
Genetic Progress Achieved during 10 Years of Selective Breeding for Honeybee Traits of Interest to the Beekeeping Industry


Abstract
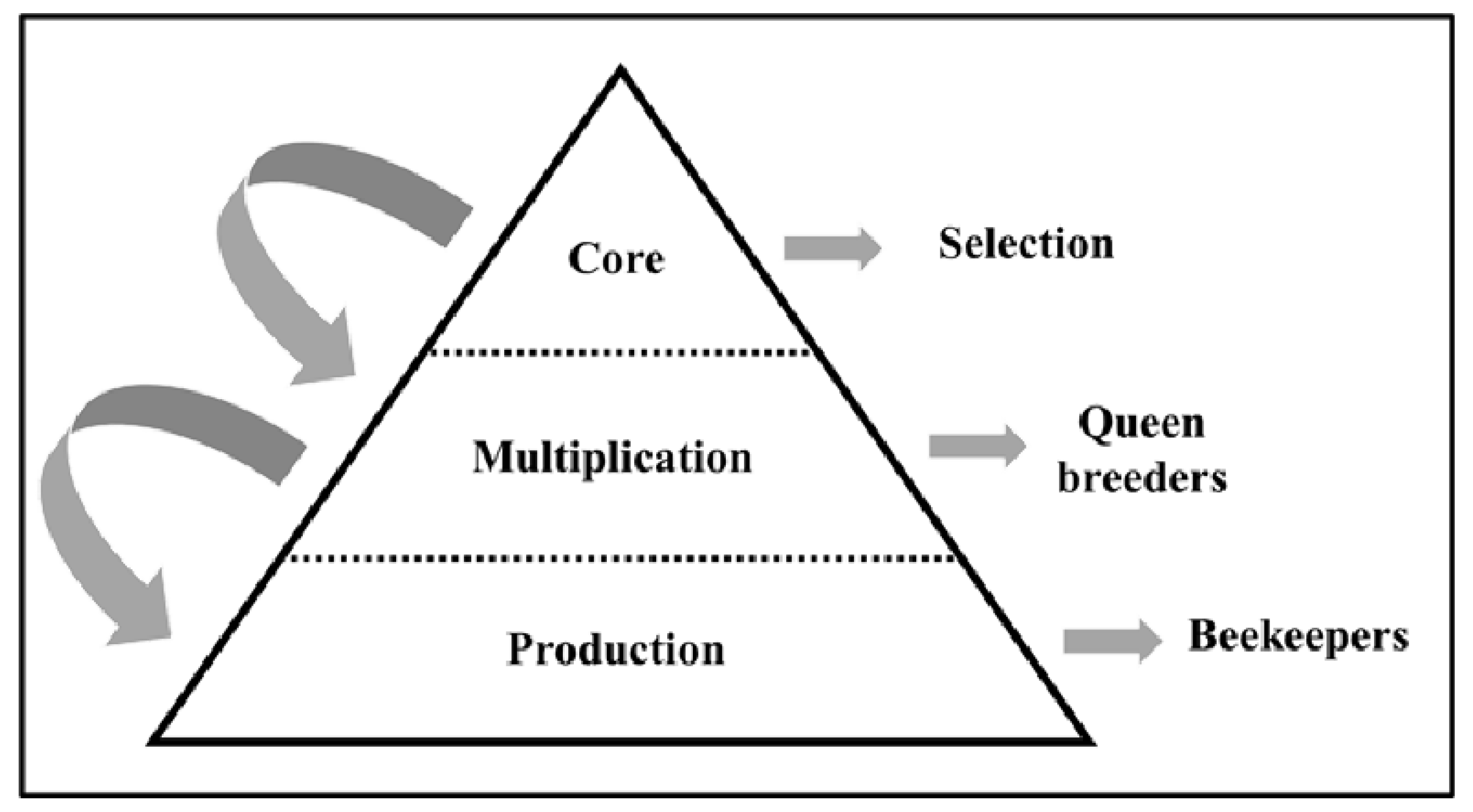
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Traits Measured in Colonies
2.2.1. Hygienic Behavior
2.2.2. Honey Production
2.2.3. Spring Development
2.3. Selection from Breeding Value and Selection Index
2.3.1. CRSAD Breeding Program Selection Plan Launched in 2010
2.3.2. Rearing and Mating Process
2.3.3. Guaranteed Selected Genetics
2.3.4. Colony Management
2.3.5. Statistical Analysis
2.3.6. Choice of Breeders
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Denis, B. La domestication: Un concept devenu pluriel. INRA Prod. Anim. 2004, 17, 161–166. [Google Scholar] [CrossRef]
- Harpur, B.A.; Minaei, S.; Kent, C.F.; Zayed, A. Management increases genetic diversity of honey bees via admixture. Mol. Ecol. 2012, 21, 4414–4421. [Google Scholar] [CrossRef]
- Sellier, P.; Boichard, D.; Verrier, E. La génétique animale à l’inra. Hist. Rech. Contemp. 2019, 8, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Verrier, E.; Jussiau, R.; Le Roy, P.; Gastinel, P.L.; Ducos, A.; Journaux, L.; Lagarrigue, S.; Mambrini, M.; Mattalia, S.; Montmeas, L.; et al. Training for animal breeding: Old questions and new challenges. Renc. Rech. Rumin. 2005, 12, 41–47. [Google Scholar]
- Kerr, R.J.; Hammond, K.; Kinghorn, B.P. Effects of multiple sire matings on genetic evaluations, selection response and rates of inbreeding. Livest. Prod. Sci. 1994, 38, 161–168. [Google Scholar] [CrossRef]
- Henderson, C. Unbiased methods for G and R unknown. In Applications of Linear Models in Animal Breeding, 1st ed.; University of Guelph: Guelph, ON, Canada, 1984. [Google Scholar]
- Cauia, E.; Siceanu, A.; Patruica, S.; Bura, M.; Sapcaliu, A.; Magdici, M. A standardized data base for honeybee colonies evaluation, with application in honeybee breeding programs. Econ. Manag. Financ. Mark. 2011, 6, 1146–1151. [Google Scholar]
- Page, R.E.; Laidlaw, H.H. Honey bee genetics and breeding. In Hive and the Honey Bee, 3rd ed.; Graham, J., Ed.; Dadant & Sons: Hamilton, IL, USA, 2008; pp. 235–267. [Google Scholar]
- Sauvager, B. Hérédité chez l’Abeille et les Colonies d’Abeilles, 1st ed.; Anercea, au Cœur de L’élevage: Surgères, France, 2019; pp. 28–30. [Google Scholar]
- Chevalet, C.; Cornuet, J.M. Evolution de la consanguinité dans une population d’abeilles. Apidologie 1982, 13, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Willam, A. Parameter- und Zuchtwertschätzung für die Honigbiene (Apis mellifera carnica). Ph.D. Thesis, University of Agricultural Sciences, Vienna, Austria, 1991. [Google Scholar]
- Cobey, S.; Sheppard, W.S.; Tarpy, D.R. Status of breeding practices and genetic diversity in domestic, U.S. honey bees. In Honey Bee Colony Health: Challenges and Sustainable Solutions, 1st ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 39–53. [Google Scholar]
- Bienefeld, K.; Reinhardt, F.; Pirchner, F. Inbreeding effects of queen in the honey bee colony traits. Apidologie 1989, 20, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Smith, S. Estimating quantitative genetic parameters in haplodiploid organisms. Heredity 2000, 85, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Bienefeld, K. Breeding success or genetic diversity in honey bees? Bee World 2016, 93, 40–44. [Google Scholar] [CrossRef]
- Plate, M.; Bernstein, R.; Hoppe, A.; Bienefeld, K. Long-term evaluation of breeding scheme alternatives for endangered honeybee subspecies. Insects 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Bienefeld, K.; Ehrhardt, K.; Reinhardt, F. Genetic evaluation in the honey bee considering queen and worker effects—A BLUP-animal model approach. Apidologie 2007, 38, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Oxley, P.R.; Oldroyd, B.P. The genetic architecture of honeybee breeding. Adv. Insect Physiol. 2010, 39, 83–118. [Google Scholar]
- Jamieson, C.A. Facts about beekeeping in Canada. Bee World 1958, 39, 232–236. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee, 1st ed.; Harvard University Press: Cambridge, MA, USA, 1987; pp. 250–254. [Google Scholar]
- Canadian Association of Professional Apiculturist. Available online: https://capabees.com/shared/2012/10/2012capawintloss1.pdf (accessed on 29 April 2021).
- Canadian Honey Council. Honey promotions through partnerships. Hivelights 2015, 28, 6–8. [Google Scholar]
- Parker, R.; Melathopoulos, A.P.; White, R.; Pernal, S.F.; Guarna, M.M.; Foster, L.J. Ecological adaptation of diverse honey bee (Apis mellifera) populations. PLoS ONE 2010, 5, e11096. [Google Scholar] [CrossRef]
- Currie, R.; Pernal, S.F.; Guzman-Novoa, E. Honey bee colony losses in Canada. J. Apic. Res. 2010, 49, 104–106. [Google Scholar] [CrossRef]
- Canadian Association of Professional Apiculturists. Available online: https://capabees.com/shared/CAPA-Statement-on-Colony-Losses-2020.pdf (accessed on 29 April 2021).
- Kevan, P.G.; Guzman, E.; Skinner, A.; van Engelsdorp, D. Colony collapse disorder in Canada: Do we have a problem? Hivelights 2007, 1, 14–16. [Google Scholar]
- Canadian Association of Professional Apiculturists. Available online: https://www.agrireseau.net/apiculture/documents/93309/rapport-sur-les-mortalites-hivernales-des-colonies-d_abeilles-au-canada-acpa-2016?a=1&r=Association+canadienne+des+professionnels+de+l%E2%80%99apiculture+Rapport+sur+la+mortalit%C3%A9+hivernale+de+colonies+d%E2%80%99abeilles+au+Canada+%282016%29 (accessed on 29 April 2021).
- Bixby, M.A. Canadian beekeeping perspective on colony health and growing our local queen supply. Hivelights 2017, 30, 13–15. [Google Scholar]
- Bixby, M.; Mcafee, A. Queens rule! Growing Canada’s queens breeding industry: Results from the 2016–2017 queen breeder survey. Am. Bee J. 2017, 157, 893–895. [Google Scholar]
- Meixner, M.D.; Buchler, R.; Costa, C.; Francis, R.M.; Hatjina, F.; Kryger, P.; Uzunov, A.; Carreck, N. Honey bee genotypes and the environment. J. Apic. Res. 2014, 53, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Seeley, T.D.; Tarpy, D.R. Queen promiscuity lowers disease within honeybee colonies. Proc. R. Soc. B 2007, 274, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Leveaux, G.; van Laere, O. Beebreed. Eu programme de sélection. Abeille Cie. 2014, 2, 33–34. [Google Scholar]
- Hoppe, A.; Du, R.; Bernstein, R.; Tiesler, F.-K.; Kacher, M.; Bienefeld, K. Substantial genetic progress in the international Apis mellifera carnica population since the implementation of genetic evaluation. Insects 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Buckfast Denmark. Available online: https://buckfast.dk/index.php/en/ (accessed on 7 June 2021).
- Fédération des Apiculteurs. Available online: https://www.apiculteursduquebec.com/documents/Plan_strategique.pdf (accessed on 29 April 2021).
- Programme Canadien D’adaptation Agricole. Available online: https://docplayer.fr/193446657-Pcaa-programme-canadien-d-adaptation-agricole.html (accessed on 29 April 2021).
- Maucourt, S.; Fortin, F.; Robert, C.; Giovenazzo, P. Genetic parameters of honey bee colonies traits in a Canadian selection program. Insects 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Spivak, M.; Downey, D.L. Field assays for hygienic behavior in honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1998, 91, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Spivak, M.; Reuter, G.S. Performance of hygienic honey bee colonies in a commercial apiary. Apidologie 1998, 29, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Guarna, M.; Hoover, S.; Huxter, E.; Higo, H.; Moon, K.-M.; Domanski, D.; Bixby, M.; Melathopoulos, A.; Ibrahim, A.; Peirson, M.; et al. Peptide biomarkers used for the selective breeding of a complex polygenic trait in honey bees. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovenazzo, P.; Dubreuil, P. Evaluation of spring organic treatments against Varroa destructor (Acari: Varroidae) in honey bee Apis mellifera (Hymenoptera: Apidae) colonies in eastern Canada. Exp. Appl. Acarol. 2011, 55, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Delaplane, K.S.; van der Steen, J.; Guzman-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Chevalet, C.; Cornuet, J. Etude théorique sur la selection du caractère “production de miel” chez l’abeille II. Plan de sélection combinée de reines en fécondation naturelle. Apidologie 1987, 18, 253–266. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman, P., Ed.; Longman: Burnt Mill, UK, 1996; pp. 135–165. [Google Scholar]
- Minvielle, F. Que Sais-Je? La Selection Animale, 1st ed.; Les Presses Universitaires de France: Paris, France, 1998; pp. 16–24. [Google Scholar]
- Mrode, R.A. Linear Models for the Prediction of Animal Breeding Values, 1st ed.; CABI Publishing: Oxford, England, 2005; pp. 60–95. [Google Scholar]
- Brascamp, E.; Bijma, P. Methods to estimate breeding values in honey bees. Genet. Sel. Evol. 2014, 46, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Connor, L.J. Bee Sex Essentials; Wicwas Press: Kalamazoo, MI, USA, 2008; pp. 152–153. [Google Scholar]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J.; Hatjina, F. Standard methods for rearing and selection of Apis mellifera queens. J. Apic. Res. 2013, 52, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Laidlaw, H.H.; Page, R.E. Queen Rearing and Bee Breeding, 1st ed.; Wicwas Press: Cheshire, CT, USA, 1997; pp. 135–145. [Google Scholar]
- Sorel, A.; Martin, G.; Houle, E.; Giovenazzo, P. Finding DCA’s. Available online: https://www.beeculture.com/finding-dcas/ (accessed on 29 April 2021).
- Hellmich, R.L.; Collins, A.M.; Danka, R.G.; Rinderer, T.E. Influencing matings of European honey bees in areas with Africanized honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1988, 81, 796–799. [Google Scholar] [CrossRef]
- Hellmich, R.L.; Waller, G.D. Preparing for africanized honey bees: Evaluating control in mating apiaries. Am. Bee J. 1990, 130, 537–542. [Google Scholar]
- Guzman-Novoa, E. Elemental Genetics and Breeding for the Honeybee, 1st ed.; Association, O.B., Ed.; Guelph University Press: Bayfield, ON, Canada, 2007; pp. 47–49. [Google Scholar]
- Plate, M.; Bernstein, R.; Hoppe, A.; Bienefeld, K. The importance of controlled mating in honeybee breeding. Genet. Sel. Evol. 2019, 51, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrose, J.T. Management for honey production. In Hive and the Honey Bee, 3rd ed.; Graham, J., Ed.; Dadant & Sons: Hamilton, IL, USA, 2008; Volume 1, pp. 602–654. [Google Scholar]
- Smartbees. Available online: https://www.smartbees-fp7.eu/resources/Publications/2016/FRE_SMARTBEES-Protocol-for-performance-testing_2016.pdf (accessed on 29 April 2021).
- Baker, R.J. Selection indexes without economic weights for animal breeding. Can. J. Anim. Sci. 1974, 54, 1–8. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Otis, G.W. Application of a modified selection index for honey bess (Hymenoptera: Apidae). J. Econ. Entomol. 2000, 93, 1606–1612. [Google Scholar] [CrossRef] [PubMed]
- Sammatero, D.; Gerson, U.; Needham, G. Parasitic mites of honey bees: Life, history, implications, and impact. Annu. Rev. Entomol. 2000, 45, 519–548. [Google Scholar] [CrossRef]
- Statistique Canada. Available online: https://www.agr.gc.ca/resources/prod/doc/pdf/honey_2016-eng.pdf (accessed on 29 April 2021).
- Canadian Honey Council. Available online: http://honeycouncil.ca/wp-content/uploads/2016/12/BHRT-RoD-January-22-2019-FINAL-FR.pdf (accessed on 29 April 2021).
- Bienefeld, K.; Ehrhardt, K.; Reinhardt, F. Bee breeding around the world—Noticeable success in honey bee selection after the introduction of genetic evaluation using BLUP. Am. Bee J. 2008, 8, 739–742. [Google Scholar]
- Cobey, S.; Lawrence, T. A successful application of the Page/Laidlaw breeding program. Glean. Bee Cult. 1988, 274–276. [Google Scholar]
- De Rochambeau, H. Les bases de la génétique quantitative: Le progrès génétique et sa réalisation dans les expériences de sélection. In Elément de Génétique Quantitative et Application aux Populations Animales; special ed.; INRA Production Animales: Paris, France, 1992; Volume 1, pp. 83–86. [Google Scholar]
- Hallais, J. Available online: https://docplayer.fr/16360791-Bases-de-genetique-et-de-selection-animale.html (accessed on 29 April 2021).
- Wiener, G.; Rouvier, R. L’amélioration Génétique Animale, 1st ed.; Presses Agronomique de Gembloux: Gembloux, France, 2009; Volume 1, pp. 186–190. [Google Scholar]
- Tedeschi, L.O. Assessment of the adequacy of mathematical models. Agric. Syst. 2006, 89, 225–247. [Google Scholar] [CrossRef]
- Néron, F.; Guéguen, R. Petit Précis d’Elevage, 1st ed.; Agricole, F., Ed.; Agriproduction: Paris, France, 2018; pp. 310–443. [Google Scholar]
- Westell, R.A.; Quaas, R.L.; van Vleck, L.D. Genetic group in an animal model. J. Dairy Sci. 1988, 71, 1310–1318. [Google Scholar] [CrossRef]
- Perez-Enciso, M.; Fernando, R.L. Genetic evaluation with uncertain parentage: A comparison of methods. Theor. Appl. Genet. 1992, 84, 173–179. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation | Number of Colonies | Number of Queen-Producing Colonies for Breeding Next Generation | |
|---|---|---|---|
| Selection based on phenotypic measurements | 2010 | 26 | 7 |
| 2011 | 60 | 11 | |
| 2012 | 38 | 12 | |
| 2013 | 45 | 13 | |
| 2014 | 109 | 11 | |
| 2015 | 144 | 14 | |
| Selection with BLUP-animal model | 2016 | 97 | 9 |
| 2017 | 85 | 11 | |
| 2018 | 152 | 9 | |
| 2019 | 134 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maucourt, S.; Fortin, F.; Robert, C.; Giovenazzo, P. Genetic Progress Achieved during 10 Years of Selective Breeding for Honeybee Traits of Interest to the Beekeeping Industry. Agriculture 2021, 11, 535. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060535
Maucourt S, Fortin F, Robert C, Giovenazzo P. Genetic Progress Achieved during 10 Years of Selective Breeding for Honeybee Traits of Interest to the Beekeeping Industry. Agriculture. 2021; 11(6):535. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060535
Chicago/Turabian StyleMaucourt, Ségolène, Frédéric Fortin, Claude Robert, and Pierre Giovenazzo. 2021. "Genetic Progress Achieved during 10 Years of Selective Breeding for Honeybee Traits of Interest to the Beekeeping Industry" Agriculture 11, no. 6: 535. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060535