
Natural Feed Supplements Improve Growth, Non-Specific Immune Responses and Resistance against Vibrio alginolyticus in Lates calcarifer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish Rearing
2.2. Experimental Diets Preparation
2.3. Feeding Trial
2.4. Immunity Tests
2.5. Challenge Test
2.6. Statistical Analyses
3. Results
3.1. Improvement of Growth by Probiotics and Plant Products
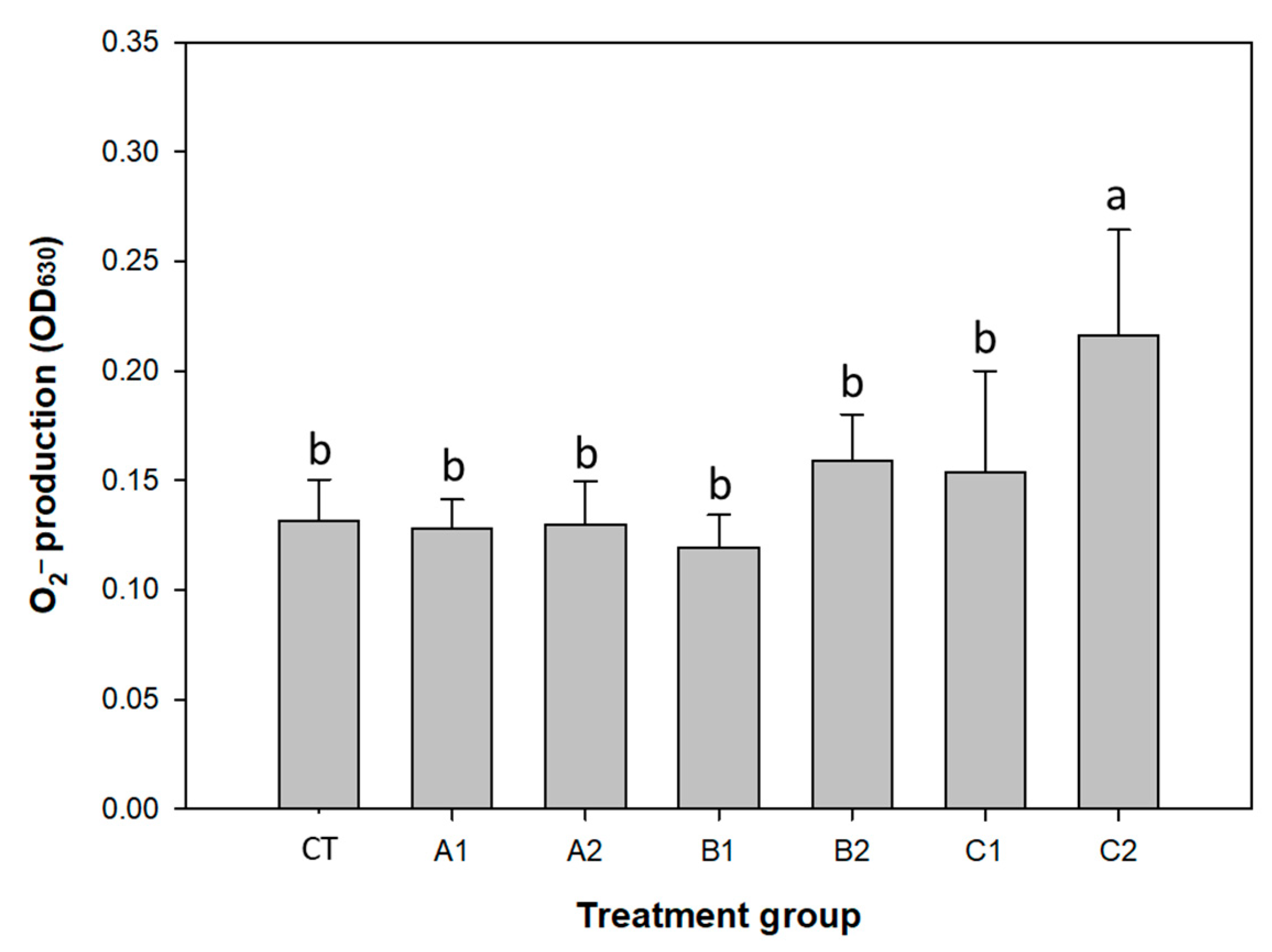
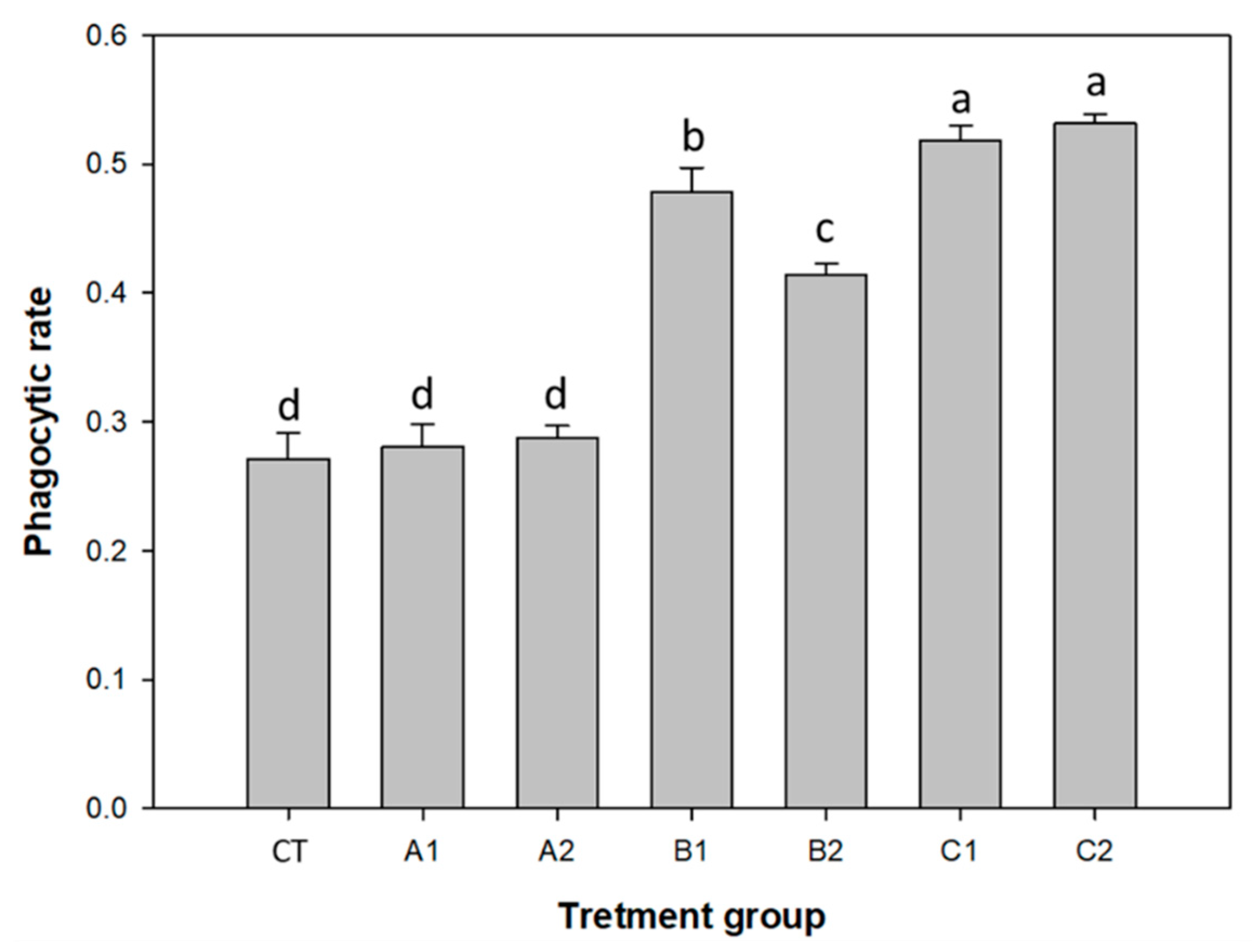
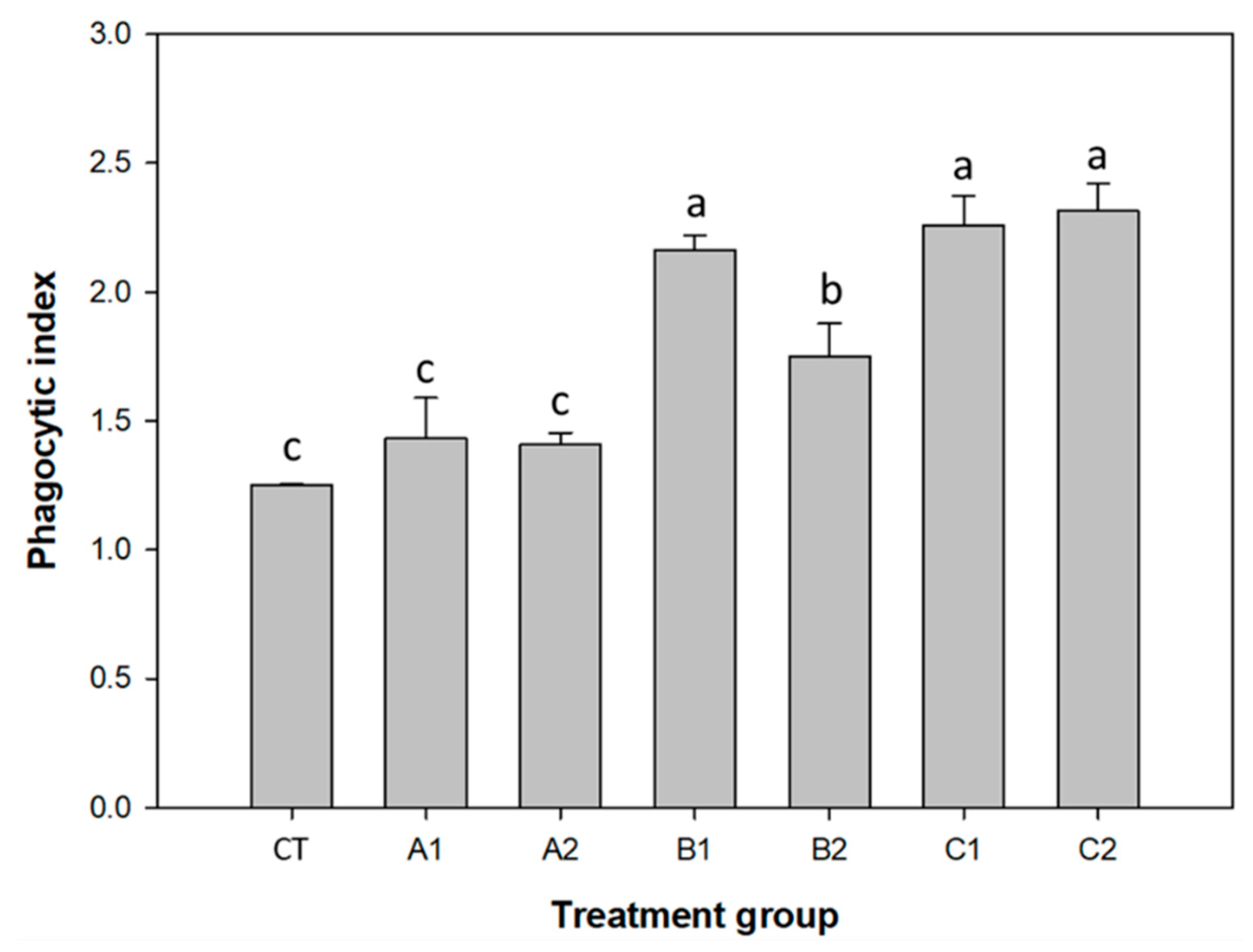
3.2. Improvement of Immunity by Probiotics and Plant Products
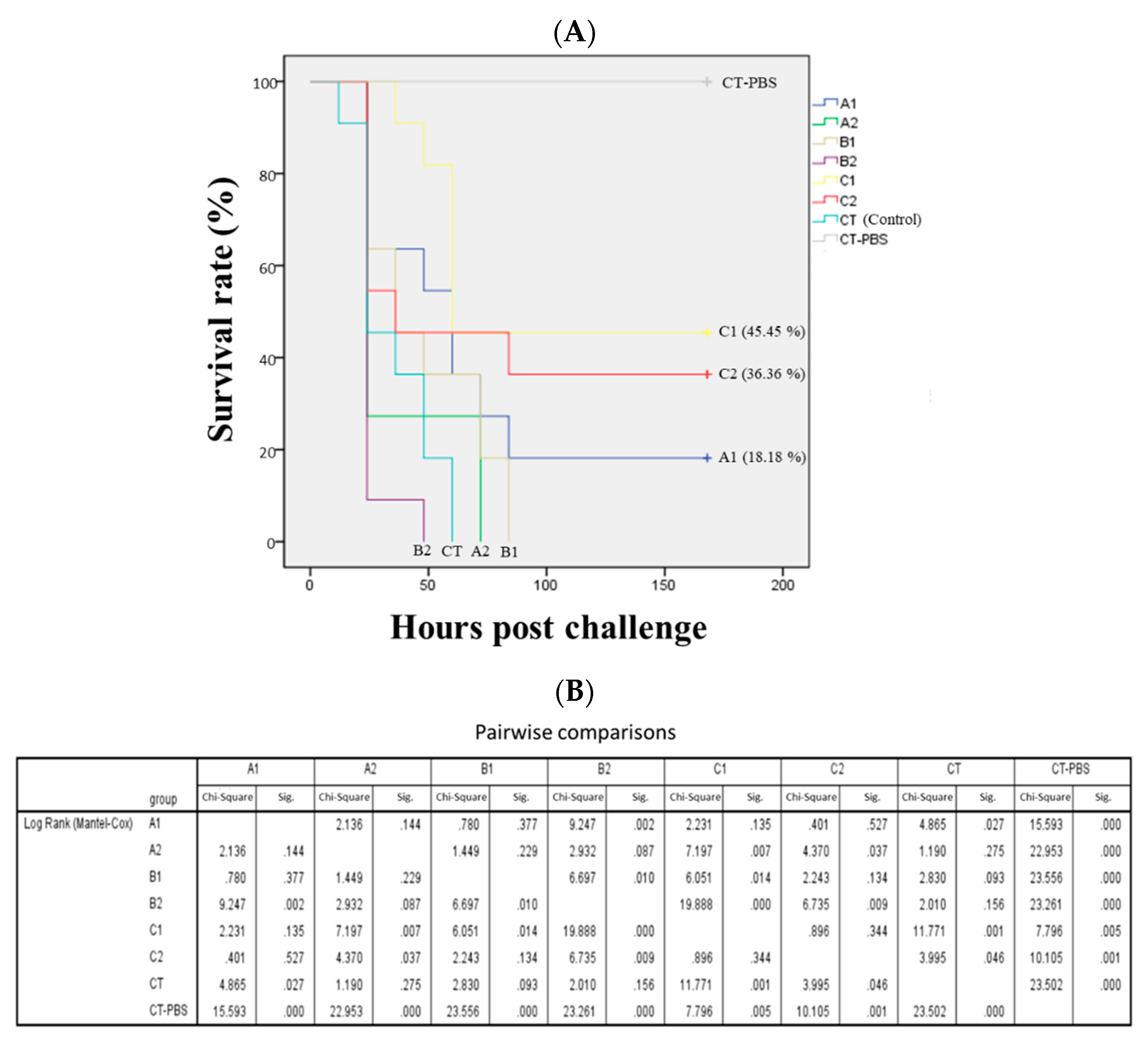
3.3. Improvement of Disease Resistance by Probiotics and Plant Products
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ina-Salwany, M.Y.; Al-saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in fish: A review on disease development and prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.G.; Shiu, Y.L.; Nguyen, T.P.; Truong, Q.P.; Chen, J.C.; Liu, C.H. Current applications, selection, and possible mechanisms of actions of synbiotics in improving the growth and health status in aquaculture: A review. Fish Shellfish Immunol. 2017, 64, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef] [PubMed]
- De, B.C.; Meena, D.K.; Behera, B.K.; Das, P.; Mohapatra, P.K.D.; Sharma, A.P. Probiotics in fish and shellfish culture: Immunomodulatory and ecophysiological responses. Fish Physiol. Biochem. 2014, 40, 921–971. [Google Scholar]
- Olmos, J.; Acosta, M.; Mendoza, G.; Pitones, V. Bacillus subtilis, an ideal probiotic bacterium to shrimp and fish aquaculture that increase feed digestibility, prevent microbial diseases, and avoid water pollution. Arch. Microbiol. 2020, 202, 427–435. [Google Scholar] [CrossRef]
- Ringø, E.; Doan, H.V.; Lee, S.H.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, lactic acid bacteria and bacilli: Interesting supplementation for aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef] [Green Version]
- Satora, M.; Magdziarz, M.; Rząsa, A.; Rypuła, K.; Płoneczka-Janeczko, K. Insight into the intestinal microbiome of farrowing sows following the administration of garlic (Allium sativum) extract and probiotic bacteria cultures under farming conditions. BMC Vet. Res. 2020, 16, 442. [Google Scholar] [CrossRef]
- Elbaz, A.M.; Ibrahim, N.S.; Shehata, A.M.; Mohamed, N.G.; Abdel-Moneim, A.E. Impact of multi-strain probiotic, citric acid, garlic powder or their combinations on performance, ileal histomorphometry, microbial enumeration and humoral immunity of broiler chickens. Trop. Anim. Health Prod. 2021, 53, 115. [Google Scholar] [CrossRef]
- Glencross, B. The nutritional management of barramundi, Lates calcarifer-a review. Aquac. Nutr. 2006, 12, 291–309. [Google Scholar] [CrossRef]
- Liao, Z.H.; Ibarra-Arana, M.J.; Chen, J.C.; Huang, H.T.; Lin, Y.J.; Nan, F.H. The effects of dietary supplement of Ulva fasciata, fermented soybean and probiotics on the growth of Apostichopus japonicas. Aquac. Nutr. 2021, 27, 1363–1369. [Google Scholar] [CrossRef]
- Lee, P.T.; Liao, Z.H.; Huang, H.T.; Chuang, C.Y.; Nan, F.H. β-glucan alleviates the immunosuppressive effects of oxytetracycline on the non-specific immune responses and resistance against Vibrio alginolyticus infection in Epinephelus fuscoguttatus × Epinephelus lanceolatus hybrids. Fish Shellfish Immunol. 2020, 100, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Hasan, K.N.; Banerjee, G. Recent studies on probiotics as beneficial mediator in aquaculture: A review. J. Basic Appl. Zool. 2020, 81, 53. [Google Scholar] [CrossRef]
- Chen, S.W.; Liu, C.H.; Hu, S.Y. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Thy, H.T.T.; Tri, N.N.; Quy, O.M.; Fotedar, R.; Kannika, K.; Unajak, S.; Areechon, N. Effects of the dietary supplementation of mixed probiotic spores of Bacillus amyloliquefaciens 54A, and Bacillus pumilus 47B on growth, innate immunity and stress responses of striped catfish (Pangasianodon hypophthalmus). Fish Shellfish Immunol. 2017, 60, 391–399. [Google Scholar]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; EI-Basuini, M.F.; Hossain, M.S.; Nhu, T.H.; Dossou, S.; Moss, A.S. Effects of dietary supplementation of Lactobacillus rhamnosus or/and Lactococcus lactis on the growth, gut microbiota and immune responses of red sea bream, Pagrus major. Fish Shellfish Immunol. 2016, 49, 275–285. [Google Scholar] [CrossRef]
- Xia, Y.; Lu, M.X.; Chen, G.; Cao, J.M.; Gao, F.Y.; Wang, M.; Liu, Z.G.; Zhang, D.F.; Zhu, H.P.; Yi, M.M. Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2018, 76, 368–379. [Google Scholar] [CrossRef]
- Pourgholam, M.A.; Khara, H.; Safari, R.; Sadati, M.A.Y.; Aramli, M.S. Dietary administration of Lactobacillus plantarum enhanced growth performance and innate immune response of Siberian sturgeon, Acipenser baerii. Probiotics Antimicro. Prot. 2016, 8, 1–7. [Google Scholar] [CrossRef]
- Son, V.M.; Chang, C.C.; Wu, M.C.; Guu, Y.K.; Chiu, C.H.; Cheng, W.T. Dietary administration of the probiotic, Lactobacillus plantarum, enhanced the growth, innate immune responses, and disease resistance of the grouper Epinephelus coioides. Fish Shellfish Immunol. 2009, 26, 691–698. [Google Scholar] [CrossRef]
- Adorian, T.J.; Jamali, H.; Farsani, H.G.; Darvishi, P.; Hasanpour, S.; Bagheri, T.; Roozbehfar, R. Effects of probiotic bacteria Bacillus on growth performance, digestive enzyme activity, and hematological parameters of Asian sea bass, Lates calcarifer (Bloch). Probiotics Antimicro. Prot. 2019, 11, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.L.; Shiu, Y.L.; Chiu, C.S.; Huang, S.L.; Liu, C.H. Screening probiotic candidates for a mixture of probiotics to enhance the growth performance, immunity, and disease resistance of Asian seabass, Lates calcarifer (Bloch), against Aeromonas hydrophila. Fish Shellfish Immunol. 2017, 60, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Yu, W.L.; Wu, J.P. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.Q.; Wu, Z.L.; Dai, Z.L.; Wang, G.H.; Wu, G.Y. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Abdelwahab, A.M.; EI-Bahr, S.M.; AI-Khamees, S. Influence of dietary garlic (Allium sativum) and/or ascorbic acid on performance, feed utilization, body composition and hemato-biochemical parameters of juvenile Asian sea bass (Lates calcarifer). Animals 2020, 10, 2396. [Google Scholar] [CrossRef]
- Valenzuela-Gutierrez, R.; Lago-Leston, A.; Vargas-Albores, F.; Cicala, F.; Martinez-Porchas, M. Exploring the garlic (Allium sativum) properties for fish aquaculture. Fish Physiol. Biochem. 2021, 47, 1179–1198. [Google Scholar] [CrossRef]
- Nya, E.J.; Austin, B. Use of garlic, Allium sativum, to control Aeromonas hydrophila infection in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2009, 32, 963–970. [Google Scholar] [CrossRef]
- Nya, E.J.; Austin, B. Development of immunity in rainbow trout (Oncorhynchus mykiss, Walbaum) to Aeromonas hydrophila after the dietary application of garlic. Fish Shellfish Immunol. 2011, 30, 845–850. [Google Scholar] [CrossRef]
- Huang, H.T.; Lee, P.T.; Liao, Z.H.; Huang, C.T.; Chen, H.Y.; Huang, Z.G.; Wu, Y.S.; Nan, F.H. Raw garlic (Allium sativum) improves nonspecific immune responses and resistance against Vibrio alginolyticus infection in grouper (Epinephelus coioides). J. Mar. Sci. Tech. 2021, 29, 757–766. [Google Scholar] [CrossRef]
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S. Dietary astaxanthin augments disease resistance of Asian seabass, Lates calcarifer (Bloch, 1790), against Vibrio alginolyticus infection. Fish Shellfish Immunol. 2021, 114, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Talpur, A.D.; Ikhwanuddin, M. Dietary effects of garlic (Allium sativum) on haemato-immunological parameters, survival, growth, and disease resistance against Vibrio harveyi infection in Asian sea bass, Lates calcarifer (Bloch). Aquaculture 2012, 364–365, 6–12. [Google Scholar] [CrossRef]
- Sunu, P.; Sunarti, D.; Mahfudz, L.D.; Yunianto, V.D. Prebiotic activity of garlic (Allium sativum) extract on Lactobacillus acidophilus. Vet. World. 2019, 12, 2046–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.X.; Song, C.Y.; Xie, J.; Ge, X.P.; Liu, B.; Xia, S.L.; Yang, S.; Wang, Q.; Zhu, S.H. Effects of fish meal replacement by soybean peptide on growth performance, digestive enzyme activities, and immune responses of yellow catfish Pelteobagrus fulvidraco. Fish. Sci. 2016, 82, 665–673. [Google Scholar] [CrossRef]
- Ragaza, J.A.; Mamauag, R.E.P.; Yokoyama, S.; Ishikawa, M.; Koshio, S. Dietary soy peptide enhances thermotolerance and survival of juvenile Japanese flounder, Paralichthys olivaceus. J. World Aquac. Soc. 2015, 46, 129–138. [Google Scholar] [CrossRef]
- Park, Y.; Moniruzzaman, M.; Lee, S.; Hong, J.; Won, S.; Lee, J.M.; Yun, H.; Kim, K.W.; Ko, D.; Bai, S.C. Comparison of the effects of dietary single and multi-probiotics on growth, non-specific immune responses and disease resistance in starry flounder, Platichthys stellatus. Fish Shellfish Immunol. 2016, 59, 351–357. [Google Scholar] [CrossRef]
- Aly, S.M.; Ahmed, Y.A.; Ghareeb, A.A.; Mohamed, M.F. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008, 25, 128–136. [Google Scholar] [CrossRef]
- Yeh, S.P.; Chiu, C.H.; Shiu, Y.L.; Huang, Z.L.; Liu, C.H. Effects of diets supplemented with either individual or combined probiotics, Bacillus subtilis E20 and Lactobacillus plantarum 7-40, on the immune response and disease resistance of the mud crab, Scylla paramamosain (Estampador). Aquac. Res. 2014, 45, 1164–1175. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Initial Weight (g) | Final Weight (g) | WG (%) | SGR (%) | FCR | FI | SR (%) |
|---|---|---|---|---|---|---|---|
| CT | 49.82 ± 4.88 a | 84.42 ± 8.57 b | 69.60 ± 1.91 b | 1.89 ± 0.04 b | 1.20 ± 0.03 a | 41.55 | 100 |
| A1 | 50.27 ± 4.89 a | 100.04 ± 11.64 a | 99.01 ± 3.93 a | 2.46 ± 0.07 a | 0.91 ± 0.03 b | 45.49 | 100 |
| A2 | 50.79 ± 4.59 a | 96.26 ± 11.44 a | 89.49 ± 3.08 a | 2.28 ± 0.06 a | 1.00 ± 0.04 b | 45.54 | 100 |
| B1 | 50.58 ± 5.00 a | 98.63 ± 12.60 a | 95.00 ± 1.71 a | 2.39 ± 0.03 a | 0.95 ± 0.02 b | 45.56 | 100 |
| B2 | 51.96 ± 4.26 a | 98.31 ± 13.51 a | 89.20 ± 2.04 a | 2.28 ± 0.04 a | 0.99 ± 0.03 b | 45.82 | 100 |
| C1 | 50.92 ± 5.06 a | 98.27 ± 11.16 a | 92.98 ± 1.93 a | 2.35 ± 0.04 a | 0.98 ± 0.04 b | 46.25 | 100 |
| C2 | 50.75 ± 5.34 a | 97.25 ± 14.74 a | 91.63 ± 4.01 a | 2.32 ± 0.07 a | 0.99 ± 0.03 b | 45.95 | 100 |
| F6,7 | 4.08 | 27.43 | 22.42 | 24.62 | 16.23 | ||
| p | 0.0441 | 0.0002 | 0.0003 | 0.0002 | 0.0009 |
| Groups | Initial Weight (g) | Final Weight (g) | WG (%) | SGR (%) | FCR | FI | SR (%) |
|---|---|---|---|---|---|---|---|
| CT | 49.82 ± 4.88 a | 117.53 ± 14.21 b | 136.59 ± 8.10 b | 1.54 ± 0.06 b | 1.32 ± 0.08 a | 89.56 | 100 |
| A1 | 50.27 ± 4.89 a | 134.51 ± 18.28 a | 167.57 ± 7.93 a | 1.76 ± 0.05 a | 1.23 ± 0.06 a | 103.11 | 100 |
| A2 | 50.79 ± 4.59 a | 136.96 ± 19.06 a | 169.66 ± 14.51 a | 1.77 ± 0.10 a | 1.20 ± 0.10 a | 103.16 | 100 |
| B1 | 50.58 ± 5.00 a | 133.61 ± 19.24 a | 164.23 ± 5.51 a | 1.73 ± 0.04 a | 1.24 ± 0.04 a | 103.17 | 100 |
| B2 | 51.96 ± 4.26 a | 130.47 ± 19.32 ab | 150.97 ± 8.53 ab | 1.64 ± 0.06 ab | 1.32 ± 0.08 a | 103.43 | 100 |
| C1 | 50.92 ± 5.06 a | 137.03 ± 17.48 a | 169.10 ± 4.93 a | 1.77 ± 0.03 a | 1.21 ± 0.04 a | 103.87 | 98 |
| C2 | 50.75 ± 5.34 a | 134.30 ± 25.70 a | 164.64 ± 20.99 a | 1.74 ± 0.14 a | 1.27 ± 0.16 a | 105.13 | 100 |
| F6,7 | 4.08 | 3.54 | 18.47 | 19.61 | 0.63 | ||
| p | 0.0441 | 0.0021 | <0.0001 | <0.0001 | 0.7023 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Z.-H.; Huang, H.-T.; Lin, Y.-R.; Chen, B.-Y.; Lee, Y.-F.; Lin, Y.-H.; Chuang, C.-Y.; Nan, F.-H. Natural Feed Supplements Improve Growth, Non-Specific Immune Responses and Resistance against Vibrio alginolyticus in Lates calcarifer. J. Mar. Sci. Eng. 2022, 10, 692. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050692
Liao Z-H, Huang H-T, Lin Y-R, Chen B-Y, Lee Y-F, Lin Y-H, Chuang C-Y, Nan F-H. Natural Feed Supplements Improve Growth, Non-Specific Immune Responses and Resistance against Vibrio alginolyticus in Lates calcarifer. Journal of Marine Science and Engineering. 2022; 10(5):692. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050692
Chicago/Turabian StyleLiao, Zhen-Hao, Huai-Ting Huang, Yu-Ru Lin, Bo-Ying Chen, Yi-Fan Lee, Yuan-Heng Lin, Chieh-Yuan Chuang, and Fan-Hua Nan. 2022. "Natural Feed Supplements Improve Growth, Non-Specific Immune Responses and Resistance against Vibrio alginolyticus in Lates calcarifer" Journal of Marine Science and Engineering 10, no. 5: 692. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050692