
Repeated Application of Rice Straw Stabilizes Soil Bacterial Community Composition and Inhibits Clubroot Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Measurement of the Plant Quality and Disease Incidence
2.4. Soil Sampling and Chemical Properties Analysis
2.5. DNA Extraction and Quantitative Polymerase Chain Reaction (PCR) Analysis
2.6. Illumina MiSeq Sequencing and Data Processing
2.7. Statistical Analysis
3. Results and Discussion
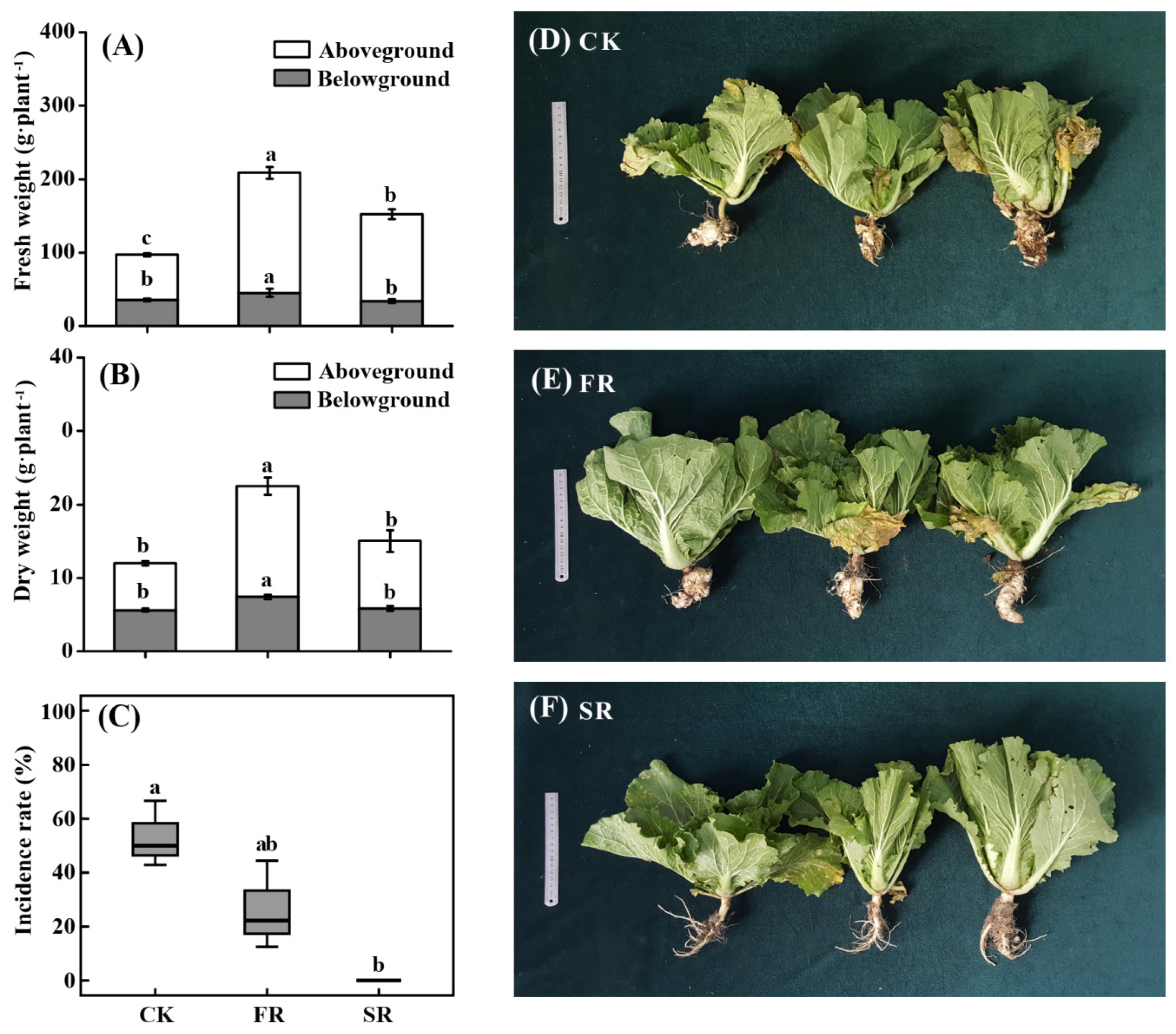
3.1. The Plant Growth Responses to the Addition of Rice Straw
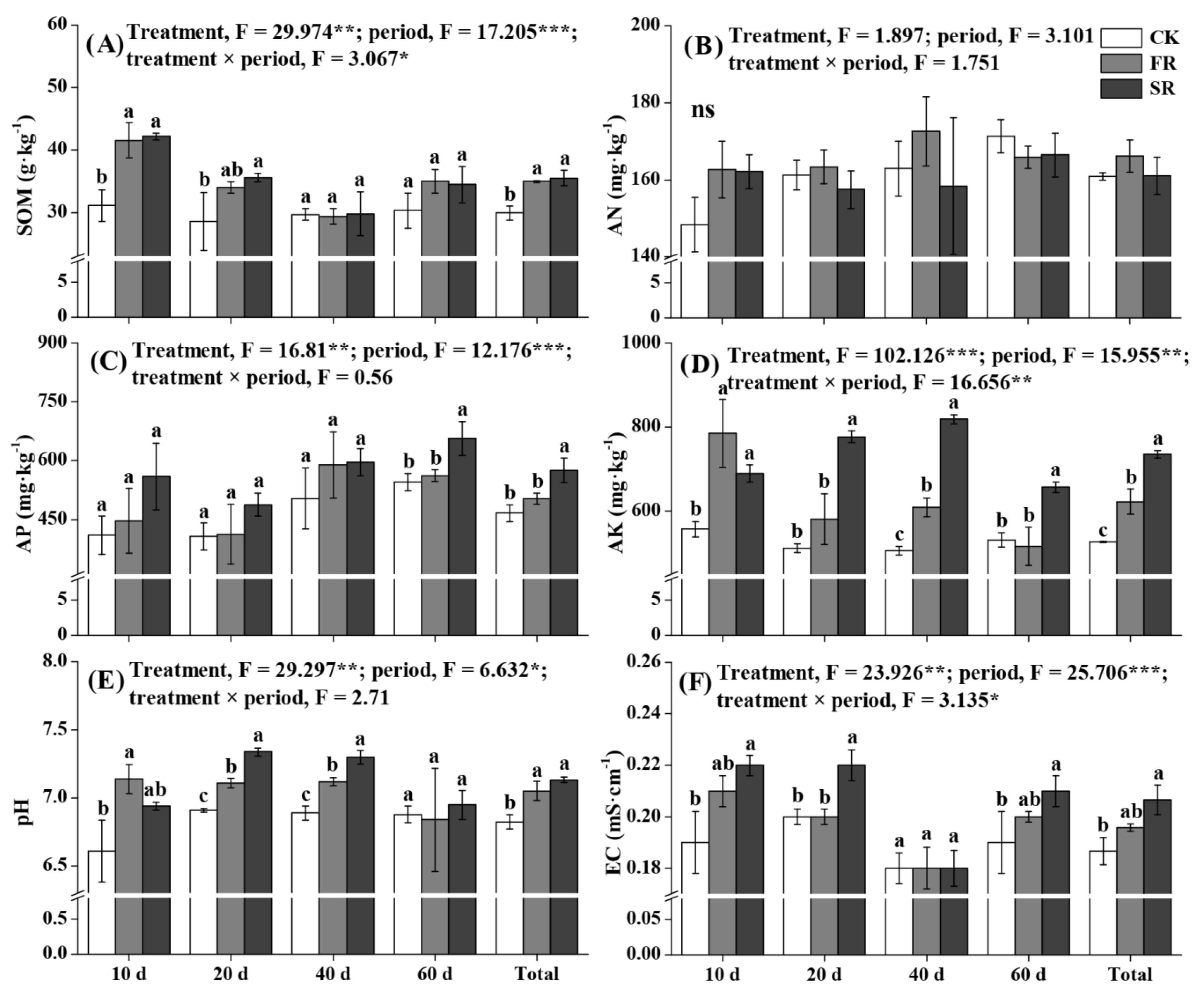
3.2. The Soil Chemical Properties’ Response to the Addition of Rice Straw
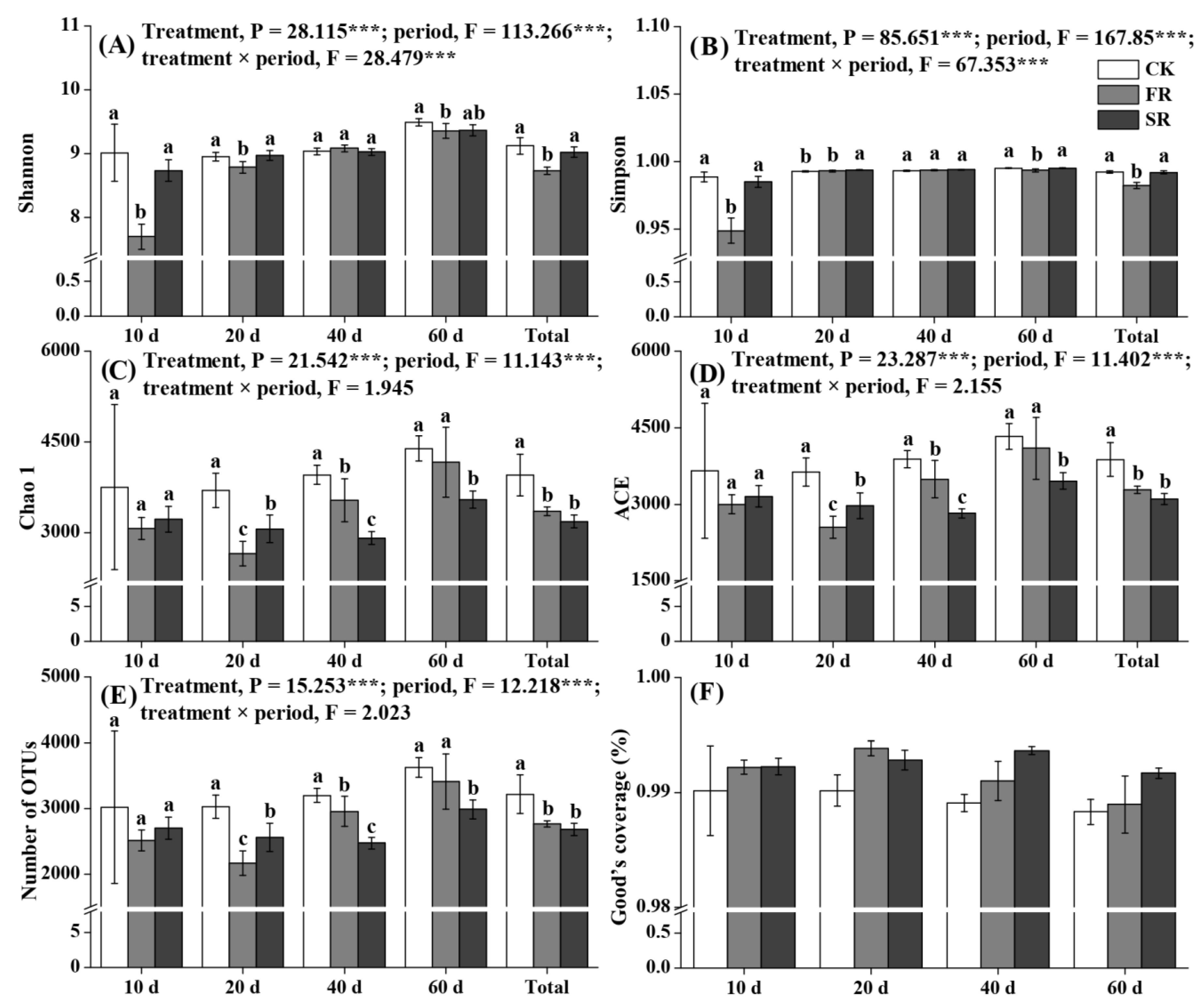
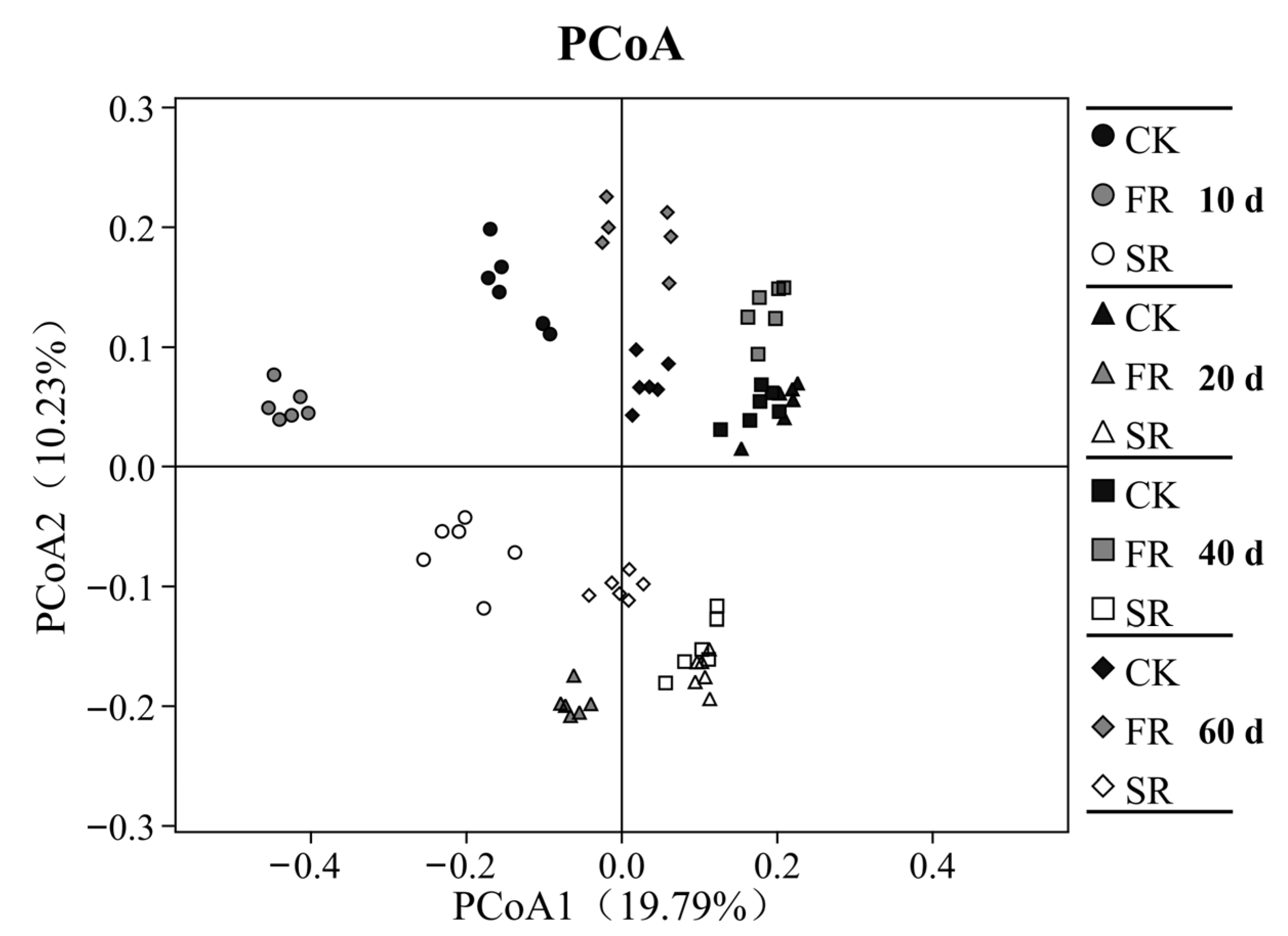
3.3. Modulation of Bacterial Community Diversity in Soils
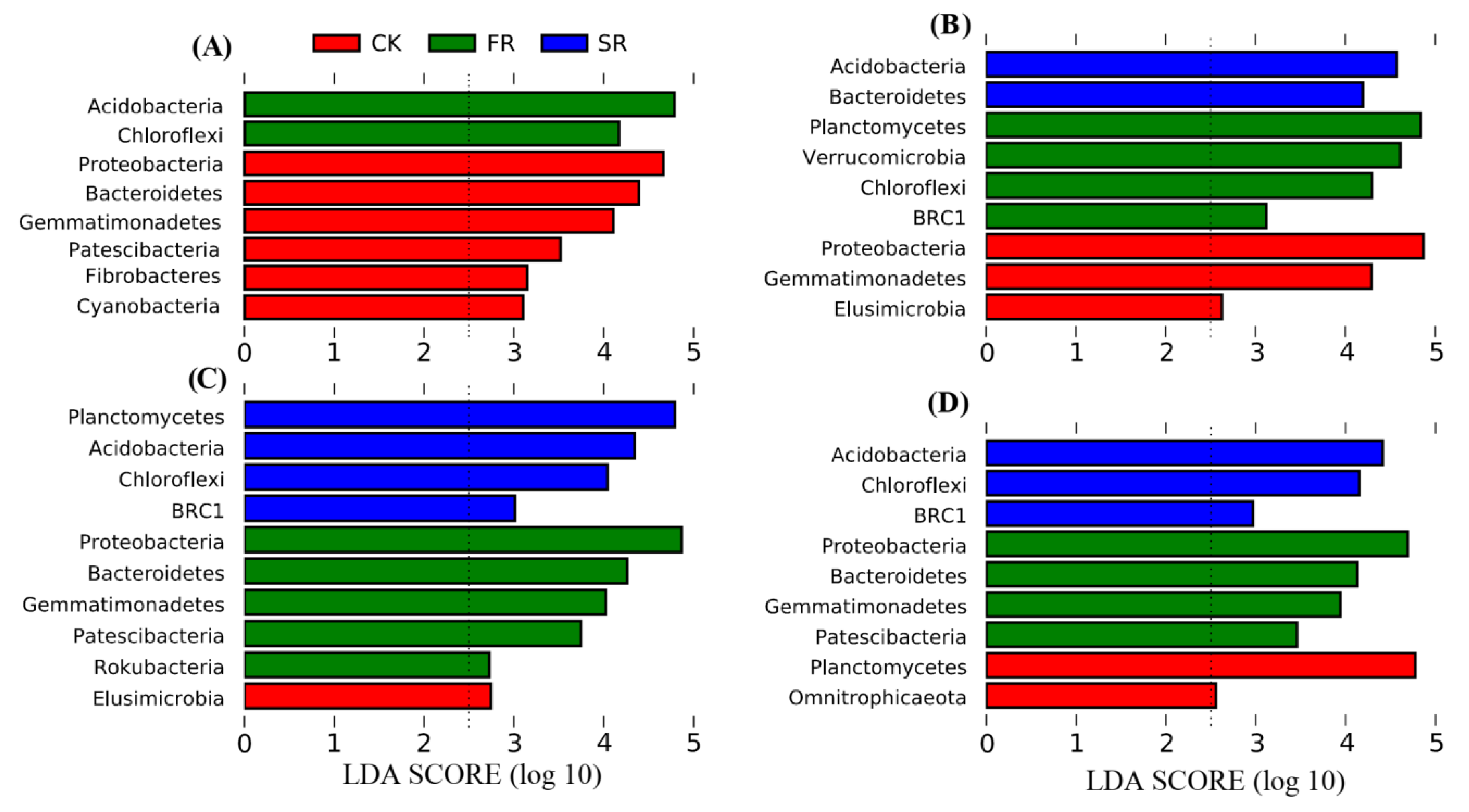
3.4. Response of Bacterial Phyla Composition to the Addition of Rice Straw
3.5. Response of the Bacterial Genera Composition to the Addition of Rice Straw
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hwang, S.F.; Strelkov, S.E.; Feng, J.; Gossen, B.D.; Howard, R.J. Plasmodiophora brassicae: A review of an emerging pathogen of the Canadian canola (Brassica napus) crop. Mol. Plant Pathol. 2012, 13, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Chai, A.L.; Xie, X.W.; Shi, Y.X.; Li, B.J. Research status of clubroot (Plasmodiophora brassicae) on cruciferous crops in China. Can. J. Plant Pathol. 2014, 36, 142–153. [Google Scholar] [CrossRef]
- Donald, C.; Porter, I. Integrated Control of Clubroot. J. Plant Growth Regul. 2009, 28, 289–303. [Google Scholar] [CrossRef]
- Sharma, K.; Gossen, B.; McDonald, M.R. Effect of temperature on primary infection by Plasmodiophora brassicae. Plant Pathol. 2011, 60, 830–838. [Google Scholar] [CrossRef]
- Samuel, G.; Garrett, S.D. The infected root-hair count for estimating the activity of Plasmodiophora brassicae Woron. in the soil. Ann. Appl. Biol. 2010, 32, 96–101. [Google Scholar] [CrossRef]
- Dixon, G.R. The Occurrence and Economic Impact of Plasmodiophora brassicae and Clubroot Disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Ahmed, H.U.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; Peng, G.; Howard, R.J.; Turnbull, G.D. Assessment of bait crops to reduce inoculum of clubroot (Plasmodiophora brassicae) of canola. Can. J. Plant Sci. 2011, 91, 545–551. [Google Scholar] [CrossRef]
- Hwang, S.F.; Ahmed, H.U.; Zhou, Q.; Turnbull, G.D.; Strelkov, S.E.; Gossen, B.D.; Peng, G. Effect of host and non-host crops on Plasmodiophora brassicae resting spore concentrations and clubroot of canola. Plant Pathol. 2015, 64, 1198–1206. [Google Scholar] [CrossRef]
- Murakami, H.; Tsushima, S.; Shishido, Y. Soil suppressiveness to clubroot disease of Chinese cabbage caused by Plasmodiophora brassicae. Soil Biol. Biochem. 2000, 32, 1637–1642. [Google Scholar] [CrossRef]
- Ney, L.; Franklin, D.; Mahmud, K.; Cabrera, M.; Fatzinger, B. Rebuilding Soil Ecosystems for Improved Productivity in Biosolarized Soils. Int. J. Agron. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Hwang, S.F.; Howard, R.J.; Strelkov, S.E.; Gossen, B.D.; Peng, G. Management of clubroot (Plasmodiophora brassicae) on canola (Brassica napus) in western Canada. Can. J. Plant Pathol. 2014, 36, 49–65. [Google Scholar] [CrossRef]
- Narisawa, K.; Tokumasu, S.; Hashiba, T. Suppression of clubroot formation in Chinese cabbage by the root endophytic fungus, Heteroconium chaetospira. Plant Pathol. 2010, 47, 206–210. [Google Scholar] [CrossRef]
- Mahmud, K.; Franklin, D.; Ney, L.; Cabrera, M.; Habteselassie, M.; Hancock, D.; Newcomer, Q.; Subedi, A.; Dahal, S. Improving inorganic nitrogen in soil and nutrient density of edamame bean in three consecutive summers by utilizing a locally sourced bio-inocula. Org. Agric. 2021. [Google Scholar] [CrossRef]
- Peng, G.; Lahlali, R.; Hwang, S.-F.; Pageau, D.; Hynes, R.K.; McDonald, M.R.; Gossen, B.D.; Strelkov, S.E. Crop rotation, cultivar resistance, and fungicides/biofungicides for managing clubroot (Plasmodiophora brassicae) on canola. Can. J. Plant Pathol. 2014, 36, 99–112. [Google Scholar] [CrossRef]
- Yang, X.X.; Huang, X.Q.; Wen-Xian, W.U.; Xiang, Y.J.; Liu, Y. Effects of different rotation patterns on the occurrence of clubroot disease and diversity of rhizosphere microbes. J. Integr. Agric. 2020, 19, 2265–2273. [Google Scholar] [CrossRef]
- Howard, R.J.; Strelkov, S.E.; Harding, M.W. Clubroot of cruciferous crops—New perspectives on an old disease. Can. J. Plant Pathol. 2010, 32, 43–57. [Google Scholar] [CrossRef]
- Rifai, S.W.; Markewitz, D.; Borders, B. Twenty years of intensive fertilization and competing vegetation suppression in loblolly pine plantations: Impacts on soil C, N, and microbial biomass. Soil Biol. Biochem. 2010, 42, 713–723. [Google Scholar] [CrossRef]
- Chen, S.C.; Zhou, X.G.; Yu, H.J.; Wu, F.Z. Root exudates of potato onion are involved in the suppression of clubroot in a Chinese cabbage-potato onion-Chinese cabbage crop rotation. Eur. J. Plant Pathol. 2018, 150, 765–777. [Google Scholar] [CrossRef]
- Liu, C.; Yang, Z.; He, P.; Munir, S.; Wu, Y.; Ho, H.; He, Y. Deciphering the bacterial and fungal communities in clubroot-affected cabbage rhizosphere treated with Bacillus Subtilis XF-1. Agric. Ecosyst. Environ. 2018, 256, 12–22. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An Examination of the Degtjareff Method for Determining Soil Organic Matter, and A Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Li, N.; Gao, D.; Zhou, X.; Chen, S.; Wu, F. Intercropping with Potato-Onion Enhanced the Soil Microbial Diversity of Tomato. Microorganisms 2020, 8, 834. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Xia, Y.; Fan, C.; Kou, J.; Wu, F.; Li, W.; Pan, K. Control of Fusarium wilt by wheat straw is associated with microbial network changes in watermelon rhizosphere. Sci. Rep. 2020, 10, 12736. [Google Scholar] [CrossRef] [PubMed]
- Sundelin, T.; Christensen, C.B.; Larsen, J.; Mller, K.; Lübeck, M.; Bdker, L.; Jensen, B. In Planta Quantification of Plasmodiophora brassicae Using Signature Fatty Acids and Real-Time PCR. Plant Dis. Int. J. Appl. Plant Pathol. 2010, 94, 432–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowther, T.W.; Maynard, D.S.; Leff, J.W.; Oldfield, E.E.; Mcculley, R.L.; Fierer, N.; Bradford, M.A. Predicting the responsiveness of soil biodiversity to deforestation: A cross-biome study. Glob. Change Biol. 2014, 20, 2983–2994. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Gloeckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Nie, S.; Li, W.; Fan, C.; Wang, S.; Wu, F.; Pan, K. Wheat straw increases the defense response and resistance of watermelon monoculture to Fusarium wilt. BMC Plant Biol. 2019, 19, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.Y.; Fu, X.P.; Zhou, X.G.; Wu, F.Z.; Liu, S.W. Effects of plant residues on cucumber (Cucumis sativus L) growth, soil enzyme activities and microbial communities. Allelopath. J. 2016, 38, 147–157. [Google Scholar]
- Zhou, X.; Wu, F. Changes in soil chemical characters and enzyme activities during continuous monocropping of cucumber (Cucumis sativus). Pak. J. Bot. 2015, 47, 691–697. [Google Scholar]
- Zhang, Y.; Li, Z.L.; Wang, S.W.; Wu, F.Z.; Pan, K. Effects of crop straws on root knot nematodes and soil fungal community in continuous cropping of tomato. Allelopath. J. 2017, 42, 79–92. [Google Scholar] [CrossRef]
- Hwang, S.F.; Ahmed, H.U.; Strelkov, S.E.; Gossen, B.D.; Turnbull, G.D.; Peng, G.; Howard, R.J. Seedling age and inoculum density affect clubroot severity and seed yield in canola. Can. J. Plant Sci. 2011, 91, 183–190. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, L.; Lin, Q.; Yuan, M.; Xu, D.; Yu, H.; Hu, Y.; Duan, J.; Li, X.; He, Z.; et al. Responses of the functional structure of soil microbial community to livestock grazing in the Tibetan alpine grassland. Glob. Change Biol. 2013, 19, 637–648. [Google Scholar] [CrossRef]
- Ye, R.; Doane, T.A.; Morris, J.; Horwath, W.R. The effect of rice straw on the priming of soil organic matter and methane production in peat soils. Soil Biol. Biochem. 2015, 81, 98–107. [Google Scholar] [CrossRef]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Kleinman, P.J.A.; Pimentel, D.; Bryant, R.B. The ecological sustainability of slash-and-burn agriculture. Agric. Ecosyst. Environ. 1995, 52, 235–249. [Google Scholar] [CrossRef]
- Mourtzinis, S.; Gaska, J.; Diallo, T.; Lauer, J.G.; Conley, S. Legacy Effects of Three Management Practices on Corn, Soybean, and Wheat Yield. Agron. J. 2018, 110, 418–424. [Google Scholar] [CrossRef]
- Dendooven, L.; Alcántara-Hernández, R.J.; Valenzuela-Encinas, C.; Luna-Guido, M.; Perez-Guevara, F.; Marsch, R. Dynamics of carbon and nitrogen in an extreme alkaline saline soil: A review. Soil Biol. Biochem. 2010, 42, 865–877. [Google Scholar] [CrossRef]
- Cheng, W.; Padre, A.T.; Shiono, H.; Sato, C.; Nguyen-Sy, T.; Tawaraya, K.; Kumagai, K. Changes in the pH, EC, available P, SOC and TN stocks in a single rice paddy after long-term application of inorganic fertilizers and organic matters in a cold temperate region of Japan. J. Soils Sediments 2017, 17, 1834–1842. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, P.; Wang, K.; Ding, R.; Yang, B.; Nie, J.; Jia, Z.; Han, Q. Effects of Wheat Straw Incorporation on the Availability of Soil Nutrients and Enzyme Activities in Semiarid Areas. PLoS ONE 2015, 10, e0120994. [Google Scholar] [CrossRef] [Green Version]
- Niwa, R.; Kumei, T.; Nomura, Y.; Yoshida, S.; Osaki, M.; Ezawa, T. Increase in soil pH due to Ca-rich organic matter application causes suppression of the clubroot disease of crucifers. Soil Biol. Biochem. 2007, 39, 778–785. [Google Scholar] [CrossRef] [Green Version]
- Green, S.J.; Prakash, O.; Jasrotia, P.; Overholt, W.A.; Cardenas, E.; Hubbard, D.; Tiedje, J.M.; Watson, D.B.; Schadt, C.W.; Brooks, S.C. Denitrifying bacteria from the genus Rhodanobacter dominate bacterial communities in the highly contaminated subsurface of a nuclear legacy waste site. Appl. Environ. Microbiol. 2012, 78, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Lv, J.L.; Yu, M.; Ma, Z.H.; Xi, H.; Kou, C.L.; He, Z.C.; Shen, A.L. Long-term decomposed straw return positively affects the soil microbial community. J. Appl. Microbiol. 2019, 128, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Liu, Y.F.; Zhang, G.X.; Guo, X.O.; Sun, Z.P.; Li, T.L. The Effects of Rice Straw and Biochar Applications on the Microbial Community in a Soil with a History of Continuous Tomato Planting History. Agronomy Basel 2018, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef]
- Ludwig-Mueller, J.; Prinsen, E.; Rolfe, S.A.; Scholes, J.D. Metabolism and Plant Hormone Action During Clubroot Disease. J. Plant Growth Regul. 2009, 28, 229–244. [Google Scholar] [CrossRef]
- Saraiva, A.L.d.R.F.; Bhering, A.d.S.; do Carmo, M.G.F.; Andreote, F.D.; Dias, A.C.F.; Coelho, I.d.S. Bacterial composition in brassica-cultivated soils with low and high severity of clubroot. J. Phytopathol. 2020, 168, 613–619. [Google Scholar] [CrossRef]
- Vida, C.; de Vicente, A.; Cazorla, F.M. The role of organic amendments to soil for crop protection: Induction of suppression of soilborne pathogens. Ann. Appl. Biol. 2020, 176, 1–15. [Google Scholar] [CrossRef]
- Lee, S.-H.; Ka, J.-O.; Cho, J.-C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2008, 285, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyashita, N.T. Contrasting soil bacterial community structure between the phyla Acidobacteria and Proteobacteria in tropical Southeast Asian and temperate Japanese forests. Genes Genet. Syst. 2015, 90, 61–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadnikov, V.V.; Mardanov, A.V.; Beletsky, A.V.; Rakitin, A.L.; Frank, Y.A.; Karnachuk, O.V.; Ravin, N.V. Phylogeny and physiology of candidate phylum BRC1 inferred from the first complete metagenome-assembled genome obtained from deep subsurface aquifer. Syst. Appl. Microbiol. 2019, 42, 67–76. [Google Scholar] [CrossRef]
- Larsbrink, J.; McKee, L.S. Bacteroidetes bacteria in the soil: Glycan acquisition, enzyme secretion, and gliding motility. In Advances in Applied Microbiology; Gadd, G.M., Sariaslani, S., Eds.; Academic Press: Cambridge, MA, USA, 2020; Chapter 2; Volume 110, pp. 63–98. [Google Scholar]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global Biogeography and Quantitative Seasonal Dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Naumoff, D.G.; Dedysh, S.N. Bacteria from Poorly Studied Phyla as a Potential Source of New Enzymes: β-Galactosidases from Planctomycetes and Verrucomicrobia. Microbiology 2018, 87, 796–805. [Google Scholar] [CrossRef]
- Jifu, L.; Jianwei, L.; Xiaokun, L.; Tao, R.; Rihuan, C.; Li, Z. Dynamics of Potassium Release and Adsorption on Rice Straw Residue. PLoS ONE 2014, 9, e90440. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Buenger, W.; Burbano, C.S.; Sabale, M.; Hurek, T. Roots Shaping Their Microbiome: Global Hotspots for Microbial Activity. Annu. Rev. Phytopathol. 2015, 53, 403–424. [Google Scholar] [CrossRef]
- Espana, M.; Rasche, F.; Kandeler, E.; Brune, T.; Rodriguez, B.; Bending, G.D.; Cadisch, G. Identification of active bacteria involved in decomposition of complex maize and soybean residues in a tropical Vertisol using N-15-DNA stable isotope probing. Pedobiologia 2011, 54, 187–193. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, S.X.; Fang, S.; Tian, Y. Contrasting decomposition rates and nutrient release patterns in mixed vs singular species litter in agroforestry systems. J. Soils Sediments 2014, 14, 1071–1081. [Google Scholar] [CrossRef]
- Tang, H.M.; Xiao, X.P.; Tang, W.G.; Lin, Y.C.; Wang, K.; Yang, G.L.; Hauke, S. Effects of Winter Cover Crops Residue Returning on Soil Enzyme Activities and Soil Microbial Community in Double-Cropping Rice Fields. PLoS ONE 2014, 9, e100443. [Google Scholar] [CrossRef]
- Li, J.; Philp, J.; Li, J.; Wei, Y.; Wang, Y. Trichoderma harzianum Inoculation Reduces the Incidence of Clubroot Disease in Chinese Cabbage by Regulating the Rhizosphere Microbial Community. Microorganisms 2020, 8, 1325. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Hu, X.M.; Deng, W.; Li, Y.; Xiong, C.; Ye, H.; Han, G.M.; Li, X. Changes in soil microbial community structure and functional diversity in the rhizosphere surrounding mulberry subjected to long-term fertilization. Appl. Soil Ecol. 2015, 86, 30–40. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Hu, S.; Bruggen, A.H.C.V. Short-term cover crop decomposition in organic and conventional soils: Characterization of soil C, N, microbial and plant pathogen dynamics. Eur. J. Plant Pathol. 2000, 106, 37–50. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Z.; Liu, X.; Zhang, X.H. Microbial assembly, interaction, functioning, activity and diversification: A review derived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Z.; Di, C.; Rahman, M.K.u.; Gao, D.; Wu, F.; Pan, K. Repeated Application of Rice Straw Stabilizes Soil Bacterial Community Composition and Inhibits Clubroot Disease. Agriculture 2021, 11, 108. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020108
Han Z, Di C, Rahman MKu, Gao D, Wu F, Pan K. Repeated Application of Rice Straw Stabilizes Soil Bacterial Community Composition and Inhibits Clubroot Disease. Agriculture. 2021; 11(2):108. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020108
Chicago/Turabian StyleHan, Zhe, Chengqian Di, Muhammad Khashi u Rahman, Danmei Gao, Fengzhi Wu, and Kai Pan. 2021. "Repeated Application of Rice Straw Stabilizes Soil Bacterial Community Composition and Inhibits Clubroot Disease" Agriculture 11, no. 2: 108. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020108