Fish Assemblage Structure Comparison between Freshwater and Estuarine Habitats in the Lower Nakdong River, South Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Ecological Guild and Habitat Type of Fishes
2.3. Data Analyses
3. Results
3.1. Environmental Variables
3.2. Fish Species Composition
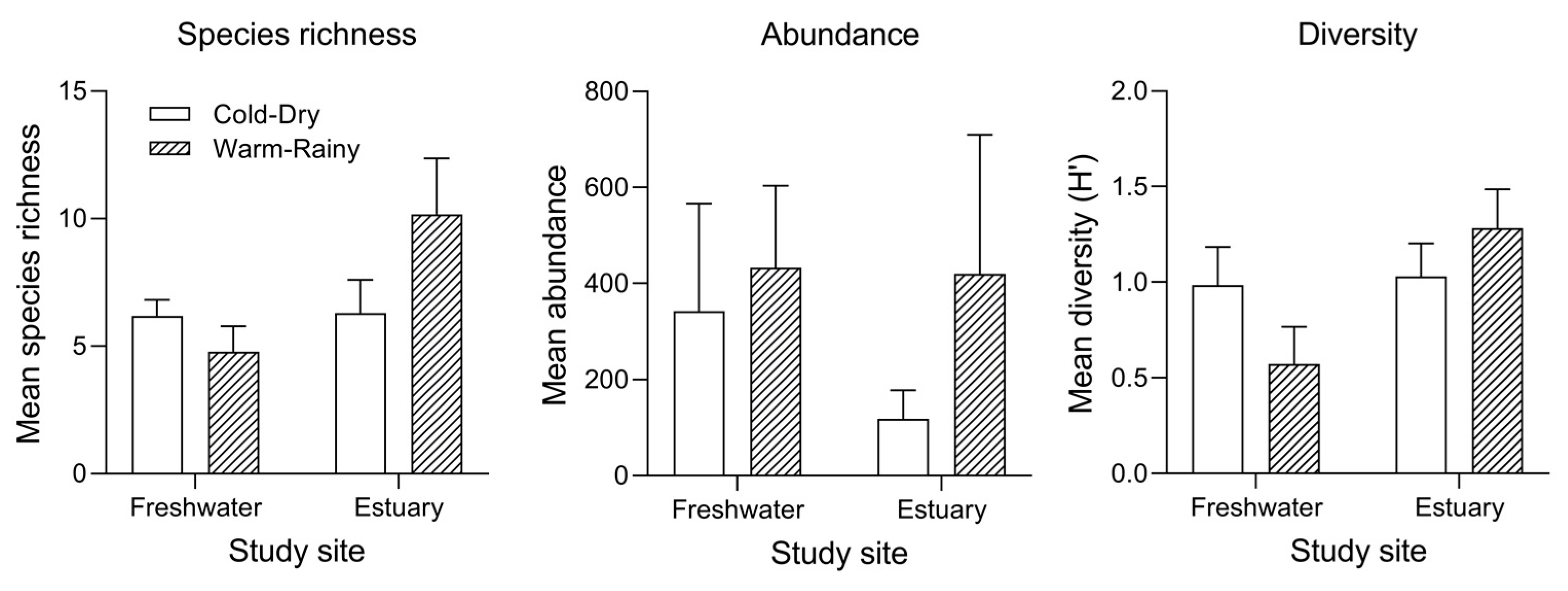
3.3. Spatial and Seasonal Variation in Species Richness, Abundance and Diversity
3.4. Fish Assemblage Structure
3.5. Ecological Guild and Habitat Type of Fish Assemblages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Day, J.W., Jr.; Kemp, W.M.; Yáñez-Arancibia, A.; Crump, B.C. Estuarine Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Dolbeth, M.; Cardoso, P.G.; Ferreira, S.M.; Verdelhos, T.; Raffaelli, D.; Pardal, M.A. Anthropogenic and natural disturbance effects on a macrobenthic estuarine community over a 10-year period. Mar. Pollut. Bull. 2007, 54, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaonkar, U.V.; Sivadas, S.K.; Ingole, B.S. Effect of tropical rainfall in structuring the macrobenthic community of Mandovi estuary, west coast of India. J. Mar. Biol. Assoc. U. K. 2013, 93, 1727–1738. [Google Scholar] [CrossRef] [Green Version]
- Schuchardt, B.; Haesloop, U.; Schirmer, M. The tidal freshwater reach of the Weser estuary: Riverine or estuarine? Neth. J. Aquat. Ecol. 1993, 27, 215–226. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Raffaelli, D.; Lillebo, A.I.; Verdelhos, T.; Pardal, M.A. The impact of extreme flooding events and anthropogenic stressors on the macrobenthic communities’ dynamics. Estuar. Coast. Shelf Sci. 2008, 76, 553–565. [Google Scholar] [CrossRef] [Green Version]
- McDowall, R.M. The evolution of diadromy in fishes (revisited) and its place in phylogenetic analysis. Rev. Fish. Biol. Fish. 1997, 7, 443–462. [Google Scholar] [CrossRef]
- Kim, I.S.; Choi, Y.; Lee, C.; Lee, Y.; Kim, B.; Kim, J.; Choi, Y.; Lee, Y.; Kim, B. Illustrated Book of Korean Fishes; Kyo-Hak: Seoul, Korea, 2005. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Version (12/2019). Available online: https://www.fishbase.org/ (accessed on 16 April 2020).
- Hwang, J.-S. The fish migration through fishlock at Youngam lake. J. Korea Water Resour. Assoc. 2000, 33, 561–568. [Google Scholar]
- Kang, E.-J.; Yang, H.; Lee, H.-H.; Kim, K.-S.; Kim, C.-H. Characteristics of fish fauna collected from near estuaries bank and fish-way on the bank of Naktong river. Korean J. Ichthyol. 2012, 24, 201–219. [Google Scholar]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities-the roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar] [CrossRef]
- Joy, M.K.; Death, R.G. Control of freshwater fish and crayfish community structure in Taranaki, New Zealand: Dams, diadromy or habitat structure? Freshw. Biol. 2001, 46, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Gillanders, B.M.; Kingsford, M.J. Impact of changes in flow of freshwater on estuarine and open coastal habitats and the associated organisms. Oceanogr. Mar. Biol. An Annu. Rev. 2002, 40, 233–309. [Google Scholar]
- Gehrke, P.C.; Gilligan, D.M.; Barwick, M. Changes in fish communities of the Shoalhaven River 20 years after construction of Tallowa Dam, Australia. River Res. Appl. 2002, 18, 265–286. [Google Scholar] [CrossRef]
- Rolls, R.J. The role of life-history and location of barriers to migration in the spatial distribution and conservation of fish assemblages in a coastal river system. Biol. Conserv. 2011, 144, 339–349. [Google Scholar] [CrossRef]
- Han, M.; Fukushima, M.; Kameyama, S.; Fukushima, T.; Matsushita, B. How do dams affect freshwater fish distributions in Japan? Statistical analysis of native and nonnative species with various life histories. Ecol. Res. 2008, 23, 735–743. [Google Scholar] [CrossRef]
- Jeon, S. Studies on the fish fauna of the estuary area of Naktong river, Korea. Bull. Kor. Assoc. Conser. Nat. 1987, 9, 77–90. [Google Scholar]
- Flemer, D.A.; Champ, M.A. What is the future fate of estuaries given nutrient over-enrichment, freshwater diversion and low flows? Mar. Pollut. Bull. 2006, 52, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.-K.; Kim, C.-H. A study on the changes of the molluscan and crustacean fauna after the construction of the Naktong estuary barrage. Korean J. Fish. Aquat. Sci. 1992, 25, 265–281. [Google Scholar]
- Kwak, S.N.; Huh, S.-H. Changes in species composition of fishes in the Nakdong River Estuary. Korean J. Fish. Aquat. Sci. 2003, 36, 129–135. [Google Scholar]
- Park, S.; Lee, J.; Choi, J.U.; Heo, N.; An, S. Study on the long-term changes in water quality and benthic ecology and evaluation on effect of the barrage in Nakdong River estuary. J. Wetl. Res. 2016, 18, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Verhelst, P.; Buysse, D.; Reubens, J.; Pauwels, I.; Aelterman, B.; Van Hoey, S.; Goethals, P.; Coeck, J.; Moens, T.; Mouton, A. Downstream migration of European eel (Anguilla anguilla L.) in an anthropogenically regulated freshwater system: Implications for management. Fish. Res. 2018, 199, 252–262. [Google Scholar] [CrossRef] [Green Version]
- Potter, I.C.; Claridge, P.N.; Warwick, R.M. Consistency of Seasonal-Changes in an Estuarine Fish Assemblage. Mar. Ecol. Prog. Ser. 1986, 32, 217–228. [Google Scholar] [CrossRef]
- Hood, W.G. Indirect environmental effects of dikes on estuarine tidal channels: Thinking outside of the dike for habitat restoration and monitoring. Estuaries 2004, 27, 273–282. [Google Scholar] [CrossRef]
- Zampatti, B.P.; Bice, C.M.; Jennings, P.R. Temporal variability in fish assemblage structure and recruitment in a freshwater-deprived estuary: The Coorong, Australia. Mar. Freshw. Res. 2010, 61, 1298–1312. [Google Scholar] [CrossRef]
- Hwang, S.W.; Lee, T.W.; Hwang, H.B.; Do Hwang, S. Temporal occurrence patterns and habitat use of fish in the Mangyeong Estuary on the western coast of Korea in 2003, before dike enclosure. Ocean Sci. J. 2011, 46, 307–314. [Google Scholar] [CrossRef]
- Park, J.M.; Huh, S.H.; Baeck, G.W. Temporal variations of fish assemblage in the surf zone of the Nakdong River Estuary, southeastern Korea. Anim. Cells Syst. 2015, 19, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Mariani, S. Can spatial distribution of ichthyofauna describe marine influence on coastal lagoons? A central Mediterranean case study. Estuar. Coast. Shelf Sci. 2001, 52, 261–267. [Google Scholar] [CrossRef]
- Ayvazian, S.; Hyndes, G. Surf-zone fish assemblages in south-western Australia: Do adjacent nearshore habitats and the warm Leeuwin Current influence the characteristics of the fish fauna? Mar. Biol. 1995, 122, 527–536. [Google Scholar] [CrossRef]
- Akin, S.; Buhan, E.; Winemiller, K.O.; Yimaz, H. Fish assemblage structure of Koycegiz Lagoon-Estuary, Turkey: Spatial and temporal distribution patterns in relation to environmental variation. Estuar. Coast. Shelf Sci. 2005, 64, 671–684. [Google Scholar] [CrossRef]
- Potter, I.C.; Bird, D.J.; Claridge, P.; Clarke, K.; Hyndes, G.; Newton, L. Fish fauna of the Severn Estuary. Are there long-term changes in abundance and species composition and are the recruitment patterns of the main marine species correlated? J. Exp. Mar. Biol. Ecol. 2001, 258, 15–37. [Google Scholar] [CrossRef]
- Contente, R.F.; Stefanoni, M.F.; Spach, H.L. Fish assemblage structure in an estuary of the Atlantic Forest biodiversity hotspot (southern Brazil). Ichthyol. Res. 2011, 58, 38–50. [Google Scholar] [CrossRef]
- Fausch, K.D.; Torgersen, C.E.; Baxter, C.V.; Li, H.W. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes: A continuous view of the river is needed to understand how processes interacting among scales set the context for stream fishes and their habitat. BioScience 2002, 52, 483–498. [Google Scholar] [CrossRef] [Green Version]
- Durance, I.; Lepichon, C.; Ormerod, S.J. Recognizing the importance of scale in the ecology and management of riverine fish. River Res. Appl. 2006, 22, 1143–1152. [Google Scholar] [CrossRef]
- Chyung, M.K. The Fishes of Korea; Il Ji Sa Publishing Co.: Seoul, Korea, 1977. [Google Scholar]
- Shannon, C.; Weaver, W. The Mathematical Theory of Communication; Illinois University Press: Urbana, IL, USA, 1949. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research; Freeman New York: New York, NY, USA, 1981. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Plymouth Marine Laboratory: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.; Gorley, R. PRIMER V7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Martino, E.J.; Able, K.W. Fish assemblages across the marine to low salinity transition zone of a temperate estuary. Estuar. Coast. Shelf Sci. 2003, 56, 969–987. [Google Scholar] [CrossRef]
- De Moura, P.M.; Vieira, J.P.; Garcia, A.M. Fish abundance and species richness across an estuarine-freshwater ecosystem in the Neotropics. Hydrobiologia 2012, 696, 107–122. [Google Scholar] [CrossRef]
- Yoklavich, M.M.; Cailliet, G.M.; Barry, J.P.; Ambrose, D.A.; Antrim, B.S. Temporal and Spatial Patterns in Abundance and Diversity of Fish Assemblages in Elkhorn Slough, California. Estuaries 1991, 14, 465–480. [Google Scholar] [CrossRef]
- Sosa-Lopez, A.; Mouillot, D.; Ramos-Miranda, J.; Flores-Hernandez, D.; Chi, T.D. Fish species richness decreases with salinity in tropical coastal lagoons. J. Biogeogr. 2007, 34, 52–61. [Google Scholar] [CrossRef]
- Vasconcelos, R.P.; Henriques, S.; Franca, S.; Pasquaud, S.; Cardoso, I.; Laborde, M.; Cabral, H.N. Global patterns and predictors of fish species richness in estuaries. J. Anim. Ecol. 2015, 84, 1331–1341. [Google Scholar] [CrossRef]
- Martinho, F.; Leitão, R.; Neto, J.M.; Cabral, H.N.; Marques, J.C.; Pardal, M.A. The use of nursery areas by juvenile fish in a temperate estuary, Portugal. Hydrobiologia 2007, 587, 281–290. [Google Scholar] [CrossRef]
- Ikejima, K.; Tongnunui, P.; Medej, T.; Taniuchi, T. Juvenile and small fishes in a mangrove estuary in Trang province, Thailand: Seasonal and habitat differences. Estuar. Coast. Shelf Sci. 2003, 56, 447–457. [Google Scholar] [CrossRef]
- Vidy, G.; Darboe, F.S.; Mbye, E.M. Juvenile fish assemblages in the creeks of the Gambia Estuary. Aquat. Living Resour. 2004, 17, 56–64. [Google Scholar] [CrossRef]
- Plavan, A.A.; Passadore, C.; Gimenez, L. Fish assemblage in a temperate estuary on the uruguayan coast: Seasonal variation and environmental influence. Braz. J. Oceanogr. 2010, 58, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Weitkamp, L.A.; Bentley, P.J.; Litz, M.N.C. Seasonal and interannual variation in juvenile salmonids and associated fish assemblage in open waters of the lower Columbia River estuary. Fish. Bull. 2012, 110, 426–450. [Google Scholar]
- Hwang, S.-W.; Hwang, H.-B.; Noh, H.-S.; Lee, T.-W. Seasonal variation in species composition of fish collected by a bag net in the Geum River estuary, Korea. Korean J. Fish. Aquat. Sci. 2005, 38, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Han, K.; Kang, C.; Kim, J. Commercial Fishes of the Coastal and Offshore Waters in Korea; Hangel: Busan. Korea, 2004. [Google Scholar]
- Sung, N.J.; Lee, E.H.; Ha, B.S. Studies on the food from fresh water fish-3. Composition in amino acids and nucleotides of the dorsal muscle of some fresh water fishes, Sweet Fish, Cornet Fish, Mandarin Fish and Read Fish. J. Korean Soc. Food Sci. Nutr. 1984, 13, 163–168. [Google Scholar]
- Yoon, J.D.; Kim, J.H.; In, D.S.; Yu, J.J.; Hur, M.; Chang, K.H.; Jang, M.H. Evaluation of movement pattern of Erythroculter erythropterus inhabit in the mid-lower part of Nakdong River using acoustic telemetry. Korean J. Ecol. Environ. 2012, 45, 403–411. [Google Scholar] [CrossRef]
- Jang, S.-H.; Zhang, C.-I.; Na, J.-H.; Lee, J.-H. Analysis of trophic structures and energy flows in aquatic ecosystem of the lower reaches of the Nakdong River. Korean J. Environ. Biol. 2008, 26, 292–302. [Google Scholar]
- Jeong, J.M.; Park, J.M.; Huh, S.H.; Ye, S.J.; Kim, H.J.; Baeck, G.W. Seasonal variation in the species composition of fish assemblages in the coastal waters off Gadeok-do, South sea, Korea. Korean J. Fish. Aquat. Sci. 2013, 46, 948–956. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Kawaguchi, Y.; Yamanaka, H.; Okunaka, T.; Nakajima, J.; Mitani, Y.; Shimatani, Y.; Mukai, T.; Onikura, N. Predicting the spatial distribution of the invasive piscivorous chub (Opsariichthys uncirostris uncirostris) in the irrigation ditches of Kyushu, Japan: A tool for the risk management of biological invasions. Biol. Invasions 2010, 12, 3677–3686. [Google Scholar] [CrossRef]
- Williams, J.R.; Dellapenna, T.M.; Lee, G.H. Shifts in depositional environments as a natural response to anthropogenic alterations: Nakdong Estuary, South Korea. Mar. Geol. 2013, 343, 47–61. [Google Scholar] [CrossRef]
- Park, S.R.; Kim, J.H.; Kang, C.K.; An, S.; Chung, I.K.; Kim, J.H.; Lee, K.S. Current status and ecological roles of Zostera marina after recovery from large-scale reclamation in the Nakdong River estuary, Korea. Estuar. Coast. Shelf Sci. 2009, 81, 38–48. [Google Scholar] [CrossRef]
- Ju, K.J. Changes in fish fauna and evaluation of migratory fishes before and after the construction of the Nakdong River Estuary weir. Nakdong River Res. 2005, 2–14. [Google Scholar]
- Yang, H.J.; Kim, K.H.; Kum, J.D. The fish fauna and migration of the fishes in the fish way of the Nakdong River Mouth Dam. J. Ecol. Environ. 2001, 34, 251–258. [Google Scholar]
- O’Brien, T. Overcoming Physical Barriers to Fish Migration. In Proceedings of the Fish Movement and Migration, Australian Society for Fish Biology Workshop Proceedings, Bendigo, Victoria, Australia, 27–29 September 1999; pp. 129–134. [Google Scholar]
- Gillson, J.; Scandol, J.; Suthers, I. Estuarine gillnet fishery catch rates decline during drought in eastern Australia. Fish. Res. 2009, 99, 26–37. [Google Scholar] [CrossRef]
- Hoshikawa, H.; Kuwahara, H.; Tajima, K.; Kawai, T.; Kaneta, T.; Tsuda, F. Characteristics of a pacific herring Clupea pallasii spawning bed off Minedomari, Hokkaido, Japan. Fish. Sci. 2004, 70, 772–779. [Google Scholar] [CrossRef]
- Funamoto, T.; Aoki, I.; Wada, Y. Reproductive characteristics of Japanese anchovy, Engraulis japonicus, in two bays of Japan. Fish. Res. 2004, 70, 71–81. [Google Scholar] [CrossRef]
- Cha, H.K.; Lee, J.B.; Kang, S.K.; Chang, D.S.; Choi, J.H. Reproduction of the jack mackerel, Trachurus japonicus Temminck et Schlegel in the coastal waters around Jeju Island, Korea: Maturation and spawning. J. Korean Soc. Fish. Technol. 2009, 45, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Park, J.M.; Baeck, G.W.; Huh, S.H. Variations in demersal fish assemblage in the southern coast of East Sea, Korea. Korean J. Fish. Aquat. Sci. 2018, 51, 426–434. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | EG | HT | Fr | Es | Fw | Family | Species | EG | HT | Fr | Es | Fw |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acropomatidae | Acropoma japonicum | MJ | P | ● | Hapalogenyidae | Hapalogenys nigripinnis | MJ | P | ● | ||||
| Ammodytidae | Ammodytes personatus | MV | P | ● | Hemitripteridae | Hemitripterus villosus | MJ | B | ● | ||||
| Apogonidae | Apogon lineatus | MJ | P | ● | Lateolabracidae | Lateolabrax japonicus | MV | P | ● | ● | ● | ||
| Bagridae | Tachysurus nitidus | FW | B | ● | ● | Leiognathidae | Equulites rivulatus | MJ | P | ● | |||
| Callionymidae | Callionymus beniteguri | MV | B | ● | ● | Nuchequula nuchalis | MV(J) | P | ● | ||||
| Callionymus curvicornis | MV | B | ● | Liparidae | Liparis tanakae | MV | B | ● | |||||
| Callionymus lunatus | MJ | B | ● | Liparis tessellatus | MJ | B | ● | ||||||
| Callionymus valenciennei | MV | B | ● | Lophiidae | Lophius litulon | MJ | B | ● | |||||
| Repomucenus virgis | MJ | B | ● | ● | Monacanthidae | Rudarius ercodes | MJ | P | ● | ||||
| Carangidae | Trachurus japonicus | MJ | P | ● | Mugilidae | Mugil cephalus | DM | P | ● | ● | ● | ||
| Caranx sexfasciatus | MV | P | ● | Mullidae | Upeneus japonicus | MJ | B | ● | |||||
| Centrarchidae | Lepomis macrochirus | FW | P | ● | ● | Muraenesocidae | Muraenesox cinereus | MV | B | ● | |||
| Micropterus salmoides | FW | P | ● | Paralichthyidae | Paralichthys olivaceus | MJ | B | ● | |||||
| Centrolophidae | Psenopsis anomala | MJ | P | ● | Tarphops oligolepis | MJ | B | ● | |||||
| Clupeidae | Clupea pallasii | MJ | P | ● | Pholidae | Pholis fangi | MV | P | ● | ||||
| Clupeidae juvenile sp. | MJ | P | ● | Pholis nebulosa | MV | P | ● | ● | |||||
| Konosirus punctatus | MJ | P | ● | Platycephalidae | Inegocia japonicus | MV | B | ● | |||||
| Spratelloides gracilis | MJ | P | ● | Platycephalus indicus | MV | B | ● | ||||||
| Congridae | Conger myriaster | MV(J) | B | ● | Pleuronectidae | Kareius bicoloratus | MV | B | ● | ● | |||
| Cynoglossidae | Cynoglossus joyneri | MV | B | ● | Pleuronectes yokohamae | MV(J) | B | ● | ● | ||||
| Paraplagusia japonica | ER | B | ● | Rajidae | Okamejei acutispina | MV | B | ● | |||||
| Cyprinidae | Acheilognathus macropterus | FW | P | ● | Okamejei kenojei | MV | B | ● | |||||
| Carassius auratus | FW | P | ● | Sciaenidae | Johnius belengerii | MJ | P | ● | ● | ||||
| Chanodichthys erythropterus | FW | P | ● | ● | Nibea albiflora | MJ | P | ● | |||||
| Cyprinus carpio | FW | P | ● | Pennahia argentata | MV | P | ● | ||||||
| Hemibarbus labeo | FW | P | ● | ● | Scombridae | Scomber japonicus | MJ | P | ● | ||||
| Hemiculter eigenmanni | FW | P | ● | Sillaginidae | Sillago japonica | MV | P | ● | |||||
| Microphysogobio jeoni | FW | P | ● | Siluridae | Silurus asotus | FW | B | ● | ● | ||||
| Opsariichthys uncirostris | FW | P | ● | ● | Sparidae | Acanthopagrus schlegelii | MJ | P | ● | ● | |||
| Pseudogobio esocinus | FW | B | ● | Sphyraenidae | Sphyraena pinguis | MV | P | ● | |||||
| Squalidus gracilis | FW | P | ● | Stromateidae | Pampus argenteus | MV | P | ● | |||||
| Engraulidae | Coilia nasus | DM | P | ● | ● | Tetraodontidae | Lagocephalus wheeleri | MJ | P | ● | |||
| Engraulis japonicus | MJ | P | ● | Takifugu niphobles | ER | P | ● | ||||||
| Thryssa adelae | MV | P | ● | Trichiuridae | Trichiurus lepturus | MJ | P | ● | |||||
| Thryssa hamiltonii | MV | P | ● | Triglidae | Chelidonichthys spinosus | MJ | B | ● | |||||
| Thryssa kammalensis | MV | P | ● | Lepidotrigla guentheri | MJ | B | ● | ||||||
| Gobiidae | Acanthogobius flavimanus | ER | B | ● | ● | Triglidae sp. | MJ | B | ● | ||||
| Favonigobius gymnauchen | ER | B | ● | ● | Zeidae | Zeus faber | MV(J) | P | ● | ||||
| Tridentiger brevispinis | ER | B | ● | ● | Zoarcidae | Zoarces gillii | MV(J) | B | ● | ● |
| Source | df | Species Richness | Abundance | Diversity | |||
|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | ||
| Site | 1 | 9.056 | 0.004 | 7.775 | 0.007 | 7.840 | 0.007 |
| Season | 1 | 1.815 | 0.184 | 4.000 | 0.051 | 0.348 | 0.558 |
| Site × Season | 1 | 8.262 | 0.006 | 0.823 | 0.369 | 6.093 | 0.017 |
| Residual | 53 | ||||||
| Source | df | MS | Pseudo-F | P | COV |
|---|---|---|---|---|---|
| Si | 1 | 70860.0 | 30.243 | 0.007 | 50.884 |
| Se | 1 | 7596.3 | 6.333 | 0.025 | 15.548 |
| St (Si) | 3 | 2353.8 | 1.227 | 0.131 | 6.207 |
| Si × Se | 1 | 7611.4 | 6.346 | 0.023 | 22.014 |
| St (Si) × Se | 3 | 1181.2 | 0.616 | 0.988 | −11.419 |
| Residuals | 47 | 1918.3 | 43.798 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.M.; Riedel, R.; Ju, H.H.; Choi, H.C. Fish Assemblage Structure Comparison between Freshwater and Estuarine Habitats in the Lower Nakdong River, South Korea. J. Mar. Sci. Eng. 2020, 8, 496. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse8070496
Park JM, Riedel R, Ju HH, Choi HC. Fish Assemblage Structure Comparison between Freshwater and Estuarine Habitats in the Lower Nakdong River, South Korea. Journal of Marine Science and Engineering. 2020; 8(7):496. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse8070496
Chicago/Turabian StylePark, Joo Myun, Ralf Riedel, Hyun Hee Ju, and Hee Chan Choi. 2020. "Fish Assemblage Structure Comparison between Freshwater and Estuarine Habitats in the Lower Nakdong River, South Korea" Journal of Marine Science and Engineering 8, no. 7: 496. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse8070496