Antioxidant Response to Salinity in Salt-Tolerant and Salt-Susceptible Cultivars of Date Palm




1
Department of Biology, College of Sciences, Sultan Qaboos University, P.O. Box 36, 123 Muscat, Oman
2
Department of Crop Sciences, College of Agricultural and Marine Sciences, Sultan Qaboos University, P.O. Box 34, 123 Muscat, Oman
*
Author to whom correspondence should be addressed.
Agriculture 2019, 9(1), 8; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010008
Submission received: 28 November 2018
/
Revised: 19 December 2018
/
Accepted: 22 December 2018
/
Published: 2 January 2019
(This article belongs to the Special Issue Response and Tolerance of Agricultural Crops to Salinity Stress)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The salinity tolerance mechanism in date palm through antioxidation has not been completely deciphered to date. Therefore, this study aimed to investigate the role of various antioxidants in salinity tolerance. Two date palm cultivars, contrasting in salinity tolerance, were used as model plants in a comparative study designed to detect changes in growth, Na+ and K+ uptake, reactive oxygen species (ROS), and antioxidant accumulations, when plants were exposed to salt stress. The results showed that salinity treatment had a more substantial negative effect on the growth and photosynthetic pigmentation of the susceptible ‘Zabad’ cultivar than on the tolerant ‘Umsila’ cultivar, probably due to the ability of ‘Umsila’ to accumulate less Na+ and more K+, to maintain a normal concentration of ROS and to produce more non-enzymatic antioxidants, including glutathione, phenolic compounds, flavonoids, and proline. Under salinity, ‘Umsila’ could also activate more superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX) than ‘Zabad’. These results suggest that the tolerance of ‘Umsila’ is partially due to the balanced Na+ and K+ uptake and to the relatively high concentration of ROS-scavenging metabolites. Together, these results indicate that the antioxidant mechanism is crucial for salinity tolerance in date palms. However, other mechanisms may also be involved in this trait.
1. Introduction
Soil salinization is a global problem, damaging agricultural lands mainly due to improper irrigation [1]. In Oman, particularly in Al Batinah region, slight soil salinization has given rise to the need to grow salt-tolerant crops, such as date palm. Although date palm is classified as a salt-tolerant plant, differences in tolerance have been observed among different cultivars [2,3,4,5,6]. Ramoliya and Pandey [7] reported that some date palm cultivars can tolerate high levels of soil salinity, up to 12.8 dSm−1, without showing significant growth defects. However, excessive salts in the soil can impair growth.
Plants can tolerate stress triggered by soil salinity by producing antioxidant agents [8]. In addition, salt tolerance in plants can occur through mechanisms such as osmotic adjustment and the coordination between water, potassium ions (K+), and some compatible solutes at the cellular level. Salinity tolerance can also occur via sodium ion (Na+) exclusion via the xylem stream or the apoplast barrier of the root or shoot tissues and the tolerance of tissues to a higher Na+ influx [9,10]. Plants can adopt other possible mechanisms, such as avoidance through anatomical changes in the root or leaf tissues or accumulative metabolites that can cause changes in specific metabolic pathways under salt stress [11,12,13].
All these mechanisms involve gene expression alterations [14,15], epigenetic changes [16,17,18], and community changes in the microbes associated with the plants [19,20,21,22,23,24].
In plants, salinity causes oxidative stress and subsequently leads to the production of reactive oxygen species (ROS) or free radicals, such as superoxide (O2•−), singlet oxygen (1O), and hydrogen peroxide (H2O2) [25]. Excessive salinity disturbs the osmotic balance and the water status, resulting in stomatal closure, which decreases the CO2/O2 ratio and subsequently reduces plant growth [26] through over-reduction of the electron transport chain (ETC), which leads to an unbalanced ATP/NADPH ratio [27,28]. This procedure induces excessive electron transfer to molecular oxygen and greatly reduces the ROS generated mostly in Photosystem I (PSI) and Photosystem II (PSII), or in the plastoquinone pool (PQ pool) [29]. These radicals can cause serious damage to cellular components and DNA and may subsequently lead to metabolic dysfunction and plant growth impairment [30,31].
Plants tolerate oxidative stress caused by salinity through antioxidant mechanisms [32,33,34]. These are subdivided into enzymatic and non-enzymatic antioxidant mechanisms. A number of non-enzymatic antioxidants have been well characterized; these enzymes catalyze redox reactions and rely on electron donation via reduction of low-molecular-weight antioxidants, such as glutathione, flavonoids, phenols and free proline [25,35]. Enzymatic antioxidants, such as superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX), have been previously reported as the major antioxidant enzymes that serve as an antioxidant defense system in many plants under salt stress [36,37,38,39].
Several previous publications reported the positive effects of enzymatic and non-enzymatic agents on salinity tolerance in plant species, including halophytes. For example, in a study on the response of the tolerant sapodilla rootstock (Manilkara zapota (L.) P.Royen) to salinity, significant increases in CAT and APX activities were observed when the rootstocks were exposed to 12 dSm−2 of diluted seawater [40]. SOD and CAT activities increased significantly in the leaves of olive (Olea europaea L.) subjected to 200 mM NaCl, suggesting a key role for antioxidants under salt stress [41]. In another report, it was demonstrated that there was a significant increase in SOD, CAT, and APX activities in the Chinese bayberry tree (Myrica rubra Siebold and Zucc.) when grown under oxidative stress [42]. Similarly, the non-enzymatic role of proline was increased in grapevine (Vitis vinifera L.) [43], and in citrus rootstock Carrizo citrange, when grown under saline conditions [44].
However, little research has been conducted on the role of antioxidant agent activity in salinity tolerance in date palms [45]. Therefore, the present study was carried out to investigate the role of antioxidant agents in the salinity tolerance mechanisms of date palm. In this study two date palm cultivars with contrasting salinity tolerance were used, and the enzymatic and non-enzymatic antioxidants were quantified in leaf and root tissues, in response to salinity treatment. The results showed that antioxidant agents play a crucial role in salinity tolerance by increasing their accumulation in salinity-tolerant cultivars.
2. Materials and Methods
2.1. Seed Germination and Plant Growth Measurements
The two date palm cultivars (Phoenix dactylifera L.), ‘Umsila’ (salt tolerant) and ‘Zabad’ (salt susceptible), were chosen based on previous observations and screening of various local date palm cultivars for salinity tolerance [6]. The experiments were carried out at the Agricultural Experimental Station (AES) of Sultan Qaboos University.
The seeds were germinated in sterilized moist vermiculite (no salt added) at 35 °C. After radicle emergence, the seeds were transferred into 2-L pots filled with sand containing 0.5 g NPK (20:5:10) slow-release fertilizer. The fertilizer included essential macronutrients, such as calcium and magnesium, together with ferrous and micronutrient traces. The pots were irrigated to field capacity with distilled water (0 mM NaCl) for the control treatment, or with NaCl solution at 240 mM for the salt treatment. Initially, all seedlings were irrigated with distilled water for 10 days, then the concentration of salt in the solution was gradually increased every week for five weeks until it reached the final concentration of 240 mM NaCl. The pots were placed in a glasshouse under natural sunlight conditions, and the plants were grown at 30 °C. Each treatment had three biological replicates. The selection of 240 mM NaCl concentration was based on the observations obtained from a previous study on screening of date palm cultivars for salinity tolerance [6]. This concentration was the threshold for non-lethal stress for these cultivars.
For growth measurement, the plants were separated into leaves and roots and their fresh weights were recorded. Subsequently, the samples were dried at 80 °C in an oven for 48 h and their dry weights were recorded.
At the end of the experiments, samples were collected, flashed directly into liquid nitrogen (LN2), and used for physiological and biochemical analyses.
The photosynthetic pigments in the leaves were extracted with 80% acetone and the concentrations of chlorophyll, carotenoids and anthocyanins were measured by determining the absorbance via spectrophotometry at 646, 480 and 567 nm respectively. Pigment concentrations in mg/g FW were then calculated following the method of Porra et al. [46].
2.2. Measurement of Na+ and K+ Concentrations
Leaf and root samples were digested with 69% HNO3 acid and H2O2 (5:1 v/v). The Na+ and K+ concentrations were quantified using flame photometry (Microprocessor Flame Photometer, Electronics India, Model 1382, Parwanoo, Himachal Pradesh, India) against Na+ and K+ standards of known concentration, following a previously described method [47].
2.3. Determination of Hydrogen Peroxidase and Lipid Peroxidation
Quantification of H2O2 in plant tissues was carried out following a previously established protocol [48]. Briefly, 0.5-g samples of leaf and root tissues were ground in LN2 and a potassium phosphate buffer (KPB) (pH 6.8). The extracts of the various samples were centrifuged at 6000× g for 25 min at 4 °C. A 100-µL aliquot of the supernatant was added to 1 mL of xylenol solution, thoroughly mixed and allowed to stand for 30 min. The intensity of the color, which directly represents the amount of H2O2 in the sample, was measured by spectrophotometer at 560 nm.
Lipid peroxidation was determined by measuring the amount of malondialdehyde (MDA), a product of lipid peroxidation, using the calorimetric method as described by Stewart and Bewley [49]. One gram of fresh leaf or root tissue was ground in LN2. Then, the ground tissues were suspended in 0.5% thiobarbituric acid in 20% trichloroacetic acid solution. Subsequently, the mixture was heated to 95 °C for 30 min and cooled quickly in an ice bath. The mixture was then centrifuged at 10,000× g for 10 min, and the absorbance of the supernatant was determined spectrophotometrically at 532 nm and at 600 nm for the non-specific absorbance value. The MDA concentration was calculated from the extinction coefficient at 155 mM−1 cm−1.
2.4. Determination of Antioxidant Enzymatic Activity
SOD enzyme activity was assayed according to the previously published protocol of Beauchamp and Fridovich [50], which is based on inhibition of the photochemical reduction of nitro blue tetrazolium (NBT). Briefly, 0.5-g samples of leaf or root extracts were prepared by grinding in 5 mL of potassium phosphate buffer (pH 7), mixed with ethylenediaminetetraacetic acid (EDTA) (pH 7.8) and 1% PVPP. The extract was centrifuged at 6000× g for 10 min. The resultant supernatant obtained in this experiment was the enzyme extract. The reaction mixture contained 100 mM KPB, 100 mM methionine, 2 mM NBT, 2 mM riboflavin, 445.7 µL of water, and 50 µL of enzyme extract. The reactants were then placed under a 20-W fluorescent lamp for 15 min and the samples in the tubes were covered with a black cloth. At the end of the reaction, the absorbance at 560 nm was determined.
CAT was measured in the samples based on a previously established protocol [51]. Briefly, leaf and root samples were ground in LN2 and added to 5 mL of the extraction buffer (25 KPB, 0.5 mM EDTA, 2% (w/v) PVPP (pH 7.8)). Enzyme extract was centrifuged at 15,000× g at 4 °C for 15 min. The reaction was carried out using an aliquot of 50 µL of the enzyme extract, 500 mM KPB (pH 7.0), 100 mM H2O2, and 750 µL of water. Absorbance was measured using a UV spectrophotometer at 240 nm with an extinction coefficient of 0.04 mM−1 cm−1, for a period of 1 min.
APX activity was measured using the method of Nakano and Asada [52]. Plant tissues (0.5 g) were ground in 0.1 M of phosphate buffer (pH 7.0). Enzyme extract was centrifuged at 11,000× g for 25 min at 4 °C. The reaction included 600 µL of the enzyme extract, 0.1 M phosphate buffer, 5 mM ascorbate, and 0.5 mM H2O2. Absorbance was measured using a UV spectrophotometer at 290 nm with an extinction coefficient of 2.8 mM−1 cm−1, for a period of 1 min.
2.5. Determination of Non-Enzymatic Antioxidants
Total phenolic content (TPC) concentration was measured in leaf and root samples following a previously established protocol [53]. Briefly, 0.5-g samples of plant tissue were powdered in LN2 and then extracted by shaking for 3 h at room temperature, with a 100% methanol solution. Subsequently, the extract was centrifuged at 10,000× g for 10 min at 4 °C. The TPC reaction included 100 µL of the plant extract, 900 µL dH2O, 1 mL of diluted phenol reagent (1:2), and 0.4 M sodium carbonate (Na2CO3). The reaction mixture was incubated for 5 min at 50 °C, then cooled and placed in the dark for 2 h at room temperature. Absorbance was measured spectrophotometrically at 765 nm. The standard curve was prepared using gallic acid and the TPC was calculated in mg gallic acid equivalent/g of fresh weight of the plant tissues.
Total flavonoid content (TFC) was estimated using the AlCl3 method, as previously established [54]. Samples of 0.5 g of leaf or root tissue were ground in LN2 and then in 80% methanol. Solutions of 2.9 mL of methanol, 10% AlCl3.6H2O, 5% sodium potassium tartrate, and 0.5 mL dH2O were sequentially added to a 100-µL aliquot of the extract. The mixture was vigorously shaken, and the absorbance measured by spectrometer using light of wavelength 415 nm. The standard curve was prepared using catechin and the results were expressed as mg catechin equivalent/g of fresh weight of the plant tissues.
Total glutathione (GS) concentration was determined as previously described [55]. The reaction component included 500 µL of the plant extract, 0.2 M Tris buffer (pH 8.2), 0.1 M 5,5-dithiobis-(2-nitrobenzoic acid) (DTNB), and 3.9 mL of methanol. The components were shaken for 15 min at room temperature and the absorbance measured by spectrophotometer at 412 nm. The GS concentration was estimated, and the standard curve was prepared, using a stock solution of reduced glutathione (GSH), and the total GS concentration was expressed as mg/g of fresh weight of the plant tissues. Proline concentration was determined by the colorimetric method, according to Bates et al. [56].
2.6. Data Analysis
Data were analyzed using one-way analysis of variance (ANOVA) and the SPSS statistical package version 21 (IBM Corp., Armonk, NY, USA). Tests of significance were carried out using Duncan’s multiple range test (DMRT) at p = 0.05.
3. Results
3.1. The Effect of Salinity on Growth and Na+ and K+ Uptake
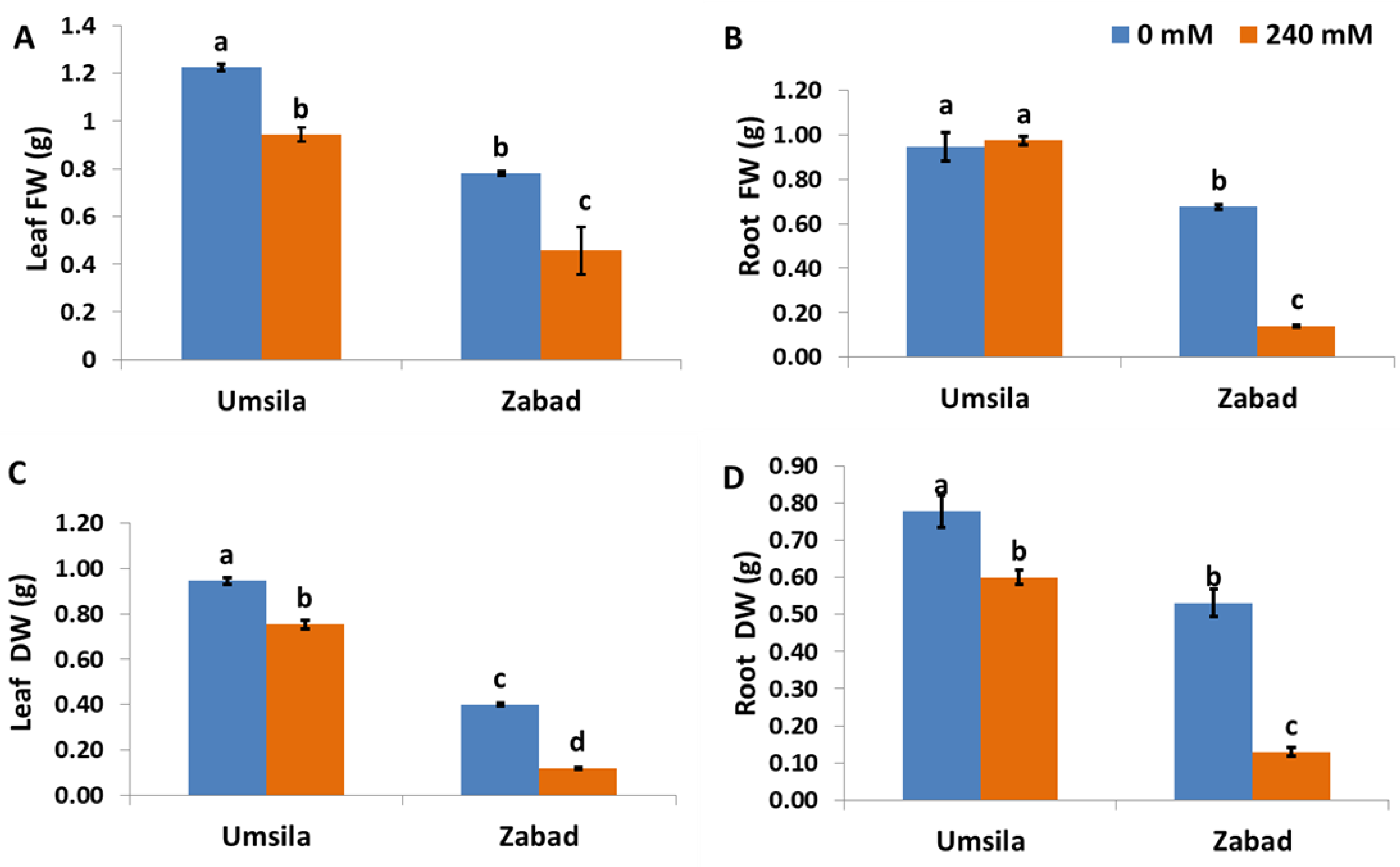
The changes in biomass under the different treatments were represented as fresh and dry weight changes. These changes were used as an indicator of growth inhibition due to salinity. The results showed that under control conditions, ‘Umsila’ exhibited a significantly (p ≤ 0.05) higher fresh and dry weight (Figure 1). The results also showed a significant reduction (p ≤ 0.05) in the fresh and dry weights of leaf tissues and the dry weight of root tissues in both cultivars (Figure 1). Although salinity showed a significant (p ≤ 0.05) negative effect on the fresh weight of ‘Zabad’, there was no significant (p ≤ 0.05) effect on the root fresh weight of ‘Umsila’ (Figure 1B). It was observed that salinity has a higher negative impact on fresh and dry weights of the leaves and root tissues in ‘Zabad’ than on these tissues in ‘Umsila’.
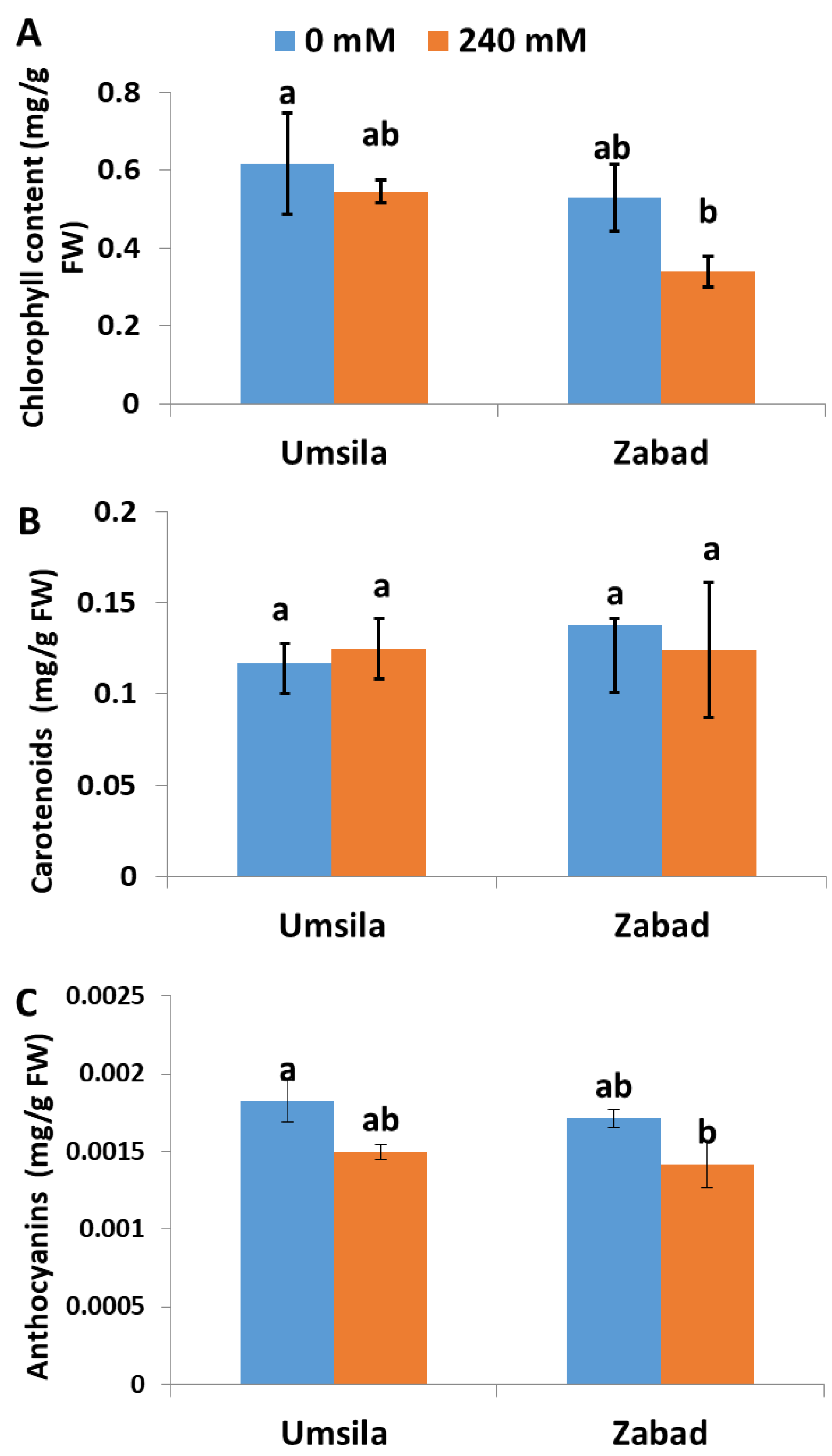
Plants often alter the amounts of chlorophyll, carotenoid and anthocyanin pigments produced in their leaf tissues, in order to protect the photosynthetic system against abiotic stress. In this study, we aimed at understanding the response of the pigments when date palm seedlings grew under saline conditions. The analysis of the photosynthetic pigments in date palm seedlings revealed that salinity had no significant (p ≤ 0.05) impact on the accumulation of chlorophyll in ‘Umsila’. However, there was a significant (p ≤ 0.05) negative effect on the chlorophyll concentrations in ‘Zabad’ (Figure 2A). Similarly, salinity did not show a significant (p ≤ 0.05) effect on carotenoid accumulation in either cultivar (Figure 2B), but salinity showed a significant (p ≤ 0.05) negative effect on the accumulation of anthocyanin in the leaves of ‘Zabad’ (Figure 2C).
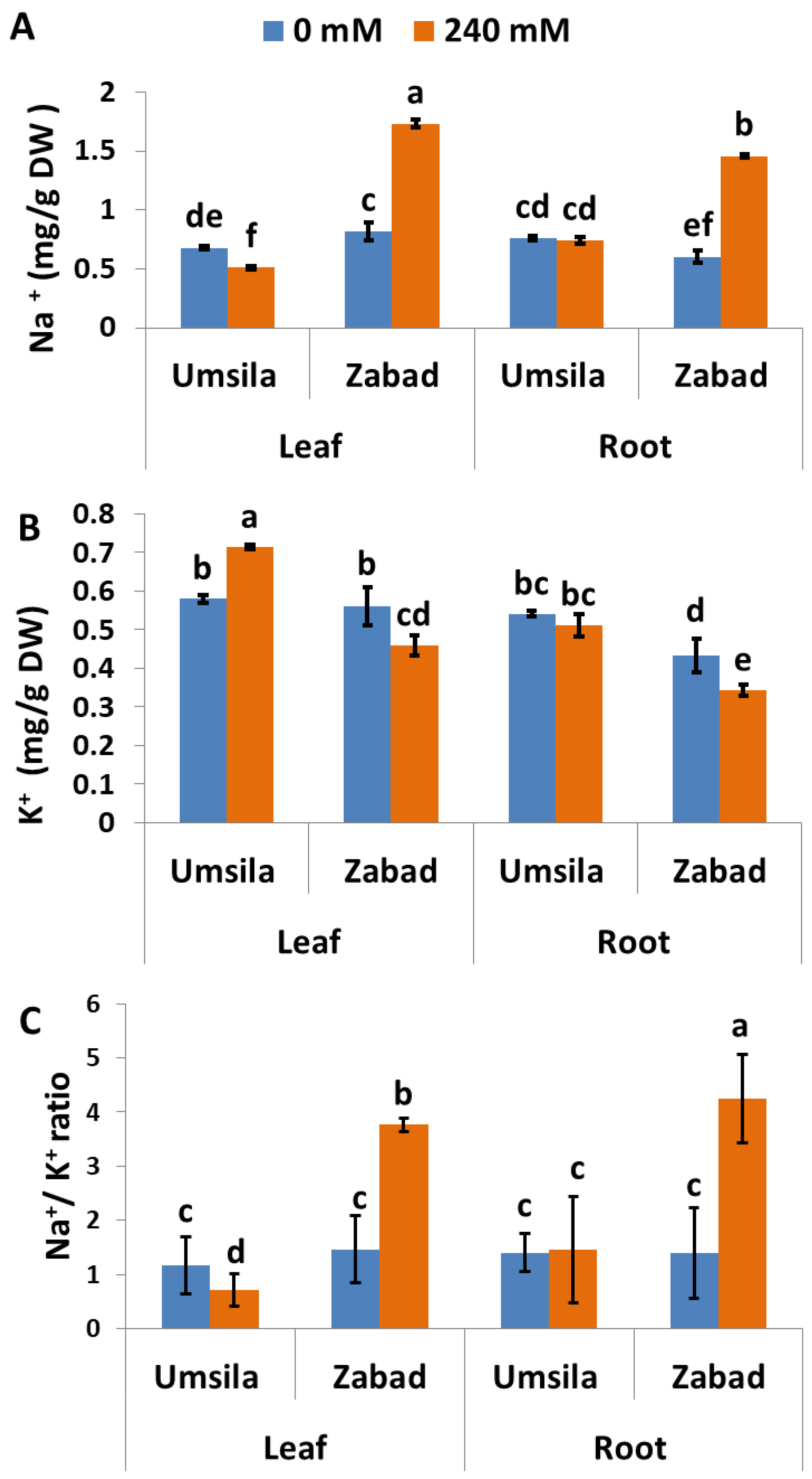
The salinity tolerance capacity is partially dependent on the ability of the plants to avoid Na+ and to enhance K+ accumulation. In order to understand the mechanism behind salinity tolerance in ‘Umsila’, Na+ and K+ concentrations in leaf and root tissues were quantified. The results showed that ‘Umsila’ leaf tissues accumulated less Na+ than ‘Zabad’ leaf tissues, when grown under control conditions. When exposed to salinity, the leaf tissues of ‘Umsila’ had the ability to significantly (p ≤ 0.05) reduce the amount of Na+ they contained, though the amount was not significantly changed in the root tissues. Although the amount of Na+ in ‘Zabad’ tissues was significantly (p ≤ 0.05) increased, Na+ concentration in ‘Umsila’ root tissues was not affected (Figure 3A) after exposure to salinity.
Although K+ concentration was significantly (p ≤ 0.05) increased in the leaf tissues of ‘Umsila’ under the high-salinity treatment, this concentration was significantly decreased in ‘Zabad’ leaf and root tissues. However, this treatment had no significant effect on the concentration of K+ in root tissues of ‘Umsila’ (Figure 3B). The increase in Na+ and the low K+ uptake in ‘Zabad’ resulted in an increase in the Na+/K+ ratio in ‘Zabad’ (Figure 3C).
3.2. The Effects of Salinity on the Accumulation of H2O2 and MDA
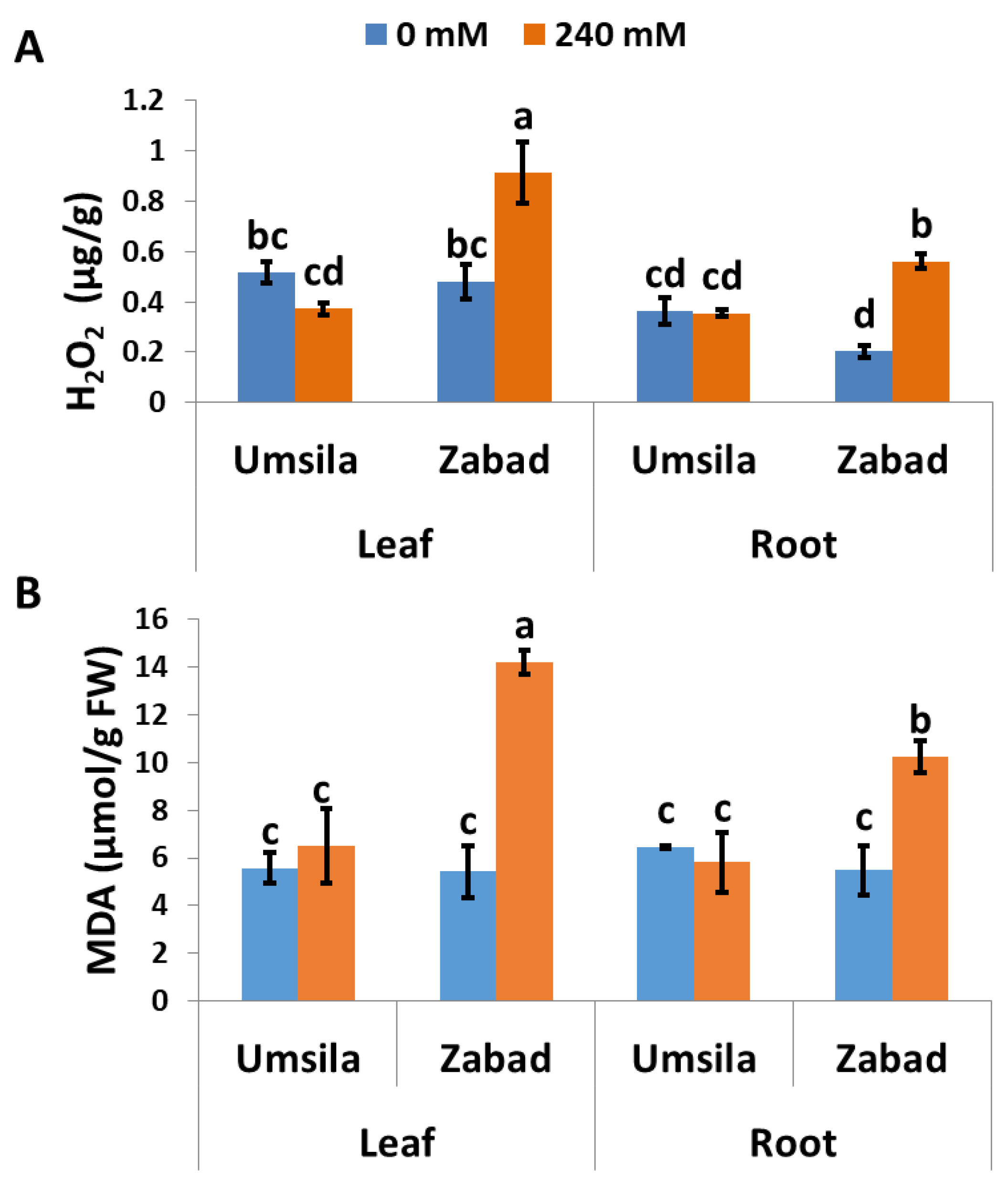
To investigate the role of H2O2 in salinity tolerance in date palm seedlings, the H2O2 concentrations in leaf and root tissues of ‘Umsila’ and ‘Zabad’ were quantified when the plants grew under normal and saline conditions. The results revealed that under normal conditions, the H2O2 concentrations of the leaf tissues of ‘Umsila’ and ‘Zabad’ were similar. However, the H2O2 concentration in the root tissues of ‘Umsila’ was significantly (p ≤ 0.05) higher than in ‘Zabad’. The results also showed that while the H2O2 concentration was not affected in the root tissues of ‘Umsila’, this concentration was significantly (p ≤ 0.05) reduced in the leaf tissues of the same cultivar under salinity treatment (Figure 4A). However, the results showed that, due to salinity, the amounts of H2O2 were significantly (p ≤ 0.05) increased in the leaf and root tissues of ‘Zabad’ (Figure 4A).
MDA plays an important role in disturbing membrane integrity and in controlling programmed cell death in plants. Therefore, maintaining the minimum amounts of this molecule under salinity stress is a key factor in keeping plants healthy and in their subsequent acquisition of tolerance to salinity. Quantification of MDA in leaf and root tissues of date palm seedlings grown under normal and saline conditions revealed that the amount of MDA in the leaf and root tissues of ‘Umsila’ and ‘Zabad’ is similar when the seedlings grew under normal conditions. While the MDA concentration of ‘Umsila’ cultivar was not affected, the MDA concentration was significantly (p ≤ 0.05) increased in the leaf and root tissues of ‘Zabad’ under salinity conditions (Figure 4B).
3.3. The Effects of Salinity on Antioxidant Enzymatic Activity
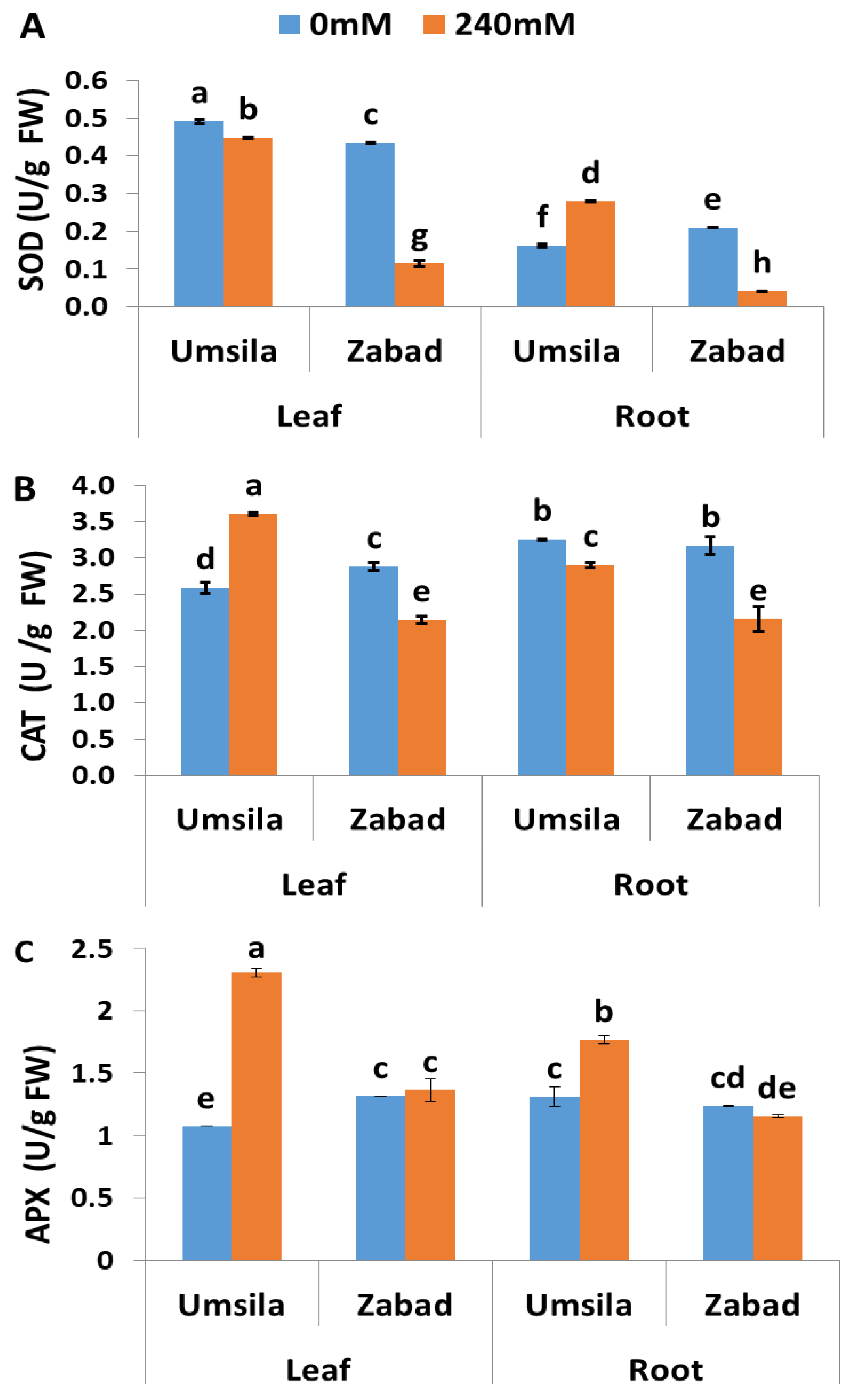
The enzymatic antioxidant activities of SOD, CAT, and APX were measured in ‘Umsila’ and ‘Zabad’ seedlings in order to investigate the contribution of these enzymes to the salinity tolerance mechanism of this plant. In general, it was observed that leaf tissues showed a significantly (p ≤ 0.05) higher concentration of SOD activity than root tissues (Figure 5A). Under normal conditions, SOD activity was significantly (p ≤ 0.05) higher in the leaves of ‘Umsila’ than in the leaves of ‘Zabad’. However, it was significantly (p ≤ 0.05) lower in the roots of ‘Umsila’ than in the roots of ‘Zabad’. When exposed to salinity, SOD activity was significantly (p ≤ 0.05) decreased in the leaves of ‘Umsila’ and ‘Zabad’ and in the roots of ‘Zabad’. However, it was significantly (p ≤ 0.05) increased in the roots of ‘Umsila’ (Figure 5A).
Quantification of the enzymatic activity of CAT revealed that ‘Zabad’ had significantly (p ≤ 0.05) higher enzymatic activity than ‘Umsila’ in the leaf tissues. However, the activity was very similar in the root tissues of both cultivars when the seedlings were grown under normal conditions (Figure 5B). The results showed that there was a significant (p ≤ 0.05) increase in CAT activity in the leaf tissues of ‘Umsila’. However, there was a significant (p ≤ 0.05) decrease in CAT activity in the leaf and root tissues of ‘Zabad’, as well as in the root tissues of ‘Umsila’, in response to salinity (Figure 5B).
The results of this study showed that salinity stress significantly (p ≤ 0.05) increased the activity of APX in the leaf and root tissues of ‘Umsila’. However, salinity did not enhance the APX activity in ‘Zabad’ (Figure 5C). The increase in APX activity in the leaf tissues of ‘Umsila’ due to salinity was remarkable, indicating a significant involvement of this enzyme in salinity stress tolerance in date palms.
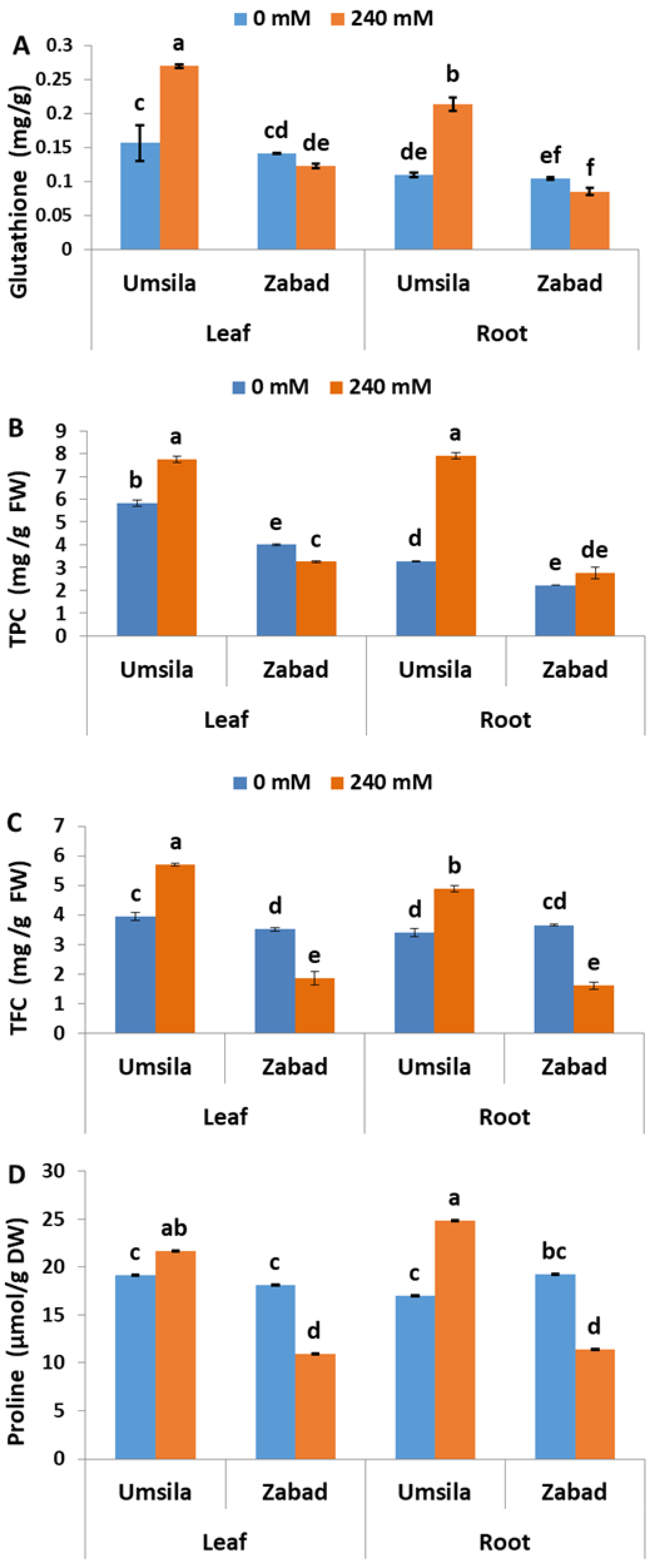
The role of the non-enzymatic antioxidants GS, TPC, TFC, and free proline in salinity tolerance was investigated in date palm seedlings subjected to control and saline conditions. Glutathione was accumulated similarly in ‘Umsila’ and ‘Zabad’ tissues, when the plants grew under normal conditions (Figure 6A). Under saline conditions, glutathione was significantly (p ≤ 0.05) increased in the leaf and root tissues of ‘Umsila’. However, salinity had no significant (p ≤ 0.05) effect on the accumulation of glutathione in the leaf tissues of ‘Zabad’ seedlings (Figure 6A).
The analysis of TPC in date palm seedlings grown under normal conditions revealed that ‘Umsila’ leaf tissues contained the highest amount of TPC, compared to other tissues. The analysis also showed that salinity significantly (p ≤ 0.05) increased the accumulation of TPC in the leaf and root tissues of ‘Umsila’. However, it significantly (p ≤ 0.05) decreased the accumulation of TPC in the leaf tissues of ‘Zabad’ and had an insignificant (p ≤ 0.05) effect on accumulation in the root tissues (Figure 6B).
Quantification of TFC in date palm seedlings showed that leaf and root tissues of both cultivars contained similar amounts when the plants were grown under control conditions. The analysis also revealed that the accumulation of these chemicals was significantly (p ≤ 0.05) enhanced in the ‘Umsila’ leaf and root tissues when grown under salt stress. However, the accumulation of TFC was significantly (p ≤ 0.05) inhibited in ‘Zabad’ under the same environmental conditions (Figure 6C). Similarly, free proline accumulation analysis showed that salinity significantly increased the accumulation of proline in the leaf and root tissues of ‘Umsila’, but the amount of proline was significantly (p ≤ 0.05) decreased in the leaf and the root tissues of ‘Zabad’, when exposed to salinity (Figure 6D). Interestingly, under normal conditions, the amount of proline in ‘Umsila’ and ‘Zabad’ tissues was similar.
4. Discussion
Salinity tolerance is an important trait for plants such as date palms that grow in arid and semi-arid areas where water has high concentrations of salts [57]. Plant species and cultivars vary in their ability to tolerate salinity, due to changes in their genetic and epigenetic makeup which took a long time to evolve [58]. In date palms, the ‘Umsila’ cultivar had previously been identified as a salinity-tolerant cultivar and ‘Zabad’ as a salinity-susceptible cultivar [6]. However, the mechanisms behind this salt tolerance trait are yet to be identified. Tolerance may involve a single mechanism or several mechanisms, such as the ability to avoid salts in the soil, the ability to compartmentalize Na+ ions among different tissues and cells, or the ability to deal with the consequences of excessive amounts of salt in cells by producing additional quantities of antioxidants [26].
In this study, we focused on the role of antioxidants in salt tolerance in date palms. ‘Umsila’ showed an ability to grow better and to maintain fresh and dry weights, compared with the salt-susceptible ‘Zabad’ cultivar. The results showed that salinity did not have a negative effect on the fresh weight of ‘Umsila’. This could be attributed to the ability of this tolerant cultivar to maintain enough water in the root tissues in response to salinity. Despite the fact that salinity showed a negative effect on both cultivars in terms of the dry weight of tissues, the reduction in ‘Zabad’ was much higher than that observed in ‘Umsila’ (Figure 1). In fact, salt-tolerant date palms [3,6], as well as other plant species, are able to maintain growth vigor, in comparison with susceptible plant species [26], especially when they are able to maintain the photosynthetic machinery by protecting the photosynthetic pigments chlorophyll, carotenoid, and anthocyanin. This protection is further enhanced when these plants are able to exhibit a functional oxidative protective mechanism. This was clearly observed in ‘Umsila’ seedlings when grown under saline conditions (Figure 2).
Our previous screening study results showed no effect of salinity stress on the quantum yield efficiency of PSII (Qy) in the ‘Umsila’ plants, indicating optimal functioning of PSII. Hence, the photosynthetic machinery is well protected, and leaf and root growth maintained.
The ability of ‘Umsila’ to exhibit better growth than ‘Zabad’ probably stems from the fact that it can better control the accumulation of Na+ and K+ ions in its tissues (Figure 3). While the amounts of Na+ ions in the leaf and root tissues of ‘Umsila’ did not increase, the amounts of Na+ ions in the leaf and root tissues of ‘Zabad’ increased enormously when the plants were grown under saline conditions (Figure 3). Unlike in ‘Umsila’, amounts of K+ ions were decreased in ‘Zabad’ under the same conditions. Excessive amounts of cellular Na+ ions have several negative impacts on plants and may cause cellular toxicity [59], a situation which ‘Umsila’ was able to avoid by reducing the absorbance of Na+ ions and increasing the accumulation of K+ ions in its leaf tissues, thereby maintaining a healthy Na+/K+ ratio balance in the cytosol [60]. Although ‘Umsila’ was able to achieve or maintain a low concentration of H2O2 and MDA, the amount of these ROS increased in ‘Zabad’ under saline conditions (Figure 4). This could be the reason behind the susceptibility of ‘Zabad’ to salt stress. Several reports have previously shown that high salinity induces oxidative stress in various plant species [37,61]. Various studies have reported that salt stress alters the structure and the composition of the lipids in the plasma membrane. This alteration affects the degree of saturation of free fatty acids and free sterols, which may eventually lead to a decrease in the fluidity of the cell membrane [62,63]. A salt-tolerant genotype, such as ‘Umsila’, produces a lower concentration of H2O2 and MDA than ‘Zabad’, which may contribute to the greater salt tolerance.
The salt tolerance in ‘Umsila’ may also involve an overproduction of SOD, CAT, and APX when the seedlings are exposed to salinity. This pattern of antioxidant enzymatic activity was not observed in ‘Zabad’, where salinity led to a reduction in the activity of these enzymes (Figure 5). This reduction could be due to damage in the antioxidant system. This system is essential, when active, in facilitating the conversion of superoxide anions and singlet oxygen from the Mehler reaction in the chloroplast, to hydrogen peroxide in the thylakoid [64].
Consistent with our findings, various studies have previously reported that SODs increase in response to abiotic stresses in plants [65,66], including salinity [67].
‘Umsila’ produced higher levels of active CAT scavenger enzyme than ‘Zabad’ leaf tissues when exposed to salinity (Figure 5). Recently it was reported that H2O2 formation is inhibited by the photosynthetic electron transport inhibitor 3-(3,4-dichlorophenyl)-1,1-diethylurea (DCMU), where plastid terminal oxidase (PTOX) receives electrons from plastoquinol and reduces oxygen to water instead of H2O2 in PSII [64]. As in ‘Umsila’, it was previously reported that CAT activity increased in the leaf of Calendula officinalis L. under saline conditions [68]. The low level of CAT activity in the root of ‘Umsila’ could be because the H2O2 concentration did not significantly increase in the root system, compared to the leaf system, in response to salinity (Figure 4A), or possibly because photosynthesis mainly occurs in the leaf tissues and, therefore, there is a low probability of an electron imbalance in the roots. Therefore, the concentration of H2O2 in the root tissues did not reach a critical level that could activate CAT, since this enzyme has a low affinity to H2O2 [69].
The results obtained from this study revealed that there was a significant increase in the levels of APX activity in both leaf and root tissues of ‘Umsila’ (Figure 5C). This is another indication that the salinity-tolerant ‘Umsila’ cultivar can decompose the elevated concentrations of ROS produced in the cells due to salinity, with higher efficiency than the susceptible ‘Zabad’ cultivar. The high level of APX activity could be due to the presence of substantial amounts of ascorbate [70], which may donate the electron in the ascorbate-glutathione (ASC-GSH) cycle, thus detoxifying the H2O2 in the chloroplast to water molecules. The high level of APX activity in the ‘Umsila’ tissues may not only be used to scavenge the generated H2O2 but it may also function by disposing of the excessive excitation energy of the electrons. It has been reported that the ASC-GSH cycle, which protects against stress, also acts as an alternative energy and electron sink in the chloroplast [71,72]. In a study on Eutrema salsugineum (Pall.) Al-Shehbaz & Warwick, 2005 grown under saline conditions, it was reported that activation of the ASC-GSH cycle is an avoidance mechanism for the single-oxygen-mediated damage in PSII, and also helps to maintain highly reduced states of the PQ pool [73].
In addition to the increase in the enzymatic antioxidants in ‘Umsila’, there was also an increase in non-enzymatic antioxidants such as GS, TPC, TFC, and proline, due to salinity. In the roots of ‘Umsila’, there was a significant increase in the GS (Figure 6A), probably as a result of the activation of a redox homeostasis mechanism such as the ASC-GSH cycle, in response to salt stress [74,75,76]. The root-specific increase of GS in ‘Umsila’ could be due to an increase in the demand for sulfur ions under salinity stress. Therefore, there is a requirement to synthesize sulfur-containing compounds, such as GS [77].
Date palm plants are rich in flavonoids, phenolic acid, sterols, carotenoids, procyanidins, and anthocyanins, and their concentrations depend on the soil conditions [78,79,80]. The increase in TPC in the leaf and root tissues of ‘Umsila’ could be a defensive mechanism developed by this cultivar in order to tolerate salinity through an affective ROS scavenging pathway. On the other hand, there was a slight increase in TPC in the root tissues of ‘Zabad’ as a result of the significant increase in Na+ ions and ROS accumulations. However, this increase was apparently not enough to ameliorate the salinity stress. A study on Salacca zalacca (Gaertn.) Voss has reported that phenolic compounds have redox properties and play an antioxidant role in abiotic stress tolerance [81,82,83]. Salinity has induced the accumulation of TPC in Setaria italica (L.) P. Beauv. [84], as well as in a number of halophytes [85,86], and in the tolerant genotypes of Morus alba L. [87].
The results obtained from this study revealed that the high-salinity treatment of date palm seedlings induced the accumulation of TFC only in the ‘Umsila’ cultivar (Figure 6C). A previous study has shown that biosynthesis of antioxidant flavonoids is triggered under stress conditions, when the activity of antioxidant enzymes against ROS starts to decline [88,89]. However, their role in salinity tolerance mechanisms is unclear [90,91].
In addition to being a signal molecule, proline amino acid is considered an antioxidant and an osmolytic agent that plays an important role in salinity tolerance in plants [92]. The accumulation of proline was also enhanced in the ‘Umsila’ tissues in response salinity (Figure 6D). Despite the fact that proline is a common response element for most abiotic stresses in date palm cv. Khalas [45], surprisingly, it was not accumulated in response to salinity in ‘Zabad’. This could be due to malfunctioning in the glutamate or other signaling pathways leading to synthesis of proline in this cultivar under high-salinity conditions. Low amounts of proline production in ‘Zabad’ may provide this plant with a lower antioxidant capability and subject the cells of the plant to a higher probability of electrolyte leakage.
5. Conclusions
Our results suggest that the salinity tolerance of ‘Umsila’ is due to the ability of this cultivar to take up low amounts of Na+ and high amounts of K+ ions. In addition, the tolerance can be attributed to the ability of ‘Umsila’ to protect the photosynthetic pigments and, thus, maintain photosynthates, thereby minimizing any loss in biomass due to salinity. ‘Umsila’ also has the capacity to minimize the amount of H2O2 and MDA produced due to salt stress, to activate the antioxidant enzymes SOD, CAT, and APX, and to increase the accumulation of non-enzymatic antioxidant reagents such as GS, TPC, TFC, and proline, hence maintaining the oxidative stress balance between ROS and scavengers. These activities are poorly managed in the salt-susceptible ‘Zabad’ cultivar. Therefore, antioxidation could represent a key mechanism for salt tolerance in date palms. Further studies are needed to describe other components of this mechanism.
Author Contributions
L.A.K. conceived, designed, performed the experiments, analyzed data, and wrote the manuscript; R.A.Y. supervised the work; and M.W.Y. designed the experiment, supervised the work, wrote the manuscript, and contributed reagents/materials/analysis tools.
Funding
This study is supported by the generous grant number 151 from the research council (TRC), Oman to M.W.Y.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flowers, T.; Flowers, S. Why does salinity pose such a difficult problem for plant breeders? Agric. Water Manag. 2005, 78, 15–24. [Google Scholar] [CrossRef]
- Furr, J. Water and salinity problems of Abadan Island date gardens. Date Growers’ Inst. 1975, 52, 14–17. [Google Scholar]
- Sperling, O.; Lazarovitch, N.; Schwartz, A.; Shapira, O. Effects of high salinity irrigation on growth, gas-exchange, and photoprotection in date palms (Phoenix dactylifera L., cv. Medjool). Environ. Exp. Bot. 2014, 99, 100–109. [Google Scholar] [CrossRef]
- Ramawat, K.G. Desert Plants: Biology and Biotechnology; Springer Science & Business Media: Berlin, Germany, 2009. [Google Scholar]
- Youssef, T.; Awad, M.A. Mechanisms of enhancing photosynthetic gas exchange in date palm seedlings (Phoenix dactylifera L.) under salinity stress by a 5-aminolevulinic acid-based fertilizer. J. Plant Growth Regul. 2008, 27, 1. [Google Scholar] [CrossRef]
- Al Kharusi, L.; Assaha, D.; Al-Yahyai, R.; Yaish, M. Screening of Date Palm (Phoenix dactylifera L.) Cultivars for Salinity Tolerance. Forests 2017, 8, 136. [Google Scholar] [CrossRef]
- Ramoliya, P.; Pandey, A. Soil salinity and water status affect growth of Phoenix dactylifera seedlings. N. Z. J. Crop Horticult. Sci. 2003, 31, 345–353. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signaling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. AoB Plants 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plants cope with water stress in the field? Photosynthesis and growth. AoB Plants 2002, 89, 907–916. [Google Scholar] [CrossRef]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlowski, T. Responses of woody plants to flooding and salinity. Tree Physiol. 1997, 17, 490. [Google Scholar] [CrossRef]
- Yaish, M.W.; Patankar, H.V.; Assaha, D.V.; Zheng, Y.; Al-Yahyai, R.; Sunkar, R. Genome-wide expression profiling in leaves and roots of date palm (Phoenix dactylifera L.) exposed to salinity. BMC Genom. 2017, 18, 246. [Google Scholar] [CrossRef] [PubMed]
- Patankar, H.V.; Al-Harrasi, I.; Al-Yahyai, R.; Yaish, M.W. Identification of Candidate Genes Involved in the Salt Tolerance of Date Palm (Phoenix dactylifera L.) Based on a Yeast Functional Bioassay. DNA Cell Biol. 2018, 3, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M.W.; Sunkar, R.; Zheng, Y.; Ji, B.; Al-Yahyai, R.; Sardar, F.A. A genome-wide identification of the miRNAome in response to salinity stress in date palm (Phoenix dactylifera L.). Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M.W.; Al-Lawati, A.; Al-Harrasi, I.; Patankar, H.V. Genome-wide DNA Methylation analysis in response to salinity in the model plant caliph medic (Medicago truncatula). BMC Genom. 2018, 19, 78. [Google Scholar] [CrossRef]
- Al-Harrasi, I.; Al-Yahyai, R.; Yaish, M.W. Differential DNA methylation and transcription profiles in date palm roots exposed to salinity. PLoS ONE 2018, 13, e0191492. [Google Scholar] [CrossRef]
- Yaish, M.W.; Antony, I.; Glick, B.R. Isolation and characterization of endophytic plant growth-promoting bacteria from date palm tree (Phoenix dactylifera L.) and their potential role in salinity tolerance. Anton. Leeuwenhoek 2015, 107, 1519–1532. [Google Scholar] [CrossRef]
- Yaish, M.W.; Al-Lawati, A.; Jana, G.A.; Patankar, H.V.; Glick, B.R. Impact of soil salinity on the structure of the bacterial endophytic community identified from the roots of caliph medic (Medicago truncatula). PLoS ONE 2016, 11, e0159007. [Google Scholar] [CrossRef]
- Yaish, M.W. Draft genome sequence of endophytic bacterium Enterobacter asburiae PDA134, isolated from date palm (Phoenix dactylifera L.) roots. Genome Announc. 2016, 4. [Google Scholar] [CrossRef]
- Jana, G.A.; Al-Yahyai, R.; Yaish, M.W. Genome Sequencing of Microbacterium sp. Yaish 1, a Bacterial Strain Isolated from the Rhizosphere of Date Palm Trees Affected by Salinity. Genome Announc. 2017, 5. [Google Scholar] [CrossRef]
- Yaish, M.W. Draft genome sequence of the endophytic Bacillus aryabhattai strain SQU-R12, identified from Phoenix dactylifera L. roots. Genome Announc. 2017, 5. [Google Scholar] [CrossRef]
- Yaish, M.W.; Al-Harrasi, I.; Alansari, A.S.; Al-Yahyai, R.; Glick, B.R. The use of high throughput DNA sequence analysis to assess the endophytic microbiome of date palm roots grown under different levels of salt stress. Int. Microbiol. 2016, 19, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Lokhande, V.H.; Nikam, T.D.; Patade, V.Y.; Ahire, M.L.; Suprasanna, P. Effects of optimal and supra-optimal salinity stress on antioxidative defence, osmolytes and in vitro growth responses in Sesuvium portulacastrum L. Plant Cell Tissue Organ. Cult. 2011, 104, 41–49. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Srivastava, A.K.; Srivastava, S.; Nikam, T.D.; Suprasanna, P. Regulated alterations in redox and energetic status are the key mediators of salinity tolerance in the halophyte Sesuvium portulacastrum (L.) L. Plant Growth Regul. 2011, 65, 287–298. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of reactive oxygen species by photosystem II. Biochim. Biophys. Acta Bioenergy 2009, 1787, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. AoB Plants 2009, 103, 551–560. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, A.; Dietz, K.-J. Reactive Oxygen Species and the Redox-Regulatory Network in Cold Stress Acclimation. Antioxidants 2018, 7, 169. [Google Scholar] [CrossRef] [PubMed]
- Symes, A.; Shavandi, A.; Zhang, H.; Mohamed Ahmed, I.A.; Al-Juhaimi, F.Y.; Bekhit, A.E.-D.A. Antioxidant Activities and Caffeic Acid Content in New Zealand Asparagus (Asparagus officinalis) Roots Extracts. Antioxidants 2018, 7, 52. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Mohanty, P. Defense potentials to NaCl in a mangrove, Bruguiera parviflora: Differential changes of isoforms of some antioxidative enzymes. J. Plant Physiol. 2004, 161, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Chawla, S.; Jain, S.; Jain, V. Salinity induced oxidative stress and antioxidant system in salt-tolerant and salt-sensitive cultivars of rice (Oryza sativa L.). J. Plant Biochem. Biotechnol. 2013, 22, 27–34. [Google Scholar] [CrossRef]
- Wang, N.; Qiao, W.; Liu, X.; Shi, J.; Xu, Q.; Zhou, H.; Yan, G.; Huang, Q. Relative contribution of Na+/K+ homeostasis, photochemical efficiency and antioxidant defense system to differential salt tolerance in cotton (Gossypium hirsutum L.) cultivars. Plant Physiol. Biochem. 2017, 119, 121–131. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Mohammadi, Z.; Rastegar, S.; Abdollahi, F.; Hosseini, Y. Morphological and antioxidant enzymatic activity responses of sapodilla rootstock to salinity stress. J. Plant Process Funct. 2018, 6, 23–28. [Google Scholar]
- Valderrama, R.; Corpas, F.J.; Carreras, A.; GÓMEZ-RODRÍGUEZ, M.V.; Chaki, M.; Pedrajas, J.R.; Fernandez-Ocana, A.; DEL RÍO, L.A.; Barroso, J.B. The dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants. Plant Cell Environ. 2006, 29, 1449–1459. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Yang, H.; Chang, Y.; Cheng, J.; Bai, S.; Yin, J. Effects of nitric oxide on reactive oxygen species and antioxidant capacity in Chinese Bayberry during storage. Sci. Horticult. 2012, 135, 106–111. [Google Scholar] [CrossRef]
- Ozden, M.; Demirel, U.; Kahraman, A. Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci. Horticult. 2009, 119, 163–168. [Google Scholar] [CrossRef]
- Arbona, V.; Flors, V.; Jacas, J.; García-Agustín, P.; Gómez-Cadenas, A. Enzymatic and Non-enzymatic Antioxidant Responses of Carrizo citrange, a Salt-Sensitive Citrus Rootstock, to Different Levels of Salinity. Plant Cell Physiol. 2003, 44, 388–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaish, M.W. Proline accumulation is a general response to abiotic stress in the date palm tree (Phoenix dactylifera L.). Genet. Mol. Res. 2015, 14, 9943–9950. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenergy 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Ren, Z.-H.; Gao, J.-P.; Li, L.-G.; Cai, X.-L.; Huang, W.; Chao, D.-Y.; Zhu, M.-Z.; Wang, Z.-Y.; Luan, S.; Lin, H.-X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-H.; Offler, C.E.; Ruan, Y.-L. A simple, rapid, and reliable protocol to localize hydrogen peroxide in large plant organs by DAB-mediated tissue printing. Front. Plant Sci. 2014, 5, 745. [Google Scholar] [CrossRef]
- Stewart, R.R.; Bewley, J.D. Lipid peroxidation associated with accelerated aging of soybean axes. Plant Physiol. 1980, 65, 245–248. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinhem, Germany, 1983; pp. 273–286. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Lamaison, J.; Carant, A. The amount of main flavonoids in flowers and leaves of Crataegus monogyna Jacq. and Crataegus laevigata (Poiret) DC. (Rosacea). Pharm. Acta Helv. 1996, 65, 315–320. [Google Scholar]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yaish, M.W.; Kumar, P.P. Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M. Epigenetic Modifications Associated with Abiotic and Biotic Stresses in Plants: An Implication for Understanding Plant Evolution. Front. Plant Sci. 2017, 8, 1983. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Assaha, D.V.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Shabala, S.; Munns, R. Salinity stress: Physiological constraints and adaptive mechanisms. Plant Stress Physiology, 2nd ed.; CABI: Wallingford, UK, 2017; pp. 24–63. [Google Scholar]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.-E.; Rajashekar, C.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses role of phospholipase Dα in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef]
- Plumb, W.; Townsend, A.J.; Rasool, B.; Alomrani, S.; Razak, N.; Karpinska, B.; Ruban, A.V.; Foyer, C.H. Ascorbate-mediated regulation of growth, photoprotection, and photoinhibition in Arabidopsis thaliana. J. Exp. Bot. 2018, 69, 2823–2835. [Google Scholar] [CrossRef]
- Luis, A.; Corpas, F.J.; López-Huertas, E.; Palma, J.M. Plant Superoxide Dismutases: Function Under Abiotic Stress Conditions. In Antioxidants and Antioxidant Enzymes in Higher Plants; Springer: Berlin, Germany, 2018; pp. 1–26. [Google Scholar]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—Mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef] [PubMed]
- Qiu-Fang, Z.; Yuan-Yuan, L.; Cai-Hong, P.; Cong-Ming, L.; Bao-Shan, W. NaCl enhances thylakoid-bound SOD activity in the leaves of C3 halophyte Suaeda salsa L. Plant Sci. 2005, 168, 423–430. [Google Scholar] [CrossRef]
- Chaparzadeh, N.; D’Amico, M.L.; Khavari-Nejad, R.-A.; Izzo, R.; Navari-Izzo, F. Antioxidative responses of Calendula officinalis under salinity conditions. Plant Physiol. Biochem. 2004, 42, 695–701. [Google Scholar] [CrossRef]
- Mizuno, M.; Kamei, M.; Tsuchida, H. Ascorbate peroxidase and catalase cooperate for protection against hydrogen peroxide generated in potato tubers during low-temperature storage. IUBMB Life 1998, 44, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N.; Pallanca, J. Ascorbate metabolism in relation to oxidative stress. Biochem. Soc. Trans. 1996, 24, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The water–water cycle as alternative photon and electron sinks. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1419–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiciarz, M.; Niewiadomska, E.; Kruk, J. Effects of salt stress on low molecular antioxidants and redox state of plastoquinone and P700 in Arabidopsis thaliana (glycophyte) and Eutrema salsugineum (halophyte). Photosynthetica 2018, 56, 811–819. [Google Scholar] [CrossRef]
- Bela, K.; Riyazuddin, R.; Horváth, E.; Hurton, Á.; Gallé, Á.; Takács, Z.; Zsigmond, L.; Szabados, L.; Tari, I.; Csiszár, J. Comprehensive analysis of antioxidant mechanisms in Arabidopsis glutathione peroxidase-like mutants under salt-and osmotic stress reveals organ-specific significance of the AtGPXL’s activities. Environ. Exp. Bot. 2018, 150, 127–140. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef]
- Noshi, M.; Yamada, H.; Hatanaka, R.; Tanabe, N.; Tamoi, M.; Shigeoka, S. Arabidopsis dehydroascorbate reductase 1 and 2 modulate redox states of ascorbate-glutathione cycle in the cytosol in response to photooxidative stress. Biosci. Biotechnol. Biochem. 2017, 81, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Baliga, M.S.; Baliga, B.R.V.; Kandathil, S.M.; Bhat, H.P.; Vayalil, P.K. A review of the chemistry and pharmacology of the date fruits (Phoenix dactylifera L.). Food Res. Int. 2011, 44, 1812–1822. [Google Scholar] [CrossRef]
- Mansouri, A.; Embarek, G.; Kokkalou, E.; Kefalas, P. Phenolic profile and antioxidant activity of the Algerian ripe date palm fruit (Phoenix dactylifera). Food Chem. 2005, 89, 411–420. [Google Scholar] [CrossRef]
- Al-Farsi, M.; Alasalvar, C.; Al-Abid, M.; Al-Shoaily, K.; Al-Amry, M.; Al-Rawahy, F. Compositional and functional characteristics of dates, syrups, and their by-products. Food Chem. 2007, 104, 943–947. [Google Scholar] [CrossRef]
- Suica-Bunghez, I.; Teodorescu, S.; Dulama, I.; Voinea, O.; Ion, R. Antioxidant activity and phytochemical compounds of snake fruit (Salacca Zalacca). IOP Conf. Ser. Mater. Sci. Eng. 2016, 133. [Google Scholar] [CrossRef]
- Bunghez, I.; Barbinta Patrascu, M.; Badea, N.; Doncea, S.; Popescu, A.; Ion, R. Antioxidant silver nanoparticles green synthesized using ornamental plants. J. Optoelectron. Adv. Mater. 2012, 14, 1016–1022. [Google Scholar]
- Aires, A.; Carvalho, R. Profiling of Polyphenol Composition and Antiradical Capacity of Erica cinerea. Antioxidants 2017, 6, 72. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Grimm, B.; Wobus, U.; Weschke, W. Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiol. Plant 2000, 109, 435–442. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouysegu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Kefu, Z.; Hai, F.; San, Z.; Jie, S. Study on the salt and drought tolerance of Suaeda salsa and Kalanchoe claigremontiana under iso-osmotic salt and water stress. Plant Sci. 2003, 165, 837–844. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Agastian, P.; Kingsley, S.; Vivekanandan, M. Effect of salinity on photosynthesis and biochemical characteristics in mulberry genotypes. Photosynthetica 2000, 38, 287–290. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, I.; Alegre, L.; Van Breusegem, F.; Munné-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Di Ferdinando, M.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as antioxidants in plants under abiotic stresses. In Abiotic Stress Responses in Plants; Springer: Berlin, Germany, 2012; pp. 159–179. [Google Scholar]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The effect of salinity on the fresh weight (A,B) and dry weight (C,D) of date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).
Figure 1.
The effect of salinity on the fresh weight (A,B) and dry weight (C,D) of date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).

Figure 2.
The effect of salinity on the total chlorophyll (A), total carotenoid (B), and anthocyanin concentrations (C) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).
Figure 2.
The effect of salinity on the total chlorophyll (A), total carotenoid (B), and anthocyanin concentrations (C) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).

Figure 3.
The effect of salinity on sodium (Na+) (A) and potassium (K+) (B) accumulation and on the Na+/K+ ratio (C) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).
Figure 3.
The effect of salinity on sodium (Na+) (A) and potassium (K+) (B) accumulation and on the Na+/K+ ratio (C) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).

Figure 4.
The effect of salinity on the activity of hydrogen peroxidase (A) and malondialdehyde (B), when date palm seedlings were exposed to control (0 mM NaCl) and high-salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).
Figure 4.
The effect of salinity on the activity of hydrogen peroxidase (A) and malondialdehyde (B), when date palm seedlings were exposed to control (0 mM NaCl) and high-salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).

Figure 5.
Differential enzymatic antioxidant activity of superoxide dismutase (SOD) (A), catalase (CAT) (B) and ascorbate peroxidase (APX) (C) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).
Figure 5.
Differential enzymatic antioxidant activity of superoxide dismutase (SOD) (A), catalase (CAT) (B) and ascorbate peroxidase (APX) (C) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).

Figure 6.
The effect of salinity on the concentrations of the non-enzymatic antioxidants total glutathione (GS) (A), total phenolic content (TPC) measured as mg gallic acid equivalent/g of fresh weight of the plant tissues (B), total flavonoid content (TFC) measured as mg catechin equivalent/g of fresh weight of the plant tissues (C) and proline (D) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).
Figure 6.
The effect of salinity on the concentrations of the non-enzymatic antioxidants total glutathione (GS) (A), total phenolic content (TPC) measured as mg gallic acid equivalent/g of fresh weight of the plant tissues (B), total flavonoid content (TFC) measured as mg catechin equivalent/g of fresh weight of the plant tissues (C) and proline (D) in date palm seedlings when subjected to control (0 mM NaCl) and salinity (240 mM NaCl) conditions. Bars represent the mean ± SE (n = 3).

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Al Kharusi, L.; Al Yahyai, R.; Yaish, M.W. Antioxidant Response to Salinity in Salt-Tolerant and Salt-Susceptible Cultivars of Date Palm. Agriculture 2019, 9, 8. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010008
AMA Style
Al Kharusi L, Al Yahyai R, Yaish MW. Antioxidant Response to Salinity in Salt-Tolerant and Salt-Susceptible Cultivars of Date Palm. Agriculture. 2019; 9(1):8. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010008
Chicago/Turabian StyleAl Kharusi, Latifa, Rashid Al Yahyai, and Mahmoud W. Yaish. 2019. "Antioxidant Response to Salinity in Salt-Tolerant and Salt-Susceptible Cultivars of Date Palm" Agriculture 9, no. 1: 8. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010008
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.