
A First Attempt to Combine NIRS and Plenoptic Cameras for the Assessment of Grasslands Functional Diversity and Species Composition

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Sites and Design
- (a)
- Experimental sites and pedoclimatic conditions
- (b)
- Design of the plant mixtures and technical itinerary
2.2. Field Measurements
- (a)
- Plenoptic images
- (b)
- Measurement of functional traits
- (c)
- Biomass NIRS measurements and species composition
2.3. Images and NIRS Processing
- -
- A segmentation approach to separate the images into different categories based on plant species. This enabled us to evaluate the extent of the cover of each species;
- -
- A color approach to evaluate the greenness of the cover.
- (a)
- Image segmentation
- (b)
- Color Analysis
- (c)
- NIRS Analysis
2.4. Statistical Analysis
- (a)
- Calculation of vegetation descriptors
- (b)
- Correlation between the different variables 2 by 2 using the maximum information coefficients
- (c)
- Evaluating the complementarity and redundancy between the variables using variance partitioning
- (d)
- Predicting capacity using partial least squares regression
3. Results
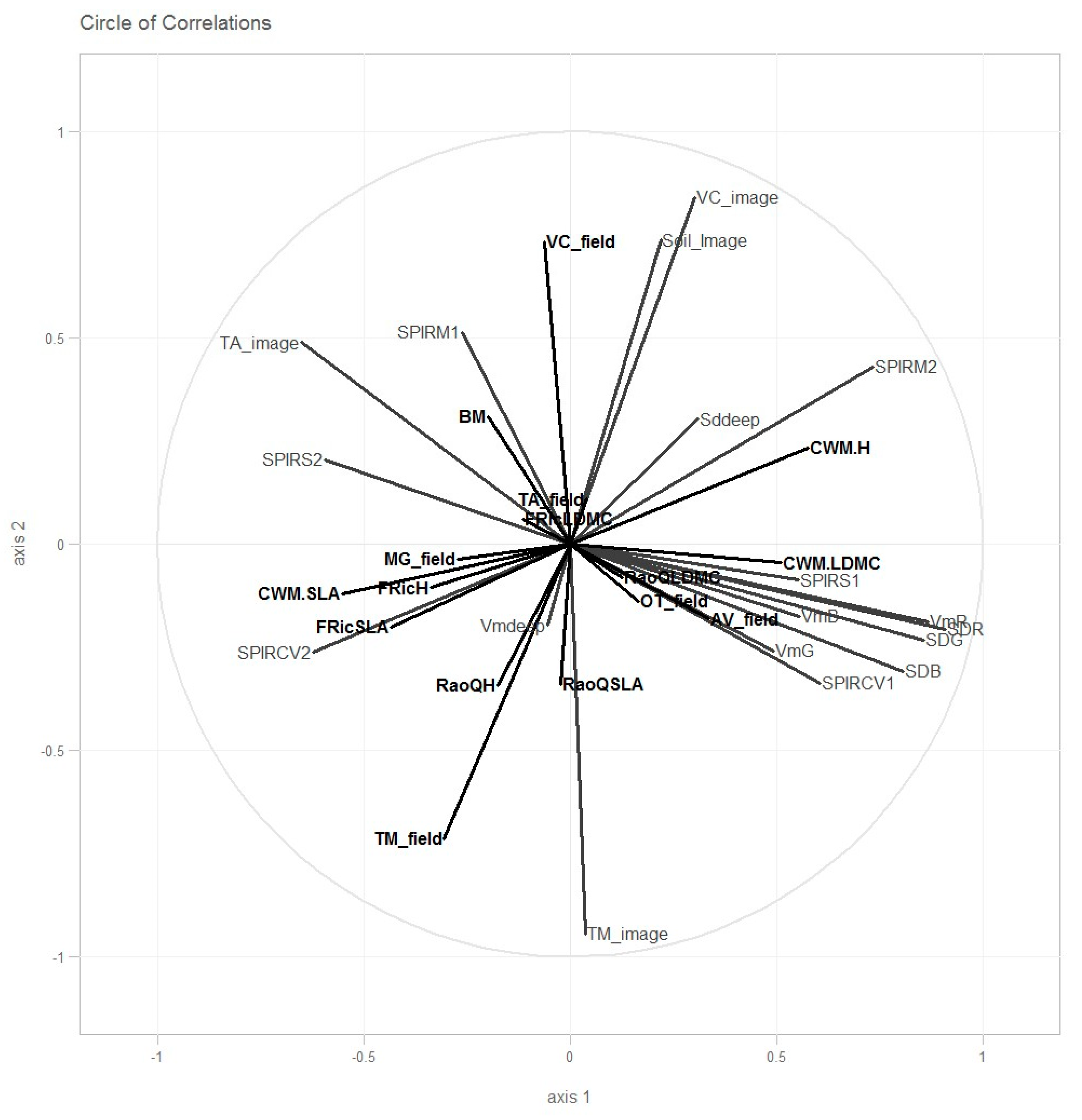
3.1. MIC Correlation
- (a)
- Vegetation height descriptors
- (b)
- Descriptors of leaf dry matter content
- (c)
- Descriptors of specific leaf area
- (d)
- Biomass and species composition
3.2. Variance Partition
3.3. Partial Least Squares (PLS) Regressions
4. Discussion
4.1. Limits of This First Attempt
4.2. Evaluating Functional Traits and Diversity Using Optical Captors
4.3. Future Outlook on the Use of Plenoptic Images
4.4. Combination of Several Tools
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Panunzi, E. Are Grasslands under Threat? Brief Analysis of FAO Statistical Data on Pasture and Fodder Crops; UN Food and Agriculture Organization: Rome, Italy, 2008. [Google Scholar]
- Givens, D.I.; Owen, E.; Omed, H.; Axford, R. Forage Evaluation in Ruminant Nutrition; CABI: Wallingford, UK, 2000. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- McMahon, L.R.; McAllister, T.A.; Berg, B.P.; Majak, W.; Acharya, S.N.; Popp, J.D.; Coulman, B.E.; Wang, Y.; Cheng, K.-J. A review of the effects of forage condensed tannins on ruminal fermentation and bloat in grazing cattle. Can. J. Plant Sci. 2000, 80, 469–485. [Google Scholar] [CrossRef] [Green Version]
- Spiegelberger, T.; Gillet, F.; Amiaud, B.; Thébault, A.; Mariotte, P.; Buttler, A. How do plant community ecologists consider the complementarity of observational, experimental and theoretical modelling approaches? Plant Ecol. Evol. 2012, 145, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user’s guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef] [Green Version]
- McGill, B.J.; Enquist, B.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- De Bello, F.; Lavorel, S.; Díaz, S.; Harrington, R.; Cornelissen, J.H.C.; Bardgett, R.D.; Berg, M.P.; Cipriotti, P.; Feld, C.K.; Hering, D.; et al. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers. Conserv. 2010, 19, 2873–2893. [Google Scholar] [CrossRef]
- Wachendorf, M.; Fricke, T.; Möckel, T. Remote sensing as a tool to assess botanical composition, structure, quantity and quality of temperate grasslands. Grass Forage Sci. 2018, 73, 1–14. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the great plains with ERTS. In Proceedings of the Third ERTS Symposium, SP-351, Washington, DC, USA, 10–14 December 1973. [Google Scholar]
- Chen, J.; Gu, S.; Shen, M.; Tang, Y.; Matsushita, B. Estimating aboveground biomass of grassland having a high canopy cover: An exploratory analysis of in situ hyperspectral data. Int. J. Remote Sens. 2009, 30, 6497–6517. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A. Integrating imaging spectroscopy and neural networks to map grass quality in the Kruger National Park, South Africa. Remote Sens. Environ. 2004, 90, 104–115. [Google Scholar] [CrossRef]
- De Boever, J.; Cottyn, B.; Vanacker, J.; Boucqué, C. The use of NIRS to predict the chemical composition and the energy value of compound feeds for cattle. Anim. Feed Sci. Technol. 1995, 51, 243–253. [Google Scholar] [CrossRef]
- Wachendorf, M.; Ingwersen, B.; Taube, F. Taube Prediction of the clover content of red clover- and white clover-grass mixtures by near-infrared reflectance spectroscopy. Grass Forage Sci. 1999, 54, 87–90. [Google Scholar] [CrossRef]
- Pilon, R.; Klumpp, K.; Carrère, P.; Picon-Cochard, C. Determination of Aboveground Net Primary Productivity and Plant Traits in Grasslands with Near-Infrared Reflectance Spectroscopy. Ecosystems 2010, 13, 851–859. [Google Scholar] [CrossRef]
- Bonnal, L.; Julien, L.; Delalande, M.; Bastianelli, D. How can a dry forage database be used to predict fresh grass composition by NIR spectroscopy? Data transfer vs. spectra transfer. In Proceedings of the International Conference on Near Infrared Spectroscopy, La Grande-Motte, France, 2–7 June 2013; pp. 685–688. [Google Scholar]
- Onyango, C.; Marchant, J.; Grundy, A.; Phelps, K.; Reader, R. Image Processing Performance Assessment Using Crop Weed Competition Models. Precis. Agric. 2005, 6, 183–192. [Google Scholar] [CrossRef]
- Hague, T.; Tillett, N.D.; Wheeler, H. Automated Crop and Weed Monitoring in Widely Spaced Cereals. Precis. Agric. 2006, 7, 21–32. [Google Scholar] [CrossRef]
- Gebhardt, S.; Kühbauch, W. A new algorithm for automatic Rumex obtusifolius detection in digital images using colour and texture features and the influence of image resolution. Precis. Agric. 2007, 8, 1–13. [Google Scholar] [CrossRef]
- Gebhardt, S.; Schellberg, J.; Lock, R.; Kühbauch, W. Identification of broad-leaved dock (Rumex obtusifolius L.) on grassland by means of digital image processing. Precis. Agric. 2006, 7, 165–178. [Google Scholar] [CrossRef]
- Himstedt, M.; Fricke, T.; Wachendorf, M. Determining the Contribution of Legumes in Legume-Grass Mixtures Using Digital Image Analysis. Crop Sci. 2009, 49, 1910–1916. [Google Scholar] [CrossRef]
- Himstedt, M.; Fricke, T.; Wachendorf, M. The Relationship between Coverage nd Dry Matter Contribution of Forage Legumes in Binary Legume–Grass Mixtures. Crop Sci. 2010, 50, 2186–2193. [Google Scholar] [CrossRef]
- Ng, R.; Levoy, M.; Brédif, M.; Duval, G.; Horowitz, M.; Hanrahan, P. Light Field Photography with a Hand-Held Plenoptic Camera; Computer Science Tech Report CSTR 2005-02; Stanford University: Stanford, CA, USA, 2005; pp. 1–11. [Google Scholar]
- Hahne, C.; Aggoun, A.; Velisavljevic, V.; Fiebig, S.; Pesch, M. Refocusing distance of a standard plenoptic camera. Opt. Express 2016, 24, 21521–21540. [Google Scholar] [CrossRef] [Green Version]
- Huguenin, J.; Julien, L.; Capron, J.; Desclaux, D.; Lesnoff, M.; Crespo, D. Multispecies pastures in Mediterranean zones: Agro-ecological resilience of forage production subject to climatic variations. Grassl. Resour. Extensive Farming Syst. Marg. Lands Major Driv. Future Scenar. 2017, 2017, 566–569. [Google Scholar]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- WinFolia, version PRO 2001; Regent Instruments Inc.: Quebec, QC, Canada, 2001.
- Arganda-Carreras, I.; Kaynig, V.; Rueden, C.; Eliceiri, K.W.; Schindelin, J.; Cardona, A.; Seung, H.S. Trainable Weka Segmentation: A machine learning tool for microscopy pixel classification. Bioinformatics 2017, 33, 2424–2426. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. Package FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. 2014. Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 13 April 2022).
- Reshef, D.N.; Reshef, Y.A.; Finucane, H.K.; Grossman, S.R.; McVean, G.; Turnbaugh, P.J.; Lander, E.S.; Mitzenmacher, M.; Sabeti, P.C. Detecting Novel Associations in Large Data Sets. Science 2011, 334, 1518–1524. [Google Scholar] [CrossRef] [Green Version]
- Speed, T. A Correlation for the 21st Century. Science 2011, 334, 1502–1503. [Google Scholar] [CrossRef]
- Albanese, D.; Filosi, M.; Visintainer, R.; Riccadonna, S.; Jurman, G.; Furlanello, C. Minerva and minepy: A C engine for the MINE suite and its R, Python and MATLAB wrappers. Bioinformatics 2013, 29, 407–408. [Google Scholar] [CrossRef] [Green Version]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the Spatial Component of Ecological Variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. 2012. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 13 April 2022).
- Costa, F.R.C.; Lang, C.; Almeida, D.R.A.; Castilho, C.V.; Poorter, L. Near-infrared spectrometry allows fast and extensive predictions of functional traits from dry leaves and branches. Ecol. Appl. 2018, 28, 1157–1167. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant Functional Markers Capture Ecosystem Properties during Secondary Succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field. Methodol. Matters Funct. Ecol. 2008, 22, 134–147. [Google Scholar]
- Pakeman, R.; Quested, H. Sampling plant functional traits: What proportion of the species need to be measured? Appl. Veg. Sci. 2007, 10, 91–96. [Google Scholar] [CrossRef]
- Walker, B.; Kinzig, A.; Langridge, J. Original Articles: Plant Attribute Diversity, Resilience, and Ecosystem Function: The Nature and Significance of Dominant and Minor Species. Ecosystems 1999, 2, 95–113. [Google Scholar] [CrossRef]
- Mouillot, D.; Bellwood, D.R.; Baraloto, C.; Chave, J.; Galzin, R.; Harmelin-Vivien, M.; Kulbicki, M.; Lavergne, S.; Lavorel, S.; Mouquet, N.; et al. Rare Species Support Vulnerable Functions in High-Diversity Ecosystems. PLoS Biol. 2013, 11, e1001569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violle, C.; Thuiller, W.; Mouquet, N.; Munoz, F.; Kraft, N.J.; Cadotte, M.W.; Livingstone, S.; Mouillot, D. Functional Rarity: The Ecology of Outliers. Trends Ecol. Evol. 2017, 32, 356–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violle, C.; Borgy, B.; Choler, P. Trait databases: Misuses and precautions. J. Veg. Sci. 2015, 26, 826–827. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.C.; Falster, D.S.; Garnier, E.; Hikosaka, K.; Lamont, B.B.; Lee, W.; Oleksyn, J.; Osada, N.; et al. Assessing the generality of global leaf trait relationships. New Phytol. 2005, 166, 485–496. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.C.; Falster, D.S.; Groom, P.K.; Hikosaka, K.; Lee, W.; Lusk, C.H.; Niinemets, Ü.; Oleksyn, J.; et al. Modulation of leaf economic traits and trait relationships by climate. Glob. Ecol. Biogeogr. 2005, 14, 411–421. [Google Scholar] [CrossRef]
- Tran, H.; Salgado, P.; Lecomte, P. Species, climate and fertilizer effects on grass fibre and protein in tropical environments. J. Agric. Sci. 2009, 147, 555–568. [Google Scholar] [CrossRef]
- Navas, M.-L.; Roumet, C.; Bellmann, A.; Laurent, G.; Garnier, E. Suites of plant traits in species from different stages of a Mediterranean secondary succession. Plant Biol. 2010, 12, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Prošek, J.; Šímová, P. UAV for mapping shrubland vegetation: Does fusion of spectral and vertical information derived from a single sensor increase the classification accuracy? Int. J. Appl. Earth Obs. Geoinf. 2019, 75, 151–162. [Google Scholar] [CrossRef]
- Waser, L.; Baltsavias, E.; Ecker, K.; Eisenbeiss, H.; Feldmeyer-Christe, E.; Ginzler, C.; Küchler, M.; Zhang, L. Assessing changes of forest area and shrub encroachment in a mire ecosystem using digital surface models and CIR aerial images. Remote Sens. Environ. 2008, 112, 1956–1968. [Google Scholar] [CrossRef]
- Aasen, H.; Burkart, A.; Bolten, A.; Bareth, G. Generating 3D hyperspectral information with lightweight UAV snapshot cameras for vegetation monitoring: From camera calibration to quality assurance. ISPRS J. Photogramm. Remote Sens. 2015, 108, 245–259. [Google Scholar] [CrossRef]
- Frey, J.; Kovach, K.; Stemmler, S.; Koch, B. UAV Photogrammetry of Forests as a Vulnerable Process. A Sensitivity Analysis for a Structure from Motion RGB-Image Pipeline. Remote Sens. 2018, 10, 912. [Google Scholar] [CrossRef] [Green Version]
- Bossoukpe, M.; Ndiaye, O.; Diatta, S.; Diatta, O.; Diouf, A.A.; Assouma, M.H.; Faye, E.; Taugourdeau, S. Ground based photogrammetry to assess herbaceous biomass in Sahelian rangelands. In Proceedings of the European Grassland Federation, Helsinki, Finland, 19–21 October 2020. [Google Scholar]
- Dale, L.-M. L’utilisation de la spectrométrie (NIR) et l’imagerie hyperspectrale (NIR-HIS) proche infrarouge pour étudier la composition chimique et botanique de des fourrages. Ph.D. Thesis, Université of Liege, Liege, Belgique, 2014. [Google Scholar]
- Moeckel, T.; Safari, H.; Reddersen, B.; Fricke, T.; Wachendorf, M. Fusion of Ultrasonic and Spectral Sensor Data for Improving the Estimation of Biomass in Grasslands with Heterogeneous Sward Structure. Remote Sens. 2017, 9, 98. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species | Mix1 in kg·ha−1 | Mix2 in kg·ha−1 | |
|---|---|---|---|
| Grass | Avena sativa | 10 | 10 |
| Legumes | Vicia sativa | 15 | 0 |
| Trifolium alexandrinum, | 5 | 5 | |
| Medicago truncatula | 2.5 | 2.5 | |
| Trifolium michelianum | 0 | 5 |
| 16 June | 29 June | ||||
|---|---|---|---|---|---|
| General Average | Mix1 | Mix2 | Mix1 | Mix2 | |
| CWM. H in cm | 42.67 | 40.84 | 32.29 | 48.07 | 49.49 |
| CWM.LDMC in mg·g−1 | 336.68 | 322.32 | 287.05 | 345.51 | 391.83 |
| CWM.SLA in cm2·g−1 | 21.01 | 21.13 | 25.36 | 19.73 | 17.84 |
| FRicH | 1.70 | 1.82 | 2.06 | 1.45 | 1.45 |
| FRicLDMC | 1.68 | 2.66 | 1.41 | 0.97 | 1.69 |
| FRicSLA | 2.09 | 2.43 | 2.70 | 1.34 | 1.88 |
| RaoQH | 0.34 | 0.26 | 0.55 | 0.27 | 0.30 |
| RaoQLDMC | 0.41 | 0.51 | 0.26 | 0.21 | 0.68 |
| RaoQSLA | 0.53 | 0.49 | 0.66 | 0.25 | 0.70 |
| Biomass in g | 200.31 | 206.53 | 217.69 | 227.50 | 149.50 |
| Ta biomass in g | 120.95 | 100.56 | 129.23 | 153.00 | 101.00 |
| Tm biomass in g | 17.79 | 0.00 | 53.65 | 0.00 | 17.50 |
| Mg biomass in g | 0.04 | 0.05 | 0.10 | 0.00 | 0.00 |
| Av biomass in g | 8.30 | 4.28 | 5.91 | 9.50 | 13.50 |
| Vc biomass in g | 34.06 | 80.22 | 0.00 | 56.00 | 0.00 |
| Weed biomass in g | 23.91 | 13.75 | 25.89 | 21.50 | 34.50 |
| NIRS | Colors | Segmentation | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NIRSM1 | NIRSM2 | NIRSS1 | NIRSS2 | NIRSCV1 | NIRSCV2 | Vmdeep | Sddeep | VmR | SDR | VmB | SDB | VmG | SDG | TA | TM | VC | Soil | |
| CWM. H | 0.33 | 0.59 | 0.40 | 0.37 | 0.29 | 0.38 | 0.31 | 0.32 | 0.59 | 0.69 | 0.28 | 0.34 | 0.31 | 0.39 | 0.31 | 0.36 | 0.34 | 0.48 |
| FRicH | 0.27 | 0.28 | 0.32 | 0.33 | 0.22 | 0.21 | 0.37 | 0.25 | 0.34 | 0.41 | 0.24 | 0.46 | 0.25 | 0.37 | 0.39 | 0.25 | 0.26 | 0.39 |
| RaoQH | 0.26 | 0.33 | 0.25 | 0.34 | 0.18 | 0.31 | 0.30 | 0.33 | 0.31 | 0.41 | 0.24 | 0.26 | 0.22 | 0.21 | 0.31 | 0.28 | 0.28 | 0.49 |
| CWM.LDMC | 0.23 | 0.48 | 0.29 | 0.31 | 0.40 | 0.29 | 0.20 | 0.33 | 0.45 | 0.50 | 0.36 | 0.26 | 0.48 | 0.28 | 0.27 | 0.27 | 0.24 | 0.38 |
| FRicLDMC | 0.37 | 0.18 | 0.43 | 0.23 | 0.39 | 0.23 | 0.41 | 0.23 | 0.20 | 0.25 | 0.30 | 0.33 | 0.36 | 0.31 | 0.20 | 0.24 | 0.24 | 0.24 |
| RaoQLDMC | 0.33 | 0.20 | 0.29 | 0.23 | 0.48 | 0.22 | 0.61 | 0.32 | 0.28 | 0.31 | 0.26 | 0.24 | 0.32 | 0.31 | 0.25 | 0.19 | 0.19 | 0.26 |
| CWM.SLA | 0.25 | 0.36 | 0.36 | 0.28 | 0.30 | 0.28 | 0.27 | 0.17 | 0.38 | 0.37 | 0.39 | 0.26 | 0.30 | 0.28 | 0.31 | 0.28 | 0.28 | 0.28 |
| FRicSLA | 0.23 | 0.34 | 0.23 | 0.24 | 0.32 | 0.26 | 0.29 | 0.31 | 0.32 | 0.35 | 0.23 | 0.28 | 0.25 | 0.43 | 0.31 | 0.21 | 0.26 | 0.25 |
| RaoQSLA | 0.35 | 0.29 | 0.35 | 0.32 | 0.55 | 0.31 | 0.29 | 0.21 | 0.40 | 0.31 | 0.28 | 0.26 | 0.27 | 0.36 | 0.24 | 0.33 | 0.33 | 0.31 |
| Biomass in g | 0.55 | 0.30 | 0.20 | 0.36 | 0.37 | 0.30 | 0.31 | 0.21 | 0.45 | 0.43 | 0.22 | 0.23 | 0.24 | 0.22 | 0.57 | 0.48 | 0.35 | 0.39 |
| Ta biomass in g | 0.35 | 0.29 | 0.41 | 0.33 | 0.34 | 0.30 | 0.33 | 0.26 | 0.28 | 0.38 | 0.29 | 0.23 | 0.34 | 0.21 | 0.27 | 0.28 | 0.18 | 0.23 |
| Tm biomass in g | 0.31 | 0.46 | 0.32 | 0.28 | 0.32 | 0.28 | 0.21 | 0.29 | 0.53 | 0.50 | 0.30 | 0.32 | 0.22 | 0.34 | 0.53 | 1.00 | 1.00 | 0.78 |
| Mg biomass in g | 0.28 | 0.20 | 0.23 | 0.22 | 0.26 | 0.19 | 0.23 | 0.38 | 0.32 | 0.30 | 0.26 | 0.25 | 0.29 | 0.32 | 0.16 | 0.24 | 0.15 | 0.22 |
| Av biomass in g | 0.23 | 0.25 | 0.58 | 0.33 | 0.29 | 0.25 | 0.27 | 0.49 | 0.30 | 0.33 | 0.43 | 0.27 | 0.34 | 0.30 | 0.33 | 0.29 | 0.20 | 0.22 |
| Vc biomass in g | 0.28 | 0.35 | 0.32 | 0.31 | 0.22 | 0.34 | 0.22 | 0.29 | 0.53 | 0.41 | 0.20 | 0.32 | 0.30 | 0.34 | 0.41 | 1.00 | 1.00 | 0.77 |
| Weed biomass in g | 0.37 | 0.34 | 0.38 | 0.44 | 0.25 | 0.39 | 0.22 | 0.19 | 0.53 | 0.44 | 0.27 | 0.24 | 0.29 | 0.34 | 0.30 | 0.22 | 0.24 | 0.31 |
| Field Variable | C | S | N | CS | SN | CN | CSN | R |
|---|---|---|---|---|---|---|---|---|
| CWM. H in cm | 0.02 | −0.05 | 0.07 | 0.01 | 0.06 | 0.08 | 0.19 | 0.63 |
| CWM.LDMC in mg·g−1 | 0.00 | −0.14 | −0.15 | 0.05 | 0.09 | 0.17 | 0.03 | 0.96 |
| CWM.SLA in cm2·g−1 | 0.15 | −0.07 | 0.09 | 0.00 | 0.03 | 0.10 | 0.06 | 0.63 |
| FRicH | 0.23 | −0.01 | 0.13 | −0.03 | −0.10 | −0.17 | 0.13 | 0.81 |
| FRicLDMC | −0.02 | −0.09 | −0.10 | 0.01 | 0.08 | 0.09 | −0.01 | 1.05 |
| FRicSLA | 0.00 | 0.02 | 0.05 | 0.05 | −0.03 | 0.08 | 0.13 | 0.69 |
| RaoQH | 0.33 | 0.16 | 0.26 | 0.01 | −0.19 | −0.26 | 0.18 | 0.51 |
| RaoQLDMC | 0.08 | −0.05 | 0.28 | 0.02 | 0.12 | 0.05 | −0.15 | 0.65 |
| RaoQSLA | −0.08 | −0.02 | 0.06 | 0.07 | 0.04 | 0.03 | −0.03 | 0.92 |
| Biomass in g | 0.07 | 0.04 | −0.05 | 0.11 | 0.08 | 0.02 | 0.15 | 0.57 |
| Ta biomass in g | −0.03 | 0.21 | 0.22 | 0.00 | −0.09 | 0.06 | −0.04 | 0.67 |
| Tm biomass in g | 0.19 | 0.29 | 0.08 | −0.04 | 0.07 | −0.09 | 0.27 | 0.25 |
| Mg biomass in g | −0.05 | 0.06 | 0.01 | −0.02 | −0.03 | 0.19 | −0.10 | 0.94 |
| Av biomass in g | −0.21 | −0.09 | −0.11 | 0.05 | 0.11 | 0.10 | 0.06 | 1.08 |
| Vc biomass in g | −0.07 | 0.29 | 0.07 | −0.05 | 0.32 | 0.06 | −0.02 | 0.41 |
| Weed biomass in g | 0.00 | 0.03 | 0.46 | −0.02 | 0.03 | 0.15 | −0.06 | 0.40 |
| t1 | t2 | |
|---|---|---|
| CWM. H in cm | 0.244 | 0.014 |
| CWM.LDMC in mg·g−1 | 0.200 | −0.064 |
| CWM.SLA in cm2·g−1 | 0.242 | −0.049 |
| FRicH | 0.071 | −0.039 |
| FRicLDMC | −0.019 | 0.050 |
| FRicSLA | −0.012 | −0.054 |
| RaoQH | −0.011 | −0.115 |
| RaoQLDMC | 0.155 | 0.007 |
| RaoQSLA | −0.053 | 0.012 |
| Biomass in g | −0.046 | 0.035 |
| Ta biomass in g | −0.083 | −0.044 |
| Tm biomass in g | 0.022 | 0.512 |
| Mg biomass in g | 0.010 | −0.090 |
| Av biomass in g | 0.063 | −0.029 |
| Vc biomass in g | −0.020 | 0.511 |
| Weed biomass in g | −0.018 | −0.024 |
| Q2 | 0.047 | 0.043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taugourdeau, S.; Dionisi, M.; Lascoste, M.; Lesnoff, M.; Capron, J.M.; Borne, F.; Borianne, P.; Julien, L. A First Attempt to Combine NIRS and Plenoptic Cameras for the Assessment of Grasslands Functional Diversity and Species Composition. Agriculture 2022, 12, 704. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12050704
Taugourdeau S, Dionisi M, Lascoste M, Lesnoff M, Capron JM, Borne F, Borianne P, Julien L. A First Attempt to Combine NIRS and Plenoptic Cameras for the Assessment of Grasslands Functional Diversity and Species Composition. Agriculture. 2022; 12(5):704. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12050704
Chicago/Turabian StyleTaugourdeau, Simon, Mathilde Dionisi, Mylène Lascoste, Matthieu Lesnoff, Jean Marie Capron, Fréderic Borne, Philippe Borianne, and Lionel Julien. 2022. "A First Attempt to Combine NIRS and Plenoptic Cameras for the Assessment of Grasslands Functional Diversity and Species Composition" Agriculture 12, no. 5: 704. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12050704