
Short-Term Abandonment versus Mowing in a Mediterranean-Temperate Meadow: Effects on Floristic Composition, Plant Functionality, and Soil Properties—A Case Study

 and
and
Abstract
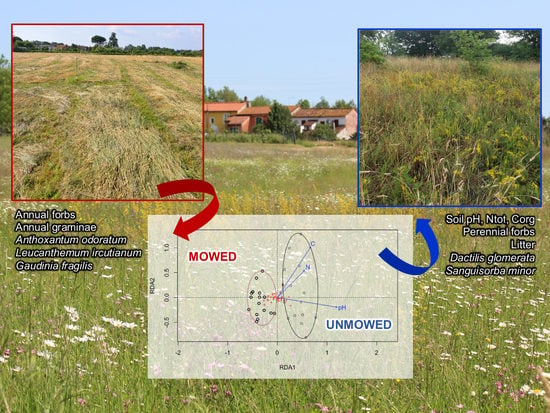
:
1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Data Collection
2.3. Soil Analysis
2.4. Vegetation Analysis
2.5. Statistical Analysis
3. Results
3.1. Plant Community Composition
3.2. Soil Properties and Plant Community
3.3. RDA
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Feurdean, A.; Ruprecht, E.; Molnár, Z.; Hutchinson, S.M.; Hickler, T. Biodiversity-rich European grasslands: Ancient, forgotten ecosystems. Biol. Conserv. 2018, 228, 224–232. [Google Scholar] [CrossRef]
- Pykälä, J.; Luoto, M.; Heikkinen, R.K.; Kontula, T. Plant species richness and persistence of rare plants in abandoned semi-natural grasslands in northern Europe. Basic Appl. Ecol. 2005, 6, 25–33. [Google Scholar] [CrossRef]
- Petermann, J.S.; Buzhdygan, O.Y. Grassland biodiversity. Curr. Biol. 2021, 31, R1195–R1201. [Google Scholar] [CrossRef] [PubMed]
- Tallowin; Jefferson Hay production from lowland semi-natural grasslands: A review of implications for ruminant livestock systems. Grass Forage Sci. 1999, 54, 99–115. [CrossRef]
- Öckinger, E.; Eriksson, A.K.; Smith, H.G. Effects of grassland abandonment, restoration and management on butterflies and vascular plants. Biol. Conserv. 2006, 133, 291–300. [Google Scholar] [CrossRef]
- Kuussaari, M.; Bommarco, R.; Heikkinen, R.K.; Helm, A.; Krauss, J.; Lindborg, R.; Öckinger, E.; Pärtel, M.; Pino, J.; Rodà, F.; et al. Extinction debt: A challenge for biodiversity conservation. Trends Ecol. Evol. 2009, 24, 564–571. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A. Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef] [Green Version]
- Haslgrübler, P.; Krautzer, B.; Tamegger, C.; Ševĉíková, M.; Tischew, S.; Rieger, E.; Kizekiva, M.; Golinski, P.; Scotton, M. SALVERE—Semi Natural Grassland as a Source of Biodiversity Improvement—A Central Europe Project. Grassl. Sci. Eur. 2011, 16, 526–528. [Google Scholar]
- Critchley, C.; Chambers, B.; Fowbert, J.; Sanderson, R.; Bhogal, A.; Rose, S. Association between lowland grassland plant communities and soil properties. Biol. Conserv. 2002, 105, 199–215. [Google Scholar] [CrossRef]
- Poeplau, C. Grassland soil organic carbon stocks along management intensity and warming gradients. Grass Forage Sci. 2021, 76, 186–195. [Google Scholar] [CrossRef]
- Chang, J.; Ciais, P.; Gasser, T.; Smith, P.; Herrero, M.; Havlík, P.; Obersteiner, M.; Guenet, B.; Goll, D.S.; Li, W.; et al. Climate warming from managed grasslands cancels the cooling effect of carbon sinks in sparsely grazed and natural grasslands. Nat. Commun. 2021, 12, 118. [Google Scholar] [CrossRef]
- Bretzel, F.; Vannucchi, F.; Romano, D.; Malorgio, F.; Benvenuti, S.; Pezzarossa, B. Wildflowers: From conserving biodiversity to urban greening—A review. Urban For. Urban Green. 2016, 20, 428–436. [Google Scholar] [CrossRef]
- Gaujour, E.; Amiaud, B.; Mignolet, C.; Plantureux, S. Factors and processes affecting plant biodiversity in permanent grasslands. A review. Agron. Sustain. Dev. 2012, 32, 133–160. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.-Y.; Pellet, J.; Buri, P.; Arlettaz, R. Does delaying the first mowing date benefit biodiversity in meadowland? Environ. Evid. 2012, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Kelemen, A.; Török, P.; Valkó, O.; Deák, B.; Miglécz, T.; Tóth, K.; Ölvedi, T.; Tóthmérész, B. Sustaining recovered grasslands is not likely without proper management: Vegetation changes after cessation of mowing. Biodivers. Conserv. 2014, 23, 741–751. [Google Scholar] [CrossRef]
- Pavlů, L.; Pavlů, V.; Gaisler, J.; Hejcman, M.; Mikulka, J. Effect of long-term cutting versus abandonment on the vegetation of a mountain hay meadow (Polygono-Trisetion) in Central Europe. Flora-Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 1020–1029. [Google Scholar] [CrossRef]
- Cachovanová, L.; Hájek, M.; Fajmonová, Z.; Marrs, R. Species Richness, Community Specialization and Soil-Vegetation Relationships of Managed Grasslands in a Geologically Heterogeneous Landscape. Folia Geobot. Phytotaxon. 2012, 47, 349–371. [Google Scholar] [CrossRef]
- Swacha, G.; Botta-Dukát, Z.; Kącki, Z.; Pruchniewicz, D.; Żołnierz, L. The effect of abandonment on vegetation composition and soil properties in Molinion meadows (SW Poland). PLoS ONE 2018, 13, e0197363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venterink, H.O.; Kardel, I.; Kotowski, W.; Peeters, W.; Wassen, M. Long-term effects of drainage and hay-removal on nutrient dynamics and limitation in the Biebrza mires, Poland. Biogeochemisty 2009, 93, 235–252. [Google Scholar] [CrossRef]
- Pavlů, L.; Pavlů, V.; Gaisler, J.; Hejcman, M. Relationship between soil and biomass chemical properties, herbage yield and sward height in cut and unmanaged mountain hay meadow (Polygono–Trisetion). Flora-Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 599–608. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes and Ecosystem Properties, 2nd ed.; John Wiley & Sons Ltd.: Sussex, UK, 2001. [Google Scholar]
- Tardella, F.M.; Di Agostino, V.M. Vegetation of the “Altipiani di Colfiorito” wetlands (central Apennines, Italy). Plant Sociol. 2020, 57, 113. [Google Scholar] [CrossRef]
- Faria, N.; Peco, B.; Carmona, C.P. Effects of haying on vegetation communities, taxonomic diversity and sward properties in mediterranean dry grasslands: A preliminary assessment. Agric. Ecosyst. Environ. 2018, 251, 48–58. [Google Scholar] [CrossRef]
- Louault, F.; Pillar, V.D.; Aufrere, J.; Garnier, E.; Soussana, J.F. Plant traits and functional types in response to reduced disturbance in a semi-natural grassland. J. Veg. Sci. 2005, 16, 151–160. [Google Scholar] [CrossRef]
- Jongen, M.; Pereira, J.; Aires, L.M.I.; Pio, C. The effects of drought and timing of precipitation on the inter-annual variation in ecosystem-atmosphere exchange in a Mediterranean grassland. Agric. For. Meteorol. 2011, 151, 595–606. [Google Scholar] [CrossRef]
- Willig, M.R. Biodiversity and Productivity. Science 2011, 333, 1709–1710. [Google Scholar] [CrossRef] [PubMed]
- Socher, S.A.; Prati, D.; Boch, S.; Müller, J.; Baumbach, H.; Gockel, S.; Hemp, A.; Schöning, I.; Wells, K.; Buscot, F.; et al. Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions. Basic Appl. Ecol. 2013, 14, 126–136. [Google Scholar] [CrossRef]
- Bartolini, A. Il Bosco di Chiusi e la Paduletta di Ramone. Indagini Naturalistiche e Attività di Ripristino Degli Habitat. Quaderni del Padule di Fucecchio n. 7; Centro di Ricerca, Documentazione e Promozione del Padule di Fucecchio; Università degli Studi di Pavia: Brescia, Italy, 2010. [Google Scholar]
- Blasi, C.; Michetti, L. The climate of Italy. In Biodiversity in Italy; Blasi, C., Boitani, L., la Posta, S., Manes, F., Marchetti, M., Eds.; Palombi Editori: Roma, Italy, 2007; pp. 57–66. Available online: http://www.pcn.minambiente.it/viewer/index.php?services=Fitoclima (accessed on 9 July 2021).
- ASA-SSSA. Methods of Soil Analysis, Part 1 and 3, Physical and Chemical Methods, 2nd ed.; ASA-SSSA: Madison, WI, USA, 1996. [Google Scholar]
- Duckworth, J.C.; Kent, M.; Ramsay, P.M. Plant functional types: An alternative to taxonomic plant community description in biogeography? Prog. Phys. Geogr. 2000, 24, 515–542. [Google Scholar] [CrossRef]
- Daubenmire, R.F. Canopy coverage method for vegetation analysis. Northwest Sci. 1959, 33, 43–64. [Google Scholar]
- Seabloom, E.W.; Borer, E.T.; Buckley, Y.; Cleland, E.E.; Davies, K.; Firn, J.; Harpole, W.S.; Hautier, Y.; Lind, E.; MacDougall, A.; et al. Predicting invasion in grassland ecosystems: Is exotic dominance the real embarrassment of richness? Glob. Chang. Biol. 2013, 19, 3677–3687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 728–814. [Google Scholar] [CrossRef] [Green Version]
- Allegrezza, M.; Biondi, E. Syntaxonomic revision of Arrhenatherum elatius grasslands of central Italy. Fitosociologia 2011, 48, 23–40. [Google Scholar]
- Foster, B.L.; Gross, K.L. Species richness in a successful grassland: Effects of nitrogen enrichment and plant litter. Ecology 1998, 79, 2593–2602. [Google Scholar] [CrossRef]
- Smart, S.M.; Thompson, K.; Marrs, R.H.; Le Duc, M.G.; Maskell, L.C.; Firbank, L. Biotic homogenization and changes in species diversity across human-modified ecosystems. Proc. R. Soc. B Boil. Sci. 2006, 273, 2659–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losvik, M.H. Plant species diversity in an old, traditionally managed hay meadow compared to abandoned hay meadows in southwest Norway. Nord. J. Bot. 1999, 19, 473–487. [Google Scholar] [CrossRef]
- Boob, M.; Elsaesser, M.; Thumm, U.; Hartung, J.; Lewandowski, I. Different management practices influence growth of small plants in species-rich hay meadows through shading. Appl. Veg. Sci. 2021, 24, e12625. [Google Scholar] [CrossRef]
- Galvánek, D.; Leps, J. The effect of management on productivity, litter accumulation and seedling recruitment in a Carpathian mountain grassland. Plant Ecol. 2012, 213, 523–533. [Google Scholar] [CrossRef]
- Klimešová, J.; Janeček, S.; Bartušková, A.; Lanta, V.; Doležal, J. How is regeneration of plants after mowing affected by shoot size in two specie-rich meadows. Folia Geobot. 2010, 45, 225–238. [Google Scholar] [CrossRef]
- Prévosto, B.; Kuiters, L.; Bernhardt-Römermann, M.; Dölle, M.; Schmidt, W.; Hoffmann, M.; van Uytvanck, J.; Bohner, A.; Kreiner, D.; Stadler, J.; et al. Impacts of Land Abandonment on Vegetation: Successional Pathways in European Habitats. Folia Geobot. 2011, 46, 303–325. [Google Scholar] [CrossRef] [Green Version]
- Grime, J.P.; Thompson, K.; Hunt, R.; Hodgson, J.G.; Cornelissen, J.H.C.; Rorison, I.H.; Hendry, G.A.F.; Ashenden, T.W.; Askew, A.P.; Band, S.R.; et al. Integrated Screening Validates Primary Axes of Specialisation in Plants. Oikos 1997, 79, 259. [Google Scholar] [CrossRef]
- White, T.A.; Moore, K.J.; Barker, D.J. The importance of local processes to landscape patterns of grassland vegetation diversity. N. Z. J. Agric. Res. 2004, 47, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Crawley, M.J.; Johnston, A.E.; Silvertown, J.; Dodd, M.; de Mazancourt, C.; Heard, M.S.; Henman, D.F.; Edwards, G.R. Determinants of Species Richness in the Park Grass Experiment. Am. Nat. 2005, 165, 179–192. [Google Scholar] [CrossRef] [PubMed]
- De Deyn, G.B.; Quirk, H.; Yi, Z.; Oakley, S.; Ostle, N.J.; Bardgett, R.D. Vegetation composition promotes carbon and nitrogen storage in model grassland communities of contrasting soil fertility. J. Ecol. 2009, 97, 864–875. [Google Scholar] [CrossRef]
- Diviaková, A.; Stašiov, S.; Pondelík, R.; Pätoprstý, V.; Novikmec, M. Environmental and Management Control over the Submontane Grassland Plant Communities in Central Slovakia. Diversity 2021, 13, 30. [Google Scholar] [CrossRef]
- Merunková, K.; Chytrý, M. Environmental control of species richness and composition in upland grasslands of the southern Czech Republic. Plant. Ecol. 2012, 213, 591–602. [Google Scholar] [CrossRef]
- Gabarrón-Galeote, M.A.; Trigalet, S.; van Wesemael, B. Effect of land abandonment on soil organic carbon fractions along a Mediterranean precipitation gradient. Geoderma 2015, 249–250, 69–78. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Functional Group | Site |
|---|---|---|---|
| Agrimonia eupatoria L. | Rosaceae | Perennial forb | AM |
| Agrostis stolonifera L. | Poaceae | Graminoid | AM |
| Anthoxanthum odoratum L. | Poaceae | Graminoid | C |
| Avena sterilis L. subsp. sterilis | Poaceae | Graminoid | C |
| Briza maxima L. | Poaceae | Graminoid | HM |
| Briza minor L. | Poaceae | Graminoid | C |
| Carex divulsa Stokes | Cyperaceae | Graminoid | C |
| Carex flacca Schreb. | Cyperaceae | Graminoid | HM |
| Carex hirta L. | Cyperaceae | Graminoid | C |
| Carex spicata Huds. | Cyperaceae | Graminoid | AM |
| Carex tomentosa L. | Cyperaceae | Graminoid | AM |
| Centaurea nigrescens Willd. | Asteraceae | Perennial forb | HM |
| Cichorium intybus L. | Asteraceae | Perennial forb | AM |
| Clinopodium nepeta (L.) Kuntze | Lamiaceae | Perennial forb | AM |
| Coleostephus myconis (L.) Cass. ex Rchb. f. | Asteraceae | Annual forb | C |
| Convolvolus arvensis L. | Convolvulaceae | Perennial forb | C |
| Convolvulus sepium L. | Convolvulaceae | Perennial forb | AM |
| Cynodon dactylon (L.) Pers | Poaceae | Graminoid | AM |
| Cyperus spp. | Cyperaceae | Graminoid | C |
| Dactylis glomerata L. | Poaceae | Graminoid | C |
| Danthonia decumbes (L.) DC. | Poaceae | Graminoid | HM |
| Daucus carota L. subsp. carota | Apiaceae | Annual forb | C |
| Dianthus armeria L. | Caryophyllaceae | Perennial forb | AM |
| Filipendula vulgaris Moench. | Rosaceae | Perennial forb | AM |
| Galium verum L. | Rubiaceae | Perennial forb | C |
| Gaudinia fragilis (L.) P. Beauv. | Poaceae | Graminoid | HM |
| Geraniumdissectum L. | Geraniaceae | Annual forb | C |
| Holcus lanatus L. | Poaceae | Graminoid | C |
| Hypericum perforatum L. | Hypericaceae | Perennial forb | AM |
| Hypochoeris radicata L. | Asteraceae | Perennial forb | C |
| Kickxia spuria (L.) Dumort. | Plantaginaceae | Annual forb | C |
| Leontodon tuberosus L. | Asteraceae | Perennial forb | HM |
| Leucanthemum ircutianum DC. | Asteraceae | Perennial forb | HM |
| Linaria vulgaris Mill. | Plantaginaceae | Perennial forb | AM |
| Linum bienne Mill. | Linaceae | Perennial forb | C |
| Lolium arundinaceum (Schreb.) Darbysh. | Poaceae | Graminoid | C |
| Lotus conrniculatus L. | Fabaceae | Legumes | AM |
| Luzula multiflora (Ehrh.) Lej. | Juncaceae | Graminoid | C |
| Lythrum salicaria L. | Lythraceae | Perennial forb | AM |
| Oenanthe pimpinelloides L. | Apiaceae | Perennial forb | AM |
| Plantago lanceolata L. | Plantaginaceae | Perennial forb | C |
| Potentilla reptans L. | Rosaceae | Perennial forb | C |
| Prunella laciniata L. (L.) | Lamiaceae | Perennial forb | C |
| Prunella vulgaris L. | Lamiaceae | Perennial forb | C |
| Prunella x intermedia Link | Lamiaceae | Perennial forb | C |
| Ranunculus bulbosus L. | Ranunculacea | Perennial forb | C |
| Rubus spp. | Rosaceae | Shrub | C |
| Sanguisorba minor Scop. | Rosaceae | Perennial forb | C |
| Silene flos-cuculi (L.) Clairv. | Caryophyllaceae | Perennial forb | C |
| Sonchus oleraceus L. | Asteraceae | Annual forb | AM |
| Trifolium campestre Schreb. | Fabaceae | Legumes | HM |
| Urospermum delachampii (L.) F.W. Schmidt | Asteraceae | Perennial forb | AM |
| Vicia sativa L. | Fabaceae | Legumes | HM |
| Biomass DW (g) | |||
|---|---|---|---|
| HM | AM | p Values | |
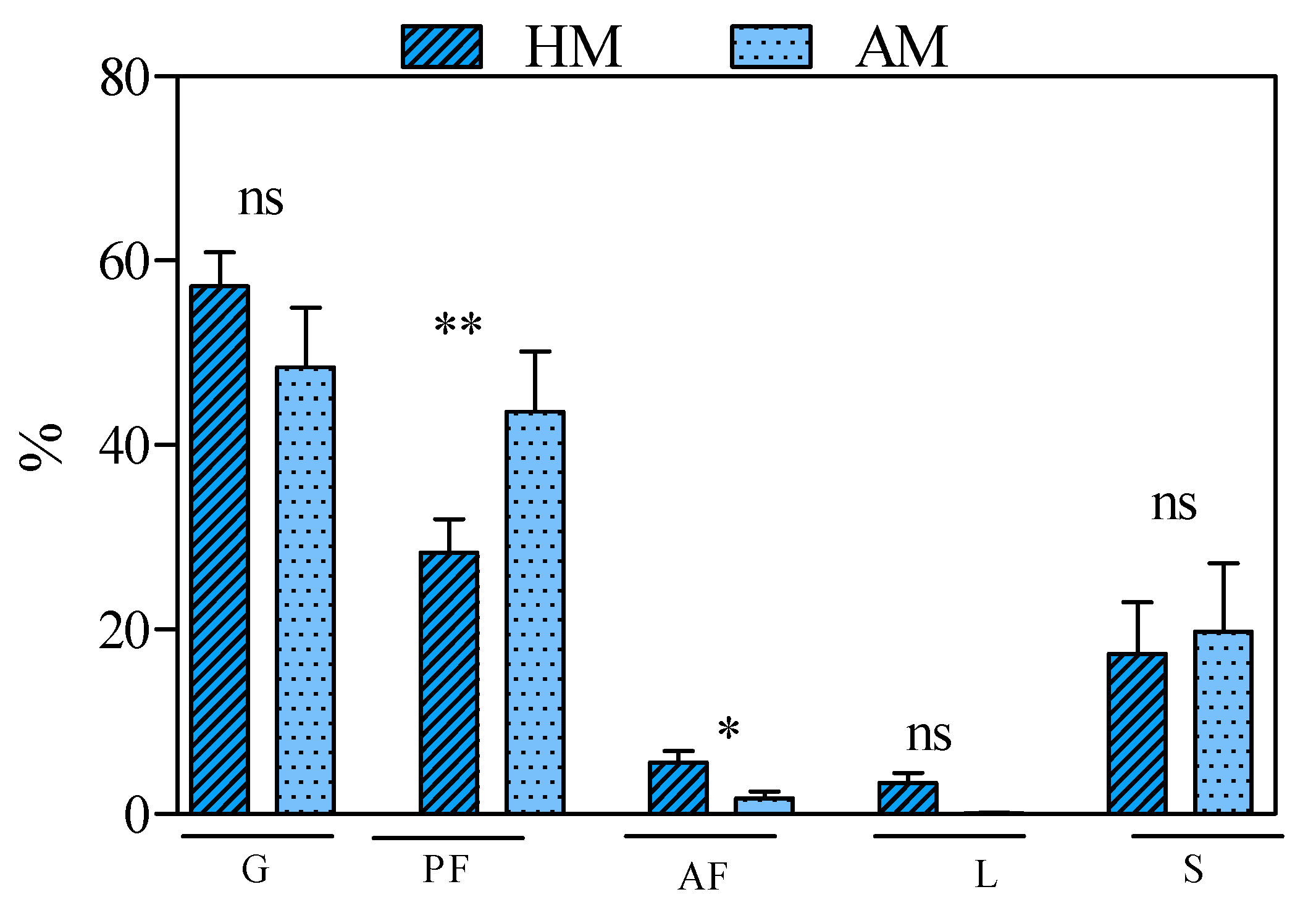
| Litter | 3.9 ± 0.83 | 26.2 ± 2.83 | *** |
| Graminoid | 46.0 ± 6.89 | 33.8 ± 5.50 | ns |
| Legumes | 0.4 ± 0.09 | 0.5 ± 0.49 | ns |
| Forbs | 11.8 ± 1.47 | 17.3 ± 3.80 | ns |
| Shrubs | 3.9 ± 1.91 | 5.6 ± 3.55 | ns |
| Soil Properties | HM | AM | p Values |
|---|---|---|---|
| pH | 5.6 ± 0.03 | 6.0 ± 0.05 | *** |
| Corg% | 1.7 ± 0.09 | 2.2 ± 0.16 | ** |
| Ntot% | 0.20 ± 0.02 | 0.26 ± 0.01 | * |
| AP | 0.66 ± 0.23 | 0.71 ± 0.21 | ns |
| CEC (cmol kg−1) | 14.1 ± 1.29 | 15.5 ± 0.68 | ns |
| EC (dS/m) | 0.20 ± 0.01 | 0.20 ± 0.01 | ns |
| pH | EC | CEC | Corg | N | Litter | Graminoid | Legumes | Forbs | Shrub | |
|---|---|---|---|---|---|---|---|---|---|---|
| pH | ||||||||||
| EC | 0.46 | |||||||||
| CEC | 0.36 | 0.36 | ||||||||
| Corg | 0.39 | 0.11 | 0.15 | |||||||
| N | 0.39 | 0.14 | 0.25 | 0.67 *** | ||||||
| Litter | 0.54 ** | 0.04 | −0.001 | 0.31 * | 0.28 | |||||
| Graminoid | −0.36 ** | −0.31 * | −0.18 | −0.10 | −0.01 | −0.17 | ||||
| legumes | 0.11 | 0.08 | 0.22 | 0.58 * | 0.42 * | 0.08 | −0.07 | |||
| Forbs | 0.43 | 0.23 | 0.11 | 0.12 | 0.30 | 0.11 | −0.001 | 0.26 | ||
| Shrub | 0.01 | 0.06 | 0.05 | 0.22 | 0.03 | −0.06 | −0.24 | −0.09 | −0.29 * | |
| SR | −0.32 * | 0.11 | −0.03 | −0.44 ** | −0.21 * | −0.66 *** | 0.21 | −0.08 | 0.14 | −0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vannucchi, F.; Lazzeri, V.; Rosellini, I.; Scatena, M.; Caudai, C.; Bretzel, F. Short-Term Abandonment versus Mowing in a Mediterranean-Temperate Meadow: Effects on Floristic Composition, Plant Functionality, and Soil Properties—A Case Study. Agriculture 2022, 12, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12010078
Vannucchi F, Lazzeri V, Rosellini I, Scatena M, Caudai C, Bretzel F. Short-Term Abandonment versus Mowing in a Mediterranean-Temperate Meadow: Effects on Floristic Composition, Plant Functionality, and Soil Properties—A Case Study. Agriculture. 2022; 12(1):78. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12010078
Chicago/Turabian StyleVannucchi, Francesca, Valerio Lazzeri, Irene Rosellini, Manuele Scatena, Claudia Caudai, and Francesca Bretzel. 2022. "Short-Term Abandonment versus Mowing in a Mediterranean-Temperate Meadow: Effects on Floristic Composition, Plant Functionality, and Soil Properties—A Case Study" Agriculture 12, no. 1: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12010078