
In-Situ Estimates of Net Ecosystem Metabolisms in the Rocky Habitats of Dokdo Islets in the East Sea of Korea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Site Selection
2.3. Aquatic Eddy Covariance System (AEC)
2.4. AEC Measurements
2.5. Data Analysis and Calculation
2.6. Sampling and Species Richness
3. Results
3.1. Site Characteristics
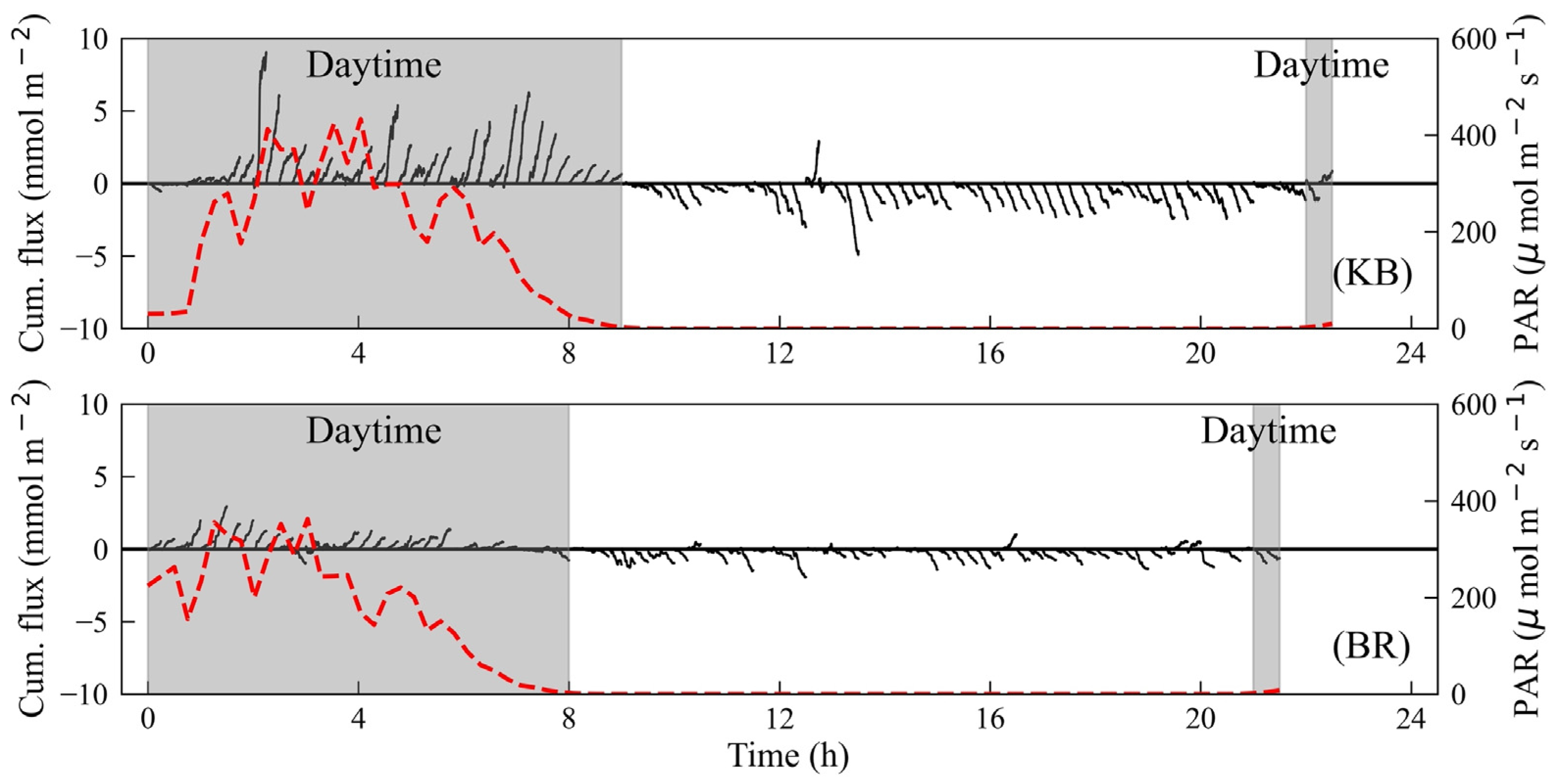
3.2. Time Series Data
3.3. Hourly Averaged O2 Flux with PAR
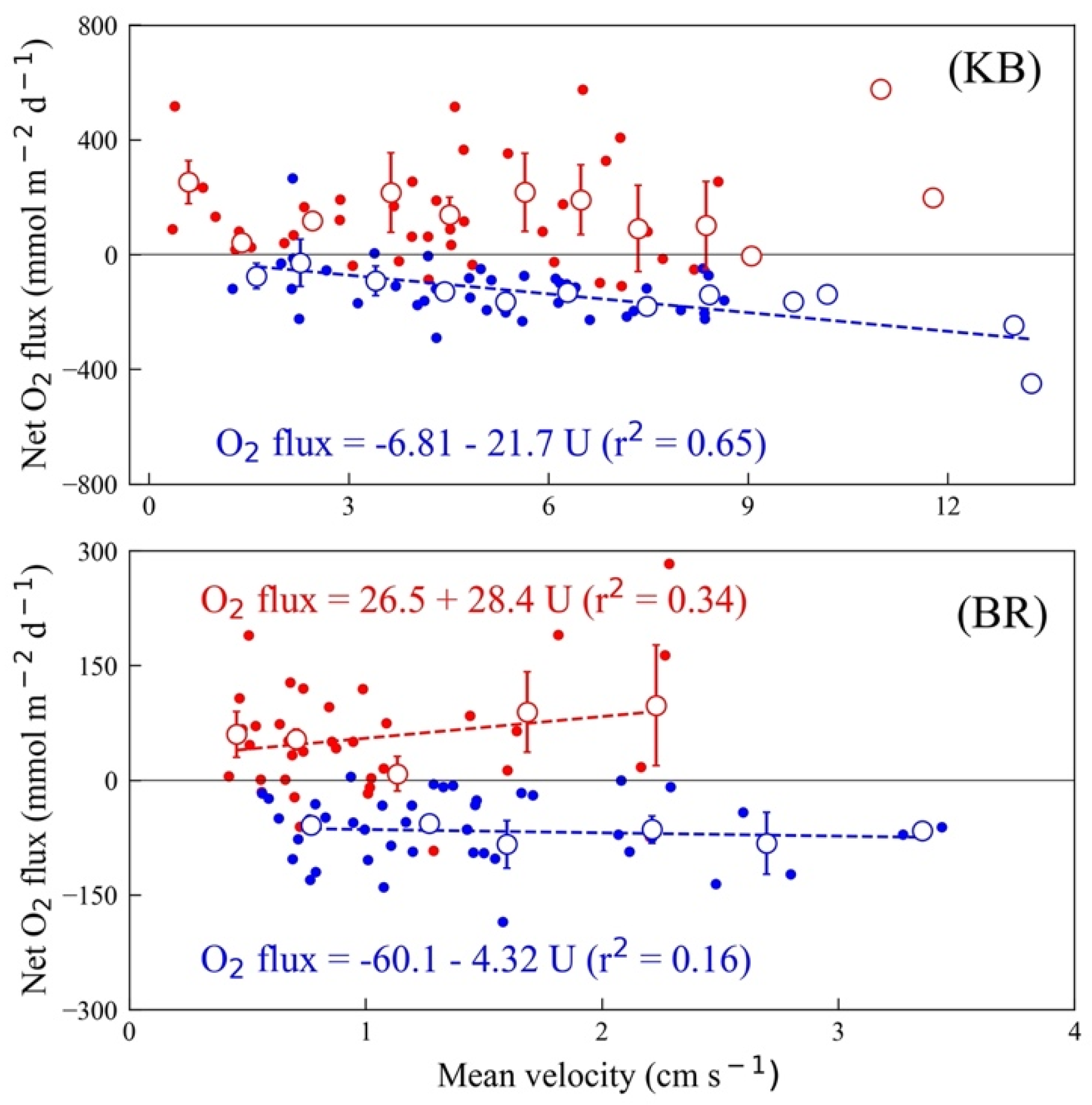
3.4. O2 Flux and Mean Current
3.5. Net Ecosystem Metabolism (NEM)
4. Discussion
4.1. O2 Flux Variability
4.2. P–I Relationship
4.3. NEM of Dokdo Islets
4.4. Implications and Further Suggestions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smale, D.A. Impacts of ocean warming on kelp forest ecosystems. New Phytol. 2020, 225, 1447–1454. [Google Scholar] [CrossRef] [Green Version]
- Teagle, H.; Hawkins, S.J.; Moore, P.J.; Smale, D.A. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. J. Exp. Mar. Biol. Ecol. 2017, 492, 81–98. [Google Scholar] [CrossRef]
- Wernberg, T.; Krumhansl, K.; Filbee-Dexter, K.; Pedersen, M.F. Status and trends for the world’s kelp forests. In World Seas: An Environmental Evaluation, 2nd ed.; Sheppard, C., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 57–78. [Google Scholar]
- Kim, M.K.; Kim, K.T. Studies on the seaweeds in the island of Ullungdo and Dokdo: I. Decrease of algal species compositions and changes of marine algal flora. Algae 2000, 15, 119–124. [Google Scholar]
- Kang, R.S.; Won, K.S.; Hong, K.P.; Kim, J.M. Population studies on the kelp Ecklonia cava and Eisenia bicyclis in Dokdo, Korea. Algae 2001, 16, 209–215. [Google Scholar]
- Tanaka, K.; Taino, S.; Haraguchi, H.; Prendergast, G.; Hiraoka, M. Warming off southwestern Japan linked to distributional shifts of subtidal canopy-forming seaweeds. Ecol. Evol. 2012, 2, 2854–2865. [Google Scholar] [CrossRef]
- Kim, M.K.; Shin, J.K.; Cha, J.H. Variation of species composition of benthic algae and whitening in the coast of Dokdo island during summer. Algae 2004, 19, 69–78. [Google Scholar] [CrossRef]
- Yoo, J.W.; Kim, H.J.; Lee, H.J.; Lee, C.G.; Kim, C.S.; Hong, J.S.; Hong, J.P.; Kim, D.S. Interaction between invertebrate grazers and seaweeds in the East Coast of Korea. Sea 2007, 12, 125–132. [Google Scholar]
- Duarte, C.M.; Cebriàn, J. The fate of marine autotrophic production. Limnol. Oceanogr. 1996, 41, 1758–1776. [Google Scholar] [CrossRef]
- Burdett, H.L.; Wright, H.; Smale, D.A. Photophysiological responses of canopy-forming kelp species to short-term acute warming. Front. Mar. Sci. 2019, 6, 516. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, K.L.; Rees, T.A.V.; Shears, N.T. A novel system for measuring in situ rates of photosynthesis and respiration of kelp. Mar. Ecol. Prog. Ser. 2015, 528, 101–115. [Google Scholar] [CrossRef] [Green Version]
- White, L.; Loisel, S.; Sevin, L.; Davoult, D. In situ estimates of kelp forest productivity in macro-tidal environments. Limnol. Oceanogr. 2021, 66, 4227–4239. [Google Scholar] [CrossRef]
- Hurd, C.L. Water motion, marine macroalgal physiology, and production. J. Phycol. 2000, 36, 453–472. [Google Scholar] [CrossRef]
- Noisette, F.; Hurd, C. Abiotic and biotic interactions in the diffusive boundary layer of kelp blades create a potential refuge from ocean acidification. Funct. Ecol. 2018, 32, 1329–1342. [Google Scholar] [CrossRef] [Green Version]
- Berg, P.; Røy, H.; Janssen, F.; Meyer, V.; Jørgensen, B.B.; Huettel, M.; DeBeer, D. Oxygen uptake by aquatic sediments measured with a novel non-invasive eddy-correlation technique. Mar. Ecol. Prog. Ser. 2003, 261, 75–83. [Google Scholar] [CrossRef]
- Hume, A.C.; Berg, P.; McGlathery, K.J. Dissolved oxygen fluxes and ecosystem metabolism in an eelgrass (Zostera marina) meadow measured with the eddy correlation technique. Limnol. Oceanogr. 2011, 56, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Long, M.H.; Berg, P.; de Beer, D.; Zieman, J.C. In situ coral reef oxygen metabolism: An eddy correlation study. PLoS ONE 2013, 8, e58581. [Google Scholar] [CrossRef] [Green Version]
- Attard, K.; Glud, R.N.; McGinnis, D.F.; Rysgaard, S. Seasonal rates of benthic primary production in a Greenland fjord measured by aquatic eddy correlation. Limnol. Oceanogr. 2014, 59, 1555–1569. [Google Scholar] [CrossRef]
- Rheuban, J.E.; Berg, P.; McGlathery, K.J. Multiple timescale processes drive ecosystem metabolism in eelgrass (Zostera marina) meadows. Mar. Ecol. Prog. Ser. 2014, 507, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Long, M.H.; Berg, P.; McGlathery, K.J.; Zieman, J.C. Sub-tropic seagrass ecosystem metabolism measured by eddy covariance. Mar. Ecol. Prog. Ser. 2015, 529, 75–90. [Google Scholar] [CrossRef]
- Rovelli, L.; Attard, K.M.; Bryant, L.D.; Flögel, S.; Stahl, H.; Roberts, J.M.; Linke, P.; Glud, R.N. Benthic O2 uptake of two cold-water coral communities estimated with the non-invasive eddy correlation technique. Mar. Ecol. Prog. Ser. 2015, 525, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Chipman, L.; Berg, P.; Huettel, M. Benthic oxygen fluxes measured by eddy covariance in permeable Gulf of Mexico shallow-water sands. Aquat. Geochem. 2016, 22, 529–554. [Google Scholar] [CrossRef]
- Volaric, M.P.; Berg, P.; Reidenbach, M.A. Oxygen metabolism of intertidal oyster reefs measured by aquatic eddy covariance. Mar. Ecol. Prog. Ser. 2018, 599, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Camillini, N.; Attard, K.M.; Eyre, B.D.; Glud, R.N. Resolving community metabolism of eelgrass Zostera marina meadows by benthic flume-chambers and eddy covariance in dynamic coastal environments. Mar. Ecol. Prog. Ser. 2021, 661, 97–114. [Google Scholar] [CrossRef]
- Berg, P.; Røy, H.; Wiberg, P.L. Eddy correlation flux measurements: The sediment surface area contributes to the flux. Limnol. Oceanogr. 2007, 52, 1672–1682. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Park, J.W.; Lee, M.H.; Park, C.H. Detailed bathymetry and submarine terraces in the coastal area of the Dokdo Volcano in the Ulleung Basin, the East Sea (Sea of Japan). J. Coast. Res. 2013, 65, 523–528. [Google Scholar] [CrossRef]
- Park, K.-A.; Ullman, D.S.; Kim, K.; Chung, Y.J.; Kim, K.-R. Spatial and temporal variability of satellite-observed subpolar front in the East/Japan Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 453–470. [Google Scholar] [CrossRef]
- Park, K.-A.; Woo, H.-J.; Ryu, J.-H. Spatial scales of mesoscale eddies from GOCI chlorophyll-a concentration images in the East/Japan Sea. Ocean Sci. J. 2012, 47, 347–358. [Google Scholar] [CrossRef]
- Kim, D.; Yang, E.J.; Kim, K.H.; Shin, C.-W.; Park, J.; Yoo, S.; Hyun, J.-H. Impact of an anticyclonic eddy on the summer nutrient and chlorophyll a distribution in the Ulleung Basin, East Sea (Japan Sea). ICES J. Mar. Sci. 2012, 69, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.; Park, J. Why is the southwest the most productive region of the East Sea/Sea of Japan? J. Mar. Syst. 2009, 78, 301–315. [Google Scholar] [CrossRef]
- Song, S.J.; Park, J.; Ryu, J.; Rho, H.S.; Kim, W.; Khim, J.S. Biodiversity hotspot for marine invertebrates around the Dokdo, East Sea, Korea: Ecological checklist revisited. Mar. Pollut. Bull. 2017, 119, 162–170. [Google Scholar] [CrossRef]
- Park, S.K.; Lee, J.R.; Heo, J.S.; An, D.S.; Lee, H.P.; Choi, H.G. Marine algae flora and ecological role of Eisenia bicyclis in Dokdo, East Sea, Korea. Korean J. Environ. Ecol. 2014, 28, 613–626. (In Korean) [Google Scholar] [CrossRef]
- Hwang, S.I.; Kim, D.K.; Sung, B.J.; Jun, S.K.; Bae, J.I.; Jeon, B.H. Effects of climate change on whitening event proliferation the Coast of Jeju. Korean J. Environ. Ecol. 2017, 31, 529–536. [Google Scholar] [CrossRef]
- Lee, J.S.; Kang, D.-J.; Hineva, E.; Slabakova, V.; Todorova, V.; Park, J.; Cho, J.-H. Estimation of net ecosystem metabolism of seagrass meadows in the coastal waters of the East Sea and Black Sea using the Noninvasive Eddy Covariance technique. Ocean Sci. J. 2017, 52, 243–256. [Google Scholar] [CrossRef]
- Berg, P.; Koopmans, D.J.; Huettel, M.; Hua, L.; Mori, K.; Wüest, A. A new robust oxygen-temperature sensor for aquatic eddy covariance measurement. Limnol. Oceanogr. Methods 2015, 14, 151–167. [Google Scholar] [CrossRef] [Green Version]
- Huettel, M.; Berg, P.; Merikhi, A. Technical note: Measurements and data analysis of sediment–water oxygen flux using a new dual-optode eddy covariance instrument. Biogeosciences 2020, 17, 4459–4476. [Google Scholar] [CrossRef]
- Rovelli, L.; Attard, K.M.; Binley, A.; Heppell, C.M.; Stahl, H.; Trimmer, M.; Glud, R.N. Reach-scale river metabolism across contrasting sub-catchment geologies: Effect of light and hydrology. Limnol. Oceanogr. 2017, 62, S381–S399. [Google Scholar] [CrossRef] [Green Version]
- Long, M.H. Aquatic biogeochemical eddy covariance fluxes in the presence wave. J. Geophys. Res. Oceans 2021, 126, e2020JC016637. [Google Scholar] [CrossRef]
- Goring, D.G.; Nikora, V.I. Despiking acoustic doppler velocimeter data. J. Hydraul. Eng. 2002, 128, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Jassby, A.D.; Platt, T. Mathematical formulation of relationship between photosynthesis and light for phytoplankton. Limnol. Oceanogr. 1976, 21, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Donis, D.; Holtappels, M.; Noss, C.; Cathalot, C.; Hancke, K.; Polsenaere, P.; McGinnis, D.F. An assessment of the precision and confidence of aquatic eddy correlation measurements. J. Atmos. Ocean. Technol. 2015, 32, 642–655. [Google Scholar] [CrossRef] [Green Version]
- Reimers, C.E.; Fogaren, K.E. Bottom boundary layer oxygen fluxes during winter on the Oregon Shelf. J. Geophys. Res. Oceans 2021, 126, e2020JC016828. [Google Scholar] [CrossRef]
- Reidenbach, M.A.; Koseff, J.R.; Monismith, S.G. Laboratory experiments of fine-scale mixing and mass transport within a coral canopy. Phys. Fluids 2007, 19, 075107. [Google Scholar] [CrossRef]
- Hansen, J.C.R.; Fourquerean, J.W. Wave and tidally driven flows in eelgrass beds and their effect on sediment suspension. Mar. Ecol. Prog. Ser. 2012, 448, 271–287. [Google Scholar] [CrossRef] [Green Version]
- Murakami, H.; Yukihiko, S.; Akira, K.; Yasutsugu, Y. Phytosynthetic performances of temperate Sargassum and Kelp species growing in the same habitat. Algae 2004, 19, 207–216. [Google Scholar] [CrossRef]
- Binzer, T.K.; Sand-Jensen, N.T.; Middelboe, A.L. Community photosynthesis of aquatic macrophytes. Limnol. Oceanogr. 2006, 51, 2722–2733. [Google Scholar] [CrossRef]
- Miller, R.J.; Reed, D.C.; Brzezinski, M.A. Partitioning of primary production among giant kelp (Macrocystis pyrifera), understory macroalgae, and phytoplankton on a temperate reef. Limnol. Oceanogr. 2011, 56, 119–132. [Google Scholar] [CrossRef]
- Barrón, C.; Apostolaki, E.T.; Duarte, C.M. Dissolved organic carbon fluxes by seagrass meadows and macroalgal beds. Front. Mar. Sci. 2014, 1, 42. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.; Konar, B.; Kim, J.-H.; Gabara, S.; Sullaway, G.; McHugh, T.; Spector, M.; Small, S. Marine deforestation leads to widespread loss of ecosystem function. PLoS ONE 2020, 15, e0226173. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Yoshida, G.; Hori, M.; Umezawa, Y.; Moki, H.; Kuwae, T. Macroalgal metabolism and lateral carbon flows can create significant carbon sinks. Biogeosciences 2020, 17, 2425–2440. [Google Scholar] [CrossRef]
- Staehr, P.A.; Testa, J.M.; Michael Kemp, W.; Cole, J.J.; Sand-Jensen, K.; Smith, S.V. The metabolism of aquatic ecosystems: History, applications, and future challenges. Aquat. Sci. 2012, 74, 15–29. [Google Scholar] [CrossRef]
- Graham, M.H.; Bourque, B.J.; Erlandson, J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Newell, R.C.; Lucas, M.I.; Velimirov, B.; Seiderer, L.J. Quantitative significance of dissolved organic losses following fragmentation of kelp (Ecklonia maxima and Laminaria pallida). Mar. Ecol. Prog. Ser. 1980, 2, 45–59. [Google Scholar] [CrossRef]
- Stuart, V.; Lucas, M.I.; Newell, R.C. Heterotrophic utilisation of particulate matter from the kelp Laminaria pallida. Mar. Ecol. Prog. Ser. 1981, 4, 337–348. [Google Scholar] [CrossRef]
- Bustamante, R.H.; Branch, G.M.; Eekhout, S. Maintenance of an exceptional grazer biomass in South Africa: Subsidy by subtidal kelps. Ecology 1995, 76, 2314–2329. [Google Scholar] [CrossRef]
- Griffiths, C.L.; Stenton-Dozey, J.M.E.; Koop, K. Kelp wrack and the flow of energy through a sandy beach ecosystem. In Sandy Beaches as Ecosystems. Developments in Hydrobiology, 1st ed.; McLachlan, A., Erasmus, T., Eds.; Springer: Dordrecht, South Africa, 1983; Volume 19, pp. 547–556. [Google Scholar] [CrossRef]
- Smale, D.A.; Pessarrodona, A.; King, N.; Moore, P. Examining the production, export, and immediate fate of kelp detritus on open-coast subtidal reeds in the Northeast Atlantic. Limnol. Oceanog. 2021, 9999, 1–14. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Duarte, C.M. Substantial role of macroalgae in marine carbon sequestration. Nat. Geosci. 2016, 9, 737–742. [Google Scholar] [CrossRef]
- NIFS. A Study on Restoration of Barren Ground in Jeju Island; NIFS: Busan, Korea, 2008. [Google Scholar]
- Kim, Y.-D.; Park, M.-S.; Yoo, H.-I.; Min, B.-H.; Moon, T.-S.; Choi, H.-G. Seasonal variation in subtidal seaweed community structure at Hajung, on the southeast coast of Korea. Korean J. Fish. Aquat. Sci. 2011, 44, 740–746. [Google Scholar] [CrossRef] [Green Version]
- Baek, J.-U.; Lee, J.S.; Kim, S.-H.; Lee, T.; Jung, S.W.; Lee, W.-C.; Kim, K.-T.; An, S.-U. Effects of Irradiance and Temperature on the Photosynthesis of the Crustose Coralline Algae Pneophyllum fragile (Corallinales, Rhodophyta) in the Coastal Waters of Korea. JMSE 2022, 10, 851. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pmax | α | Ik | Ic | Rd | GPP | R | NEM | |
|---|---|---|---|---|---|---|---|---|
| KB | 257 | 4.1 | 62 | 16 | 65 | 163 | 154 | 8 |
| BR | 168 | 0.8 | 205 | 59 | 48 | 51 | 63 | −12 |
| Location | Species | R | GPP | NEM | Method | Frequency | References |
|---|---|---|---|---|---|---|---|
| Santa Barbara, USA | Macrocystis pyrifera | −216–−54.2 | 65.1–911 | −108–130 | chamber | seasonal | [47] |
| Bolinao, (Philippines) & Trondheim, (Norway) | Caulerpa racemose Laminaria | −11.7–−185 | 41.7–243 | 9.11–107 | chamber | monthly | [48] |
| Aleutian archipelago, USA | Macrocystis pyrifera, Sargassum horneri | −166 | 150 | −19.9 | benthic isolation tents | summer | [49] |
| Heigun Island, Japan | Sargassum algae | −337–−351 | - | 393–1794 | field bag | seasonal | [50] |
| Dokdo islet, Korea | Eisenia bicyclis | −154 | 163 | 8 | AEC | autumn | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.S.; Kim, S.-H.; Min, W.-G.; Choi, D.M.; Lee, E.K.; Kim, K.-T.; An, S.-U.; Baek, J.-W.; Lee, W.-C.; Park, C.H. In-Situ Estimates of Net Ecosystem Metabolisms in the Rocky Habitats of Dokdo Islets in the East Sea of Korea. J. Mar. Sci. Eng. 2022, 10, 887. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10070887
Lee JS, Kim S-H, Min W-G, Choi DM, Lee EK, Kim K-T, An S-U, Baek J-W, Lee W-C, Park CH. In-Situ Estimates of Net Ecosystem Metabolisms in the Rocky Habitats of Dokdo Islets in the East Sea of Korea. Journal of Marine Science and Engineering. 2022; 10(7):887. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10070887
Chicago/Turabian StyleLee, Jae Seong, Sung-Han Kim, Won-Gi Min, Dong Mun Choi, Eun Kyung Lee, Kyung-Tae Kim, Sung-Uk An, Ju-Wook Baek, Won-Chan Lee, and Chan Hong Park. 2022. "In-Situ Estimates of Net Ecosystem Metabolisms in the Rocky Habitats of Dokdo Islets in the East Sea of Korea" Journal of Marine Science and Engineering 10, no. 7: 887. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10070887