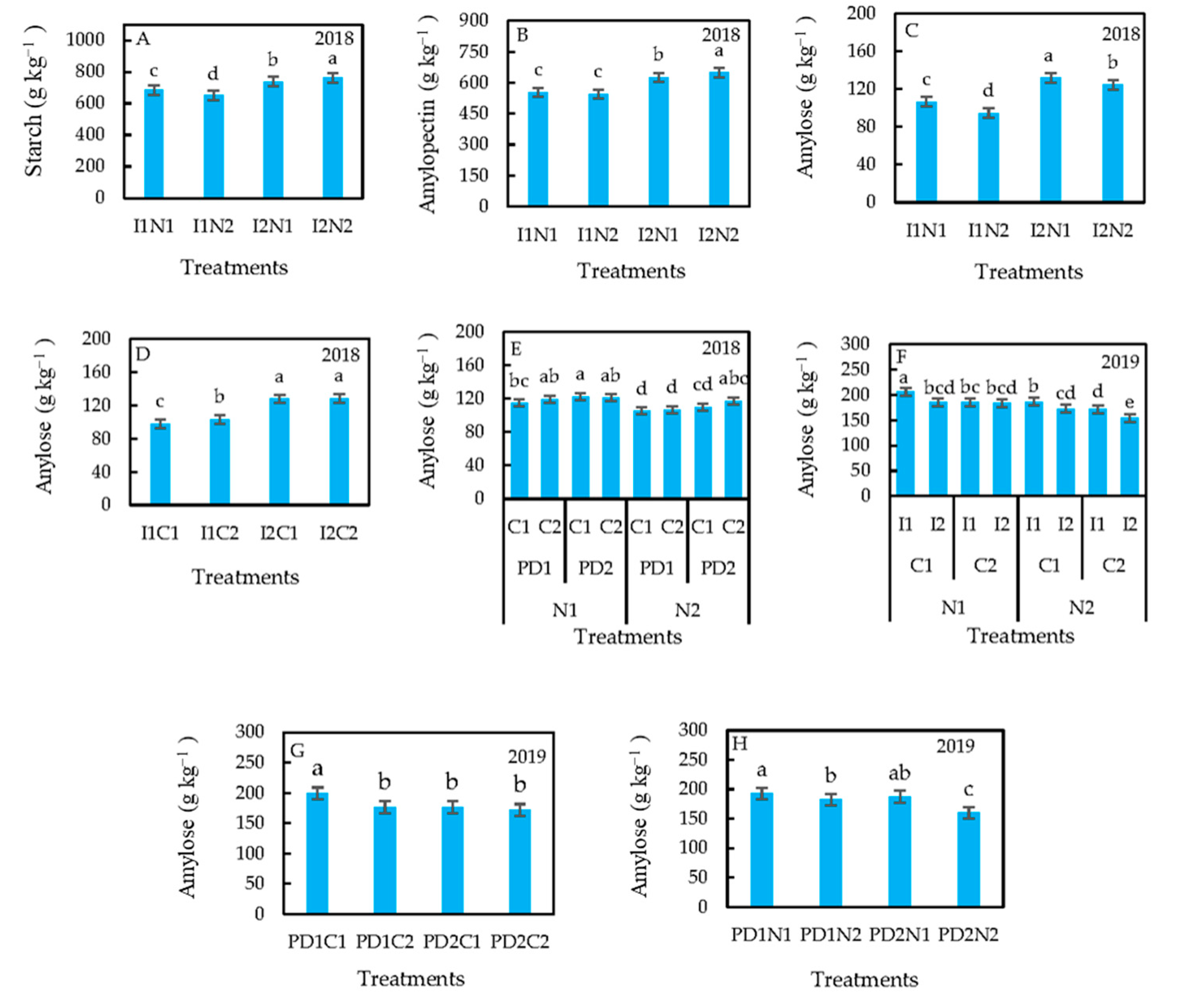
3.2. Treatment Effects on Starch Content and Composition
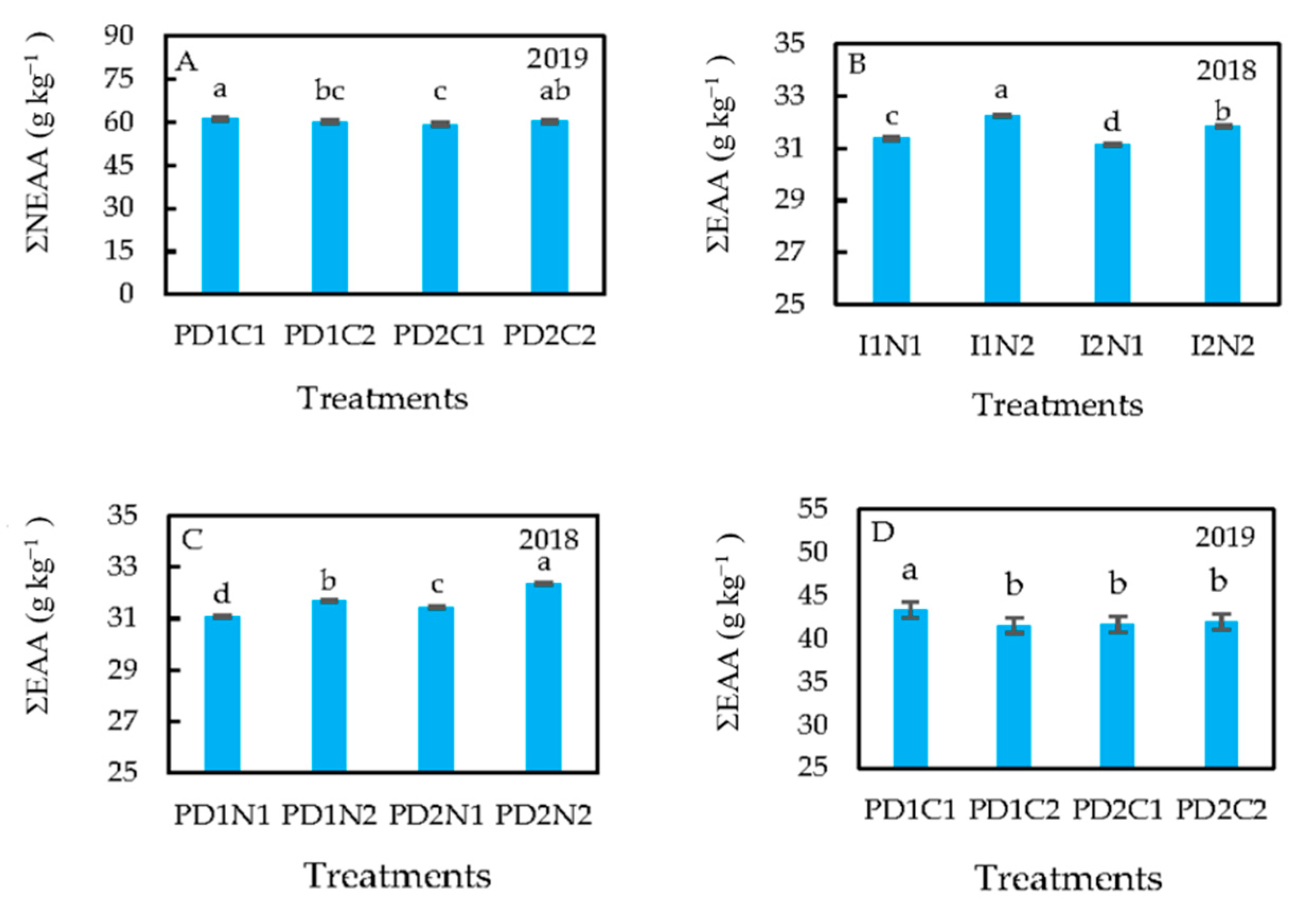
3.3. Treatment Effects on Protein Content and Composition
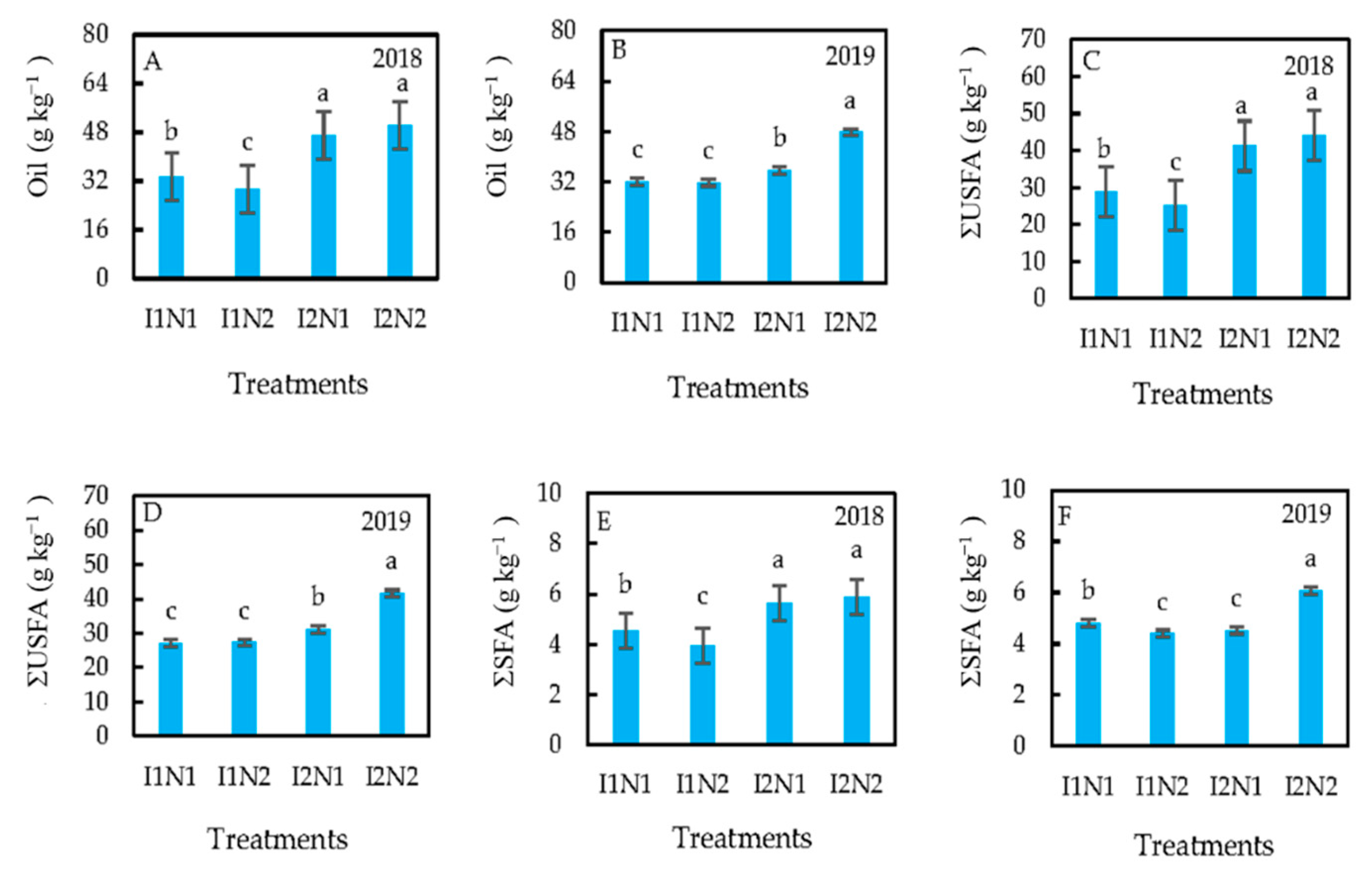
3.4. Treatment Effects on Oil Content and Composition
4. Discussion
4.1. Correlations between Variables and Treatments
4.2. Treatment Effects on Starch Content and Composition
4.3. Treatment Effects on Protein Content and Composition
4.4. Treatment Effects on Oil Content and Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darrah, L.L.; McMullen, M.; Zuber, M. Breeding, Genetics and Seed Corn Production. In Corn; Serna-Saldivar, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 19–41. [Google Scholar]
- Rausch, K.D.; Hummel, D.; Johnson, L.A.; May, J.B. Wet Milling: The Basis for Corn Biorefineries. In Corn; Serna-Saldivar, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 501–535. [Google Scholar]
- Ewu, Y.; Messing, J. Proteome balancing of the maize seed for higher nutritional value. Front. Plant Sci. 2014, 5, 240. [Google Scholar] [CrossRef] [Green Version]
- Moran, J.E.T. Starch: Granule, Amylose-Amylopectin, Feed Preparation, and Recovery by the Fowl’s Gastrointestinal Tract. J. Appl. Poult. Res. 2019, 28, 566–586. [Google Scholar] [CrossRef]
- Singh, N.; Singh, S.; Shevkani, K. Maize: Composition, Bioactive Constituents, and Unleavened Bread. In Flour and Breads and their Fortification in Health and Disease Prevention; Preedy, V., Watson, R., Patel, V., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 111–121. [Google Scholar]
- Copeland, L.; Blazek, J.; Salman, H.; Tang, M.C. Form and functionality of starch. Food Hydrocoll. 2009, 23, 1527–1534. [Google Scholar] [CrossRef]
- Cai, C.; Zhao, L.; Huang, J.; Chen, Y.; Wei, C. Morphology, structure and gelatinization properties of heterogeneous starch granules from high-amylose maize. Carbohydr. Polym. 2014, 102, 606–614. [Google Scholar] [CrossRef]
- Zhang, P.; Ma, G.; Wang, C.; Lu, H.; Li, S.; Xie, Y.; Ma, D.; Zhu, Y.; Guo, T. Effect of irrigation and nitrogen application on grain amino acid composition and protein quality in winter wheat. PLoS ONE 2017, 12, e0178494. [Google Scholar] [CrossRef]
- Allen, N.K.; Baker, D.H. Quantitative Evaluation of Nonspecific Nitrogen Sources for the Growing Chick. Poult. Sci. 1974, 53, 258–264. [Google Scholar] [CrossRef]
- Lenis, N.P.; Van Diepen, H.T.M.; Bikker, P.; Jongbloed, A.W.; Van Der Meulen, J. Effect of the ratio between essential and nonessential amino acids in the diet on utilization of nitrogen and amino acids by growing pigs. J. Anim. Sci. 1999, 77, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.-P.; Men, S.-Q.; Liu, Z.; Tong, N.-N.; Imran, M.; Shu, Q.Y. Fatty Acid Composition, Phytochemistry, Antioxidant Activity on Seed Coat and Kernel of Paeonia ostii from Main Geographic Production Areas. Foods 2019, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.P.; McMullen, M.D.; Holland, J.B.; Tian, F.; Bradbury, P.; Ross-Ibarra, J.; Buckler, E.S.; Flint-Garcia, S.A. Genetic Architecture of Maize Kernel Composition in the Nested Association Mapping and Inbred Association Panels. Plant Physiol. 2011, 158, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L. Animal Manure Fertiliser Value, Crop Utilisation and Soil Quality Impacts. Anim. Manure Recycl. 2013, 295–328. [Google Scholar] [CrossRef]
- Silva, R.R.; Zucareli, C.; Fonseca, I.C.D.B.; Riede, C.R.; Gazola, D. Nitrogen management, cultivars and growing environments on wheat grain quality. Rev. Bras. Eng. Agrícola Ambient. 2019, 23, 826–832. [Google Scholar] [CrossRef] [Green Version]
- Kresovic, B.; Gajic, B.; Tapanarova, A.; Dugalić, G. How Irrigation Water Affects the Yield and Nutritional Quality of Maize (Zea mays L.) in a Temperate Climate. Pol. J. Environ. Stud. 2018, 27, 1123–1131. [Google Scholar] [CrossRef]
- Struik, P.C. Effect of Temperature on Development, Dry-Matter Production, Dry-Matter Distribution and Quality of Forage Maize (Zea mays L.); Veenman & Zonen: Mariastraat, The Netherlands, 1983. [Google Scholar]
- Cao, X.; Sun, H.; Wang, C.; Ren, X.; Liu, H.; Zhang, Z. Effects of late-stage nitrogen fertilizer application on the starch structure and cooking quality of rice. J. Sci. Food Agric. 2018, 98, 2332–2340. [Google Scholar] [CrossRef]
- Asthir, B.; Jain, D.; Kaur, B.; Bains, N. Effect of nitrogen on starch and protein content in grain influence of nitrogen doses on grain starch and protein accumulation in diversified wheat genotypes. J. Environ. Biol. 2017, 38, 427–433. [Google Scholar] [CrossRef]
- Medic, J.; Abendroth, L.J.; Elmore, R.W.; Jane, J.-L.; Slukova, M. Effect of planting date on maize starch structure, properties, and ethanol production. Starch Stärke 2015, 68, 476–487. [Google Scholar] [CrossRef]
- Wang, J.; Hasegawa, T.; Li, L.; Lam, S.K.; Zhang, X.; Liu, X.; Pan, G. Changes in grain protein and amino acids composition of wheat and rice under short-term increased [CO2] and temperature of canopy air in a paddy from East China. New Phytol. 2019, 222, 726–734. [Google Scholar] [CrossRef]
- Dai, Z.-M.; Yin, Y.-P.; Zhang, M.; Li, W.-Y.; Yan, S.-H.; Cai, R.; Wang, Z. Distribution of Starch Granule Size in Grains of Wheat Grown Under Irrigated and Rainfed Conditions. Acta Agron. Sin. 2008, 34, 795–802. [Google Scholar] [CrossRef]
- Kaplan, M.; Kale, H.; Karaman, K.; Unlukara, A. Influence of different irrigation and nitrogen levels on crude oil and fatty acid composition of maize (Zea mays L.). Grasas Aceites 2017, 68, 207. [Google Scholar] [CrossRef] [Green Version]
- Anwar, S.; Iqbal, M.; Akram, H.M.; Niaz, M.; Rasheed, R. Influence of Drought Applied at Different Growth Stages on Kernel Yield and Quality in Maize (Zea mays L.). Commun. Soil Sci. Plant Anal. 2016, 47, 2225–2232. [Google Scholar] [CrossRef]
- Impa, S.; Perumal, R.; Bean, S.R.; Sunoj, V.J.; Jagadish, S.V.K. Water deficit and heat stress induced alterations in grain physico-chemical characteristics and micronutrient composition in field grown grain sorghum. J. Cereal Sci. 2019, 86, 124–131. [Google Scholar] [CrossRef]
- Bicego, B.; Sapkota, A.; Torrion, J.A. Differential Nitrogen and Water Impacts on Yield and Quality of Wheat Classes. Agron. J. 2019, 111, 2792–2803. [Google Scholar] [CrossRef] [Green Version]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954.
- Mehlich, A. Determination of P, Ca, Mg, K, Na, and NH4; North Carolina Soil Test Division: Raleigh, NC, USA, 1953; pp. 23–89. [Google Scholar]
- Saxton, K.E.; Rawls, W.J.; Romberger, J.S.; Papendick, R.I. Estimating Generalized Soil-water Characteristics from Texture. Soil Sci. Soc. Am. J. 1986, 50, 1031–1036. [Google Scholar] [CrossRef]
- Akbari, G.A.; Heshmati, S.; Soltani, E.; Dehaghi, M.A. Influence of Seed Priming on Seed Yield, Oil Content and Fatty Acid Composition of Safflower (Carthamus tinctorius L.) Grown Under Water Deficit. Int. J. Plant Prod. 2019, 245–258. [Google Scholar] [CrossRef]
- Estakhr, A.; Heidari, B.; Ahmadi, Z. Evaluation of kernel yield and agronomic traits of European maize hybrids in the temperate region of Iran. Arch. Agron. Soil Sci. 2015, 61, 475–491. [Google Scholar] [CrossRef]
- Jahahngirlou, R.M. Investigation of the Morphophysiological Aspects of Yield Formation in Maize (Zea Mays L.) Cultivars and the Agro-Ecological Assessment of Their Performance in Khouzestan and Fars Provinces. Master’s Thesis, Shahid Beheshti University, Tehran, Iran, 2015. [Google Scholar]
- Soufizadeh, S. GDD Calculator Program; Shahid Beheshti University: Tehran, Iran, 2012. [Google Scholar]
- Cutforth, H.; Shaykewich, C. A temperature response function for corn development. Agric. For. Meteorol. 1990, 50, 159–171. [Google Scholar] [CrossRef]
- FAOSTAT. Statistical Databases and Data-Sets of the Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org (accessed on 5 November 2020).
- Hanway, J. How a corn plant develops. Iowa State Univ. Coop. Ext. Serv. Spec. Rep. 1966, 48, 1–17. [Google Scholar]
- Megazyme, Amylose/Amylopectin. Assay Procedure for Measurement of Amylose and Amylopectin of Starch. Available online: http://www.megazyme.com (accessed on 5 November 2020).
- AOAC. Official Methods of Analysis of AOAC International, 20th ed.; AOAC International: Rockville, MD, USA, 2016; Volume 2, pp. 20–25. [Google Scholar]
- Minitab, L.L.C. SAS, v.19; Minitab. Inc.: State College, PA, USA, 2006. [Google Scholar]
- SAS Institute. The SAS System. v.9.4; SAS Institute: Cary, NC, USA, 2003. [Google Scholar]
- Hammac, W.A.; Maaz, T.M.; Koenig, R.T.; Burke, I.C.; Pan, W.L. Water and Temperature Stresses Impact Canola (Brassica napus L.) Fatty Acid, Protein, and Yield over Nitrogen and Sulfur. J. Agric. Food Chem. 2017, 65, 10429–10438. [Google Scholar] [CrossRef]
- Singletary, G.; Banisadr, R.; Keeling, P. Heat Stress During Grain Filling in Maize: Effects on Carbohydrate Storage and Metabolism. Funct. Plant Biol. 1994, 21, 829–841. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Ranjbar, G.; Pakniyat, H.; Emam, Y.; Azooz, M.; Ahmad, P. Plant-Environment Interaction: Responses and Approaches to Mitigate Stress; Wiley Blackwell: Hoboken, NJ, USA, 2016. [Google Scholar]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; Hanumantharao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saastamoinen, M.; Kumpulainen, J.; Nummela, S.; Häkkinen, U. Effect of Temperature on Oil Content and Fatty Acid Composition of Oat Grains. Acta Agric. Scand. 1990, 40, 349–356. [Google Scholar] [CrossRef]
- Elferjani, R.; Soolanayakanahally, R.Y. Canola Responses to Drought, Heat, and Combined Stress: Shared and Specific Effects on Carbon Assimilation, Seed Yield, and Oil Composition. Front. Plant Sci. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccardi, F. Protein Changes in Response to Progressive Water Deficit in Maize. Quantitative Variation and Polypeptide Identification. Plant Physiol. 1998, 117, 1253–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, M.; Peterson, C.J.; Rizza, M.D.; Dellavalle, P.D.; Vázquez, D.; Ibanez, V.; Ross, A. Influence of Heat Stress on Wheat Grain Characteristics and Protein Molecular Weight Distribution. In Molecular Breeding of Forage and Turf; Hopkins, A., Wang, Z.Y., Mian, R., Sledge, M., Barker, R.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 12, pp. 365–371. [Google Scholar]
- Butts-Wilmsmeyer, C.J.; Seebauer, J.R.; Singleton, L.; Below, F.E. Weather during Key Growth Stages Explains Grain Quality and Yield of Maize. Agronomy 2019, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Kaya, Y.; Ii, I.; Akcura, M. Effects of genotype and environment on grain yield and quality traits in bread wheat (T. aestivum L.). Food Sci. Technol. 2014, 34, 386–393. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.C.; Calderón, F.J.; Hatfield, J.L.; Sauer, T.J. Fallow Effects on Soil. In Soil Management: Building a Stable Base for Agriculture; Hatfield, J.L., Saue, T.J., Eds.; Soil Science Society of America: Madison, WI, USA, 2015; pp. 287–300. [Google Scholar]
- Rahimi, J.M.; Kambouzia, J.; Zand, E.; Rezayi, M. Investigation of grain yield and some related traits in different maize cultivars (Zea mays L.). J. Plant Physiol. 2019, 10, 150–166. [Google Scholar]
- Hurkman, W.J.; McCue, K.F.; Altenbach, S.B.; Korn, A.; Tanaka, C.K.; Kothari, K.M.; Johnson, E.L.; Bechtel, D.B.; Wilson, J.D.; Anderson, O.D.; et al. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci. 2003, 164, 873–881. [Google Scholar] [CrossRef]
- Chen, G.; Zhen, S.; Liu, Y.; Yan, X.; Zhang, M.; Yan, Y. In vivo phosphoproteome characterization reveals key starch granule-binding phosphoproteins involved in wheat water-deficit response. BMC Plant Biol. 2017, 17, 168. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Wu, D.; Yang, W.; Yang, X.; Huang, L. Sprinkler irrigation enhancing accumulation and quality properties of starch in wheat grain. Trans. Chin. Soc. Agric. Eng. 2015, 31, 97–102. [Google Scholar]
- Uribelarrea, M.; Below, F.E.; Moose, S.P. Grain Composition and Productivity of Maize Hybrids Derived from the Illinois Protein Strains in Response to Variable Nitrogen Supply. Crop. Sci. 2004, 44, 1593–1600. [Google Scholar] [CrossRef]
- Jiyun, J.; Ping, H.; Hailong, L.; Wenjuan, L.; Shaowen, H.; Xiufang, W.; Lichun, W.; Jiagui, X.; Guogang, Z. Comparison of nitrogen absorption, yield and quality between high-starch and common corn as affected by nitrogen application. J. Plant Nutr. Soil Sci. 2004, 10, 568–573. [Google Scholar]
- Kaplan, M.; Karaman, K.; Kardes, Y.M.; Kale, H. Phytic acid content and starch properties of maize (Zea mays L.): Effects of irrigation process and nitrogen fertilizer. Food Chem. 2019, 283, 375–380. [Google Scholar] [CrossRef]
- Kaufman, R.C.; Wilson, J.; Bean, S.R.; Presley, D.R.; Blanco-Canqui, H.; Mikha, M. Effect of Nitrogen Fertilization and Cover Cropping Systems on Sorghum Grain Characteristics. J. Agric. Food Chem. 2013, 61, 5715–5719. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, S.; Zhang, B.; Qiao, D.; Pu, H.; Liu, S.; Li, L. Structural features and thermal property of propionylated starches with different amylose/amylopectin ratio. Int. J. Biol. Macromol. 2017, 97, 123–130. [Google Scholar] [CrossRef]
- Jane, J.-L. Current Understanding on Starch Granule Structures. J. Appl. Glycosci. 2006, 53, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Blanco, M.; Jane, J.-L. Physicochemical properties of endosperm and pericarp starches during maize development. Carbohydr. Polym. 2007, 67, 630–639. [Google Scholar] [CrossRef]
- Pierre, C.S.; Peterson, C.J.; Ross, A.; Ohm, J.-B.; Verhoeven, M.C.; Larson, M.; Hoefer, B. White Wheat Grain Quality Changes with Genotype, Nitrogen Fertilization, and Water Stress. Agron. J. 2008, 100, 414–420. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, M.; Zheng, L.; Cheng, D.-D.; Liu, M.; Geng, Y.-Q. Controlled Release Urea Improved Nitrogen Use Efficiency, Yield, and Quality of Wheat. Agron. J. 2011, 103, 479–485. [Google Scholar] [CrossRef]
- Gous, P.W.; Warren, F.J.; Mo, O.W.; Gilbert, R.G.; Fox, G.P. The effects of variable nitrogen application on barley starch structure under drought stress. J. Inst. Brew. 2015, 121, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liang, Z.-Y.; He, X.-M.; Meng, Q.-F.; Hu, Y.; Schmidhalter, U.; Zhang, W.; Zou, C.-Q.; Chen, X. Improving grain yield and protein concentration of maize (Zea mays L.) simultaneously by appropriate hybrid selection and nitrogen management. Field Crop. Res. 2020, 249, 107754. [Google Scholar] [CrossRef]
- Zhong, Y.; Xu, D.; Hebelstrup, K.H.; Yang, D.-L.; Cai, J.; Wang, X.; Zhou, Q.; Cao, W.; Dai, T.; Jiang, D. Nitrogen topdressing timing modifies free amino acids profiles and storage protein gene expression in wheat grain. BMC Plant Biol. 2018, 18, 353. [Google Scholar] [CrossRef]
- Harrigan, G.G.; Stork, L.G.; Riordan, S.G.; Reynolds, T.L.; Taylor, J.P.; Masucci, J.D.; Cao, Y.; LeDeaux, J.R.; Pandravada, A.; Glenn, K.C. Impact of environmental and genetic factors on expression of maize gene classes: Relevance to grain composition. J. Food Compos. Anal. 2009, 22, 158–164. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, H.; Zhao, C.; Chen, G.; Zou, Y. Amino acid content in rice grains is affected by high temperature during the early grain-filling period. Sci. Rep. 2019, 9, 2700. [Google Scholar] [CrossRef] [PubMed]
- Aguirrezabal, L.A.N.; Martre, P.; Pereyrairujo, G.; Echarte, M.M.; Izquierdo, N.G. Improving grain quality: Ecophysiological and modeling tools to develop management and breeding strategies. In Crop Physiology; Sadras, V.O., Calderini, D.F., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 423–465. [Google Scholar]
- Geesing, D.; Diacono, M.; Schmidhalter, U. Site-specific effects of variable water supply and nitrogen fertilisation on winter wheat. J. Plant Nutr. Soil Sci. 2014, 177, 509–523. [Google Scholar] [CrossRef]
- Singer, S.D.; Zou, J.; Weselake, R.J. Abiotic factors influence plant storage lipid accumulation and composition. Plant Sci. 2016, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.S.; Stark, E.M.L.M.; Zonta, E.; Fernandes, M.S.; Dos Santos, A.M.; De Souza, S.R. Different nitrogen and boron levels influence the grain production and oil content of a sunflower cultivar. Acta Sci. Agron. 2017, 39, 59. [Google Scholar] [CrossRef] [Green Version]
- Obeng, E.; Obour, A.K.; Nelson, N.O.; Moreno, J.A.; Ciampitti, I.A.; Wang, D.; Durrett, T.P. Seed yield and oil quality as affected by Camelina cultivar and planting date. J. Crop. Improv. 2019, 33, 202–222. [Google Scholar] [CrossRef]
- Van Der Merwe, R.; Labuschagne, M.; Herselman, L.; Hugo, A. Effect of heat stress on seed yield components and oil composition in high- and mid-oleic sunflower hybrids. S. Afr. J. Plant Soil 2015, 32, 121–128. [Google Scholar] [CrossRef]
- Schulte, L.R.; Ballard, T.C.; Samarakoon, T.B.; Yao, L.; Vadlani, P.V.; Staggenborg, S.; Rezac, M. Increased growing temperature reduces content of polyunsaturated fatty acids in four oilseed crops. Ind. Crop. Prod. 2013, 51, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Rivas, J.M.; Sánchez-García, A.; Sicardo, M.D.; García-Díaz, M.T.; Mancha, M. Oxygen-independent temperature regulation of the microsomal oleate desaturase (FAD2) activity in developing sunflower (Helianthus annuus) seeds. Physiol. Plant. 2003, 117, 179–185. [Google Scholar] [CrossRef]
- Parsons, D.; Cherney, J.; Gauch, H.G. Alfalfa Fiber Estimation in Mixed Stands and Its Relationship to Plant Morphology. Crop. Sci. 2006, 46, 2446–2452. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Temperature (°C) | Precipitation (mm) | ||||
|---|---|---|---|---|---|---|
| Minimum | Maximum | |||||
| 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | |
| June | 20.3 | 20.8 | 38.7 | 39.1 | 0.1 | 0.0 |
| July | 24.4 | 24.2 | 42.4 | 41.9 | 0.0 | 0.0 |
| August | 22.5 | 21.0 | 40.1 | 39.3 | 0.0 | 0.0 |
| September | 17.8 | 16.9 | 35.1 | 35.3 | 0.0 | 0.0 |
| October | 12.3 | 12.2 | 26.6 | 26.9 | 0.3 | 0.8 |
| November | 6.7 | 4.4 | 16.5 | 15.6 | 0.9 | 0.6 |
| Total | 17.3 | 16.6 | 33.2 | 33.0 | 0.24 | 0.23 |
| Year | Maize Hybrids | Planting Date | Harvesting Date | Maturity Period (Days) | GDDs (°C) |
|---|---|---|---|---|---|
| 2018 | KSC704 | 20 June | 7 November | 140 | 2424 |
| 21 July | 26 November | 138 | 1839 | ||
| KSC260 | 20 June | 18 October | 120 | 2311 | |
| 21 July | 11 November | 118 | 1796 | ||
| 2019 | KSC704 | 20 June | 2 November | 135 | 2370 |
| 21 July | 28 November | 130 | 1855 | ||
| KSC260 | 20 June | 12 October | 120 | 2220 | |
| 21 July | 14 November | 116 | 1704 |
| Treatment | Starch | Amylopectin | Amylose | Protein | ΣNEAA | ΣEAA | Oil | ΣUSFA | ΣSFA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g kg−1 | ||||||||||||||||||
| 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | |
| I1 | 668.10 | 682.50 | 549.17 | 494.23 | 100.48 | 187.36 | 105.00 | 121.40 | 42.20 | 61.90 | 31.80 | 43.30 | 31.30 | 31.90 | 26.98 | 27.20 | 4.24 | 4.60 |
| I2 | 750.80 | 684.00 | 637.77 | 509.22 | 128.26 | 173.99 | 97.50 | 116.10 | 39.00 | 58.50 | 31.50 | 41.60 | 48.70 | 41.70 | 42.71 | 36.33 | 5.75 | 5.30 |
| p-value | 0.056 | 0.937 | 0.043 | 0.565 | 0.035 | 0.133 | 0.206 | 0.337 | 0.042 | 0.137 | 0.012 | 0.334 | 0.068 | 0.022 | 0.065 | 0.022 | 0.083 | 0.005 |
| SEM | 7.31 | 15.51 | 6.00 | 18.44 | 1.50 | 2.83 | 2.52 | 3.11 | 2.14 | 7.36 | 0.05 | 9.42 | 1.86 | 3.37 | 1.62 | 3.20 | 0.19 | 0.55 |
| PD1 | 705.90 | 685.60 | 594.67 | 496.33 | 111.31 | 187.59 | 101.50 | 115.10 | 40.40 | 60.70 | 31.40 | 43.00 | 43.00 | 37.20 | 37.43 | 31.95 | 5.43 | 5.06 |
| PD2 | 713.00 | 680.90 | 592.26 | 507.12 | 117.43 | 173.77 | 101.00 | 122.50 | 40.80 | 59.70 | 31.90 | 41.80 | 37.00 | 36.50 | 32.26 | 31.57 | 4.56 | 4.84 |
| p-value | 0.509 | 0.814 | 0.757 | 0.663 | 0.154 | 0.129 | 0.898 | 0.254 | 0.309 | 0.400 | 0.007 | 0.421 | 0.191 | 0.291 | 0.194 | 0.447 | 0.143 | 0.016 |
| SEM | 7.31 | 15.51 | 6.00 | 18.44 | 1.50 | 2.83 | 2.52 | 3.11 | 2.14 | 7.36 | 0.05 | 9.42 | 1.86 | 3.37 | 1.62 | 3.20 | 0.19 | 0.55 |
| C1 | 710.40 | 688.50 | 593.72 | 499.36 | 112.94 | 187.52 | 99.20 | 117.50 | 40.00 | 60.30 | 31.20 | 43.10 | 40.00 | 36.40 | 34.78 | 31.38 | 5.02 | 4.89 |
| C2 | 708.50 | 677.90 | 593.22 | 504.09 | 115.80 | 173.84 | 103.40 | 120.00 | 41.20 | 60.20 | 32.10 | 41.80 | 40.00 | 37.20 | 34.91 | 32.15 | 4.97 | 5.02 |
| p-value | 0.628 | 0.011 | 0.895 | 0.251 | 0.015 | <0.001 | 0.056 | 0.212 | <0.001 | 0.866 | <0.001 | 0.012 | 0.955 | 0.188 | 0.881 | 0.181 | 0.697 | 0.194 |
| SEM | 3.89 | 3.90 | 3.75 | 40.41 | 1.09 | 1.89 | 2.10 | 1.88 | 1.51 | 2.24 | 0.35 | 4.96 | 1.02 | 6.38 | 0.88 | 5.64 | 0.13 | 0.95 |
| N1 | 711.50 | 682.90 | 589.59 | 493.17 | 119.25 | 190.18 | 98.10 | 111.80 | 38.90 | 59.10 | 31.20 | 41.70 | 40.20 | 33.90 | 34.99 | 29.10 | 5.08 | 4.67 |
| N2 | 707.50 | 683.50 | 597.35 | 510.28 | 109.50 | 171.17 | 104.40 | 125.70 | 42.30 | 61.40 | 32.00 | 43.10 | 39.80 | 39.80 | 34.70 | 34.43 | 4.91 | 5.24 |
| p-value | 0.316 | 0.878 | 0.049 | <0.001 | <0.001 | <0.001 | 0.006 | <0.001 | <0.001 | <0.001 | <0.001 | 0.009 | 0.669 | <0.001 | 0.743 | <0.001 | 0.214 | <0.001 |
| SEM | 3.89 | 3.90 | 3.75 | 40.41 | 1.09 | 1.89 | 2.10 | 1.88 | 1.51 | 2.24 | 0.35 | 4.96 | 1.02 | 6.38 | 0.88 | 5.64 | 0.13 | 0.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahangirlou, M.R.; Akbari, G.A.; Alahdadi, I.; Soufizadeh, S.; Parsons, D. Grain Quality of Maize Cultivars as a Function of Planting Dates, Irrigation and Nitrogen Stress: A Case Study from Semiarid Conditions of Iran. Agriculture 2021, 11, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010011
Jahangirlou MR, Akbari GA, Alahdadi I, Soufizadeh S, Parsons D. Grain Quality of Maize Cultivars as a Function of Planting Dates, Irrigation and Nitrogen Stress: A Case Study from Semiarid Conditions of Iran. Agriculture. 2021; 11(1):11. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010011
Chicago/Turabian StyleJahangirlou, Maryam Rahimi, Gholam Abbas Akbari, Iraj Alahdadi, Saeid Soufizadeh, and David Parsons. 2021. "Grain Quality of Maize Cultivars as a Function of Planting Dates, Irrigation and Nitrogen Stress: A Case Study from Semiarid Conditions of Iran" Agriculture 11, no. 1: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010011