
Long-Term Fertilization Alters Mycorrhizal Colonization Strategy in the Roots of Agrostis capillaris

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Laboratory Analysis
2.3. Data Analysis
3. Results
3.1. Variations Induced by Treatments on General Mycorrhizal Colonization
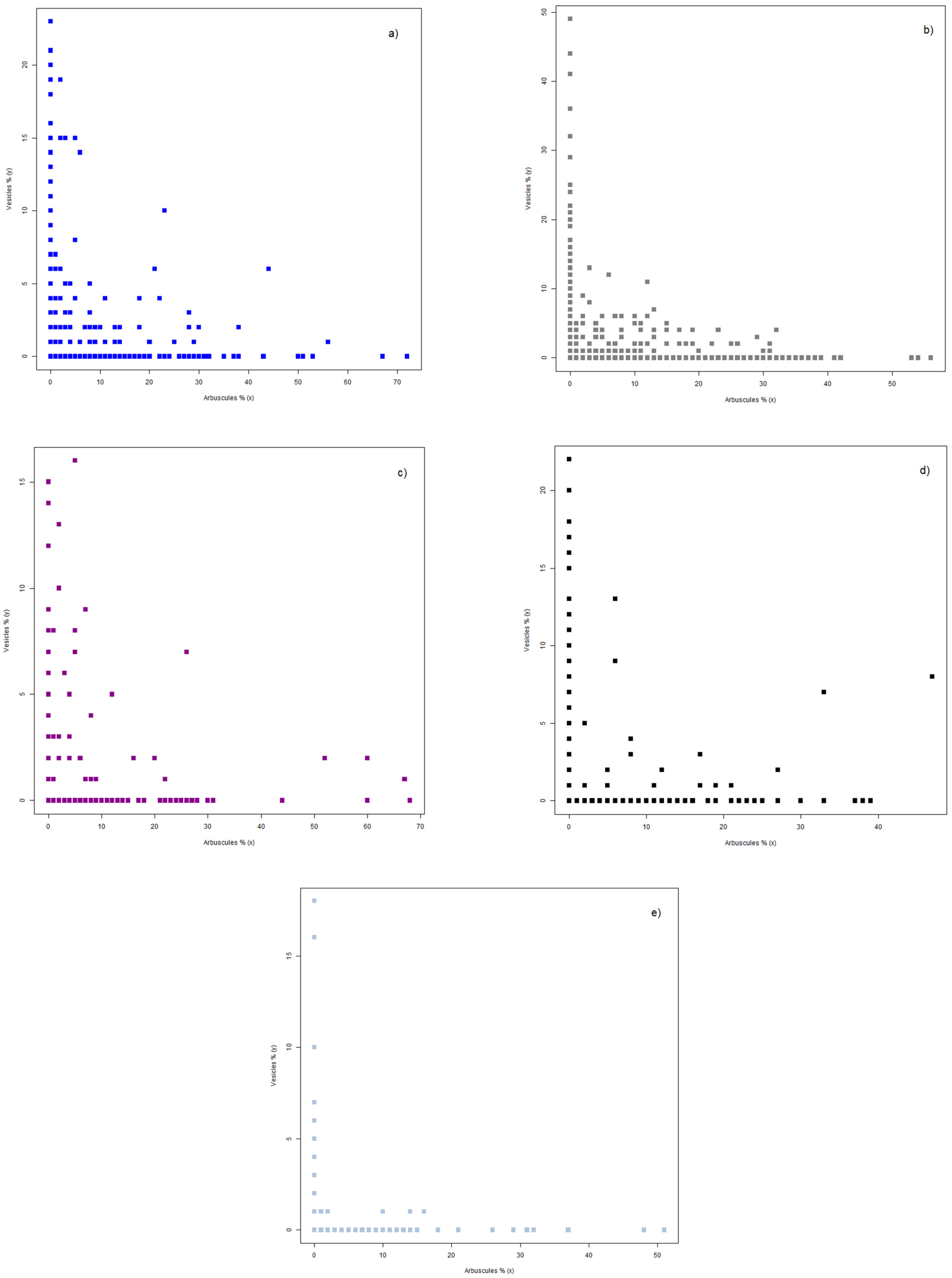
3.2. Interrelations between Colonization Parameters and Their Potential Simultaneous Presence in Colonized Roots
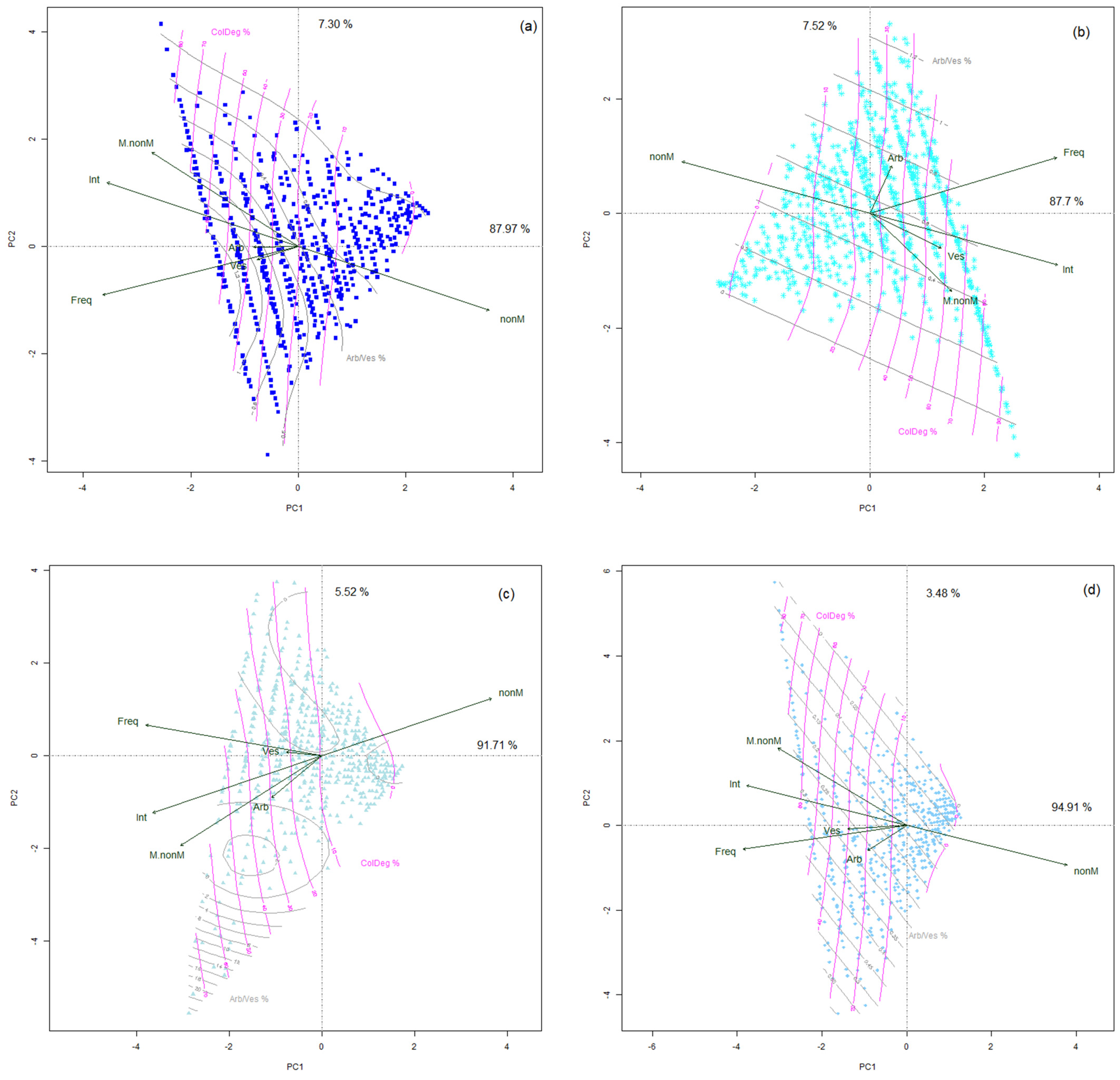
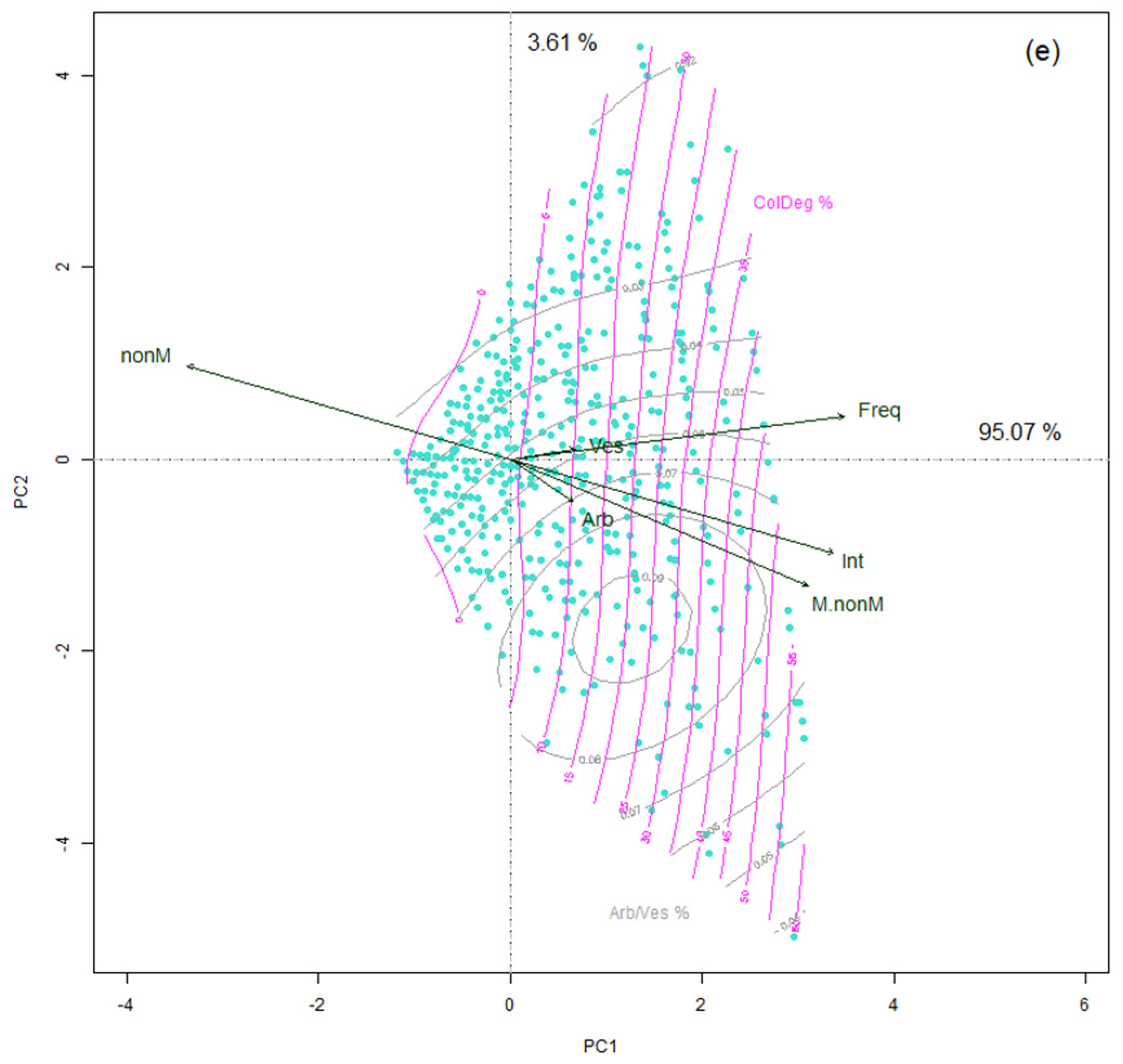
3.3. Dispersion of Mycorrhizal Colonization Data as Shaped by Applied Treatments
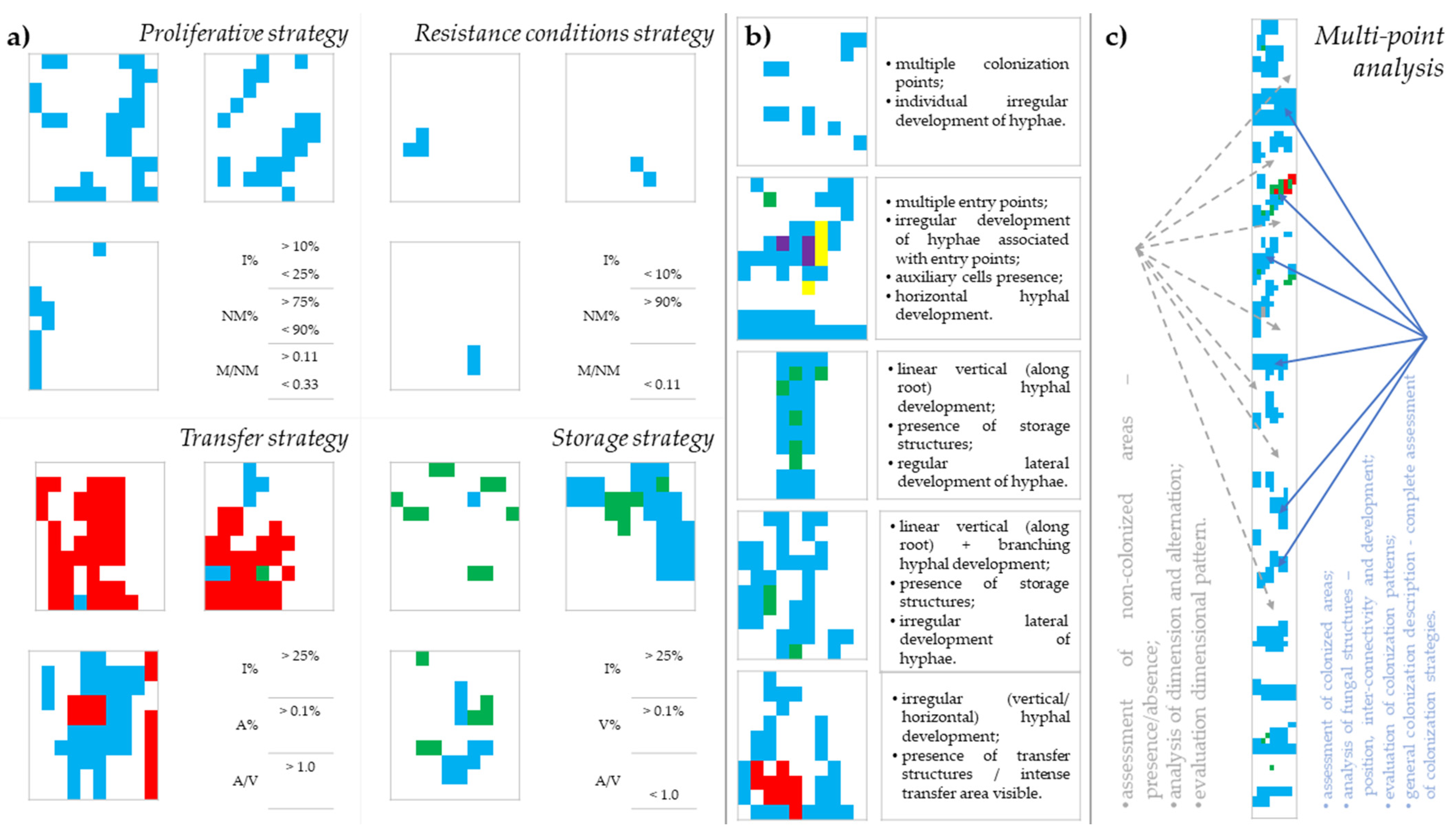
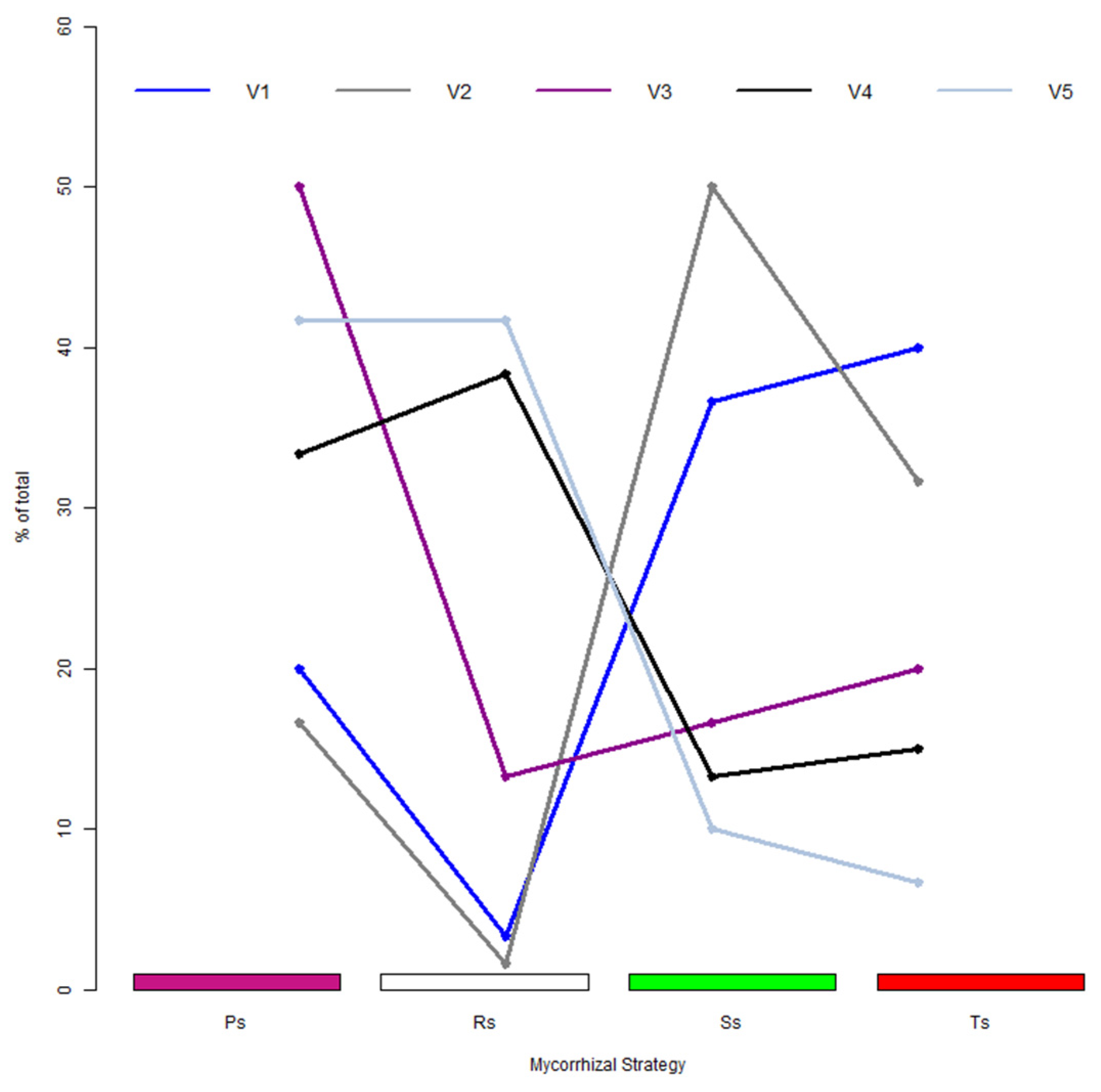
3.4. Establishment of Multi-Point Analysis as a Translation Tool in Deciphering Colonization Patterns and Strategies
3.5. Colonization Strategies Induced by the Application of Long-Term Treatments
4. Discussion
4.1. Visibility of the Treatment Effect in Variability of Colonization Parameters
4.2. Interrelations between Mycorrhizal Parameters Shape the Model of Colonization Mechanism
4.3. Linking Colonization Patterns and Fungal Strategies
4.4. Long-Term Teatments Shape Different Mycorrhizal Colonization Strategies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmitt, T.; Rákosy, L. Changes of Traditional Agrarian Landscapes and Their Conservation Implications: A Case Study of Butterflies in Romania. Divers. Distrib. 2007, 13, 855–862. [Google Scholar] [CrossRef]
- Bond, W.J.; Parr, C.L. Beyond the Forest Edge: Ecology, Diversity and Conservation of the Grassy Biomes. Biol. Conserv. 2010, 143, 2395–2404. [Google Scholar] [CrossRef]
- Carlson, M.L.; Flagstad, L.A.; Gillet, F.; Mitchell, E.A.D. Community development along a proglacial chronosequence: Are above-ground and below-ground community structure controlled more by biotic than abiotic factors? J. Ecol. 2010, 98, 1084–1095. [Google Scholar] [CrossRef]
- van Andel, J.; Aronson, J. Restoration Ecology: The New Frontier; John Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 9781118223154. [Google Scholar]
- Luo, W.; Xu, C.; Ma, W.; Yue, X.; Liang, X.; Zuo, X.; Knapp, A.K.; Smith, M.D.; Sardans, J.; Dijkstra, F.A.; et al. Effects of extreme drought on plant nutrient uptake and resorption in rhizomatous vs bunchgrass-dominated grasslands. Oecologia 2018, 188, 633–643. [Google Scholar] [CrossRef]
- Egli, T. How to live at very low substrate concentration. Water Res. 2010, 44, 4826–4837. [Google Scholar] [CrossRef]
- Duff, A.M.; Forrestal, P.; Ikoyi, I.; Brennan, F. Assessing the long-term impact of urease and nitrification inhibitor use on microbial community composition, diversity and function in grassland soil. Soil Biol. Biochem. 2022, 170, 108709. [Google Scholar] [CrossRef]
- Davison, J.; Vasar, M.; Sepp, S.K.; Oja, J.; Al-Quraishy, S.; Bueno, C.G.; Cantero, J.J.; Fabiano, E.C.; Decocq, G.; Fraser, L.; et al. Dominance, diversity, and niche breadth in arbuscular mycorrhizal fungal communities. Ecology 2022, e3761. [Google Scholar] [CrossRef]
- Le Roux, X.; Schmid, B.; Poly, F.; Barnard, R.L.; Niklaus, P.A.; Guillaumaud, N.; Habekost, M.; Oelmann, Y.; Philippot, L.; Salles, J.F.; et al. Soil Environmental Conditions and Microbial Build-Up Mediate the Effect of Plant Diversity on Soil Nitrifying and Denitrifying Enzyme Activities in Temperate Grasslands. PLoS ONE 2013, 8, e61069. [Google Scholar] [CrossRef]
- Christina, B.; Braker, G.; Matthies, D.; Reuter, A.; Engels, C.; Conrad, R. Impact of plant functional group, plant species, and sampling time on the composition of nirK-type denitrifier communities in soil. Appl. Environ. Microbiol. 2007, 73, 6876–6884. [Google Scholar]
- Debasis, M.; Braker, G.; Matthies, D.; Reuter, A.; Engels, C.; Conrad, R. Involvement of strigolactone hormone in root development, influence and interaction with mycorrhizal fungi in plant: Mini-review. Curr. Res. Microb. Sci. 2021, 2, 100026. [Google Scholar]
- Madawala, H.M.S.P. Chapter 7—Arbuscular Mycorrhizal Fungi as Biofertilizers: Current Trends, Challenges, and Future Prospects. In Biofertilizers; Rakshit, A., Meena, V.S., Parihar, M., Singh, H.B., Singh, A.K., Eds.; Woodhead Publishing: Sawston, UK, 2021; pp. 83–93. ISBN 9780128216675. [Google Scholar]
- Salmeron-Santiago, I.A.; Martínez-Trujillo, M.; Valdez-Alarcón, J.J.; Pedraza-Santos, M.E.; Santoyo, G.; Pozo, M.J.; Chávez-Bárcenas, A.T. An Updated Review on the Modulation of Carbon Partitioning and Allocation in Arbuscular Mycorrhizal Plants. Microorganisms 2022, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R. Mycorrhizal Fungi: Utilization in Agriculture and Forestry; BoD—Books on Demand: Norderstedt, Germany, 2021; ISBN 9781838819408. [Google Scholar]
- Boutaj, H.; Meddich, A.; Roche, J.; Mouzeyar, S.; el Modafar, C. The effects of mycorrhizal fungi on vascular wilt diseases. Crop Prot. 2022, 155, 105938. [Google Scholar] [CrossRef]
- Fujita, M.; Kusajima, M.; Fukagawa, M.; Okumura, Y.; Nakajima, M.; Akiyama, K.; Asami, T.; Yoneyama, K.; Kato, H.; Nakashita, H. Response of tomatoes primed by mycorrhizal colonization to virulent and avirulent bacterial pathogens. Sci. Rep. 2022, 12, 4686. [Google Scholar] [CrossRef] [PubMed]
- Dey, M.; Ghosh, S. Arbuscular mycorrhizae in plant immunity and crop pathogen control. Rhizosphere 2022, 22, 100524. [Google Scholar] [CrossRef]
- Guisande-Collazo, A.; González, L.; Souza-Alonso, P. Origin Makes a Difference: Alternative Responses of an AM-Dependent Plant to Mycorrhizal Inoculum from Invaded and Native Soils under Abiotic Stress. Plant Biol. 2022, 24, 417–429. [Google Scholar] [CrossRef]
- Bennett, A.E.; Groten, K. The Costs and Benefits of Plant–Arbuscular Mycorrhizal Fungal Interactions. Annu. Rev. Plant Biol. 2022, 73, 649–672. [Google Scholar] [CrossRef]
- Jajoo, A.; Mathur, S. Role of Arbuscular Mycorrhizal Fungi as an Underground Saviuor for Protecting Plants from Abiotic Stresses. Physiol. Mol. Biol. Plants 2021, 27, 2589–2603. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P.; Alyemeni, M.N.; Alsahli, A.A.; Ahmad, P. Arbuscular mycorrhiza in combating abiotic stresses in vegetables: An eco-friendly approach. Saudi J. Biol. Sci. 2021, 28, 1465–1476. [Google Scholar] [CrossRef]
- Mbodj, D.; Effa-Effa, B.; Kane, A.; Manneh, B.; Gantet, P.; Laplaze, L.; Diedhiou, A.G.; Grondin, A. Arbuscular mycorrhizal symbiosis in rice: Establishment, environmental control and impact on plant growth and resistance to abiotic stresses. Rhizosphere 2018, 8, 12–26. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Solís-Ramos, L.Y.; Coto-López, C.; Andrade-Torres, A. Role of Arbuscular Mycorrhizal Symbiosis in Remediation of Anthropogenic Soil Pollution. Symbiosis 2021, 84, 321–336. [Google Scholar] [CrossRef]
- Sudipti, A.; Kumar, A.; Ogita, S.; Yau, Y.-Y. (Eds.) Innovations in Environmental Biotechnology; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Dassen, S.; van der Putten, W.H.; De Deyn, G.B. Severance of arbuscular mycorrhizal fungal mycelial networks in restoration grasslands enhances seedling biomass. New Phytol. 2021, 232, 753–761. [Google Scholar] [CrossRef]
- Fernandes Figueiredo, A.; Boy, J.; Guggenberger, G. Common Mycorrhizae Network: A Review of the Theories and Mechanisms Behind Underground Interactions. Front. Fungal Biol. 2021, 2, 735299. [Google Scholar] [CrossRef]
- Semchenko, M.; Barry, K.E.; de Vries, F.T.; Mommer, L.; Moora, M.; Maciá-Vicente, J.G. Deciphering the role of specialist and generalist plant–microbial interactions as drivers of plant–soil feedback. New Phytol. 2022, 234, 1929–1944. [Google Scholar] [CrossRef]
- Cheng, J.-K.; Cao, M.-Y.; Yang, H.-R.; Yue, M.-F.; Xin, G.-R.; Chen, B.-M. Interactive effects of allelopathy and arbuscular mycorrhizal fungi on the competition between the invasive species Bidens alba and its native congener Bidens biternata. Weed Res. 2022, 1–9, in press. [Google Scholar] [CrossRef]
- Sepp, S.-K.; Davison, J.; Jairus, T.; Vasar, M.; Moora, M.; Zobel, M.; Öpik, M. Non-random association patterns in a plant–mycorrhizal fungal network reveal host–symbiont specificity. Mol. Ecol. 2019, 28, 365–378. [Google Scholar] [CrossRef]
- Prado-Tarango, D.E. Mycorrhizal Fungi on the Sagebrush Steppe: Benefits for Restoring Keystone Rangeland Plant Species; Oregon State University: Corvallis, OR, USA, 2021. [Google Scholar]
- Hernández, D.H.; Larsen, J.; González-Rodríguez, A.; Tapia-Torres, Y.; de la Barrera, E.; Luis; Eguiarte, E.; García-Oliva, F. Cooperation between Sporobolus airoides and associated arbuscular mycorrhizal fungi for phosphorus acquisition under drought conditions in an oligotrophic desert ecosystem. Rhizosphere 2020, 15, 100225. [Google Scholar] [CrossRef]
- Tiley, G.E.D. Biological Flora of the British Isles: Cirsium arvense (L.) Scop. J. Ecol. 2010, 98, 938–983. [Google Scholar] [CrossRef]
- Peeters, A.; Food and Agriculture Organization. Wild and Sown Grasses: Profiles of a Temperate Species Selection, Ecology, Biodiversity and Use; Food & Agriculture Organization: Rome, Italy, 2004; ISBN 9789251051597. [Google Scholar]
- Burescu, L.I.N.; Morar-Burescu, E.A.; Ștef, S.F.; Vlad, I.A.; Bartha, S.; Pop, I.F.; Bojinescu-Rostescu, I. Vegetation and productive potential of dominant grasslands by Festuca valesiaca and Agrostis capillaris in northwestern Romania. Rom. Agric. Res. 2021, 39, 1–14. [Google Scholar]
- Agrostis capillaris (Common Bent). Available online: https://www.cabi.org/isc/datasheet/3830?fbclid=IwAR1efyHa9vJsOtzE_r2_C5U6l6lF_uMTr92ntKIM0iPPsTUhCOXJeqs2Do (accessed on 20 May 2022).
- Yin, Y.; Yuan, Y.; Zhang, X.; Huhe; Cheng, Y.; Borjigin, S. Comparison of the Responses of Soil Fungal Community to Straw, Inorganic Fertilizer, and Compost in a Farmland in the Loess Plateau. Microbiol. Spectr. 2022, 10, e02230-21. [Google Scholar] [CrossRef]
- Emery, S.M.; Reid, M.L.; Bell-Dereske, L.; Gross, K.L. Soil mycorrhizal and nematode diversity vary in response to bioenergy crop identity and fertilization. GCB Bioenergy 2017, 9, 1644–1656. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.-A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Řezáčová, V.; Czakó, A.; Stehlík, M.; Mayerová, M.; Šimon, T.; Smatanová, M.; Madaras, M. Organic Fertilization Improves Soil Aggregation through Increases in Abundance of Eubacteria and Products of Arbuscular Mycorrhizal Fungi. Sci. Rep. 2021, 11, 12548. [Google Scholar] [CrossRef] [PubMed]
- Feilinezhad, A.; Mirzaeiheydari, M.; Babaei, F.; Maleki, A.; Rostaminya, M. The Effect of Tillage, Organic Matter and Mycorrhizal Fungi on Efficiency and Productivity Use of Nutrients in Maize. Commun. Soil Sci. Plant Anal. 2022, 53, 1–15. [Google Scholar] [CrossRef]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-El-Mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ben Ahmed, H.; Mitsui, T.; Baslam, M.; Meddich, A. The Native Arbuscular Mycorrhizal Fungi and Vermicompost-Based Organic Amendments Enhance Soil Fertility, Growth Performance, and the Drought Stress Tolerance of Quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef]
- Al-Maliki, S.; Al-zabee, M. Interactions between Biofertilizers and Chemical Fertilizers Affected Soil Biological Properties and Potato Yield. Euphrates J. Agric. Sci. 2019, 11, 1–13. [Google Scholar]
- Amir, H.; Gensous, S.; Cavaloc, Y.; Wantiez, L. Phosphorus Fertilization of an Ultramafic Soil Reduced Effects of Arbuscular Mycorrhizal Fungi but Not Mycorrhizal Colonization. J. Soil Sci. Plant Nutr. 2021, 21, 3544–3554. [Google Scholar] [CrossRef]
- Muneer, M.A.; Tarin, M.W.K.; Chen, X.; Afridi, M.S.; Iqbal, A.; Munir, M.Z.; Zheng, C.; Zhang, J.; Ji, B. Differential Response of Mycorrhizal Fungi Linked with Two Dominant Plant Species of Temperate Grassland under Varying Levels of N-Addition. Appl. Soil Ecol. 2022, 170, 104272. [Google Scholar] [CrossRef]
- McCain, K.N.S.; Wilson, G.W.T.; Blair, J.M. Mycorrhizal suppression alters plant productivity and forb establishment in a grass-dominated prairie restoration. Plant Ecol. 2011, 212, 1675–1685. [Google Scholar] [CrossRef]
- Zubek, S.; Kapusta, P.; Rożek, K.; Błaszkowski, J.; Gielas, I.; Nobis, M.; Świerszcz, S.; Nowak, A. Fungal root colonization and arbuscular mycorrhizal fungi diversity in soils of grasslands with different mowing intensities. Appl. Soil Ecol. 2022, 172, 104358. [Google Scholar] [CrossRef]
- Vaida, I. The Influence of Management on the Agronomic and Ecological Value of Mountain Grasslands. Ph.D. Thesis, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania, 2018. Available online: https://rei.gov.ro/ (accessed on 20 May 2022).
- Stoian, V.H.; Florian, V. Mycorrhiza–Benefits, Influence, Diagnostic Method. Bull. UASMV Agric. 2009, 66, 2009. [Google Scholar] [CrossRef]
- Stoian, V.; Vidican, R.; Crişan, I.; Puia, C.; Şandor, M.; Stoian, V.A.; Păcurar, F.; Vaida, I. Sensitive approach and future perspectives in microscopic patterns of mycorrhizal roots. Sci. Rep. 2019, 9, 10233. [Google Scholar] [CrossRef] [PubMed]
- Corcoz, L.; Păcurar, F.; Pop-Moldovan, V.; Vaida, I.; Stoian, V.; Vidican, R. Mycorrhizal Patterns in the Roots of Dominant Festuca rubra in a High-Natural-Value Grassland. Plants 2021, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Corcoz, L.; Păcurar, F.; Vaida, I.; Pleșa, A.; Moldovan, C.; Stoian, V.; Vidican, R. Deciphering the Colonization Strategies in Roots of Long-Term Fertilized Festuca rubra. Agronomy 2022, 12, 650. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio Inc., RStudio Team: Boston, MA, USA, 2019. [Google Scholar]
- RCore Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 1.9.12; Northwestern University: Evanston, IL, USA, 2019. [Google Scholar]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. 2019. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 20 May 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legrende, P.; McGlinn, D.; Wagner, H. 633 Vegan: Community Ecology Package; R Package Version 2.5-2; 2019; Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 May 2022).
- Wang, G.; Koziol, L.; Foster, B.L.; Bever, J.D. Microbial mediators of plant community response to long term N and P fertilization: Evidence of a role of plant responsiveness to mycorrhizal fungi. Glob. Chang. Biol. 2022, 28, 2721–2735. [Google Scholar] [CrossRef]
- Ning, Q.; Jiang, L.; Niu, G.; Yu, Q.; Liu, J.; Wang, R.; Liao, S.; Huang, J.; Han, X.; Yang, J. Mowing increased plant diversity but not soil microbial biomass under N-enriched environment in a temperate grassland. Plant Soil 2022, 1–13. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, K.; Wurzburger, N.; Zhang, J. Relationships between plant diversity and soil microbial diversity vary across taxonomic groups and spatial scales. Ecosphere 2020, 11, e02999. [Google Scholar] [CrossRef]
- Legay, N.; Grassein, F.; Binet, M.N.; Arnoldi, C.; Personeni, E.; Perigon, S.; Poly, F.; Pommier, T.; Puissant, J.; Clément, J.C.; et al. Plant species identities and fertilization influence on arbuscular mycorrhizal fungal colonisation and soil bacterial activities. Appl. Soil Ecol. 2016, 98, 132–139. [Google Scholar] [CrossRef]
- Shaji, H.; Chandran, V.; Mathew, L. Chapter 13—Organic Fertilizers as a Route to Controlled Release of Nutrients. In Controlled Release Fertilizers for Sustainable Agriculture; Lewu, F.B., Volova, T., Thomas, S., Rakhimol, K.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 231–245. ISBN 9780128195550. [Google Scholar] [CrossRef]
- Gryndler, M.; Larsen, J.; Hršelová, H.; Řezáčová, V.; Gryndlerová, H.; Kubát, J. Organic and Mineral Fertilization, Respectively, Increase and Decrease the Development of External Mycelium of Arbuscular Mycorrhizal Fungi in a Long-Term Field Experiment. Mycorrhiza 2006, 16, 159–166. [Google Scholar] [CrossRef]
- Joner, E.J. The Effect of Long-Term Fertilization with Organic or Inorganic Fertilizers on Mycorrhiza-Mediated Phosphorus Uptake in Subterranean Clover. Biol. Fertil. Soils 2000, 32, 435–440. [Google Scholar] [CrossRef]
- Bücking, H.; Liepold, E.; Ambilwade, P. The Role of the Mycorrhizal Symbiosis in Nutrient Uptake of Plants and the Regulatory Mechanisms Underlying These Transport Processes. In Plant Science; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Paparokidou, C.; Leake, J.R.; Beerling, D.J.; Rolfe, S.A. Phosphate availability and ectomycorrhizal symbiosis with Pinus sylvestris have independent effects on the Paxillus involutus transcriptome. Mycorrhiza 2021, 31, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Manoharan, L.; Rosenstock, N.P.; Olsson, P.A.; Hedlund, K. Long-term agricultural fertilization alters arbuscular mycorrhizal fungal community composition and barley (Hordeum vulgare) mycorrhizal carbon and phosphorus exchange. New Phytol. 2017, 213, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.D.; Jensen, A. Influence of vesicular-arbuscular mycorrhiza fungi on growth and uptake of various nutrients as well as uptake ratio of fertilizer P for lucerne (Medicago sativa). Plant Soil 1983, 70, 165–172. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi: A model and field test. New Phytol. 2002, 155, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Gollotte, A.; van Tuinen, D.; Atkinson, D. Diversity of Arbuscular Mycorrhizal Fungi Colonising Roots of the Grass Species Agrostis capillaris and Lolium perenne in a Field Experiment. Mycorrhiza 2004, 14, 111–117. [Google Scholar] [CrossRef]
- Olsen, E. Arbuscular mycorrhizal fungal communities and root colonisation differ in relation to plant host and soil nutrients. Fróskaparrit-Faroese Sci. J. 2017, 62, 85–101. [Google Scholar] [CrossRef]
- Weigelt, A.; King, R.; Bol, R.; Bardgett, R.D. Inter-specific variability in organic nitrogen uptake of three temperate grassland species. J. Plant Nutr. Soil Sci. 2003, 166, 606–611. [Google Scholar] [CrossRef]
- Fitter, A.H.; Graves, J.D.; Watkins, N.K.; Robinson, D.; Scrimgeour, C. Carbon transfer between plants and its control in networks of arbuscular mycorrhizas. Funct. Ecol. 1998, 12, 406–412. [Google Scholar] [CrossRef]
- Sheng, M.; Lalande, R.; Hamel, C.; Ziadi, N.; Shi, Y. Growth of Corn Roots and Associated Arbuscular Mycorrhizae Are Affected by Long-Term Tillage and Phosphorus Fertilization. Agron. J. 2012, 104, 1672–1678. [Google Scholar] [CrossRef]
- Saeed, M.; Khan, I.; Hameed, A.; Ullah, I.; Chaudhry, S.M.; Naveed-Ul-Haq, A.; Kaleem, S. Arbuscular mycorrhizal fungi (AMF) and soil chemical heterogeneity significantly alter nutritional value of tomato fruit. Pak. J. Bot. 2022, 54, 187–193. [Google Scholar] [CrossRef]
- Patreze, C.M.; Cordeiro, L. Nitrogen-Fixing and Vesicular–Arbuscular Mycorrhizal Symbioses in Some Tropical Legume Trees of Tribe Mimoseae. For. Ecol. Manag. 2004, 196, 275–285. [Google Scholar] [CrossRef]
- Islam, M.; Al-Hashimi, A.; Ayshasiddeka, M.; Ali, H.; Enshasy, H.A.E.; Dailin, D.J.; Sayyed, R.Z.; Yeasmin, T. Prevalence of Mycorrhizae in Host Plants and Rhizosphere Soil: A Biodiversity Aspect. PLoS ONE 2022, 17, e0266403. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mawdsley, J.L.; Edwards, S.; Hobbs, P.J.; Rodwell, J.S.; Davies, W.J. Plant species and nitrogen effects on soil biological properties of temperate upland grasslands. Funct. Ecol. 1999, 13, 650–660. [Google Scholar] [CrossRef]
- Aristizábal, C.; Rivera, E.L.; Janos, D.P. Arbuscular mycorrhizal fungi colonize decomposing leaves of Myrica parvifolia, M. pubescens and Paepalanthus sp. Mycorrhiza 2004, 14, 221–228. [Google Scholar] [CrossRef]
- Pietikäinen, A.; Kytöviita, M.M.; Vuoti, U. Mycorrhiza and seedling establishment in a subarctic meadow: Effects of fertilization and defoliation. J. Veg. Sci. 2005, 16, 175–182. [Google Scholar] [CrossRef]
- Johnson, N.C. Can fertilization of soil select less mutualistic mycorrhizae? Ecol. Appl. 1993, 3, 749–757. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant | Frequency (%) | Intensity (%) | Arbuscules (%) | Vesicles (%) | Non-Mycorrhizal Areas (%) | Colonization Degree (%) | Mycorrhizal/Non-Mycorrhizal Report | Arbuscules/Vesicles Report |
|---|---|---|---|---|---|---|---|---|
| V1 | 62.65 ± 0.95b | 32.81 ± 0.59b | 2.88 ± 0.26a | 1.53 ± 0.11b | 67.18 ± 0.59d | 24.8 ± 0.65b | 0.64 ± 0.02b | 0.50 ± 0.10a |
| V2 | 68.27 ± 0.92a | 38.45 ± 0.67a | 2.92 ± 0.25a | 2.32 ± 0.15a | 61.54 ± 0.67e | 31.0 ± 0.76a | 1.11 ± 0.09a | 0.54 ± 0.07a |
| V3 | 46.74 ± 1.01c | 22.05 ± 0.53c | 1.76 ± 0.21b | 0.58 ± 0.05cd | 77.94 ± 0.53c | 14.5 ± 0.51c | 0.36 ± 0.01c | 0.25 ± 0.09ab |
| V4 | 36.41 ± 1.12d | 18.59 ± 0.62d | 1.44 ± 0.17bc | 0.84 ± 0.08c | 81.39 ± 0.62b | 12.6 ± 0.60c | 0.34 ± 0.01c | 0.13 ± 0.04b |
| V5 | 30.25 ± 0.93e | 14.69 ± 0.45e | 0.68 ± 0.12c | 0.25 ± 0.03d | 85.30 ± 0.45a | 7.95 ± 0.38d | 0.21 ± 0.00c | 0.04 ± 0.02b |
| F test | 916.27 | 884.44 | 74.55 | 164.22 | 884.47 | 721.14 | 120.88 | 30.77 |
| p.val | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Parameter | Frequency | Intensity | Arbuscules | Vesicles | Non-Mycorrhizal Areas | Mycorrhizal/Non-Mycorrhizal Report | Colonization Degree | Arbuscules/Vesicles Report |
|---|---|---|---|---|---|---|---|---|
| Frequency | 0.90 | 0.23 | 0.31 | −0.90 | 0.36 | 0.88 | 0.12 | |
| Intensity | 0.22 | 0.34 | −1.00 | 0.53 | 0.97 | 0.11 | ||
| Arbuscules | −0.03 | −0.22 | 0.06 | 0.21 | 0.42 | |||
| Vesicles | −0.34 | 0.23 | 0.36 | 0.03 * | ||||
| Non-Mycorrhizal areas | −0.53 | −0.97 | −0.11 | |||||
| Mycorrhizal/non-mycorrhizal report | 0.55 | 0.04 | ||||||
| Colonization degree | 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corcoz, L.; Păcurar, F.; Pop-Moldovan, V.; Vaida, I.; Pleșa, A.; Stoian, V.; Vidican, R. Long-Term Fertilization Alters Mycorrhizal Colonization Strategy in the Roots of Agrostis capillaris. Agriculture 2022, 12, 847. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12060847
Corcoz L, Păcurar F, Pop-Moldovan V, Vaida I, Pleșa A, Stoian V, Vidican R. Long-Term Fertilization Alters Mycorrhizal Colonization Strategy in the Roots of Agrostis capillaris. Agriculture. 2022; 12(6):847. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12060847
Chicago/Turabian StyleCorcoz, Larisa, Florin Păcurar, Victoria Pop-Moldovan, Ioana Vaida, Anca Pleșa, Vlad Stoian, and Roxana Vidican. 2022. "Long-Term Fertilization Alters Mycorrhizal Colonization Strategy in the Roots of Agrostis capillaris" Agriculture 12, no. 6: 847. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12060847