
Analysis of Soil Fungal Community in Aged Apple Orchards in Luochuan County, Shaanxi Province

Abstract
:1. Introduction
2. Materials and Methods
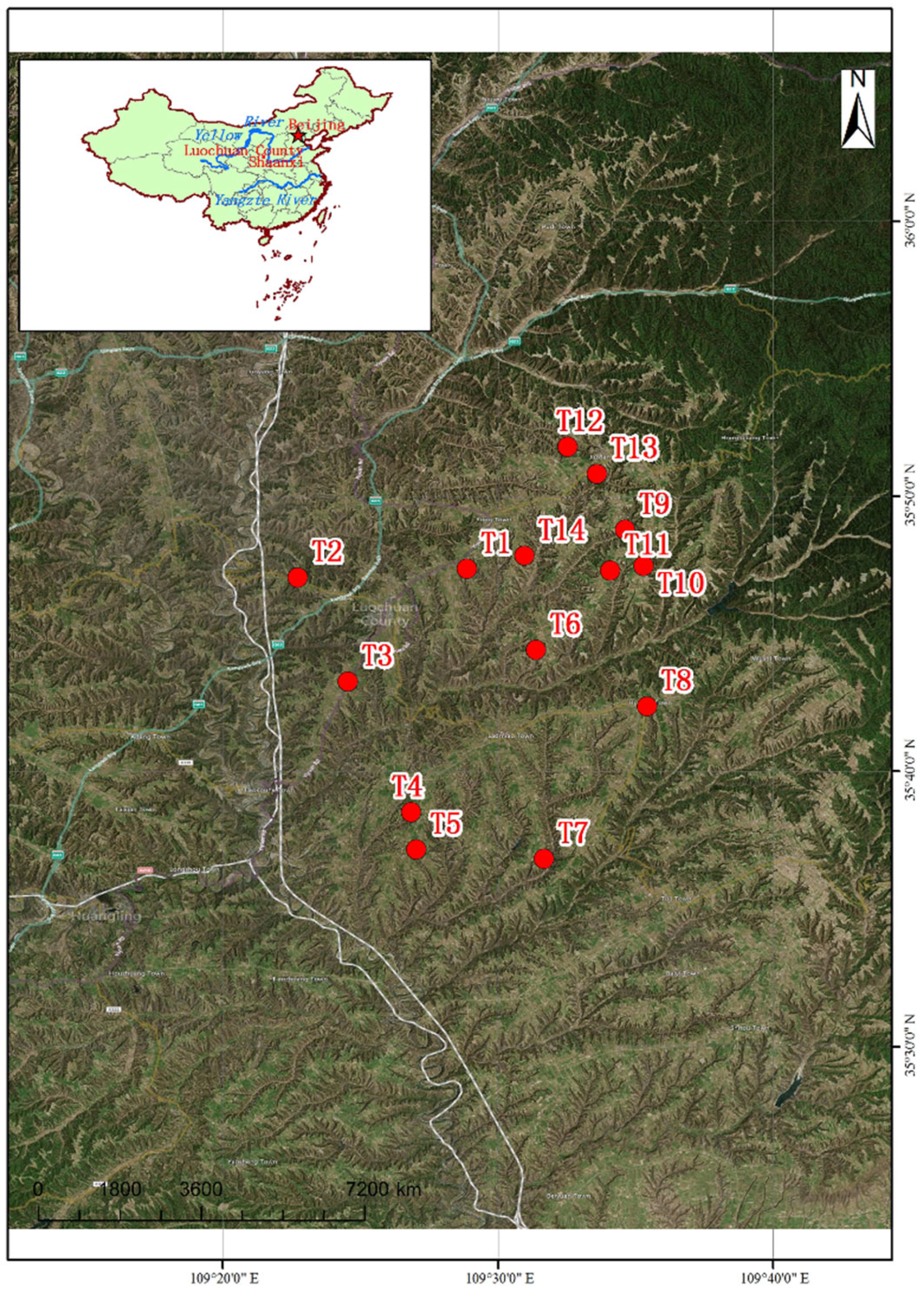
2.1. Soil Sample Collection and Processing
2.2. Determination of Soil Physical and Chemical Properties
2.3. High-Throughput Sequencing of Soil Fungi
2.3.1. DNA Extraction and PCR Amplification
2.3.2. Library Establishment and Illumina Sequencing
2.4. Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
3.1. Soil Physical and Chemical Properties
3.2. Analysis of Soil Fungal Community Structures in Different Aged Apple Orchards
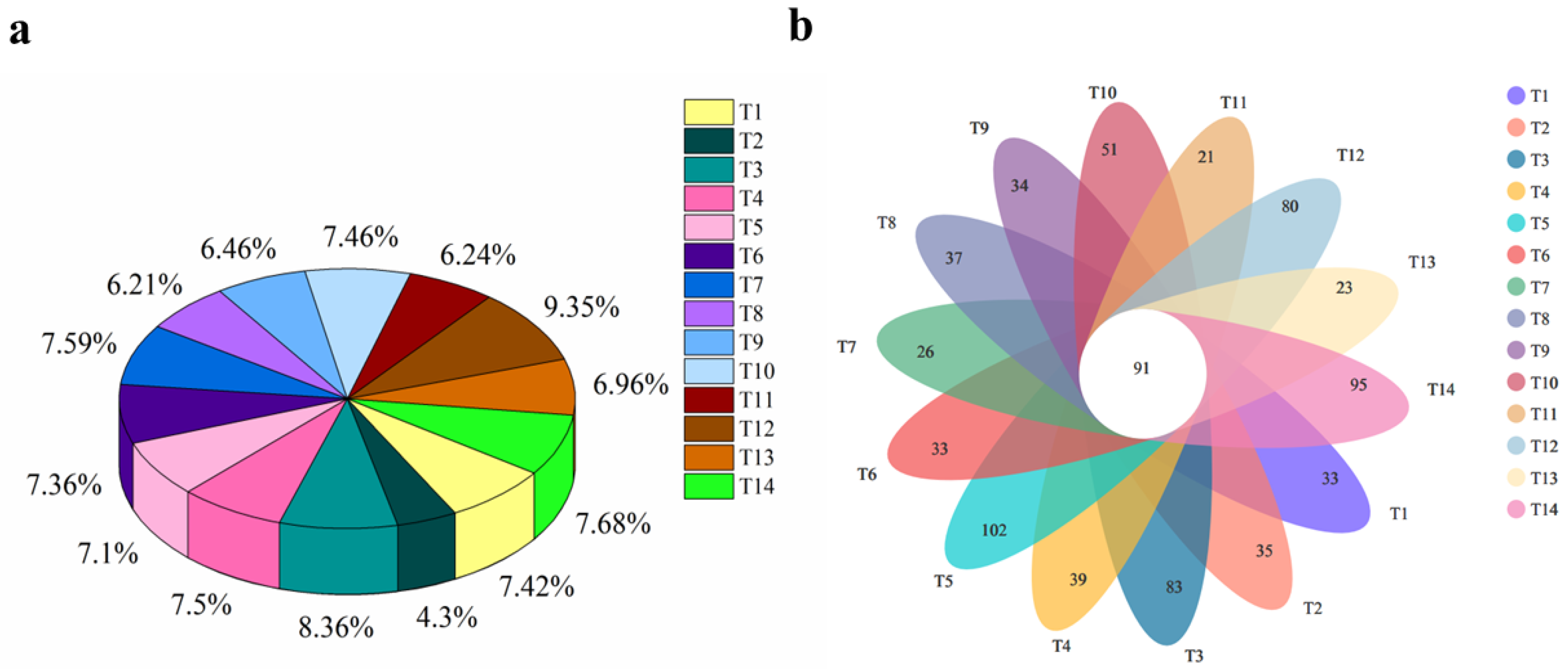
3.2.1. OTU Analysis of Soil Fungal Communities
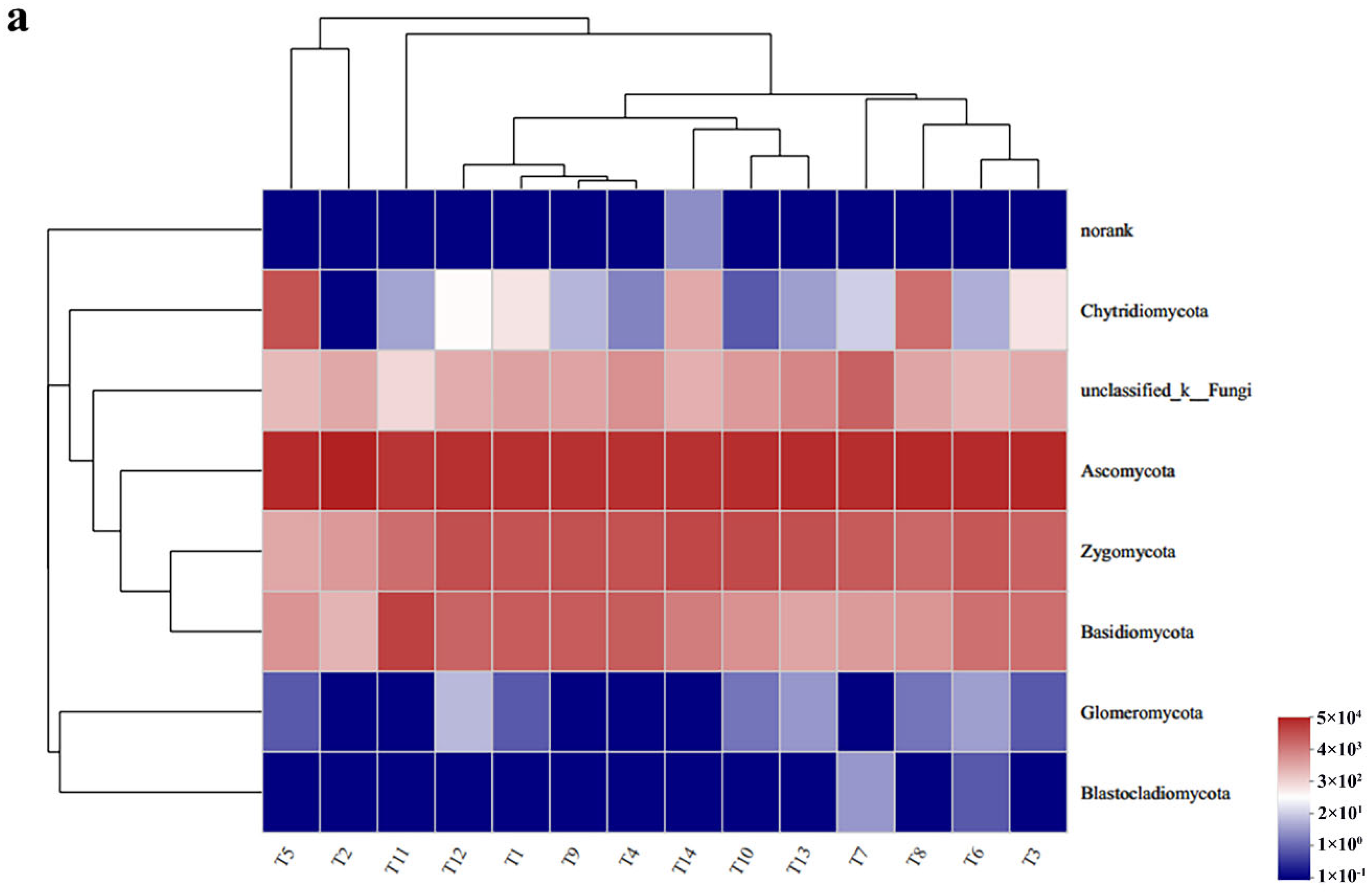
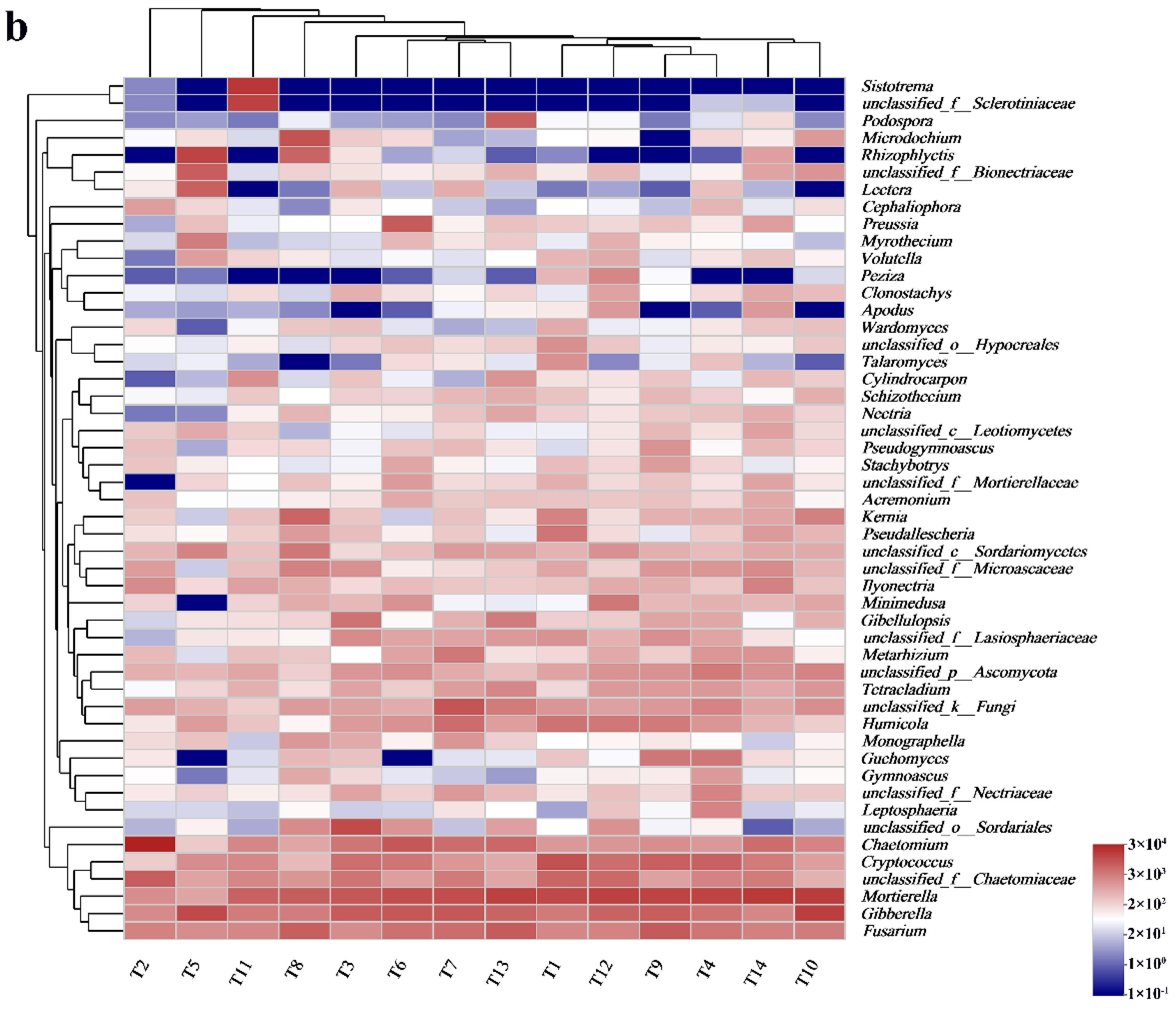
3.2.2. Soil Fungal Community Composition and Relative Abundance
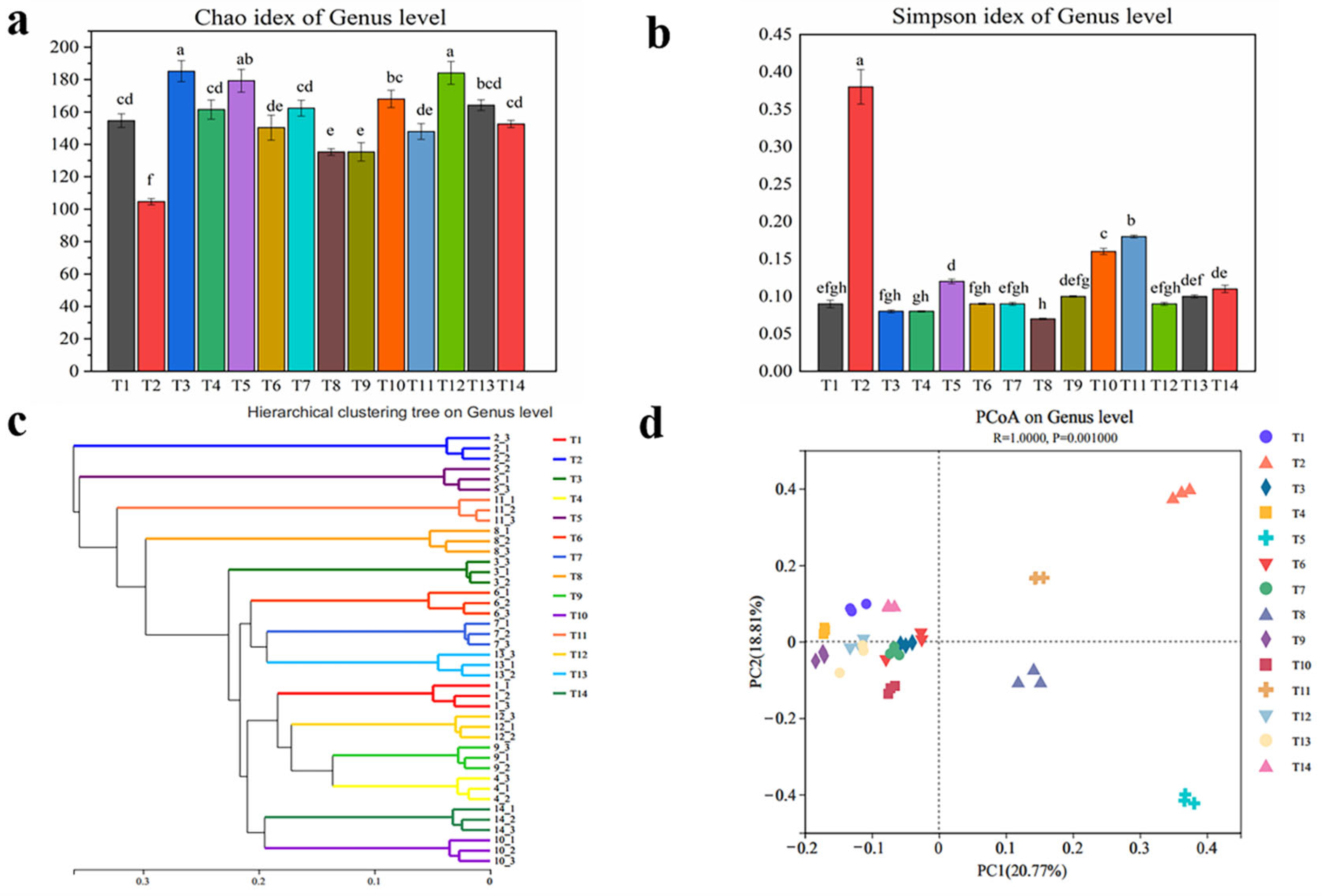
3.3. Analysis of Soil Fungal Community Diversity in Different Aged Apple Orchards
3.3.1. Alpha Diversity Analysis of Soil Fungal Communities
3.3.2. Cluster Analysis and PCoA Analysis of Soil Fungal Community
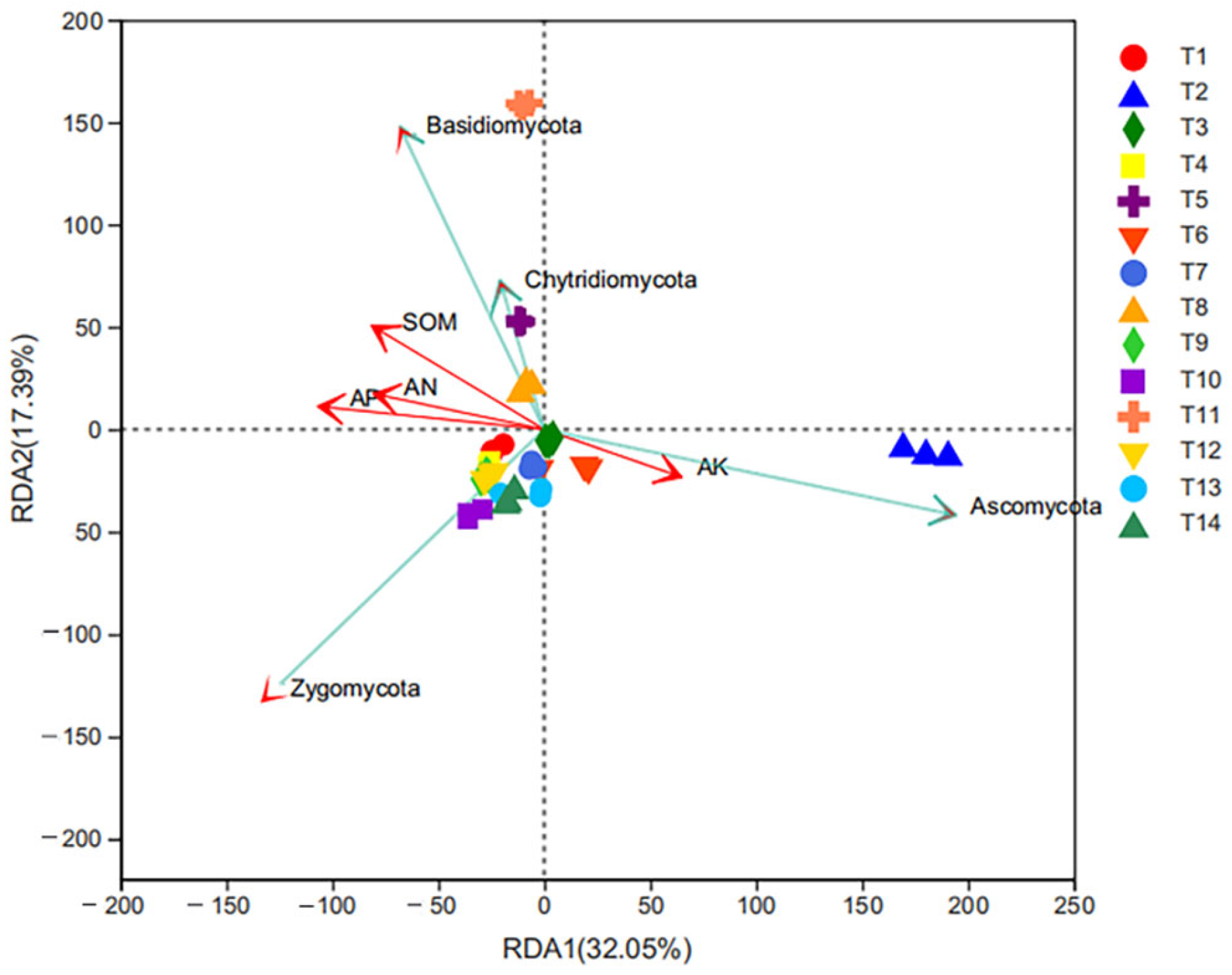
3.4. Correlation Analysis between Soil Fungal Community Structure and Soil Physicochemical Properties
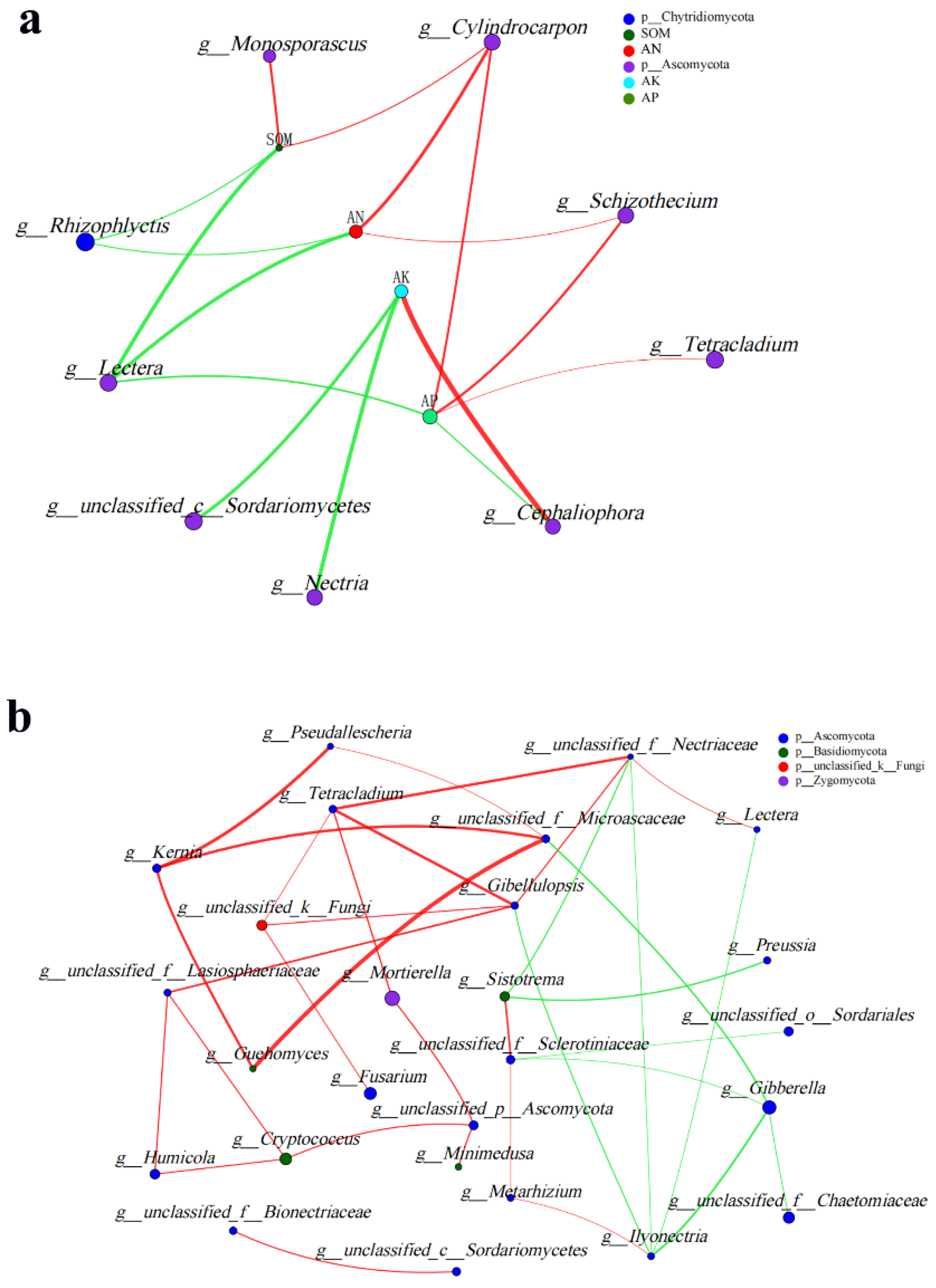
3.5. Soil Fungal Community Association Network Analysis
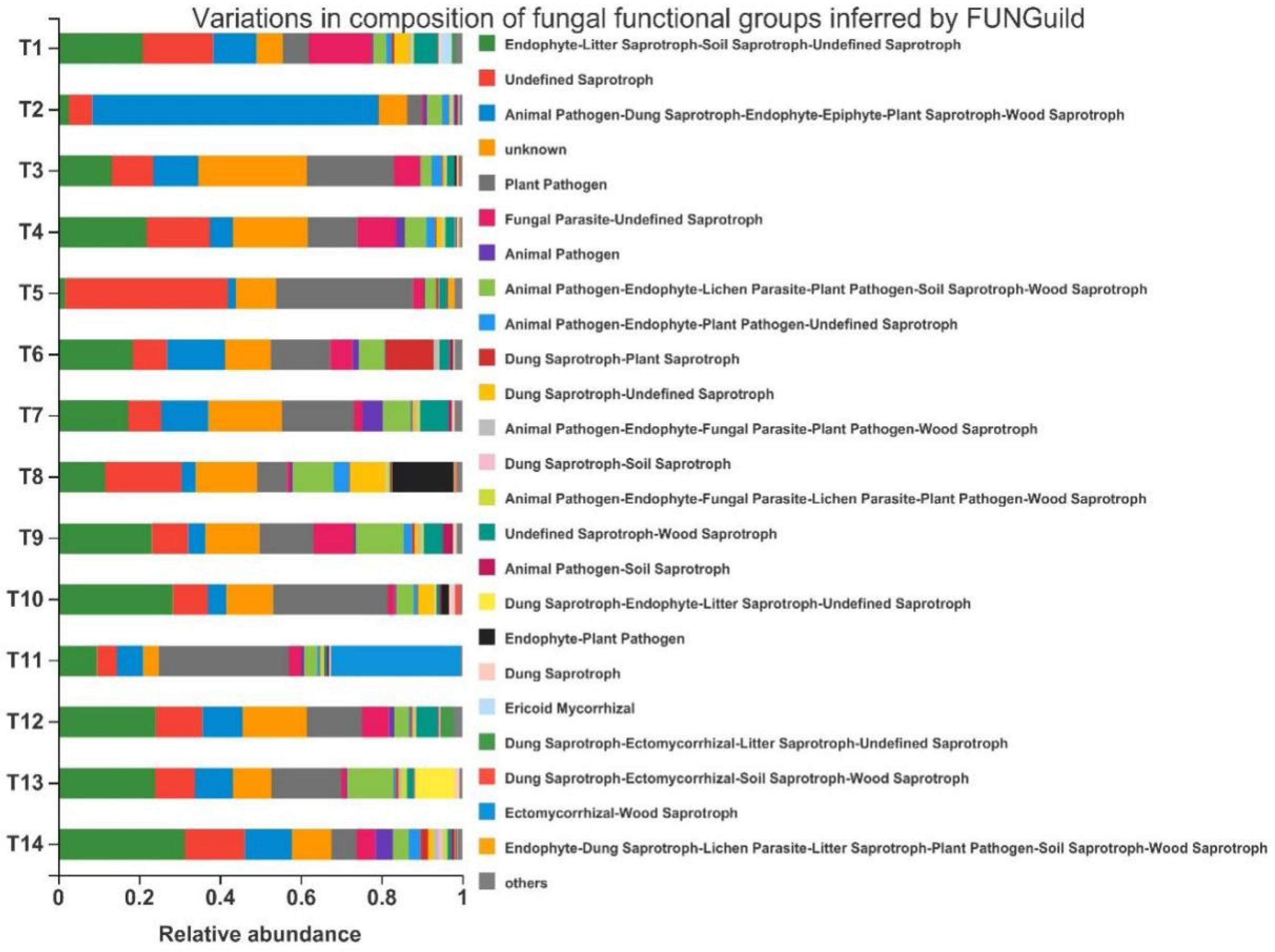
3.6. Functional Prediction Analysis of Soil Fungal Community Taxa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Q.J. Thoughts, and suggestions on the development of apple varieties in Luochuan, Shaanxi. Northwest Hortic. 2020, 5, 3–4. (In Chinese) [Google Scholar]
- Sun, C. Effects of Compound Microbial Fertilizer F25 on Soil Environment, Apple Yield and Quality in Weibei Apple Orchards; Northwest University: Xi’an, China, 2017; p. 72. [Google Scholar]
- Tian, J.; Sun, C.; Lu, P.P.; Li, F.; Shanguan, Y.Q.; Qi, F. Soil Biochemical Properties and Microbial Composition in Aged and Non-Aged Apple (Malus domestica) Orchards in Luochuan County, Loess Plateau, China. Soil Use Manag. 2020, 37, 879–890. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, C.; Li, L.X.; Yang, X.S.; Zhao, L.B.; Xia, S.J. Correlation of production constraints with the yield gap of apple cropping systems in Luochuan County, China. ScienceDirect 2019, 18, 1714–1725. [Google Scholar] [CrossRef]
- Xue, C. Manipulation of Microbial Community in Banana Rhizosphere to Suppress Fusarium wilt of Banana; Nanjing Agricultural University: Nanjing, China, 2015. (In Chinese) [Google Scholar]
- Yuan, J.J.; Zhao, Z.Y.; Zhang, L.S.; Hong, D.F.; Liu, W.L.; Shi, Q.F. Research on technique of inarching at bottom part of trunk with long shoot for renewal cultivar in old apple orchards of Loess Plateau. J. Northwest A F Univ. 2007, 35, 55–60. [Google Scholar]
- Dabrowska, G.; Baum, C.; Trejgell, A.; Hrynkiewicz, K. Impact of arbuscular mycorrhizal fungi on the growth and expression of gene encoding stress protein—Metallothionein BnMT2 in the non-host crop Brassica napus L. J. Plant Nutr. Soil Sci. 2014, 177, 459–467. [Google Scholar] [CrossRef]
- Li, M.S.; Wang, Y.L.; Yang, B.; Liu, F.T.; Zhang, Y.J.; Ren, T.B.; Zhang, F.J. Effect of biochar on community structure and function prediction of rhizosphere fungi in flue-cured tobacco at maturity. J. Agric. Resour. Environ. 2022, 39, 1041–1048. (In Chinese) [Google Scholar]
- Orgiazzi, A.; Lumini, E.; Nilsson, R.H.; Girlanda, M.; Vizzini, A.; Bonfante, P.; Bianciotto, V. Unravelling soil fungal communities from different mediterranean land-use backgrounds. PLoS ONE 2012, 7, e34847. [Google Scholar] [CrossRef]
- Liang, H.B.; Wang, X.W.; Yan, J.W.; Luo, L.X. Corrigendum: Characterizing the Intra-Vineyard Variation of Soil Bacterial and Fungal Communities. Front. Microbiol. 2019, 10, 1551. [Google Scholar] [CrossRef] [Green Version]
- Li, W.H.; Liu, Q.Z. Changes in fungal community and diversity in strawberry rhizosphere soil after 12 years in the greenhouse. J. Integr. Agric. 2019, 18, 199–209. [Google Scholar] [CrossRef]
- Pervaiz, Z.H.; Iqbal, J.; Zhang, Q.M.; Chen, D.M.; Wei, H.; Saleem, M. Continuous cropping alters multiple biotic and abiotic indicators of soil health. Soil Syst. 2020, 4, 59. [Google Scholar] [CrossRef]
- Kelderer, M.; Manici, L.M.; Caputo, F.; Thalheimer, M. Planting in the ‘inter-row’ to overcome replant disease in apple orchards: A study on the effectiveness of the practice based on microbial indicators. Plant Soil 2012, 357, 381–393. [Google Scholar] [CrossRef]
- Ma, X.X.; Zhang, X.H.; Huang, M.; Su, X.G.; Dong, B.; Zhang, X.Q.; Liang, X.Q.; Lin, Y.Y.; Zhuang, P.Y. Effects of continuous cropping of fruit trees on soil microbial diversity. Anhui Agric. Sci. 2021, 49, 15–18. (In Chinese) [Google Scholar]
- Yim, B.; Smalla, K.; Winkelmann, T. Evaluation of apple replant problems based on different soil disinfection treatments-links to soil microbial community structure? Plant Soil 2013, 366, 617–631. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, W.T.; Xu, S.Z.; Fan, H.; Chen, X.S.; Shen, X.; Yin, C.M.; Mao, Z.Q. An emerging chemical fumigant: Two-sided effects of dazomet on soil microbial environment and plant response. Environ. Sci. Pollut. Res. 2022, 29, 3022–3036. [Google Scholar] [CrossRef]
- Slykhuis, J.T.; Thomas, S.C.L. Response of apple seedlings to biocides and phosphate fertilizers in orchard soils in British Columbia. Can. J. Plant Pathol. 1985, 7, 294–301. [Google Scholar] [CrossRef]
- Munro, P.; Forge, T.A.; Jones, M.D.; Nelson, L.M. Soil biota from newly established orchards are more beneficial to early growth of cherry trees than biota from older orchards. Appl. Soil Ecol. 2020, 155, 103658. [Google Scholar] [CrossRef]
- Egidi, E.; Delgadobaquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M.; Manici, L.M. Apple Replant Disease: Role of Microbial Ecology in Cause and Control. Annu. Rev. Phytopathpl. 2012, 50, 45–65. [Google Scholar] [CrossRef]
- Sheng, Y.F.; Wang, H.Y.; Wang, M.; Li, H.H.; Xiang, L.; Pan, F.B.; Chen, X.S.; Shen, X.; Yin, C.M.; Mao, Z.Q. Effects of Soil Texture on the Growth of Young Apple Trees and Soil Microbial Community Structure under Replanted Conditions. Hortic. Plant J. 2020, 6, 123–131. [Google Scholar] [CrossRef]
- Ozimek, E.; Jaroszuk-Scisel, J.; Bohacz, J.; Kornillowicz-Kowalska, T.; Tyskiewicz, R.; Slomka, A.; Nowak, A.; Hanaka, A. Synthesis of Indoleacetic Acid, Gibberellic Acid and ACC-Deaminase by Mortierella Strains Promote Winter Wheat Seedlings Growth under Different Conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef] [Green Version]
- Arafat, Y.; Tayyab, M.; Khan, M.U.; Chen, T.; Amjad, H.; Awais, S.; Lin, X.M.; Lin, W.X.; Lin, S. Long-Term Monoculture Negatively Regulates Fungal Community Composition and Abundance of Tea Orchards. Agronomy 2019, 9, 466. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.M.; Wang, G.S.; Li, Y.Y.; Chen, X.S.; Wu, S.J.; Mao, Z.Q. T-RFLP analysis of soil fungi in continuous cropping apple orchard. J. Ecol. 2014, 3, 4837–4846. [Google Scholar]
- Wang, J.G.; Tian, T.; Wang, H.J.; Cui, J.; Shi, X.Y.; Song, J.H.; Li, T.S.; Li, W.D.; Zhong, M.T.; Zhang, W.X. Chitosan-coated compound fertilizer application and crop rotation alleviate continuouscotton cropping obstacles by modulating root exudates. Rhizosphere 2022, 23, 100581. [Google Scholar] [CrossRef]
- Sun, K.N.; Fu, L.Y.; Song, Y.; Yuan, L.; Zhang, H.R.; Wen, D.; Yang, N.; Wang, X.; Yue, Y.Q.; Li, X.H.; et al. Effects of continuous cucumber cropping on crop quality and soil fungal community. Environ. Monit. Assess. 2021, 193, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Liu, Q.Z.; Chen, P. Effect of long-term continuous cropping of strawberry on soil bacterial community structure and diversity. ScienceDirect 2018, 17, 2570–2582. [Google Scholar] [CrossRef]
- Li, X.Y.; Lewis, E.E.; Liu, Q.Z.; Li, H.Q.; Bai, C.Q.; Wang, Y.Z. Effects of long-term continuous cropping on soil nematode community and soil condition associated with replant problem in strawberry habitat. Sci. Rep. 2016, 6, 30466–30478. [Google Scholar] [CrossRef] [Green Version]
- Cheng, R.; Liu, K.S.; Dou, P.P.; Li, J.H.; Wang, K. The Changes in Soil Microorganisms and Soil Chemical Properties Affect the Heterogeneity and Stability of Soil Aggregates before and after Grassland Conversion. Agriculture 2022, 12, 307. [Google Scholar]
- Jiang, J.; Song, Z.; Yang, X.T.; Mao, Z.Q.; Nie, X.H.; Guo, H.; Peng, X.W. Microbial community analysis of apple rhizosphere around Bohai Gulf. Sci. Rep. 2017, 7, 8918. [Google Scholar] [CrossRef]
- Tao, Y.; Li, X.F.; Zhang, Z.Q.; Zhang, Q.Z.; Li, X. Effects of soil sterilization methods on soil nutrients and microbial community structure at different depths. J. Agric. Environ. Sci. 2022, 41, 575–584. [Google Scholar]
- Xiang, L.; Wang, M.; Jiang, W.T.; Wang, Y.F.; Chen, X.S.; Yin, C.M.; Mao, Z.Q. Key indicators for renewal and reconstruction of perennial trees soil: Microorganisms and phloridzin. Ecotoxicol. Environ. Saf. 2021, 225, 112723. [Google Scholar] [CrossRef]
- Arrigoni, E.; Albanese, D.; Longa, C.M.O.; Angeli, D.; Donati, C.; Ioriatti, C.; Pertot, I.; Perazzolli, M. Tissue age, orchard location and disease management influence the composition of fungal and bacterial communities present on the bark of apple trees. Environ. Microbiol. 2020, 22, 2080–2093. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.Z.; Wang, H.T.; Ma, F.W.; Qu, F.; Li, C.Y. Research Report on Fertilization Status of Apple Orchards in Shaanxi Province. China Fruit Tree 2021, 11, 89–94. [Google Scholar]
- Aparicio, V.; Costa, J.L. Soil quality indicators under continuous cropping systems in the Argentinean Pampas. Soil Tillage Res. 2007, 96, 155–165. [Google Scholar] [CrossRef]
- Chen, M.N.; Li, X.; Yang, Q.L.; Chi, X.Y.; Pan, L.J.; Chen, N.; Zhen, Y.; Wang, T.; Wang, M.; Yu, S.L. Soil eukaryotic microorganism succession as affected by continuous cropping of peanut-pathogenic and beneficial fungi were selected. PLoS ONE 2012, 7, e40659. [Google Scholar] [CrossRef] [Green Version]
- Finlay, B.J. Global dispersal of free-living microbial eukaryote species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Pan, F.J.; Han, X.Z.; Song, F.B.; Zhang, Z.M.; Yan, J.; Xu, L. A comprehensive analysis of the response of the fungal community structure to long-term continuous cropping in three typical upland crops. J. Integr. Agric. 2020, 19, 866–880. [Google Scholar] [CrossRef]
- Franke-Whittle, L.H.; Manici, L.M.; Insam, H.; Stres, B. Rhizosphere bacteria and fungi associated with plant growth in soils of three replanted apple orchards. Plant Soil 2015, 395, 317–333. [Google Scholar] [CrossRef]
- Ding, S.; Zhou, D.P.; Wei, H.W.; Wu, S.H.; Xie, B. Alleviating soil degradation caused by watermelon continuous cropping obstacle: Application of urban waste compost. Chemosphere 2021, 262, 128387. [Google Scholar] [CrossRef]
- Shen, Q.Q.; Yang, J.Y.; Su, D.F.; Li, Z.Y.; Xiao, W.; Wang, Y.X.; Cui, X.L. Comparative Analysis of Fungal Diversity in Rhizospheric Soil from Wild and Reintroduced Magnolia sinica Estimated via High-Throughput Sequencing. Plants 2020, 9, 600. [Google Scholar] [CrossRef]
- Yun, S.H.; Arie, T.; Kaneko, L.; Yoder, O.C.; Turgeon, B.G. Molecular organization of mating type loci in heterothallic, homothallic and asexual Gibberella/Fusarium species. Fungal Genet. Biol. 2000, 31, 7–20. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, Z.; Guo, Q.; Lei, B. First report of wilt of sugar beet caused by Gibellulopsis nigrescens in the Xinjiang region of China. Plant Dis. 2017, 101, 1318–1319. [Google Scholar] [CrossRef]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Surrey, UK, 1971; pp. 89–101. [Google Scholar]
- Wang, X.B.; Wang, G.S.; Liu, Y.S.; Chen, X.S.; Shen, X.; Yin, C.M.; Mao, Z.Q. Correlation analysis of apple replant disease and soil fungal community structure in the Northwest Loess Plateau area. Acta Ecol. Sin. 2018, 45, 855–864. (In Chinese) [Google Scholar]
- Zhao, L.; Wang, G.S.; Liu, X.; Chen, X.S.; Shen, X.; Yin, C.M.; Mao, Z.Q. Control of Apple Replant Disease Using Mixed Cropping with Brassica juncea or Allium fistulosum. Agriculture 2022, 12, 68. [Google Scholar] [CrossRef]
- Botha, A. The importance and ecology of yeasts in soil. Soil Biol. Biochem. 2010, 43, 1–8. [Google Scholar] [CrossRef]
- Yao, Q.; Xu, Y.X.; Liu, X.F.; Liu, J.J.; Huang, X.Y.; Yang, W.G.; Yang, Z.; Lan, L.; Zhou, J.M.; Wang, G.H. Dynamics of soil properties and fungal community structure in continuous-cropped alfalfa fields in Northeast China. PeerJ 2019, 7, e7127. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.N.; Zhou, Z.C.; Chen, T.; Wei, Y.Y.; Zheng, J.; Gao, R.X.; Chen, H. Biomarkers of antibiotic resistance genes during seasonal changes in wastewater treatment systems. Environ. Pollut. 2018, 234, 79–87. [Google Scholar] [CrossRef]
- Deng, L.T.; Tong, T.Q.; Ge, J.P. Redundancy analysis of soil microbial distribution and ecological factors in different orchards. Chin. Agron. Bull. 2017, 33, 41–47. (In Chinese) [Google Scholar]
- Li, L.; Xu, M.; Mohammad, E.A.; Zhang, W.J.; Duan, Y.H.; Li, D.C. Factors affecting soil microbial biomass and functional diversity with the application of organic amendments in three contrasting cropland soils during a field experiment. PLoS ONE 2018, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.N.; Mao, X.X.; Zhang, M.S.; Yang, W.; Di, H.J.; Li, M.; Liu, W.J.; Li, B. The application of Bacillus Megaterium alters soil microbial community composition, bioavailability of soil phosphorus and potassium, and cucumber growth in the plastic shed system of North China. Agric. Ecosyst. Environ. 2021, 307, 107236. [Google Scholar] [CrossRef]
- Zhao, F.; Zhao, M.Z.; Wang, Y.; Guan, L.; Pang, F.H. Study on microbial community structure and diversity in strawberry rhizosphere based on high throughput sequencing. Soil 2019, 51, 51–60. [Google Scholar]
- Xie, X.F.; Pu, L.J.; Wang, Q.Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total Environ. 2017, 607, 1419–1427. [Google Scholar] [CrossRef]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2014, 8, 952. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Ma, Y.J.; Wang, D.; Shan, Y.P.; Song, X.P.; Hu, H.Y.; Ren, X.L.; Ma, X.Y.; Cui, J.J.; Ma, Y. Integrated microbiology and metabolomics analysis reveal plastic mulch film residue affects soil microorganisms and their metabolic functions. J. Hazard. Mater. 2022, 423, 127258. [Google Scholar] [CrossRef]
- Lin, Q.; Li, L.J.; Adams, J.M.; Hedenec, P.; Tu, B.; Li, C.N.; Li, T.T.; Li, X.Z. Nutrient resource availability mediates niche differentiation and temporal co-occurrence of soil bacterial communities. Appl. Soil Ecol. 2021, 163, 103965. [Google Scholar] [CrossRef]
- Liu, Z.S.; Cichocki, N.C.; Hubschmann, T.; Suring, C.; Ofiteru, I.D.; Sloan, W.T.; Grimm, V.; Muller, S. Neutral mechanisms and niche differentiation in steady-state insular microbial communities revealed by single cell analysis. Environ. Microbiol. 2019, 21, 164–181. [Google Scholar] [CrossRef]
- Pozo, M.I.; Herrera, C.M.; Lachance, M.A.; Verstrepen, K.; Lievens, B.; Jacquemyn, H. Species coexistence in simple microbial communities: Unravelling the phenotypic landscape of co-occurring Metschnikowia species in floral nectar. Environ. Microbiol. 2016, 18, 1850–1862. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher-Jenull, J.; Ceccherini, M.T.; Pietramellara, G.; Renella, G.; Schloter, M. Beyond microbial diversity for predicting soil functions: A mini review. Pedosphere 2020, 30, 5–17. [Google Scholar] [CrossRef]
- Tan, Z.T.; Li, A.D.; Yang, R.; Cai, G.J.; Wu, F.C.; Guo, C.Y. Effects of passion fruit continuous cropping on soil fungal community structure. Mol. Plant Breed. 2022, 20, 10. (In Chinese) [Google Scholar]
- Cui, Y.F.; Huang, Y.; Jiang, L.H. Research progress of several important Gibberella fungi in agricultural production. Chin. Agron. Bull. 2007, 23, 441–446. (In Chinese) [Google Scholar]
- Wang, G.S.; Yin, C.M.; Pan, F.B.; Wang, X.B.; Li, X.; Wang, Y.F.; Wang, J.Z.; Tian, C.P.; Chen, J.; Mao, Z.Q. Analysis of the Fungal Community in Apple Replanted Soil Around Bohai Gulf. Hortic. Plant J. 2018, 4, 175–181. [Google Scholar] [CrossRef]
- Zhou, J.M.; Wang, X.E.; Song, R.; Cao, L.; Zhu, X.Q.; Xie, J.; Dai, Y.J. Review on biological mechanism and biological control technology of Lily continuous cropping obstacle. Hunan Agric. Sci. 2020, 11, 104–107. (In Chinese) [Google Scholar]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.B.; Zhang, C.Z.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 20, 1642–1651. [Google Scholar] [CrossRef]
- Peng, X.W.; Ding, L.; Zhang, L.; Li, Z.J.; Gu, M.Y. The Effect of antibiotic residue organic fertilizer on the community structure and functional groups of corn soil fungi. China Resour. Compr. Util. 2020, 38, 1–11. (In Chinese) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Organic Matter (SOM) (g/kg) | Available Nitrogen (AN) (mg/kg) | Available Phosphorus (AP) (mg/kg) | Available Potassium (AK) (mg/kg) |
|---|---|---|---|---|
| T1 | 1.48 ± 0.08 bcde | 39.32 ± 5.75 cde | 131.15 ± 31.23 d | 56.90 ± 10.02 b |
| T2 | 1.21 ± 0.29 e | 30.57 ± 4.08 ef | 4.55 ± 1.89 d | 53.14 ± 16.58 bcd |
| T3 | 1.25 ± 0.14 de | 34.65 ± 2.02 de | 2.03 ± 0.21 d | 55.18 ± 4.78 bc |
| T4 | 1.47 ± 0.07 cde | 31.15 ± 2.02 ef | 8.84 ± 6.07 d | 46.92 ± 5.28 bcd |
| T5 | 1.05 ± 0.26 e | 24.15 ± 2.02 f | 4.02 ± 2.96 d | 28.62 ± 5.72 bcd |
| T6 | 1.37 ± 0.15 de | 32.32 ± 4.67 ef | 18.52 ± 11.24 d | 121.19 ± 38.89 a |
| T7 | 1.24 ± 0.03 de | 32.32 ± 1.17 ef | 167.72 ± 1.39 bc | 15.14 ± 0.11 cd |
| T8 | 1.66 ± 0.01 bcd | 35.23 ± 0.58 de | 151.65 ± 0.98 cd | 14.50 ± 0.12 cd |
| T9 | 1.92 ± 0.02 ab | 47.48 ± 0.58 abc | 177.84 ± 3.58 abc | 16.33 ± 0.30 bcd |
| T10 | 2.33 ± 0.01 a | 51.57 ± 0.58 a | 199.47 ± 0.70 ab | 20.66 ± 0.53 bcd |
| T11 | 2.33 ± 0.02 a | 49.23 ± 0.58 ab | 205.68 ± 1.48 a | 20.42 ± 0.13 bcd |
| T12 | 1.88 ± 0.08 bc | 45.73 ± 0.58 abc | 196.22 ± 3.47 ab | 19.87 ± 0.05 bcd |
| T13 | 1.85 ± 0.06 de | 42.23 ± 0.58 bcd | 198.42 ± 2.17 ab | 17.44 ± 0.38 bcd |
| T14 | 1.37 ± 0.05 de | 35.23 ± 0.58 de | 157.56 ± 2.87 cd | 12.45 ± 0.12 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Jiang, W.; Wang, G.; Ding, F.; Li, Q.; Wang, R.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. Analysis of Soil Fungal Community in Aged Apple Orchards in Luochuan County, Shaanxi Province. Agriculture 2023, 13, 63. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13010063
Xu X, Jiang W, Wang G, Ding F, Li Q, Wang R, Chen X, Shen X, Yin C, Mao Z. Analysis of Soil Fungal Community in Aged Apple Orchards in Luochuan County, Shaanxi Province. Agriculture. 2023; 13(1):63. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13010063
Chicago/Turabian StyleXu, Xin, Weitao Jiang, Gongshuai Wang, Fengxia Ding, Qianjin Li, Ruolan Wang, Xuesen Chen, Xiang Shen, Chengmiao Yin, and Zhiquan Mao. 2023. "Analysis of Soil Fungal Community in Aged Apple Orchards in Luochuan County, Shaanxi Province" Agriculture 13, no. 1: 63. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13010063