
A 3D Innovative Approach Supporting the Description of Boring Sponges of the Precious Red Coral Corallium rubrum


 , ,
, ,  ,
,  and
and 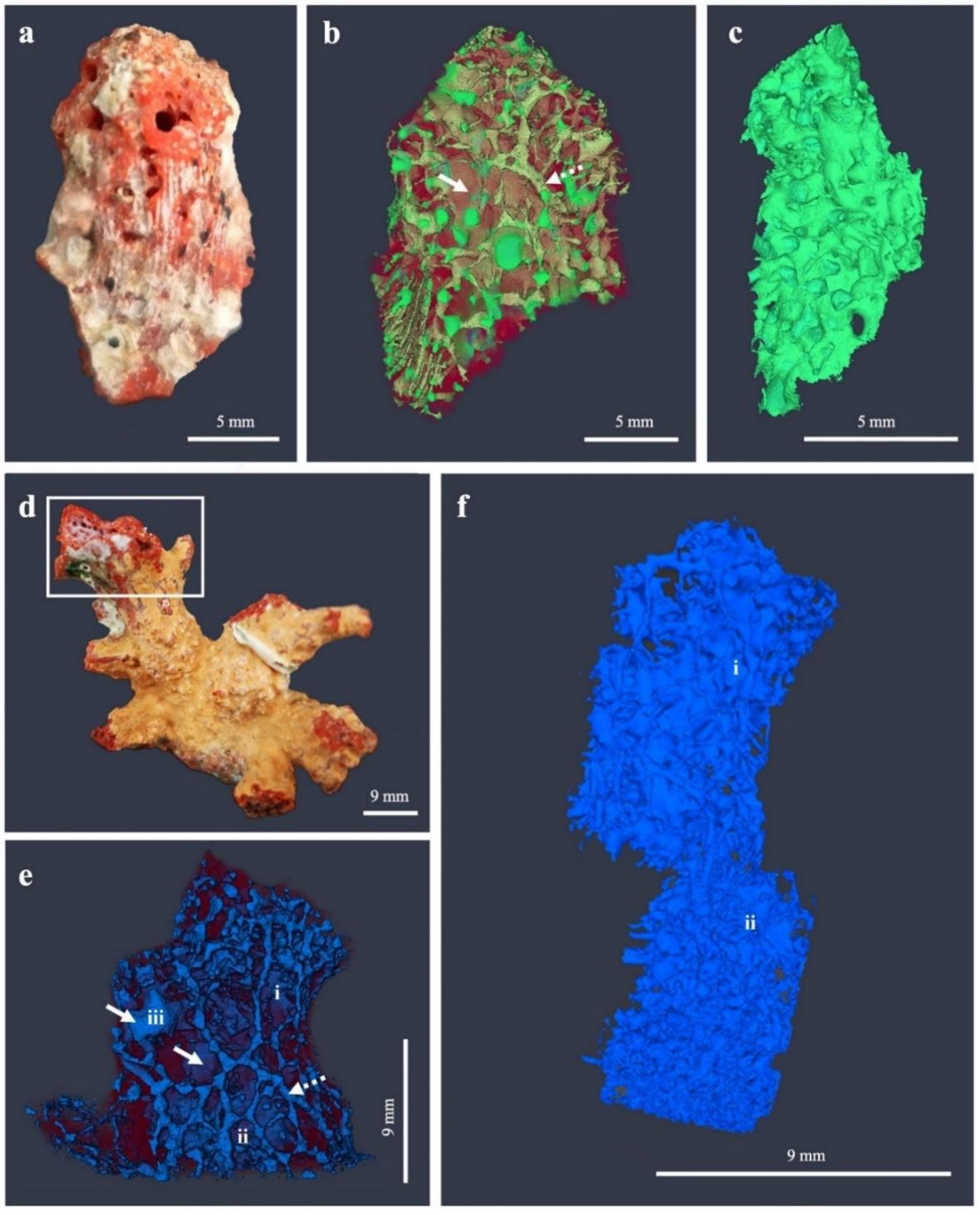{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
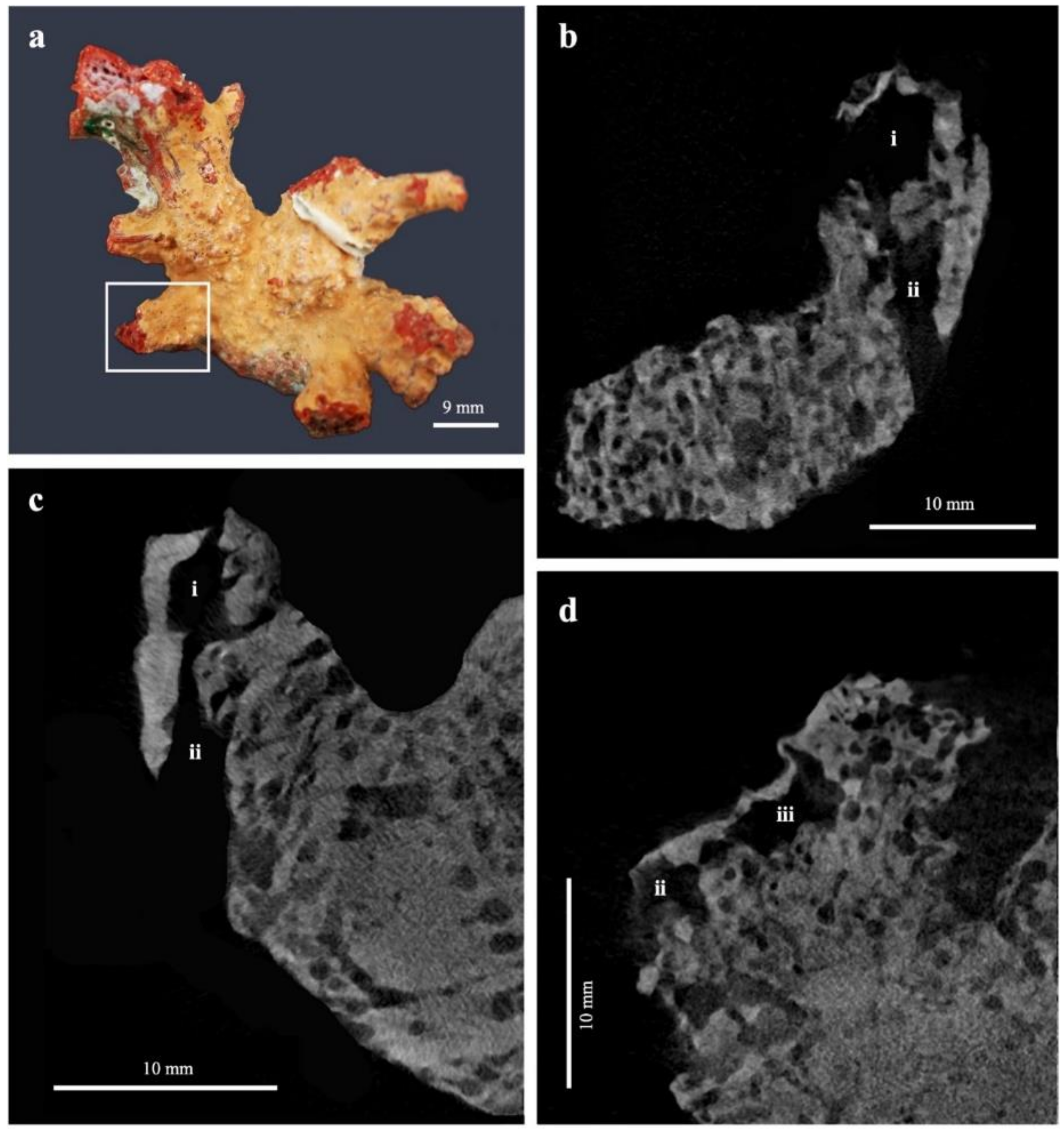
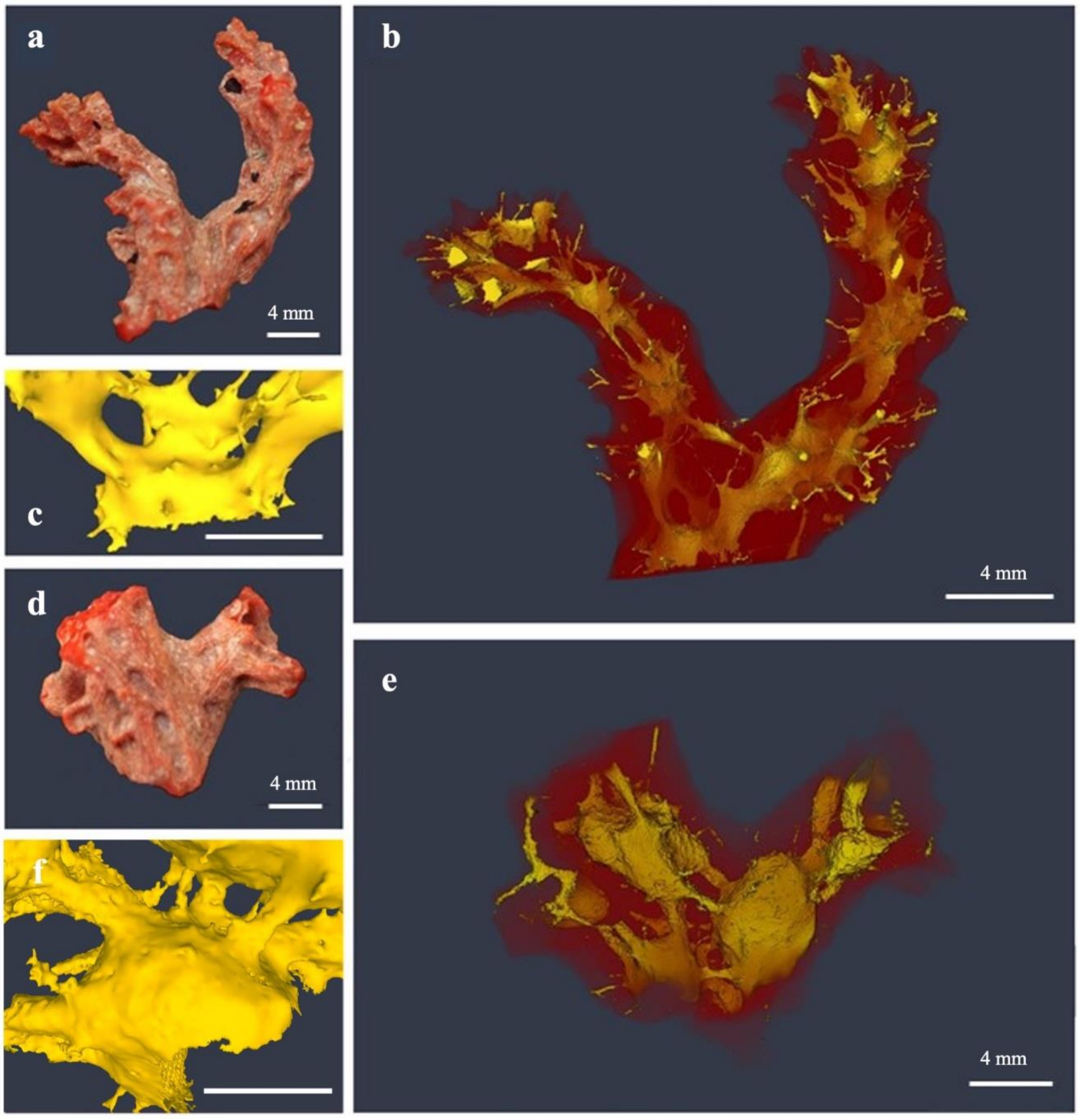
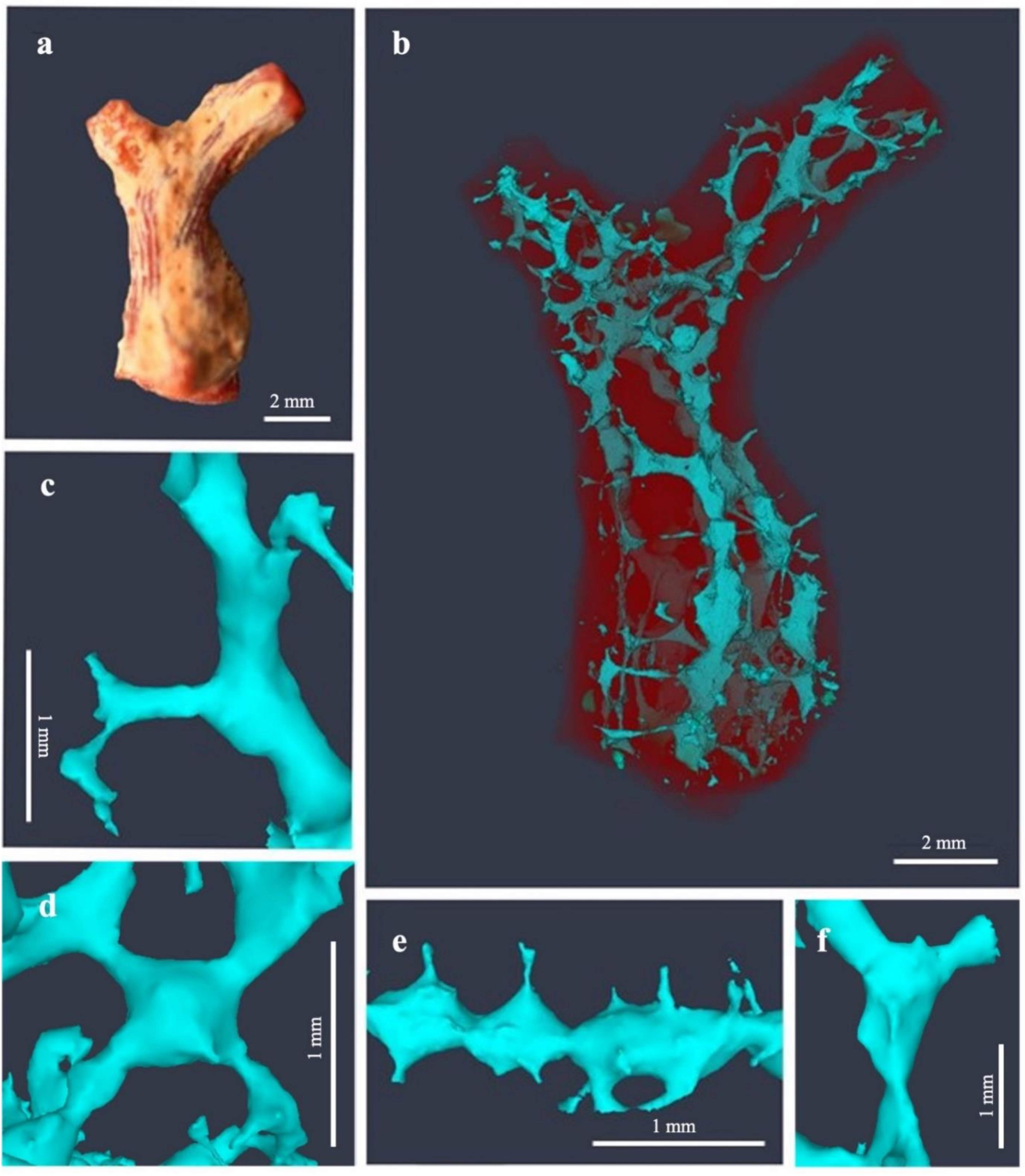
3.1. Erosion Pattern and Excavated Volume
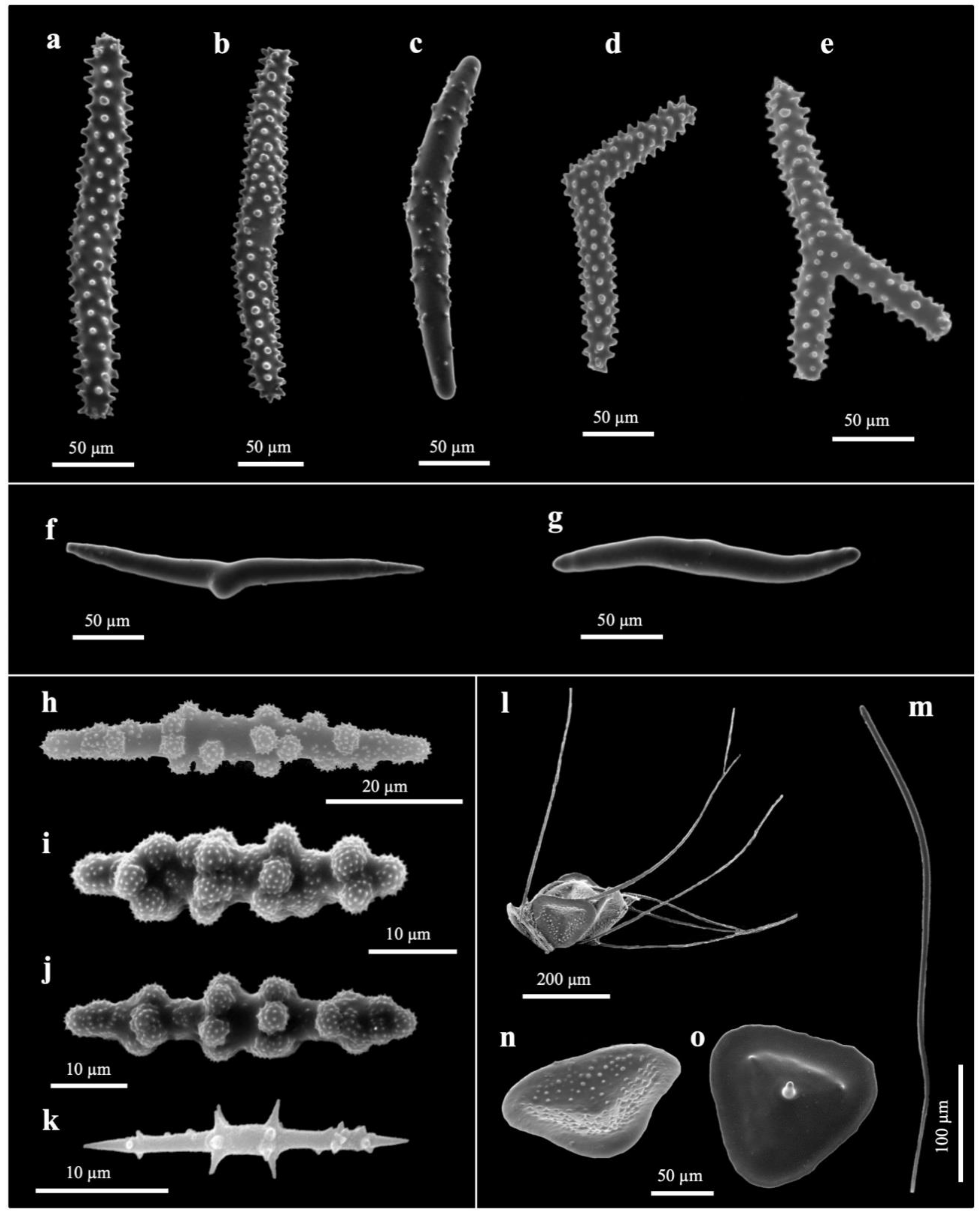
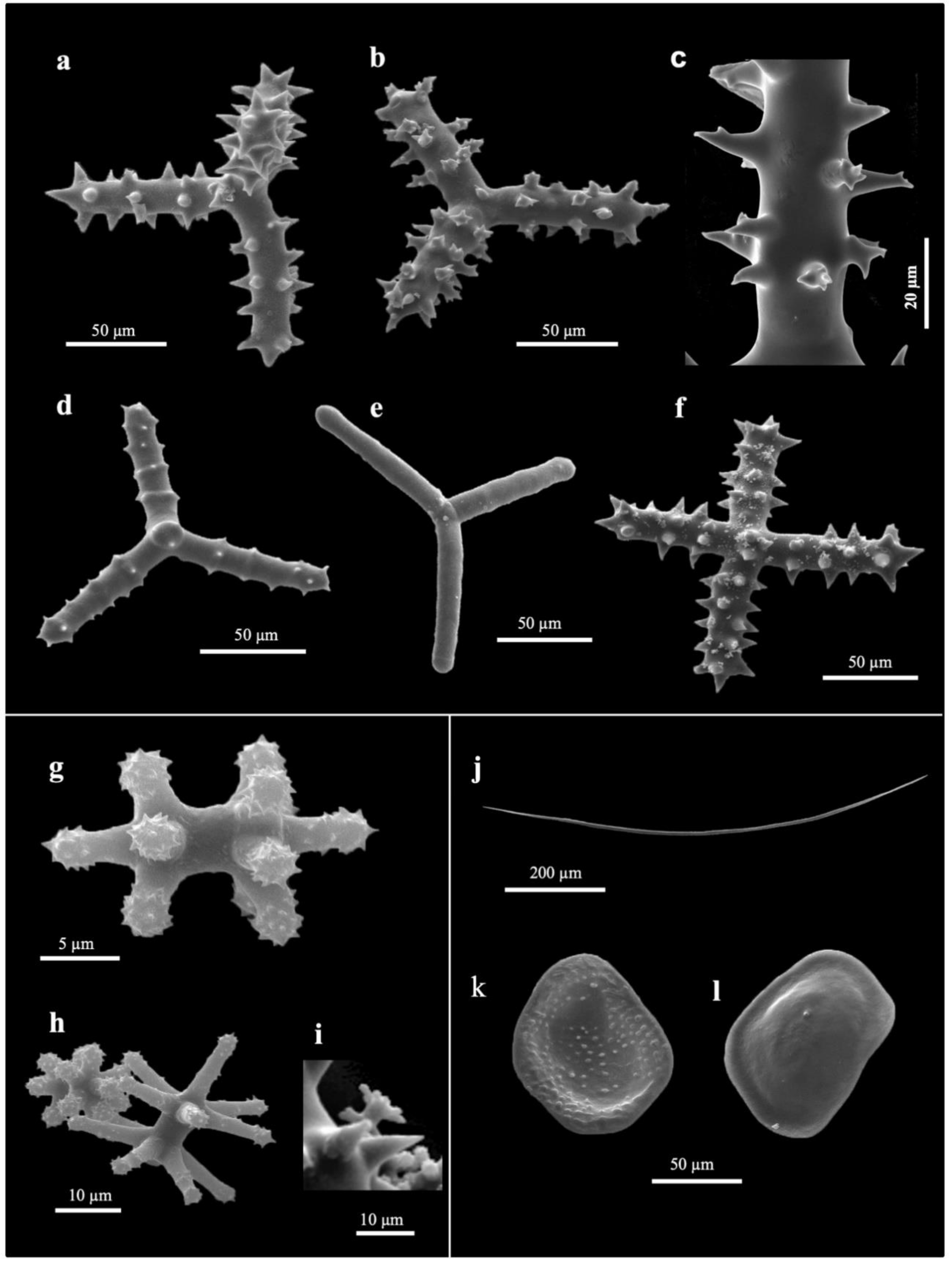
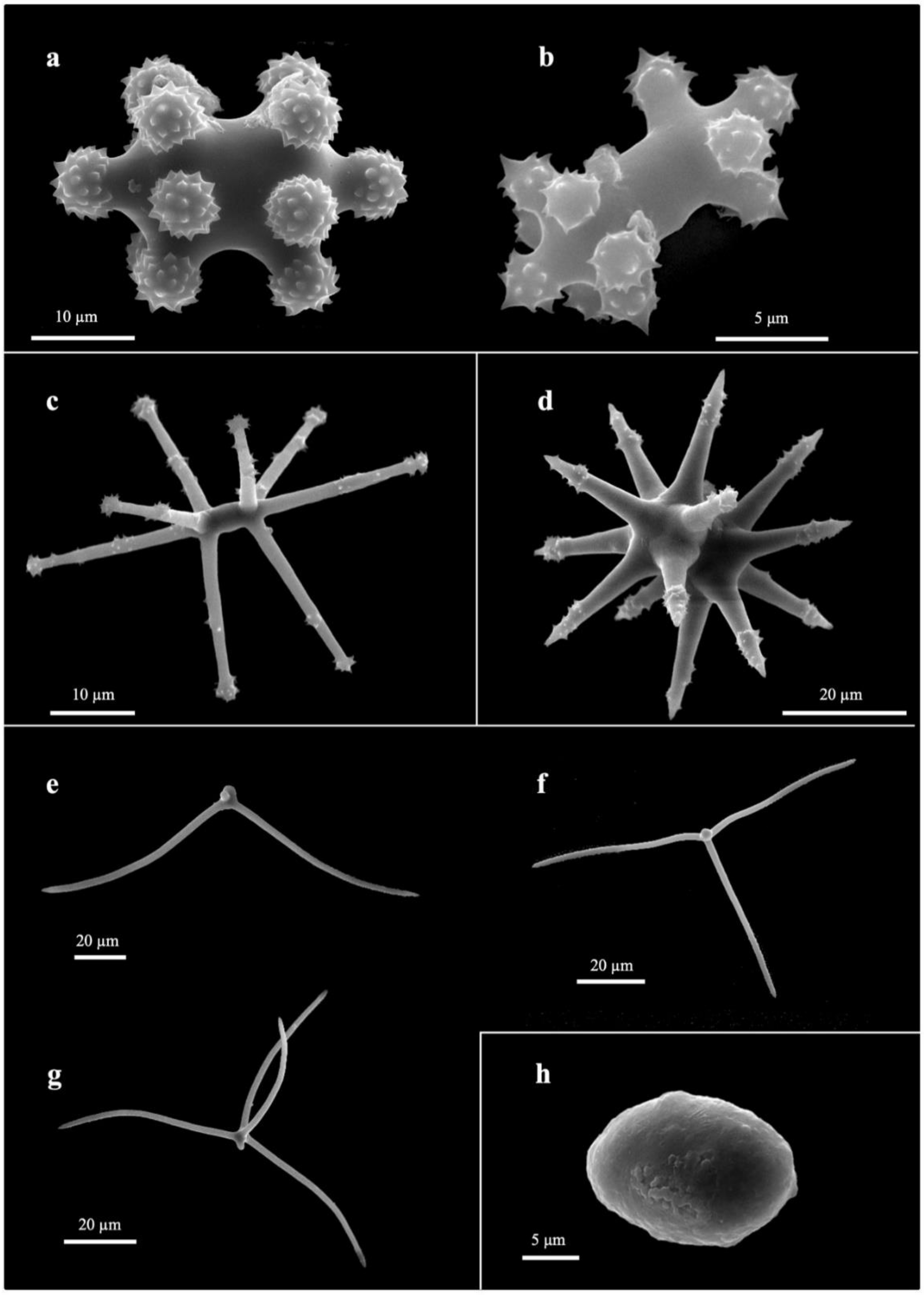
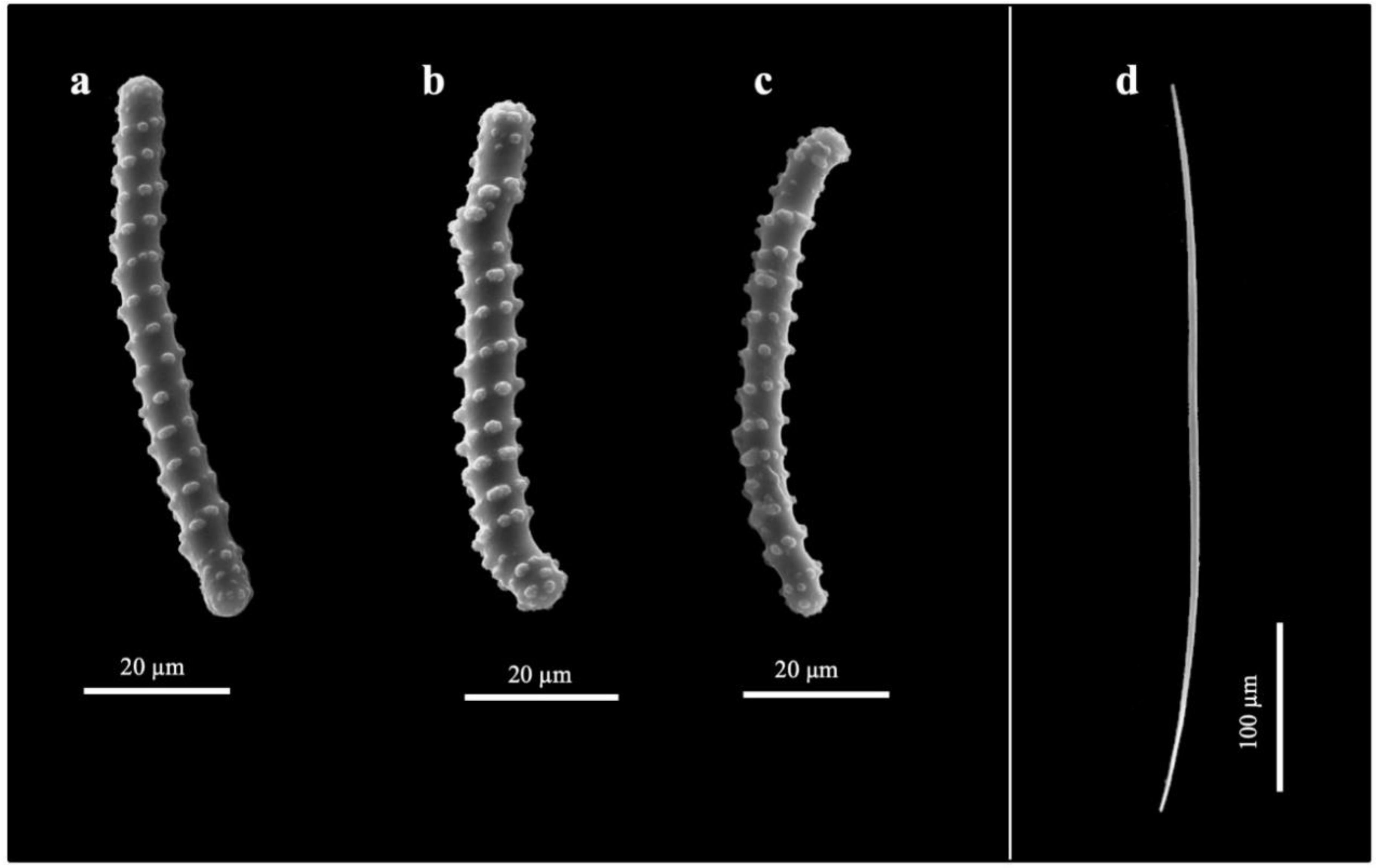
3.2. Taxonomy
4. General Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weinstein, D.K.; Maher, R.L.; Correa, A.M.S. Bioerosion. In Mesophotic Coral Ecosystems. Coral Reefs of the World; Loya, Y., Puglise, K., Bridge, T., Eds.; Springer: Cham, Switzerland, 2015; Volume 12, pp. 829–847. [Google Scholar] [CrossRef]
- Calcinai, B.; Cerrano, C.; Bavestrello, G.; Milanese, M.; Sarà, M. Il popolamento di spugne perforatrici di Corallium rubrum e di alcuni madreporari del Promontorio di Portofino. Boll. Mus. Ist. Biol. Univ. Genova 2002, 64, 53–59. [Google Scholar]
- Azzini, F.; Calcinai, B.; Iwasaki, N.; Bavestrello, G. A new species of Thoosa (Demospongiae, Hadromerida) excavating precious coral Corallium sp. From Midway. Ital. J. Zool. 2007, 74, 405–408. [Google Scholar] [CrossRef]
- Calcinai, B.; Cerrano, C.; Iwasakei, N.; Bavestrello, G. Sponges boring into precious corals: An overview with description of a new species of Alectona (Demospongiae, Alectonidae) and a world-wide identification key for the genus. Mar. Ecol. 2008, 29, 273–279. [Google Scholar] [CrossRef]
- Liverino, B. Il Corallo: Esperienze e Ricordi di un Corallaro; Bologna (Italy) Analisi: Bologna, Italy, 1984. [Google Scholar]
- Corriero, G.; Abbiati, M.; Santangelo, G. Sponges inhabiting a Mediterranean red coral population. Mar. Ecol. 1997, 18, 147–155. [Google Scholar] [CrossRef]
- Rützler, K. Family Clionaidae D’Orbigny, 1851. In Systema Porifera; Springer: Boston, MA, USA, 2002; pp. 173–185. [Google Scholar] [CrossRef]
- Vacelet, J. Planktonic armoured propagules of the excavating sponge Alectona (Porifera: Demospongiae) are larvae: Evidence from Alectona wallichii and A. mesatlantica sp. nov. Mem. Queensl. Mus. 1999, 44, 627–642. [Google Scholar]
- Calcinai, B.; Bavestrello, G.; Cerrano, C. Bioerosion micro-patterns as diagnostic characteristics in boring sponges. Boll. Mus. Ist. Biol. Univ. Genova. 2004, 68, 229–238. [Google Scholar]
- de Voogd, N.J.; Alvarez, B.; Boury-Esnault, N.; Carballo, J.L.; Cárdenas, P.; Díaz, M.-C.; Dohrmann, M.; Downey, R.; Hajdu, E.; Hooper, J.N.A.; et al. World Porifera Database. 2022. Available online: https://www.marinespecies.org/porifera (accessed on 3 February 2022).
- Schönberg, C.H.; Shields, G. Micro-computed tomography for studies on Entobia: Transparent substrate versus modern technology. In Current Developments in Bioerosion; Springer: Berlin/Heidelberg, Germany, 2008; pp. 147–164. [Google Scholar] [CrossRef]
- Beuck, L.; Vertino, A.; Stepina, E.; Karolczak, M.; Pfannkuche, O. Skeletal response of Lophelia pertusa (Scleractinia) to bioeroding sponge infestation visualised with micro-computed tomography. Facies 2007, 53, 157–176. [Google Scholar] [CrossRef]
- Färber, C.; Titschack, J.; Schönberg, C.H.L.; Ehrig, K.; Boos, K.; Baum, D.; Wisshak, M. Long-term macrobioerosion in the Mediterranean sea assessed by micro-computed tomography. Biogeosciences 2016, 13, 3461–3474. [Google Scholar] [CrossRef] [Green Version]
- Rützler, K. The burrowing sponges of Bermuda. Smithson. Cont. Zool. 1974, 165, 32. [Google Scholar] [CrossRef]
- Pica, D.; Calcinai, B.; Bertolino, M. Excavating sponges boring into the precious red coral from Cape Verde Archipelago. Biol. Mar. Mediterr. 2011, 18, 278–279. [Google Scholar]
- Carballo, J.L.; Bautista-Guerrero, E.; Cárdenas, P.; Cruz-Barraza, J.A.; Aguilar-Camacho, J.M. Molecular and morphological data from Thoosidae in favour of the creation of a new suborder of Tetractinellida. Syst. Biodivers. 2018, 16, 512–521. [Google Scholar] [CrossRef]
- Beuck, L.; Freiwald, A. Bioerosion patterns in a deep-water Lophelia pertusa (Scleractinia) thicket (Propeller Mound, northern Porcupine Seabight). In Cold-Water Corals and Ecosystems; Springer: Berlin/Heidelberg, Germany, 2005; pp. 915–936. [Google Scholar] [CrossRef]
- Bromley, R.G.; D’Alessandro, A. The ichnogenus Entobia from the Miocene, Pliocene and Pleistocene of southern Italy. Riv. Ital. Paleontol. Stratigr. 1984, 90, 227–296. [Google Scholar]
- Bromley, R.G.; D’Alessandro, A. Ichnological study of shallow marine endolithic sponges from the Italian coast. Riv. Ital. Paleontol. Stratigr. 1989, 95, 279–314. [Google Scholar]
- Bavestrello, G.; Calcinai, B.; Cerrano, C.; Sara, M. Alectona species from North-Western Pacific (Demospongiae: Clionidae). J. Mar. Biol. Ass. UK 1998, 78, 59–73. [Google Scholar] [CrossRef]
- Rosell, D.; Uriz, M.J. Excavating and endolithic sponge species (Porifera) from the Mediterranean: Species descriptions and identification key. Org. Divers. Evol. 2002, 2, 55–86. [Google Scholar] [CrossRef] [Green Version]
- Golubić, S.; Friedmann, I.; Schneider, J. The lithobiontic ecological niche, with special reference to microorganisms. J. Sediment. Res. 1981, 51, 475–478. [Google Scholar] [CrossRef]
- Wisshak, M.; Knaust, D.; Bromley, R.G. (Eds.) Trace Fossils as Indicators of Sedimentary Environments, Developments in Sedimentology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 64, pp. 213–243. [Google Scholar] [CrossRef]
- Davidson, T.M.; Altieri, A.H.; Ruiz, G.M.; Torchin, M.E. Bioerosion in a changing world: A conceptual framework. Ecol. Lett. 2018, 21, 422–438. [Google Scholar] [CrossRef] [Green Version]
- Bromley, R.G.; Tendal, O.S. Example of substrate competition and phobotropism between two clionid sponges. J. Zool. 1973, 169, 151–155. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Excavation Patterns and Spiculae Dimensions of the Boring Sponge Cliona celata from the SW Netherlands. Senckenb. Marit. 1982, 15, 55–85. [Google Scholar]
- Turicchia, E.; Abbiati, M.; Bettuzzi, M.; Calcinai, B.; Morigi, M.P.; Summers, A.P.; Ponti, M. Bioconstruction and Bioerosion in the Northern Adriatic Coralligenous Reefs Quantified by X-Ray Computed Tomography. Front. Mar. Sci. 2022, 8. [Google Scholar] [CrossRef]
- Calcinai, B.; Bavestrello, G.; Cerrano, C.; Sarà, M. Boring sponges living into precious corals from the Pacific Ocean. Ital. J. Zool. 2001, 68, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Bavestrello, G.; Calcinai, B.; Sarà, M. Two new species of Cliona (Porifera, Demospongiae) boring the scleraxis of Corallium elatius from the western Pacific. Ital. J. Zool. 1995, 62, 375–381. [Google Scholar] [CrossRef]
- Webb, A.E.; van Heuven, S.M.; de Bakker, D.M.; van Duyl, F.C.; Reichart, G.J.; de Nooijer, L.J. Combined effects of experimental acidification and eutrophication on reef sponge bioerosion rates. Front. Mar. Sci. 2017, 4, 311. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.K.; Mello-Athayde, M.A.; Schönberg, C.H.; Kline, D.I.; Hoegh-Guldberg, O.; Dove, S. Sponge biomass and bioerosion rates increase under ocean warming and acidification. Glob Chang. Biol. 2013, 19, 3581–3591. [Google Scholar] [CrossRef] [PubMed]
- Tunnicliffe, V. Breakage and propagation of the stony coral Acropora cervicornis. Proc. Nad. Acad. Sci. USA 1981, 78, 2427–2431. [Google Scholar] [CrossRef] [Green Version]
- Highsmith, R.C. Reproduction by fragmentation in corals. Mar. Ecol. Prog. Ser. 1982, 7, 207–226. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Calcinai, B.; Cattaneo-Vietti, R.; Morri, C.; Sarà, M. The role of sponge bioerosion in Mediterranean coralligenous accretion. In Mediterranean Ecosystems; Faranda, F.M., Guglielmo, L., Spezie, G.C., Eds.; Springer: Milano, Italy, 2001; pp. 235–240. [Google Scholar] [CrossRef]
- Grace, S. Winter quiescence, growth rate, and the release from competition in the temperate scleractinian coral Astrangia poculata (Ellis & Solander 1786). Northeastern Nat. 2017, 24, B119–B134. [Google Scholar] [CrossRef]
- Calcinai, B.; Bertolino, M.; Bavestrello, G.; Montori, S.; Mori, M.; Pica, D.; Valisano, L.; Cerrano, C. Comparison between the sponge fauna living outside and inside the coralligenous bioconstruction. A quantitative approach. Mediterr. Mar. Sci. 2015, 16, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Valisano, L.; Palma, M.; Pantaleo, U.; Calcinai, B.; Cerrano, C. Characterization of North–Western Mediterranean coralligenous assemblages by video surveys and evaluation of their structural complexity. Mar. Pollut. Bull. 2019, 148, 134–148. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulido Mantas, T.; Bavestrello, G.; Bertolino, M.; Cerrano, C.; Pica, D.; Roveta, C.; Calcinai, B. A 3D Innovative Approach Supporting the Description of Boring Sponges of the Precious Red Coral Corallium rubrum. J. Mar. Sci. Eng. 2022, 10, 868. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10070868
Pulido Mantas T, Bavestrello G, Bertolino M, Cerrano C, Pica D, Roveta C, Calcinai B. A 3D Innovative Approach Supporting the Description of Boring Sponges of the Precious Red Coral Corallium rubrum. Journal of Marine Science and Engineering. 2022; 10(7):868. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10070868
Chicago/Turabian StylePulido Mantas, Torcuato, Giorgio Bavestrello, Marco Bertolino, Carlo Cerrano, Daniela Pica, Camilla Roveta, and Barbara Calcinai. 2022. "A 3D Innovative Approach Supporting the Description of Boring Sponges of the Precious Red Coral Corallium rubrum" Journal of Marine Science and Engineering 10, no. 7: 868. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10070868