Gold Clusters Attenuate Inflammation in Rat Mesangial Cells via Inhibiting the Activation of NF-κB Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Gold Clusters
2.1.1. Preparation of Au25Sv9 Gold Cluster
2.1.2. Preparation of Au29SG27 Gold Clusters
2.2. Characterization of Gold Clusters
2.3. Cell Culture and Treatment
2.4. Cells Counting Kit-8 Assay for Cell Viability
2.5. Measurement of TNF-α, IL-1β, IL-6 and PGE2
2.6. Measurement of NO Release
2.7. Reverse Transcription PCR (RT-PCR) and Real-Time Quantitative PCR (RT-qPCR)
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. Characterization of Synthesized Gold Clusters
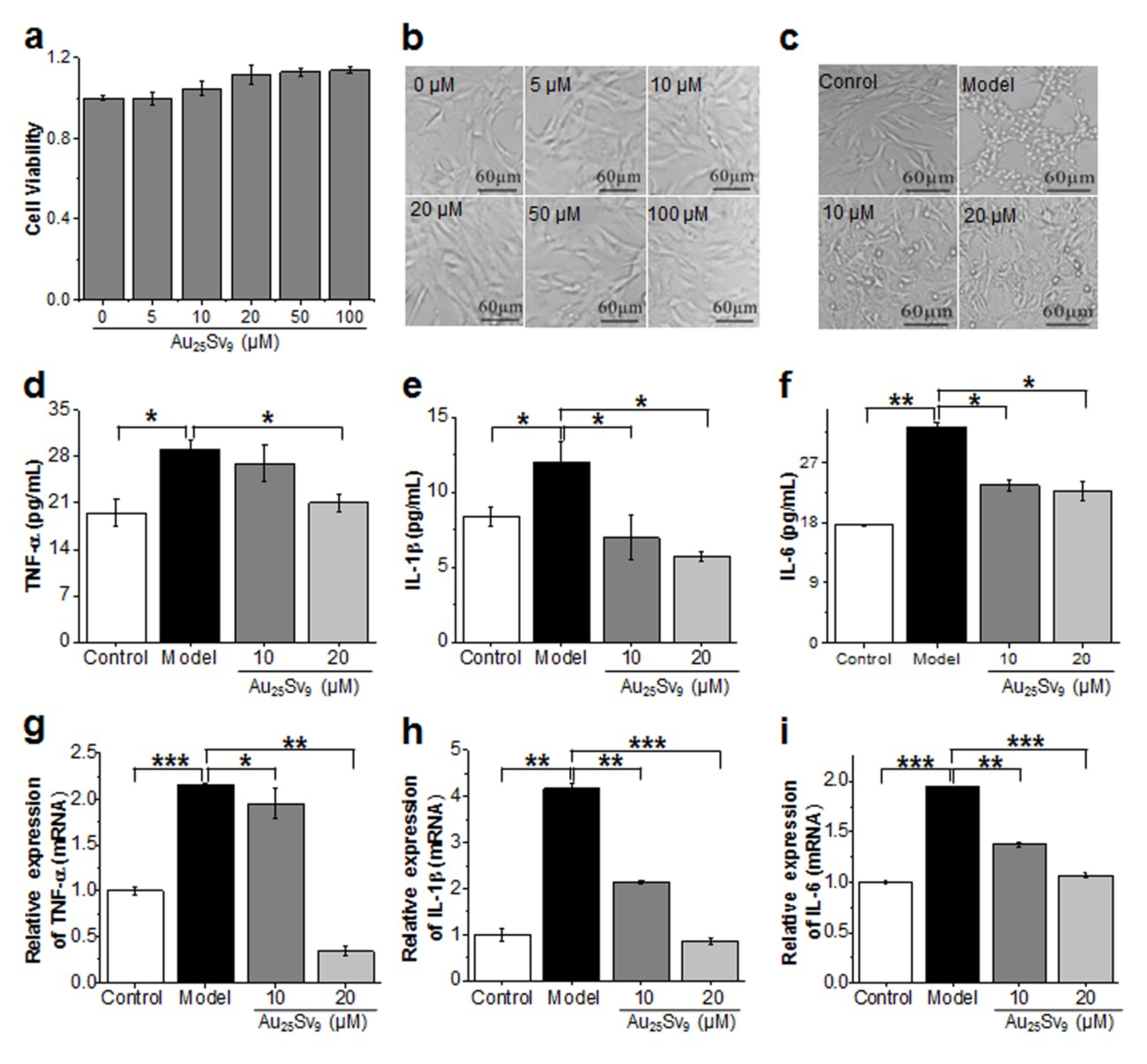
3.2. Effects of Au25Sv9 on HBZY-1 cell Viability and LPS-induced Inflammatory Cytokines Expression
3.3. Au25Sv9 Inhibited COX-2 and iNOS Expression in LPS-Stimulated HBZY-1 Cells
3.4. Au25Sv9 Inhibits Activation of NF-κB Signaling Pathway in LPS-Stimulated HBZY-1 Cells
3.5. Effects of Au29SG27 Gold Clusters on LPS-Stimulated HBZY-1 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fry, D. Sepsis, systemic inflammatory response, and multiple organ dysfunction: the mystery continues. Am. Surg. 2012, 78, 1–8. [Google Scholar] [PubMed]
- Buras, J.A.; Holzmann, B.; Sitkovsky, M. Animal Models of sepsis: setting the stage. Nat. Rev. Drug Discov. 2005, 4, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Dombrovskiy, V.Y.; Martin, A.A.; Sunderram, J.; Paz, H.L. Rapid increase in hospitalization and mortality rates for severe sepsis in the United States: A trend analysis from 1993 to 2003. Crit. Care Med. 2007, 35, 1244–1250. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in sepsis: a novel understanding of the disorder and a new therapeutic approach. Lancet Infect. Dis. 2013, 13, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, G.M.; Zamboni, D.S.; Câmara, N.O.S. The role of innate immunity in septic acute kidney injuries. Shock 2010, 34, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Bagshaw, S.M.; George, C.; Bellomo, R. Early acute kidney injury and sepsis: a multicentre evaluation. Crit. Care 2008, 12, R47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waikar, S.S.; Bonventre, J.V. Creatinine kinetics and the definition of acute kidney injury. J. Am. Soc. Nephrol. 2009, 20, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Bagshaw, S.M.; Uchino, S.; Bellomo, R.; Morimatsu, H.; Morgera, S.; Schetz, M.; Tan, I.; Bouman, C.; Macedo, E.; Gibney, R.N.; et al. Septic Acute Kidney Injury in Critically Ill Patients: Clinical Characteristics and Outcomes. Clin. J. Am. Soc. Nephrol. 2007, 2, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Mårtensson, J.; Bellomo, R. Sepsis-Induced Acute Kidney Injury. Crit. Care Clin. 2015, 31, 649–660. [Google Scholar] [CrossRef]
- Zhao, H.; Zheng, Q.; Hu, X.; Shen, H.; Li, F. Betulin attenuates kidney injury in septic rats through inhibiting TLR4/NF-κB signaling pathway. Life Sci. 2016, 144, 185–193. [Google Scholar] [CrossRef]
- Luo, Z.; Zheng, K.; Xie, J. Engineering ultrasmall water-soluble gold and silver nanoclusters for biomedical applications. Chem. Commun. 2014, 50, 5143–5155. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, Y.; Zhao, L.; Liu, R.; Gao, X. Peptide Protected Gold Clusters: Chemical Synthesis and Biomedical Applications. Nanoscale 2016, 8, 12095–12104. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yuan, Q.; Cai, P.; Gao, L.; Zhao, L.; Liu, M.; Yao, Y.; Chai, Z.; Gao, F. Au Clusters Treat Rheumatoid Arthritis with Uniquely Reversing Cartilage/Bone Destruction. Adv. Sci. 2019, 6, 1801671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Wang, Y.; Yuan, Q.; An, D.; Li, J.; Gao, X. The Au clusters induce tumor cell apoptosis via specifically targeting thioredoxin reductase 1 (TrxR1) and suppressing its activity. Chem. Commun. 2014, 50, 10687. [Google Scholar] [CrossRef]
- Yuan, Q.; Gao, F.; Yao, Y.; Cai, P.; Zhang, X.; Yuan, J.; Hou, K.; Gao, L.; Ren, X.; Gao, X. Gold clusters prevent inflammation-induced bone erosion through inhibiting the activation of NF-κB pathway. Theranostics 2019, 9, 1825–1836. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Yao, Y.; Zhang, X.; Yuan, J.; Sun, B.; Gao, X. The Gold Nanocluster Protects Neurons Directly or via Inhibiting Cytotoxic Secretions of Microglia Cell. J. Nanosci. Nanotechnol. 2019, 19, 1986–1995. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhao, Y.; Cai, P.; He, Z.; Gao, F.; Zhang, J.; Gao, X. Dose-Dependent Efficacy of Gold Clusters on Rheumatoid Arthritis Therapy. ACS Omega 2019, 4, 14092–14099. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhang, W.; Li, L.; Zheng, F.; Shao, X.; Zhou, J.; Li, H. Inhibitory effects of honokiol on lipopolysaccharide-induced cellular responses and signaling events in human renal mesangial cells. Eur. J. Pharmacol. 2011, 654, 117–121. [Google Scholar] [CrossRef]
- Li, G.; Fu, J.; Zhao, Y.; Ji, K.; Luan, T.; Zang, B. Alpha-lipoic acid exerts anti-inflammatory effects on lipopolysaccharide-stimulated rat mesangial cells via inhibition of nuclear factor kappa B (NF-κB) signaling pathway. Inflammation 2015, 38, 510–519. [Google Scholar] [CrossRef]
- Pfeilschifter, J. Nitric oxide triggers the expression of proinflammatory and protective gene products in mesangial cells and the inflamed glomerulus. Nephrol. Dial. Transplant. 2002, 17, 347–348. [Google Scholar] [CrossRef] [Green Version]
- Dinchuk, J.E.; Car, B.D.; Focht, R.J.; Johnston, J.J.; Jaffee, B.D.; Covington, M.B.; Contel, N.R.; Eng, V.M.; Collins, R.J.; Czerniak, P.M.; et al. Renal abnormalities and an altered inflammatory response in mice lacking cyclooxygenase II. Nature 1995, 378, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E. Nuclear factor—κB and its role in sepsis-associated organ failure. J. Infect. dis. 2003, 187, S364–S369. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.F.; Malik, A.B. Nf- κB activation as a pathological mechanism of septic shock and inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L622–L645. [Google Scholar] [CrossRef]
- Li, Y.M.; Zhang, J.; Su, L.J.; Kellum, J.A.; Peng, Z.Y. Downregulation of TIMP2 attenuates sepsis-induced AKI through the NF-κB pathway. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Mao, L.; Yang, L.; Zou, J.; Liu, K.; Liu, M.; Zhang, H.; Xiao, X.; Wang, K. Resveratrol protects against early polymicrobial sepsis-induced acute kidney injury through inhibiting endoplasmic reticulum stress-activated NF-κB pathway. Oncotarget 2017, 8, 36449–36461. [Google Scholar]
- Bellomo, R.; Kellum, J.A.; Ronco, C.; Wald, R.; Mårtensson, J.; Maiden, M.; Bagshaw, S.M.; Glassford, N.; Lankadeva, Y.; Vaara, S.T.; et al. Acute kidney injury in sepsis. Intensiv. Care Med. 2017, 43, 816–828. [Google Scholar] [CrossRef] [Green Version]
- Benes, J.; Chvojka, J.; Sykora, R.; Radej, J.; Krouzecky, A.; Novak, I.; Matejovic, M. Searching for mechanisms that matter in early septic acute kidney injury: an experimental study. Crit. Care 2011, 15, R256. [Google Scholar] [CrossRef] [Green Version]
- Younes-Ibrahim, M. Mesangial cells: Renal function protagonists or coadjuvants? J. Bras. Nefrol. Orgao Soc. Bras. Lat. Am. Nefrol. 2013, 35, 248–249. [Google Scholar] [CrossRef]
- Esposito, V.; Grosjean, F.; Tan, J.; Huang, L.; Zhu, L.; Chen, J.; Xiong, H.; Striker, G.E.; Zheng, F. CHOP deficiency results in elevated lipopolysaccharide-induced inflammation and kidney injury. Am. J. Physiol. Physiol. 2012, 304, F440–F450. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Bansal, S.; Falk, S.; Ljubanovic, D.; Schrier, R. Ghrelin protects mice against endotoxemia-induced acute kidney injury. Am. J. Physiol. Physiol. 2009, 297, F1032–F1037. [Google Scholar] [CrossRef] [Green Version]
- Heemskerk, S.; Masereeuw, R.; Russel, F.; Pickkers, P. Selective iNOS inhibition for the treatment of sepsis-induced acute kidney injury. Nat. Rev. Nephrol. 2009, 5, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.B.; Sanchez-Niño, M.D.; Ramos, A.M.; Moreno, J.A.; Santamaria, B.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A. NF-κB in renal inflammation. J. Am. Soc. Nephrol. 2010, 21, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Y.Y.; Hou, C.C.; Wang, W.; Huang, X.R.; Lan, H.Y. Blockade of NFκB activation and renal inflammation by ultrasound-mediated gene transfer of Smad7 in rat remnant kidney. Kidney Int. 2005, 67, S83–S91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-D.; Wu, D.; Shen, X.; Liu, P.-X.; Fan, F.-Y. In vivo renal clearance, biodistribution, toxicity of gold nanoclusters. Biomaterials 2012, 33, 4628–4638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-D.; Chen, J.; Luo, Z.; Wu, D.; Shen, X.; Song, S.-S.; Sun, Y.; Liu, P.-X.; Zhao, J.; Huo, S.; et al. Radiosensitizers: Enhanced Tumor Accumulation of Sub-2 nm Gold Nanoclusters for Cancer Radiation Therapy. Adv. Heal. Mater. 2014, 3, 152. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhai, J.; Gao, F.; Liu, R.; Gao, L.; Zhao, Y.; Chai, Z.; Gao, X. Label-Free Au Cluster Used for in Vivo 2D and 3D Computed Tomography of Murine Kidneys. Anal. Chem. 2014, 87, 343–345. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| COX-2 | ACTTACAGGAGAGAAGGAAATGGCT | GTATTGAGGAGAACAGATGGGAT |
| iNOS | AGCCGCACCACCCTCCTTGT | GCCTCTTGTCTTTGACCCAGTAG |
| TNF-α | CTCAAGCCCTGGTATGAGCC | GGCTGGGTAGAGAACGGATG |
| IL-1β | GGAAGGCAGTGTCACTCATTGTG | GGTCCTCATCCTGGAAGCTCC |
| IL-6 | TCCTACCCCAACTTCCAATGC | TAGCACACTAGGTTTGCCGAG |
| GAPDH | AGACAGCCGCATCTTCTTGT | CTTGCCGTGGGTAGAGTCAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, J.; Hou, K.; Yao, Y.; Du, Z.; Lu, C.; Yuan, Q.; Gao, X. Gold Clusters Attenuate Inflammation in Rat Mesangial Cells via Inhibiting the Activation of NF-κB Pathway. Nanomaterials 2020, 10, 712. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10040712
Yuan J, Hou K, Yao Y, Du Z, Lu C, Yuan Q, Gao X. Gold Clusters Attenuate Inflammation in Rat Mesangial Cells via Inhibiting the Activation of NF-κB Pathway. Nanomaterials. 2020; 10(4):712. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10040712
Chicago/Turabian StyleYuan, Jinling, Kaixiao Hou, Yawen Yao, Zhongying Du, Cao Lu, Qing Yuan, and Xueyun Gao. 2020. "Gold Clusters Attenuate Inflammation in Rat Mesangial Cells via Inhibiting the Activation of NF-κB Pathway" Nanomaterials 10, no. 4: 712. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10040712