
16S Amplicon Sequencing of Nitrifying Bacteria and Archaea Inhabiting Maize Rhizosphere and the Influencing Environmental Factors



Abstract
:1. Introduction
2. Materials and Method
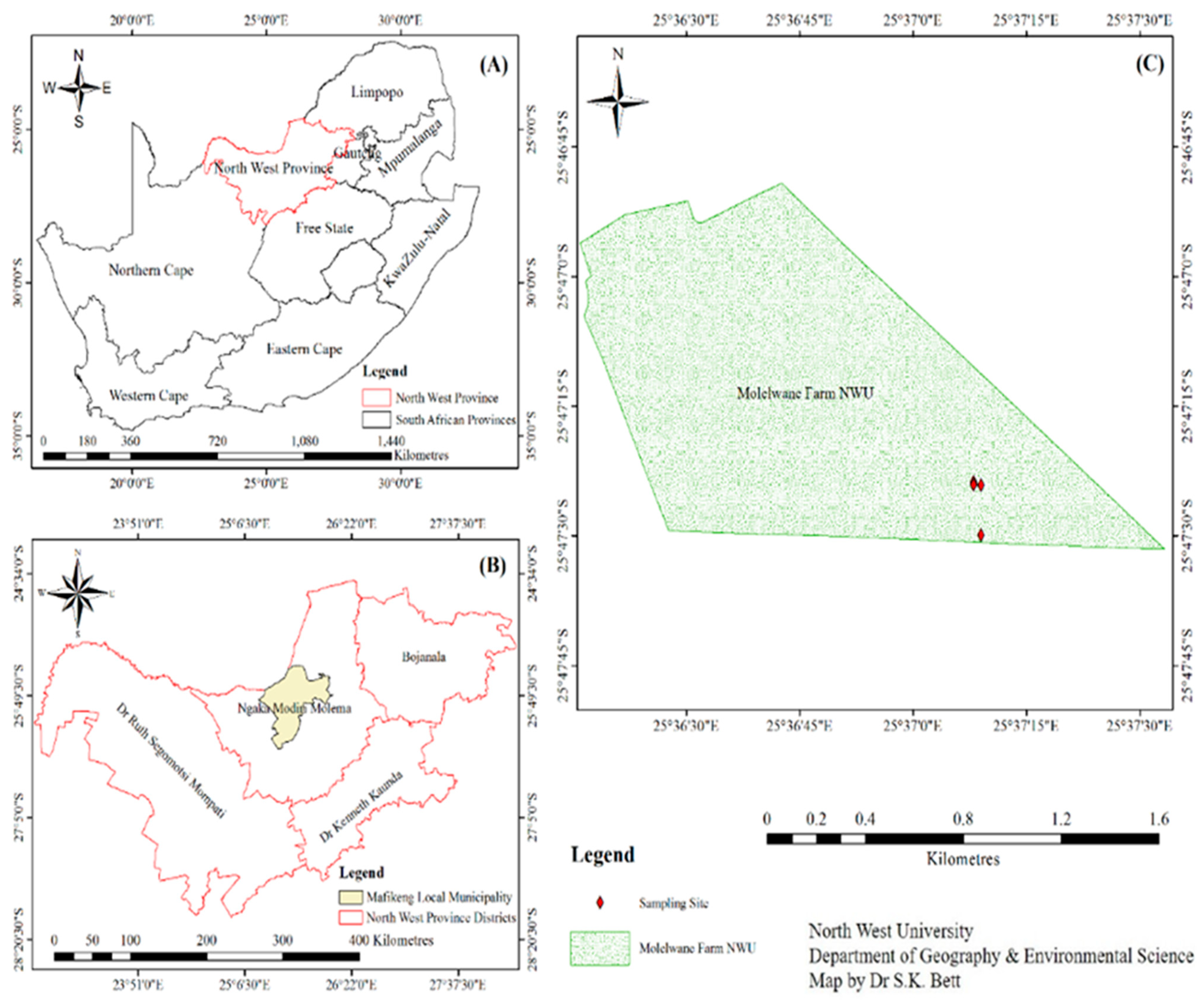
2.1. Sampling
2.2. Physio-Chemical Analysis of the Rhizosphere and Bulk Soil
2.3. DNA Extraction and 16S Amplicon Sequencing
2.4. Metagenome Assembly and Gene Annotation
2.5. Data and Statistical Analysis
3. Results
3.1. Rhizosphere Environmental Factors
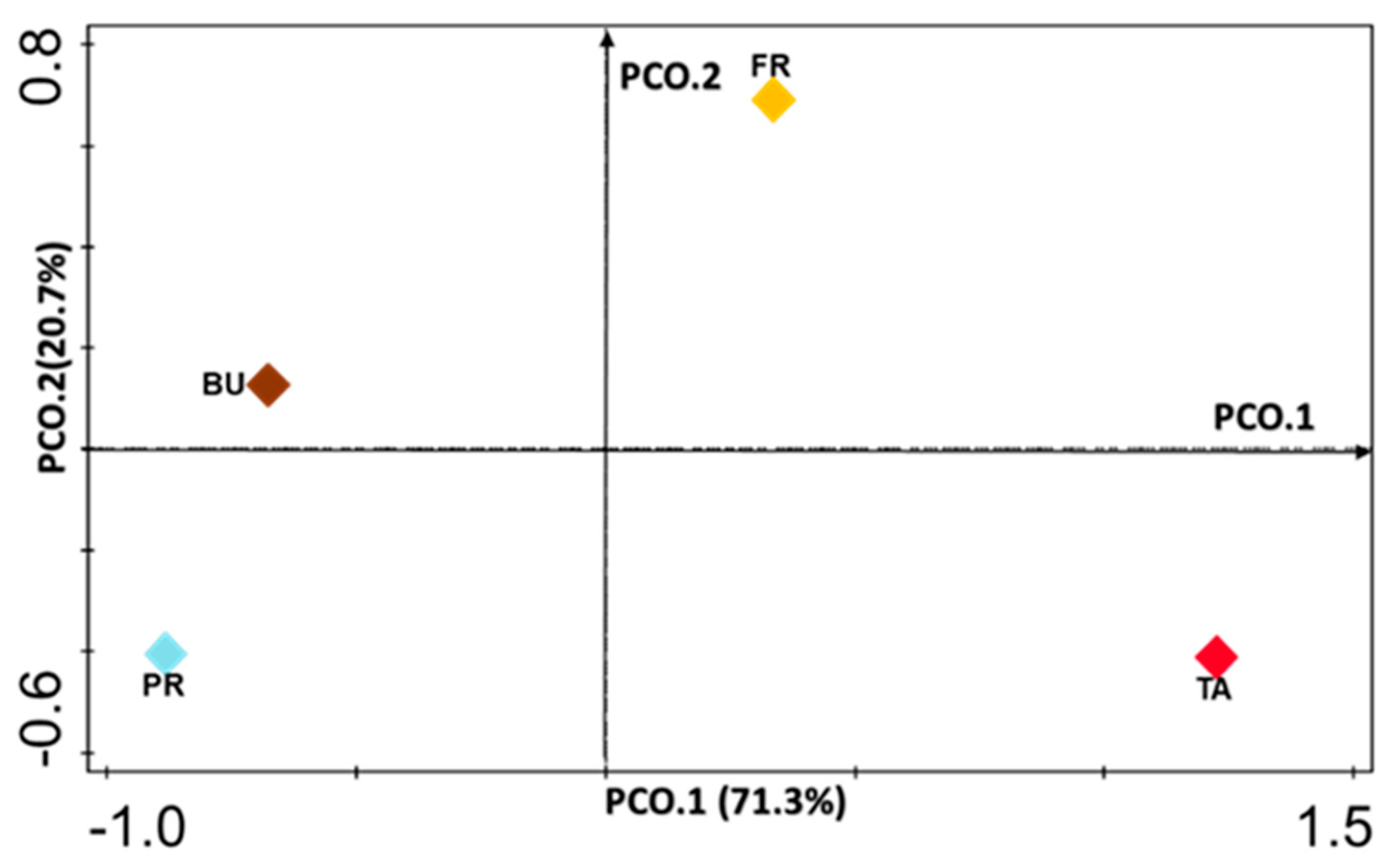
3.2. 16S Amplicon Sequencing of Maize Rhizosphere across Different Growth Stages
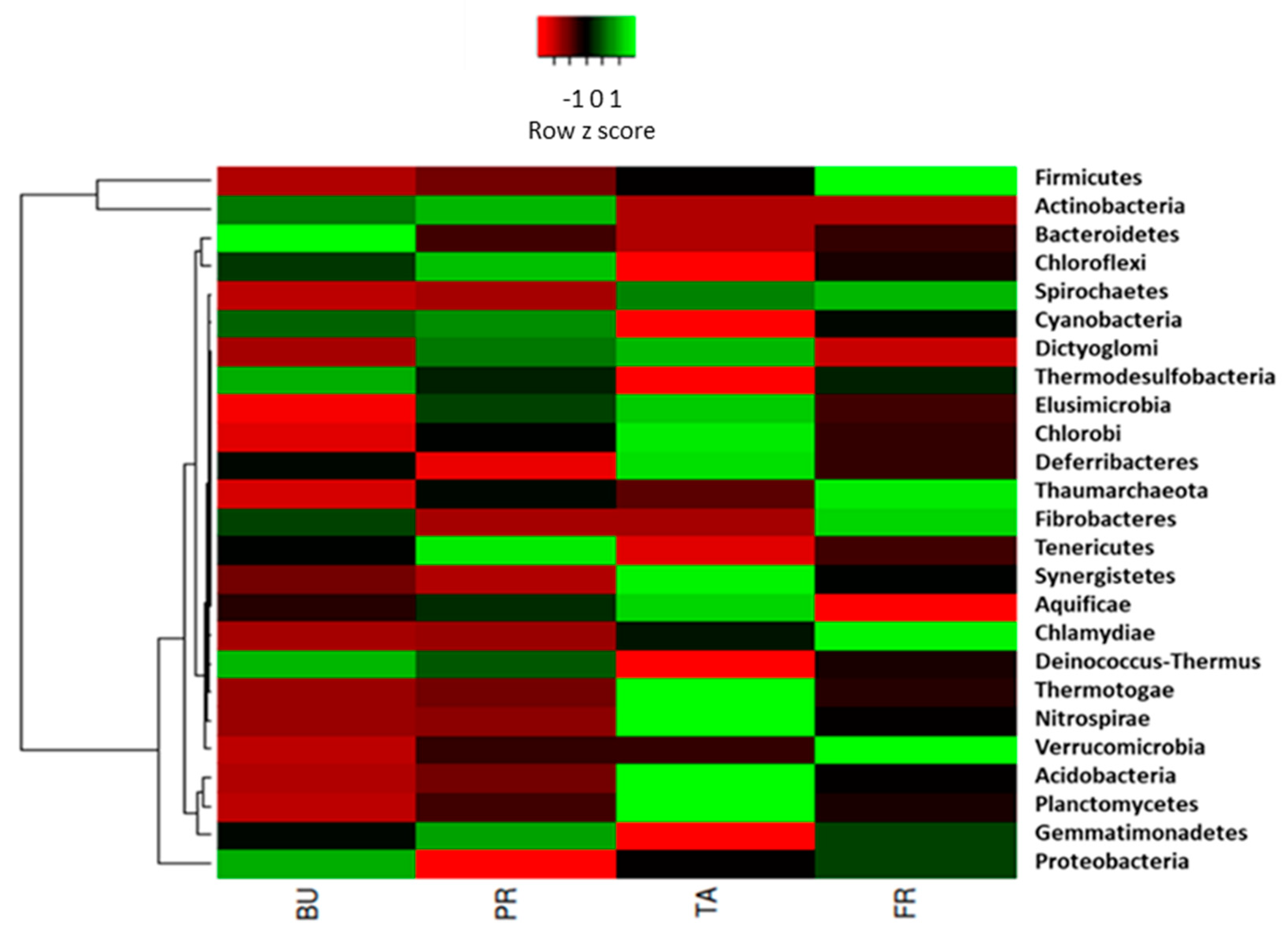
3.3. Taxonomic Profiling of Nitrifying Bacteria and Archaea Inhabiting Maize Rhizosphere across Different Vegetative Growth Stages
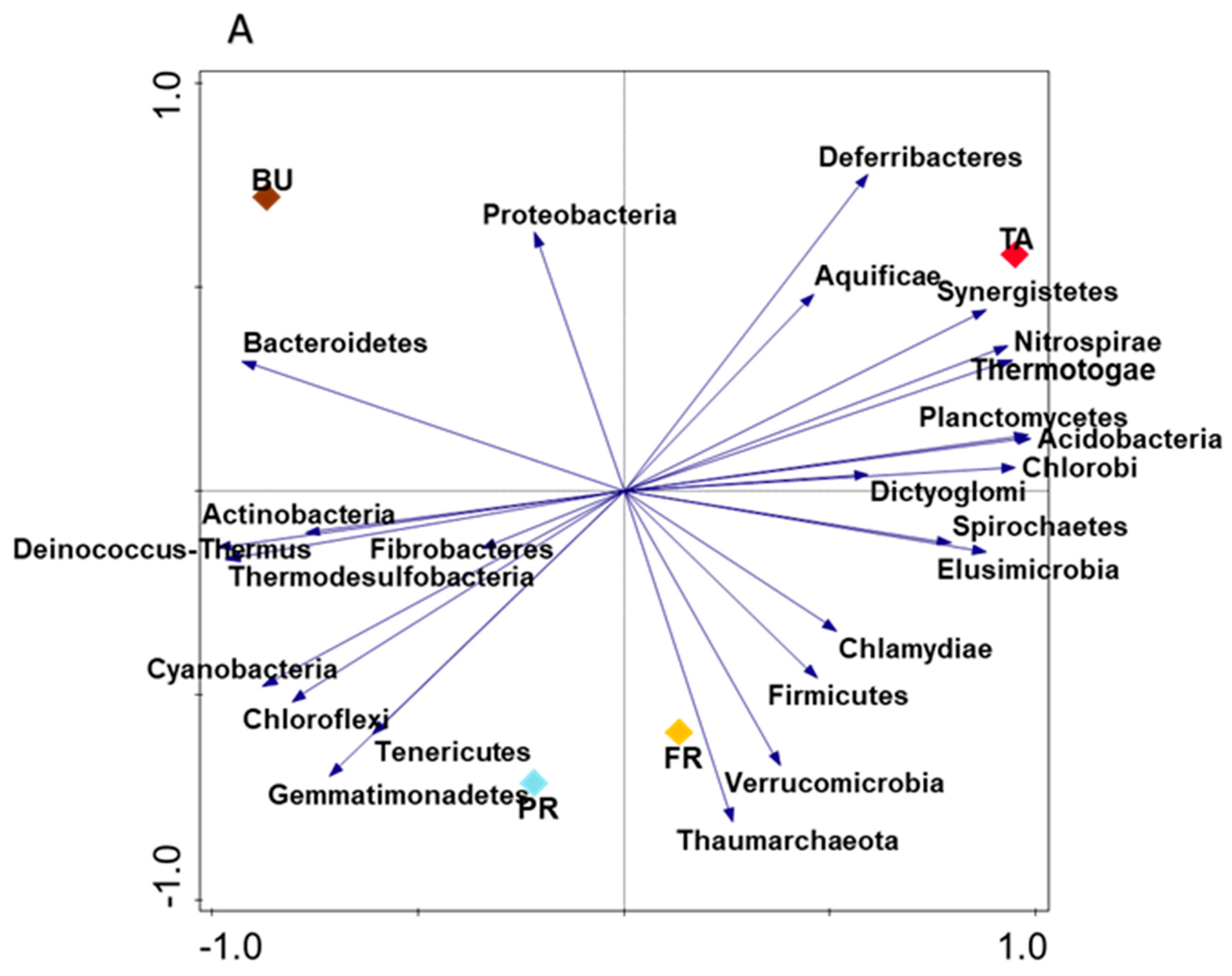
3.4. Relationship among Maize Rhizosphere Environmental Factors and Their Influence on Nitrifying Microorganism
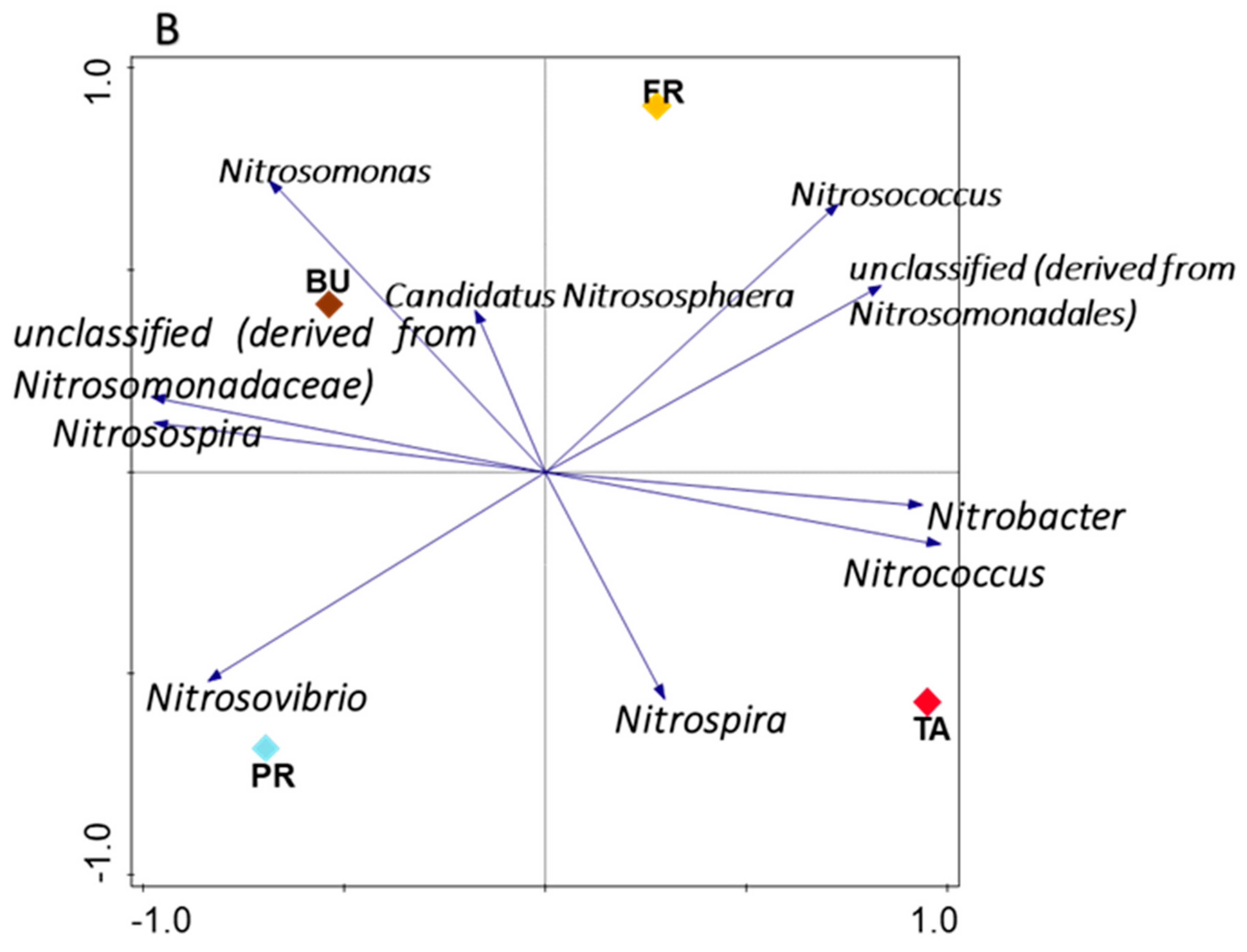
3.5. Influence of Maize Rhizosphere Environmental Factors on Nitrifying Bacteria and Archaea
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deurenberg, R.H.; Bathoorn, E.; Chlebowicz, M.A.; Couto, N.; Ferdous, M.; García-Cobos, S.; Kooistra-Smid, A.M.; Raangs, E.C.; Rosema, S.; Veloo, A.C.; et al. Application of next generation sequencing in clinical microbiology and infection prevention. J. Biotechnol. 2017, 243, 16–24. [Google Scholar] [CrossRef]
- Sharpton, T.J. An introduction to the analysis of shotgun metagenomic data. Front. Plant Sci. 2014, 5, 209. [Google Scholar] [CrossRef]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signaling in plants. J. Exp. Bot. 2016, 68, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Ayangbenro, A.S.; Olanrewaju, O.; Babalola, O.O.; Ayangbenro, A. Sulfate-Reducing Bacteria as an Effective Tool for Sustainable Acid Mine Bioremediation. Front. Microbiol. 2018, 9, 1986. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.-Y.; Ni, N.; Nkoh, J.N.; Li, J.-Y.; Xu, R.-K.; Qian, W. Beneficial dual role of biochars in inhibiting soil acidification resulting from nitrification. Chemosphere 2019, 234, 43–51. [Google Scholar] [CrossRef]
- Verma, N.; Chaudhary, S.; Goyal, S. Long Term Effects of Inorganic Fertilizers and Organic Amendments on Ammonification and Nitrification Activity of Soils under Cotton-Wheat Cropping System. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 718–724. [Google Scholar] [CrossRef]
- ZZhai, Y.; Zhao, X.; Teng, Y.; Li, X.; Zhang, J.; Wu, J.; Zuo, R. Groundwater nitrate pollution and human health risk assessment by using HHRA model in an agricultural area, NE China. Ecotoxicol. Environ. Saf. 2017, 137, 130–142. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Qiao, C.; Penton, C.R.; Xiong, W.; Liu, C.; Wang, R.; Liu, Z.; Xu, X.; Li, R.; Shen, Q. Reshaping the rhizosphere microbiome by bio-organic amendment to enhance crop yield in a maize-cabbage rotation system. Appl. Soil Ecol. 2019, 142, 136–146. [Google Scholar] [CrossRef]
- López-Carmona, D.A.; Alarcón, A.; Martinez-Romero, E.; Peña-Cabriales, J.J.; Larsen, J. Maize plant growth response to whole rhizosphere microbial communities in different mineral N and P fertilization scenarios. Rhizosphere 2019, 9, 38–46. [Google Scholar] [CrossRef]
- Tu, C.; He, T.; Lu, X.; Luo, Y.; Smith, P. Extent to which pH and topographic factors control soil organic carbon level in dry farming cropland soils of the mountainous region of Southwest China. Catena 2018, 163, 204–209. [Google Scholar] [CrossRef]
- Xiao, D.; Huang, Y.; Feng, S.; Ge, Y.; Zhang, W.; He, X.; Wang, K. Soil organic carbon mineralization with fresh organic substrate and inorganic carbon additions in a red soil is controlled by fungal diversity along a pH gradient. Geoderma 2018, 321, 79–89. [Google Scholar] [CrossRef]
- Li, X.; Cai, Y.; Liu, D.; Ai, Y.; Zhang, M.; Gao, Y.; Zhang, Y.; Zhang, X.; Yan, X.; Liu, B.; et al. Occurrence, fate, and transport of potentially toxic metals (PTMs) in an alkaline rhizosphere soil-plant (Maize, Zea mays L.) system: The role of Bacillus subtilis. Environ. Sci. Pollut. Res. 2019, 26, 5564–5576. [Google Scholar] [CrossRef] [PubMed]
- Kopáček, J.; Cosby, B.J.; Evans, C.D.; Hruška, J.; Moldan, F.; Oulehle, F.; Šantrůčková, H.; Tahovská, K.; Wright, R.F. Nitrogen, organic carbon and sulphur cycling in terrestrial ecosystems: Linking nitrogen saturation to carbon limitation of soil microbial processes. Biogeochemistry 2013, 115, 33–51. [Google Scholar] [CrossRef]
- Liu, G.; Du, Q.; Li, J. Interactive effects of nitrate-ammonium ratios and temperatures on growth, photosynthesis, and nitrogen metabolism of tomato seedlings. Sci. Hortic. 2017, 214, 41–50. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O. Biofertilizers and sustainable agriculture: Exploring arbuscular mycorrhizal fungi. Appl. Microbiol. Biotechnol. 2017, 101, 4871–4881. [Google Scholar] [CrossRef]
- Ayiti, O.E.; Ayangbenro, A.S.; Babalola, O.O. Relationship between nitrifying microorganisms and other microorganisms residing in the maize rhizosphere. Arch. Microbiol. 2022, 204, 1–11. [Google Scholar] [CrossRef]
- Kroetsch, D.; Wang, C. Particle size distribution. Soil Sampl. Methods Anal. 2008, 2, 713–725. [Google Scholar]
- Keeny, D.R.; Nelson, D.W. Nitrogen—Inorganic Forms; American Society of Soil Science Inc.: Madison, WI, USA, 1982; pp. 643–693. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter; American Societ of Agronomy: Madison, WI, USA, 1996. [Google Scholar]
- Santi, C.; Giacomo, C.; Luigi, P.D.A. Direct determination of organic carbon by dry combustion in soils with carbonates. Commun. Soil Sci. Plant Anal. 2006, 37, 155–162. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremme, J.M.; Mulvaney, C.S. Total Nitrogen. In Agronomy Monograph No. 9; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Smittenberg, J.; Harmsen, G.W.; Quispel, A.; Otzen, D. Rapid methods for determining different types of sulphur compounds in soil. Plant Soil 1951, 3, 353–360. [Google Scholar] [CrossRef]
- Sofie, T.; Michiel, O.D.B.; Bram, B.; Sascha, T.; Vincent, S.; Van, H.J.D.; Nele, W.; Jaco, V. Comparative evaluation of four bacteria-specific primer pairs for 16S rRNA gene surveys. Front. Microbiol. 2017, 8, 494. [Google Scholar] [CrossRef]
- Meyer, F.; Paarmann, D.; Souza, M.D.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kent, W.J. BLAT—The BLAST-Like Alignment Tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef]
- Wilke, A.; Harrison, T.; Wilkening, J.; Field, D.; Glass, E.M.; Kyrpides, N.; Mavrommatis, K.; Meyer, F. The M5nr: A novel non-redundant database containing protein sequences and annotations from multiple sources and associated tools. BMC Bioinform. 2012, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Paleontological Statistics Software Package for Education and Data Analysis. Palaentologia Electron. 2001, 4, 9. Available online: http://folk.uio.no/ohammer/past (accessed on 20 April 2021).
- Cajo, J.F.; Ter, B.; Petr, S. Software for Multivariate Data Exploration, Testing, and Summarization 1997. Available online: http://www.canoco5.com (accessed on 20 April 2021).
- USDA. Soil Survey Laboratory Methods Manual; United State of Agriculture, Natural Resource Conservation Service: Washington, DC, USA, 2014; p. 4.
- Burkitbayev, M.; Bachilova, N.; Kurmanbayeva, M.; Tolenova, K.; Yerezhepova, N.; Zhumagul, M.; Mamurova, A.; Turysbek, B.; Demeu, G. Effect of sulfur-containing agrochemicals on growth, yield, and protein content of soybeans (Glycine max (L.) Merr). Saudi J. Biol. Sci. 2021, 28, 891–900. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Q.; Hui, W.; Shi, J.; Lin, Q.; Chen, X.; Chen, Y. Effect of sulphur on soil Cu/Zn availability and microbial community composition. J. Hazard. Mater. 2008, 159, 385–389. [Google Scholar] [CrossRef]
- Kong, X.; Eriksen, J.; Petersen, S.O. Evaluation of the nitrification inhibitor 3,4-dimethylpyrazole phosphate (DMPP) for mitigating soil N2O emissions after grassland cultivation. Agric. Ecosyst. Environ. 2018, 259, 174–183. [Google Scholar] [CrossRef]
- Elrys, A.S.; Wang, J.; Metwally, M.A.S.; Cheng, Y.; Zhang, J.; Cai, Z.; Chang, S.X.; Müller, C. Global gross nitrification rates are dominantly driven by soil carbon-to-nitrogen stoichiometry and total nitrogen. Glob. Chang. Biol. 2021, 27, 6512–6524. [Google Scholar] [CrossRef] [PubMed]
- Doost, H.V.; Sharifi, R.S.; Farzaneh, S.; Panah, D.H. Effects of bio- and chemical-organic fertilizers on yield, some physiological traits and fatty acids composition of canola. Bangladesh J. Bot. 2019, 48, 113–122. [Google Scholar] [CrossRef]
- Vatandoost, H.; Seyed, S.R.; Kheirizadeh, A.Y. Effect of irrigation levels and plant growth promoting rhizobacteria on yield, some physiological and biochemical indices of rapeseed (Brassica napus L.). J. Crop Prod. Processing 2019, 9, 99–111. Available online: https://www.sid.ir/en/journal/ViewPaper.aspx?id=818546 (accessed on 16 May 2021).
- Zhalnina, K.V.; Dias, R.; Leonard, M.T.; de Quadros, P.D.; Camargo, F.A.O.; Drew, J.C.; Farmerie, W.G.; Daroub, S.H.; Triplett, E.W. Genome Sequence of Candidatus Nitrososphaera evergladensis from Group 1.1b Enriched from Everglades Soil Reveals Novel Genomic Features of The Ammonia Oxidizing Archaea. PLoS ONE 2014, 9, e101648. [Google Scholar] [CrossRef]
- Melnichuk, T.; Abdurashytov, S.; Andronov, E.; Abdurashytova, E.; Egovtseva, A.Y.; Gongalo, A.; Turin, E.; Pashtetskiy, V. The taxonomic structure of southern chernozem at the genus level influenced by microbial preparations and farming systems. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 6th International Conference on Agriproducts Processing and Farming, Voronezh, Russian Federation, 17–18 October 2019; IOP Publishing: Bristol, UK, 2020; p. 012101. [Google Scholar]
- Enebe, M.C.; Babalola, O.O. The influence of soil fertilization on the distribution and diversity of phosphorus cycling genes and microbes community of maize rhizosphere using shotgun metagenomics. Genes 2021, 12, 1022. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.M.; Hughes, D.J.; Fu, Q.; Abadie, M.; Hirsch, P.R. Metagenomic approaches reveal differences in genetic diversity and relative abundance of nitrifying bacteria and archaea in contrasting soils. Sci. Rep. 2021, 11, 1–9. [Google Scholar]
- Schaechter, M. Encyclopedia of Microbiology; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Fu, Q.; Abadie, M.; Blaud, A.; Carswell, A.; Misselbrook, T.H.; Clark, I.M.; Hirsch, P.R. Effects of urease and nitrification inhibitors on soil N, nitrifier abundance and activity in a sandy loam soil. Biol. Fertil. Soils 2020, 56, 185–194. [Google Scholar] [CrossRef]
- Wang, J.; Liu, L.; Gao, X.; Hao, J.; Wang, M. Elucidating the effect of biofertilizers on bacterial diversity in maize rhizosphere soil. PLoS ONE 2021, 16, e0249834. [Google Scholar] [CrossRef]
- Sun, P.; Zhao, Z.; Fan, P.; Chen, W.; Ruan, Y.; Wang, Q. Ammonia and Nitrite Oxidizing Bacteria are Dominant in Nitrification of Maize Rhizosphere Soil Following Combined Application of Biochar and Chemical Fertilizer. Front. Microbiol. 2021, 12, 715070. [Google Scholar] [CrossRef]
- Meier, M.A.; Lopez-Guerrero, M.G.; Guo, M.; Schmer, M.R.; Herr, J.R.; Schnable, J.C.; Alfano, J.R.; Yang, J. Rhizosphere Microbiomes in a Historical Maize-Soybean Rotation System Respond to Host Species and Nitrogen Fertilization at the Genus and Subgenus Levels. Appl. Environ. Microbiol. 2021, 87, e03132–e03220. [Google Scholar] [CrossRef]
- Stein, L.Y. Insights into the physiology of ammonia-oxidizing microorganisms. Curr. Opin. Chem. Biol. 2019, 49, 9–15. [Google Scholar] [CrossRef]
- Schlemper, T.R.; Leite, M.F.; Lucheta, A.R.; Shimels, M.; Bouwmeester, H.J.; van Veen, J.A.; Kuramae, E.E. Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Rocha, K.F.; Kuramae, E.E.; Borges, B.M.F.; Leite, M.F.A.; Rosolem, C.A. Microbial N-cycling gene abundance is affected by cover crop specie and development stage in an integrated cropping system. Arch. Microbiol. 2020, 202, 2005–2012. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Ke, M.; Lavoie, M.; Jin, Y.; Fan, X.; Zhang, Z.; Fu, Z.; Sun, L.; Gillings, M.; Peñuelas, J. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Xiao, Y.; Liu, Z.; Zhang, Y.; Wang, Y.; Yang, K. Trichoderma asperellum improves soil microenvironment in different growth stages and yield of maize in saline-alkaline soil of the Songnen Plain. Plant Soil Environ. 2020, 66, 639–647. [Google Scholar] [CrossRef]
- Fu, Q.; Xi, R.; Zhu, J.; Hu, H.; Xing, Z.; Zuo, J. The relative contribution of ammonia oxidizing bacteria and archaea to N2O emission from two paddy soils with different fertilizer N sources: A microcosm study. Geoderma 2020, 375, 114486. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Minimum | Maximum | Mean | Std. Deviation |
|---|---|---|---|---|
| SA | 84.00 | 86.00 | 85.00 | 1.16 |
| CL | 12.00 | 14.00 | 13.00 | 1.16 |
| pH | 5.35 | 6.22 | 5.93 | 0.41 |
| S | 246.00 | 576.00 | 336.50 | 159.85 |
| OC | 0.52 | 0.84 | 0.73 | 0.15 |
| TC | 0.52 | 0.89 | 0.73 | 0.15 |
| OM | 2.04 | 2.70 | 2.43 | 0.30 |
| TN | 0.06 | 0.09 | 0.08 | 0.01 |
| NH4 | 3.84 | 4.67 | 4.35 | 0.40 |
| NO3 | 4.02 | 9.76 | 6.12 | 2.72 |
| Diversity Indices | BU | PR | TA | FR | p-Value |
|---|---|---|---|---|---|
| Phylum | |||||
| Simpson_1-D | 0.71 ± 0.06 | 0.71 ± 0.07 | 0.74 ± 0.07 | 0.73 ± 0.06 | 0.99 |
| Shannon_H | 1.62 ± 0.19 | 1.61 ± 0.18 | 1.69 ± 0.19 | 1.69 ± 0.19 | |
| Evenness_e^H/S | 0.20 ± 0.10 | 0.21 ± 0.11 | 0.23 ± 0.10 | 0.22 ± 0.11 |
| Diversity Indices | BU | PR | TA | FR | p-Value |
|---|---|---|---|---|---|
| Genus | |||||
| Simpson_1-D | 0.52 ± 0.07 | 0.54 ± 0.11 | 0.51 ± 0.10 | 0.56 ± 0.10 | 0.99 |
| Shannon_H | 1.13 ± 0.19 | 1.19 ± 0.22 | 1.09 ± 0.20 | 1.21 ± 0.21 | |
| Evenness_e^H/S | 0.31 ± 0.10 | 0.33 ± 0.13 | 0.27 ± 0.12 | 0.28 ± 0.12 |
| Variables | SA | CL | pH | S | OC | TC | OM | TN | NH4 | NO3 | NB |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SA | 1 | ||||||||||
| CL | −1.00 | 1 | |||||||||
| pH | −0.81 | 0.81 | 1 | ||||||||
| S | −0.60 | 0.60 | 0.51 | 1 | |||||||
| OC | −0.84 | 0.84 | 0.10 | 0.54 | 1 | ||||||
| TC | −0.81 | 0.81 | 0.98 | 0.34 | 0.97 | 1 | |||||
| OM | −0.91 | 0.91 | 0.97 | 0.65 | 0.98 | 0.94 | 1 | ||||
| TN | −0.74 | 0.74 | 0.98 | 0.62 | 0.98 | 0.91 | 0.95 | 1 | |||
| NH4 | 0.27 | −0.27 | 0.06 | 0.53 | 0.06 | −0.13 | 0.04 | 0.27 | 1 | ||
| NO3 | −0.88 | 0.88 | 0.72 | 0.90 | 0.76 | 0.62 | 0.86 | 0.75 | 0.16 | 1 | |
| NB | 0.22 | −0.22 | −0.59 | 0.38 | −0.55 | −0.71 | −0.41 | −0.49 | 0.31 | 0.12 | 1 |
| Variables | SA | CL | pH | S | OC | TC | OM | TN | NH4 | NO3 |
|---|---|---|---|---|---|---|---|---|---|---|
| Nitrospira | −0.07 | 0.07 | −0.49 | 0.29 | −0.44 | −0.53 | −0.28 | −0.49 | −0.16 | 0.22 |
| Nitrosospira | −0.84 | 0.84 | 0.98 | 0.37 | 0.97 | 1.00 | 0.95 | 0.91 | −0.15 | 0.66 |
| unclassified (derived from Nitrosomonadales) | 0.97 | −0.97 | −0.65 | −0.51 | −0.68 | −0.67 | −0.78 | −0.55 | 0.43 | −0.82 |
| unclassified (derived from Nitrosomonadaceae) | −0.90 | 0.90 | 0.98 | 0.57 | 0.99 | 0.96 | 1.00 | 0.95 | −0.02 | 0.81 |
| Nitrobacter | 0.94 | −0.94 | −0.92 | −0.74 | −0.94 | −0.87 | −0.99 | −0.91 | −0.06 | −0.93 |
| Nitrosovibrio | −0.95 | 0.95 | 0.67 | 0.37 | 0.70 | 0.73 | 0.77 | 0.54 | −0.54 | 0.73 |
| Nitrosomonas | −0.41 | 0.41 | 0.84 | 0.54 | 0.82 | 0.74 | 0.76 | 0.92 | 0.56 | 0.52 |
| Candidatus Nitrososphaera | 0.20 | −0.20 | 0.33 | −0.50 | 0.28 | 0.41 | 0.11 | 0.30 | 0.01 | −0.41 |
| Nitrosococcus | 0.88 | −0.88 | −0.49 | −0.31 | −0.52 | −0.55 | −0.61 | −0.35 | 0.63 | −0.66 |
| Nitrococcus | 0.86 | −0.86 | −1.00 | −0.53 | −1.00 | −0.98 | −0.99 | −0.97 | −0.01 | −0.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayiti, O.E.; Ayangbenro, A.S.; Babalola, O.O. 16S Amplicon Sequencing of Nitrifying Bacteria and Archaea Inhabiting Maize Rhizosphere and the Influencing Environmental Factors. Agriculture 2022, 12, 1328. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12091328
Ayiti OE, Ayangbenro AS, Babalola OO. 16S Amplicon Sequencing of Nitrifying Bacteria and Archaea Inhabiting Maize Rhizosphere and the Influencing Environmental Factors. Agriculture. 2022; 12(9):1328. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12091328
Chicago/Turabian StyleAyiti, Oluwatobi Esther, Ayansina Segun Ayangbenro, and Olubukola Oluranti Babalola. 2022. "16S Amplicon Sequencing of Nitrifying Bacteria and Archaea Inhabiting Maize Rhizosphere and the Influencing Environmental Factors" Agriculture 12, no. 9: 1328. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12091328