
Functional Characteristics and Molecular Identification of Interspecific Hybrids from Genus Capsicum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
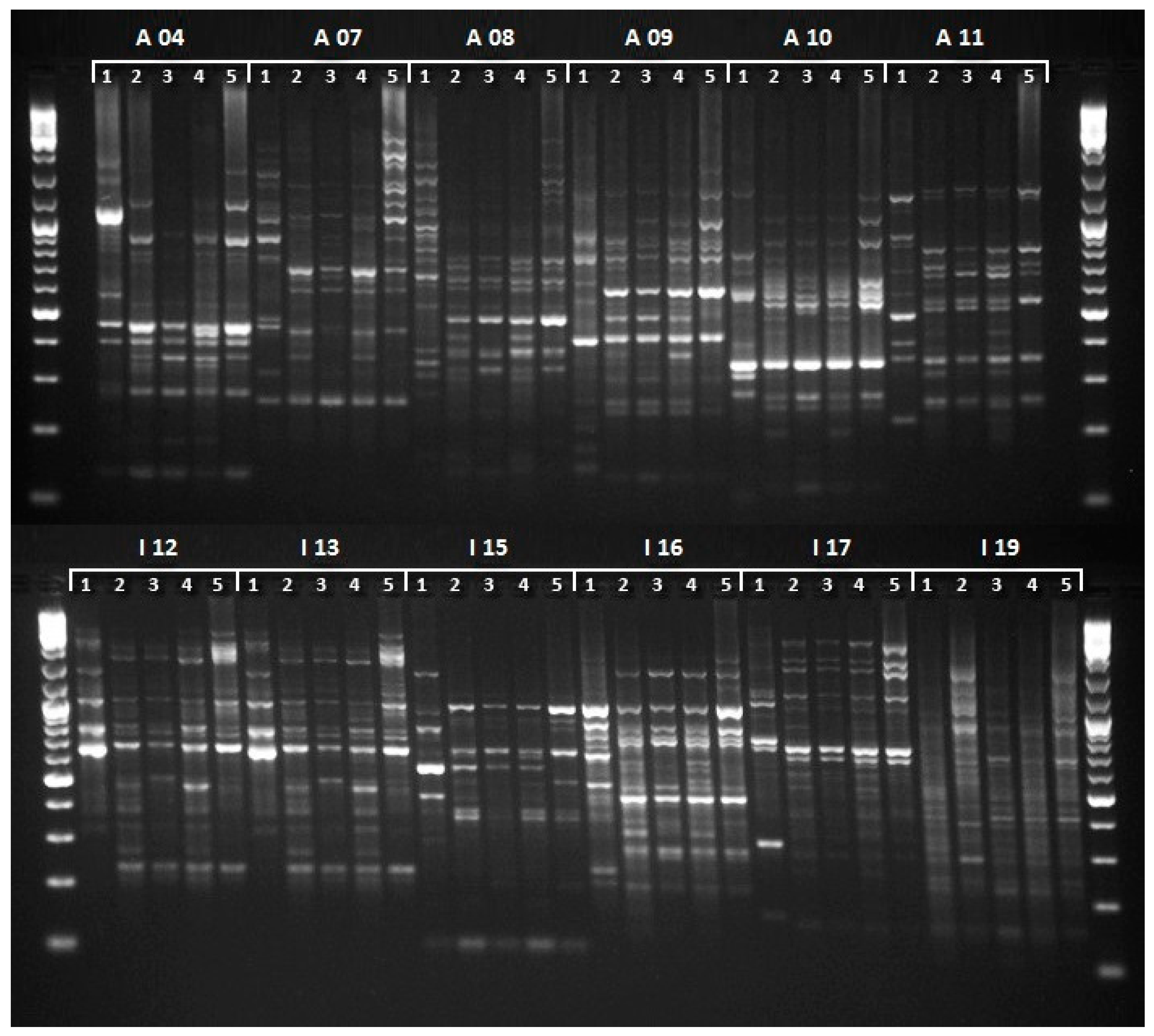
2.2. Molecular Analysis
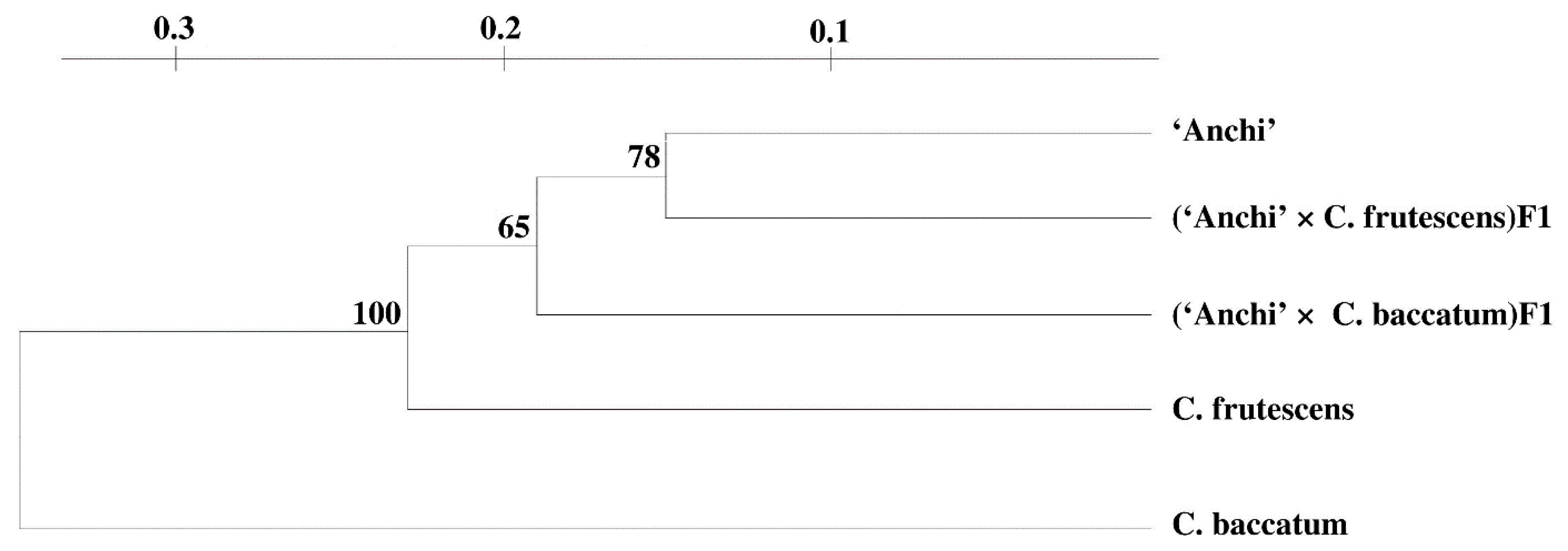
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ibiza, V.P.; Blanca, J.; Cañizares, J.; Nuez, F. Taxonomy and Genetic Diversity of Domesticated Capsicum Species in the Andean Region. Genet. Resour. Crop Evol. 2012, 59, 1077–1088. [Google Scholar] [CrossRef]
- Wang, D.; Bosland, P.W. The genes of Capsicum. Hortic. Sci. 2006, 41, 1169–1187. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.R.; Tikunov, Y.; de Vos, R.C.; Pelgrom, K.T.; Maharijaya, A.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolomics and molecular marker analysis to explore pepper (Capsicum sp.) biodiversity. Metabolomics 2013, 9, 130–144. [Google Scholar] [CrossRef] [Green Version]
- Bento, C.S.; Rodrigues, R.; Gonçalves, L.S.; Oliveira, H.S.; Santos, M.H.; Pontes, M.C.; Sudré, C.P. Inheritance of resistance to Pepper Yellow Mosaic Virus in Capsicum baccatum var. pendulum. Genet. Mol. Res. 2013, 12, 1074–1082. [Google Scholar] [CrossRef]
- Mongkolporn, O.; Taylor, P.W.J. Wild Crop Relatives: Genomic and Breeding Resources; Springer: New York, NY, USA, 2011; Volume 5, pp. 43–57. [Google Scholar]
- Mahasuk, P.; Chinthaisong, J.; Mongkolporn, O. Differential resistances to anthracnose in Capsicum baccatum as responding to two Colletotrichum pathotypes and inoculation methods. Breed. Sci. 2013, 63, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.B.; Park, H.G. Trispecies bridge crosses, (Capsicum annuum × C. chinense) × C. baccatum, as an alternative for introgression of anthracnose resistance from C. baccatum into C. annuum. Hortic. Environ. Biotechnol. 2005, 46, 5–9. [Google Scholar]
- Soler, S.; Debreczeni, D.E.; Vidal, E.; Aramburu, J.; López, C.; Galipienso, L.; Rubio, L. A new Capsicum baccatum accession shows tolerance to wild-type and resistance-breaking isolates of Tomato Spotted Wilt Virus. Ann. Appl. Biol. 2015, 167, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.V.; Lopes, R.; Lopes, M.T.G.; de Figueiredo, A.F.; Barros, W.S.; Alves, S.R.M. Cross Compatibility of Domesticated Hot Pepper and Cultivated Sweet Pepper. Crop Breed Appl. Biotechnol. 2009, 9, 37–44. [Google Scholar] [CrossRef]
- Monteiro, C.E.S.; Pereira de Campos, K. Reproductive characterization of interspecific hybrids among Capsicum species. Crop Breed Appl. Biotechnol. 2011, 11, 241–249. [Google Scholar] [CrossRef]
- Yoon, J.B.; Yang, D.C.; Do, J.W.; Park, H.G. Overcoming two post-fertilization genetic barriers in interspecific hybridization between Capsicum annuum and C. baccatum for introgression of anthracnose resistance. Breed. Sci. 2006, 56, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Martins, K.C.; Pereira, T.N.S.; Souza, S.A.M.; Rodrigues, R.; Amaral Junior, A.T.D. Crossability and Evaluation of Incompatibility Barriers in Crosses between Capsicum Species. Crop Breed Appl. Biotechnol. 2015, 15, 139–145. [Google Scholar] [CrossRef]
- Olszewska, D.; Niklas-Nowak, A.; Nowaczyk, P. Estimation of genetic divergence within androgenic regenerants of (Capsicum annuum L. ATZ1 × C. frutescens)F1 plants using Random Amplified Polymorphic DNA markers. BioTechnologia 2017, 3, 175–182. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.R.; Pereira, T.N.S.; Vitoria, A.P.; Campos, K.P.; Rodrugues, R.; Silva, D.H.; Pereira, M.G. Genetic diversity among Capsicum accessions using RAPD markers. Crop Breed Appl. Biotechnol. 2006, 6, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Orenthung, N.; Sapu, C. RAPD marker assisted study on genetic diversity of Indigenous Chilli (Capsicum sp.) landraces of Nagaland, India. Int. J. Bio-resour. Stress Manag. 2013, 4, 009–013. [Google Scholar]
- Shi, Y.H.; Hu, N.B. Acquisition, identification and analysis of an interspecific Capsicum hybrid (C. annuum x C. chinense). J. Hortic. Sci. Biotechnol. 2015, 90, 31–38. [Google Scholar]
- Ilbi, H. RAPD markers assisted varietal identification and genetic purity test in pepper, Capsicum annuum. Sci. Hortic. 2003, 97, 211–218. [Google Scholar] [CrossRef]
- Bhadragoudar, M.R.; Patil, C.G. Assessment of genetic diversity among Capsicum annuum L. Genotypes using RAPD markers. Afr. J. Biotechnol. 2011, 10, 17477–17483. [Google Scholar]
- Tilahun, S.; Paramaguru, P.; Bapu, J.R.K. Genetic diversity in certain genotypes of chilli and paprika as revelated by RAPD and SSR analysis. Asian J. Agric. Sci. 2013, 5, 25–31. [Google Scholar]
- Ahmed, M.S. Inter-simple sequence repeat (ISSR) markers in the evaluation of genetic polymorphism of Egyptian Capsicum L. hybrid. Afr. J. Biotechnol. 2013, 12, 665–669. [Google Scholar]
- Ou, L.; Zou, X. Inter simple sequence repeat analysis of genetic diversity of five cultivated pepper species. Afr. J. Biotechnol. 2012, 11, 752–757. [Google Scholar]
- Thul, S.T.; Darokar, M.P.; Shasany, A.K.; Khanuja, S.P.S. Molecular profiling for genetic variability in Capsicum species based on ISSR and RAPD markers. Mol. Biotechnol. 2012, 51, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Shapturenko, M.N.; Tarutina, L.A.; Mishin, L.A.; Kilchevsky, A.V.; Khotyleva, L.V. DNA divergence as a criterion of a sweet pepper (Capsicum annuum L.) selection for heterosis. Russ. J. Genet. 2014, 50, 123–130. [Google Scholar] [CrossRef]
- Sikora, B.; Nowaczyk, P. Application of RAPD technique for identification of interspecific hybrids from genus Capsicum. Acta Sci. Pol. Hortorum Cultus 2014, 13, 155–166. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Fruit Number per Plant Mean ± Standard Deviation | Fruit Weight per Plant (g) Mean ± Standard Deviation | Weight of Fruit without Stalk (g) Mean ± Standard Deviation |
| C. baccatum | 33.6 ± 4.9 b* | 344.0 ± 53.1 a | 11.19 ± 1.33 a |
| (Anchi × C. bac.)F1 | 15.4 ± 2.7 a | 1198.0 ± 108.1 c | 91.45 ± 6.92 c |
| ‘Anchi’ | 18.8 ± 3.5 a | 1705.1 ± 171.0 d | 111.57 ± 15.17 c |
| (Anchi × C. frut.)F1 | 14.6 ± 4.2 a | 822.1 ± 110.8 b | 65.06 ± 17.77 b |
| C. frutescens | 13.0 ± 2.5 a | 436.6 ± 71.7 a | 34.93 ± 7.47 ab |
| p-value | 0.00 | 0.00 | 0.00 |
| Genotype | Fruit Length (mm) Mean ± Standard Deviation | Fruit Width (mm) Mean ± Standard Deviation | Technological Mass (g) Mean ± Standard Deviation |
| C. baccatum | 93.78 ± 7.58 ab | 16.99 ± 1.81 a | 9.73 ± 1.47 a |
| (Anchi × C. bac.)F1 | 113.62 ± 13.94 c | 54.83 ± 7.09 b | 77.62 ± 6.53 bc |
| ‘Anchi’ | 141.23 ± 7.15 d | 57.11 ± 5.76 b | 99.15 ± 15.59 c |
| (Anchi × C. frut.)F1 | 106.81 ± 12.16 c | 47.23 ± 7.13 b | 55.99 ± 15.59 b |
| C. frutescens | 75.56 ± 10.02 a | 45.38 ± 6.91 b | 24.49 ± 13.01 a |
| p-value | 0.00 | 0.00 | 0.00 |
| Genotype | Weight of Placenta with Seeds (g) Mean ± Standard Deviation | Technological Efficiency Mean ± Standard Deviation | Wall Thickness (mm) Mean ± Standard Deviation |
| C. baccatum | 0.95 ± 0.21 a | 0.88 ± 0.01 b | 1.72 ± 0.17 a |
| (Anchi × C. bac.)F1 | 13.48 ± 3.65 c | 0.87 ± 0.01 b | 4.74 ± 0.46 c |
| ‘Anchi’ | 12.10 ± 0.63 bc | 0.89 ± 0.02 b | 5.18 ± 0.45 c |
| (Anchi × C. frut.)F1 | 8.49 ± 3.10 b | 0.85 ± 0.01 b | 3.22 ± 0.36 b |
| C. frutescens | 7.35 ± 1.39 b | 0.78 ± 0.04 a | 3.18 ± 0.52 b |
| p-value | 0.00 | 0.00 | 0.00 |
| Genotype | Weight of Seeds per Fruit (g) Mean ± Standard Deviation | Soluble Solids Mean ± Standard Deviation | Dry Matter (g) Mean ±Standard Deviation |
| C. baccatum | 1.05 ± 0.18 a | 9.55 ± 0.63 b | 13.13 ± 0.95 b |
| (Anchi × C. bac.)F1 | 2.82 ± 0.61 b | 8.94 ± 0.54 b | 11.33 ± 1.26 b |
| ‘Anchi’ | 2.29 ± 0.32 b | 6.80 ± 0.27 a | 8.59 ± 0.47 a |
| (Anchi × C. frut.)F1 | 2.30 ± 0.51 b | 9.18 ± 0.73 b | 12.17 ± 0.73 b |
| C. frutescens | 2.52 ± 0.36 b | 8.79 ± 1.56 b | 12.93 ± 2.24 b |
| p-value | 0.00 | 0.00 | 0.00 |
| Primer | Sequence | Number of Products | Range of Product Size | Polymorphic Products | % of Polymorphism |
|---|---|---|---|---|---|
| A 03 | AGTCAGCCAC | 7 | 288–1254 bp | 6 | 85.7 |
| A 04 | AATCGGGCTG | 13 | 135–1899 bp | 10 | 76.9 |
| A 05 | AGGGGTCTTG | 8 | 116–1002 bp | 6 | 75.0 |
| A 06 | GGTCCCTGAC | 7 | 167–1382 bp | 5 | 71.4 |
| A 07 | GAAACGGGTG | 13 | 234–2396 bp | 11 | 84.6 |
| A 08 | GTGACGTAGG | 16 | 253–2386 bp | 11 | 68.8 |
| A 09 | GGGTAACGCC | 14 | 132–1774 bp | 10 | 71.4 |
| A 10 | GTGATCGCAG | 9 | 123–1421 bp | 6 | 66.7 |
| A 11 | CAATCGCCGT | 14 | 240–1487 bp | 9 | 64.3 |
| A 12 | TCGGCGATAG | 8 | 249–1540 bp | 4 | 50.0 |
| A 13 | CAGCACCCAC | 6 | 186–1018 bp | 3 | 50.0 |
| A 14 | TCTGTGCTGG | 10 | 251–1295 bp | 6 | 60.0 |
| A 15 | TTCCGAACCC | 11 | 182–1827 bp | 7 | 63.6 |
| A 16 | AGCCAGCGAA | 13 | 173–2581 bp | 9 | 69.2 |
| A 17 | GACCGCTTGT | 8 | 204–2391 bp | 4 | 50.0 |
| A 18 | AGGTGACCGT | 13 | 112–1256 bp | 8 | 61.5 |
| A 19 | CAAACGTCGG | 9 | 196–1688 bp | 5 | 55.5 |
| AB 09 | GGGCGACTAC | 7 | 265–1995 bp | 4 | 57.1 |
| AE 02 | GGTGCGGGAA | 11 | 191–2394 bp | 8 | 72.7 |
| AE 10 | CTGAAGCGCA | 11 | 212–2154 bp | 6 | 54.5 |
| AE 11 | AAGAACGGGA | 12 | 106–2040 bp | 8 | 66.7 |
| AE 19 | ACGGCGTATG | 12 | 295–2482 bp | 9 | 76.9 |
| AF 05 | CCGAATTCCC | 9 | 224–2591 bp | 6 | 66.7 |
| C 15 | GACGGATCAG | 13 | 196–2118 bp | 8 | 61.5 |
| D 12 | CACCGTATCC | 14 | 265–1106 bp | 8 | 57.1 |
| P 13 | CGAGTGCCTC | 12 | 167–1840 bp | 5 | 41.7 |
| Total | 26 RAPD primers | 280 | 106–2591 bp | 182 | 65.0 |
| Primer | Sequence | Number of Products | Range of Product Size | Polymorphic Products | % of Polymorphism |
|---|---|---|---|---|---|
| I 1 | (GA)8YC | 5 | 221–562 bp | 3 | 60.0 |
| I 10 | (GA)8YT | 6 | 203–896 bp | 5 | 83.3 |
| I 11 | (CT)8GC | 4 | 508–843 bp | 3 | 75.0 |
| I 12 | (GACA)4A | 14 | 126–2116 bp | 8 | 57.1 |
| I 13 | (AG)8YC | 12 | 176–1740 bp | 5 | 41.7 |
| I 14 | (AG)8T | 5 | 217–777 bp | 3 | 60.0 |
| I 15 | (CTC)4YC | 13 | 219–2013 bp | 8 | 61.5 |
| I 16 | (AG)8C | 13 | 232–2561 bp | 9 | 69.2 |
| I 17 | (TGAG)4 | 8 | 234–2591 bp | 4 | 50.0 |
| I 19 | (GAG)3GG | 9 | 156–1681 bp | 5 | 55.5 |
| I 21 | (AC)8CT | 10 | 273–963 bp | 6 | 60.0 |
| I 25 | (CA)8AGT | 7 | 494–1385 bp | 4 | 57.1 |
| I 26 | (CA)8AT | 6 | 345–832 bp | 3 | 50.0 |
| I 27 | (GA)8CT | 5 | 189–989 bp | 2 | 40.0 |
| I 28 | (CA)8G | 6 | 225–536 bp | 5 | 83.3 |
| I 33 | (GA)8CTC | 11 | 228–1197 bp | 8 | 72.7 |
| I 40 | (GGAGA)4 | 6 | 219–1263 bp | 2 | 33.3 |
| I 46 | (GA)8T | 8 | 274–1438 bp | 6 | 75.0 |
| I 50 | (ATG)6 | 6 | 251–962 bp | 3 | 50.0 |
| I 56 | (GA)8G | 7 | 180–1832 bp | 4 | 57.1 |
| I 63 | (CCCT)4 | 9 | 396–1122 bp | 6 | 66.7 |
| I 65 | (GA)8C | 6 | 123–964 bp | 3 | 50.0 |
| I 71 | (GT)8TG | 12 | 285–739 bp | 4 | 33.3 |
| I 73 | (GT)8CG | 7 | 255–1473 bp | 6 | 85.7 |
| Total | 24 ISSR primers | 195 | 123–1832 bp | 115 | 59.0 |
| Genotypes | F* | B | AB | A | AF |
|---|---|---|---|---|---|
| F—C. frutescens | × | ||||
| B—C. baccatum | 0.717 | × | |||
| AB—(‘Anchi’ × C. baccatum)F1 | 0.537 | 0.737 | × | ||
| A–‘Anchi’ | 0.476 | 0.755 | 0.482 | × | |
| AF—(‘Anchi’ × C. frutescens)F1 | 0.452 | 0.718 | 0.381 | 0.439 | × |
| Genotypes | F* | B | AB | A | AF |
|---|---|---|---|---|---|
| F—C. frutescens | × | ||||
| B—C. baccatum | 0.489 | × | |||
| AB—(‘Anchi’ × C. baccatum)F1 | 0.431 | 0.514 | × | ||
| A—‘Anchi’ | 0.337 | 0.578 | 0.292 | × | |
| AF—(‘Anchi’ × C. frutescens)F1 | 0.281 | 0.510 | 0.283 | 0.222 | × |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olszewska, D.; Tomaszewska-Sowa, M.; Witkowska, E.; Litewka, J. Functional Characteristics and Molecular Identification of Interspecific Hybrids from Genus Capsicum. Agriculture 2021, 11, 1198. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121198
Olszewska D, Tomaszewska-Sowa M, Witkowska E, Litewka J. Functional Characteristics and Molecular Identification of Interspecific Hybrids from Genus Capsicum. Agriculture. 2021; 11(12):1198. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121198
Chicago/Turabian StyleOlszewska, Dorota, Magdalena Tomaszewska-Sowa, Emilia Witkowska, and Jakub Litewka. 2021. "Functional Characteristics and Molecular Identification of Interspecific Hybrids from Genus Capsicum" Agriculture 11, no. 12: 1198. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121198