How to Find the Right RNA-Sensing CRISPR-Cas System for an In Vitro Application


1
Computational Bioscience Research Center (CBRC), Biological and Environmental Science and Engineering (BESE), King Abdullah University of Science and Technology (KAUST), Thuwal 23955-6900, Saudi Arabia
2
Centre de Biologie Structurale (CBS), INSERM, CNRS, Université de Montpellier, F-34090 Montpellier, France
*
Authors to whom correspondence should be addressed.
Biosensors 2022, 12(2), 53; https://0-doi-org.brum.beds.ac.uk/10.3390/bios12020053
Submission received: 29 December 2021
/
Revised: 17 January 2022
/
Accepted: 18 January 2022
/
Published: 19 January 2022
(This article belongs to the Special Issue Application of CRISPR Cas Systems for Biosensing)
Abstract
:CRISPR-Cas systems have a great and still largely untapped potential for in vitro applications, in particular, for RNA biosensing. However, there is currently no systematic guide on selecting the most appropriate RNA-targeting CRISPR-Cas system for a given application among thousands of potential candidates. We provide an overview of the currently described Cas effector systems and review existing Cas-based RNA detection methods. We then propose a set of systematic selection criteria for selecting CRISPR-Cas candidates for new applications. Using this approach, we identify four candidates for in vitro RNA.
1. Introduction
The natural CRISPR-Cas system comprises two main components. The CRISPR, acronym stands for “clustered regularly interspaced short palindromic repeats” and describes the first component—an array of short DNA fragments. These DNA fragments are snippets of bacteriophage genome sequences that bacteria and archaea retain from past infections. The RNA transcripts of these sequences are then recruited by the other component, the CRISPR-associated (Cas) proteins, where the transcripts serve as guide RNA (gRNA) to recognize subsequent infections by similar viruses. Upon forming the ribonucleoprotein (RNP) complex, Cas endonuclease activity specifically destroys the DNA or RNA of the invading virus [1,2,3,4,5,6,7,8,9,10]. This adaptive immune system of prokaryotes has now been repurposed into various programmable DNA editing tools [11,12,13,14].
Offering a large range of clinical, research, and diagnostic applications, CRISPR-Cas systems have rapidly become one of the most widely used molecular biology tools. While DNA editing remains the most prominent area of CRISPR applications, bioengineers have increasingly also turned their attention to more recently discovered Cas proteins capable of targeting and cleaving RNA instead of DNA [15,16]. Such RNA-targeting CRISPR systems are now being developed into impermanent, dose-dependent in vivo genetic therapies [17,18,19,20] or are used to identify RNA–protein interactions in living cells [21,22]. However, right after the discovery of the first RNA-targeting Cas ortholog (now named LshCas13a) by Abudayyeh et al. [15], East-Seletsky et al. already demonstrated using these novel ribonucleases for detecting specific RNA sequences [16]. Consequently, in vitro RNA biosensing has become an important new application for CRISPR-Cas systems [23,24].
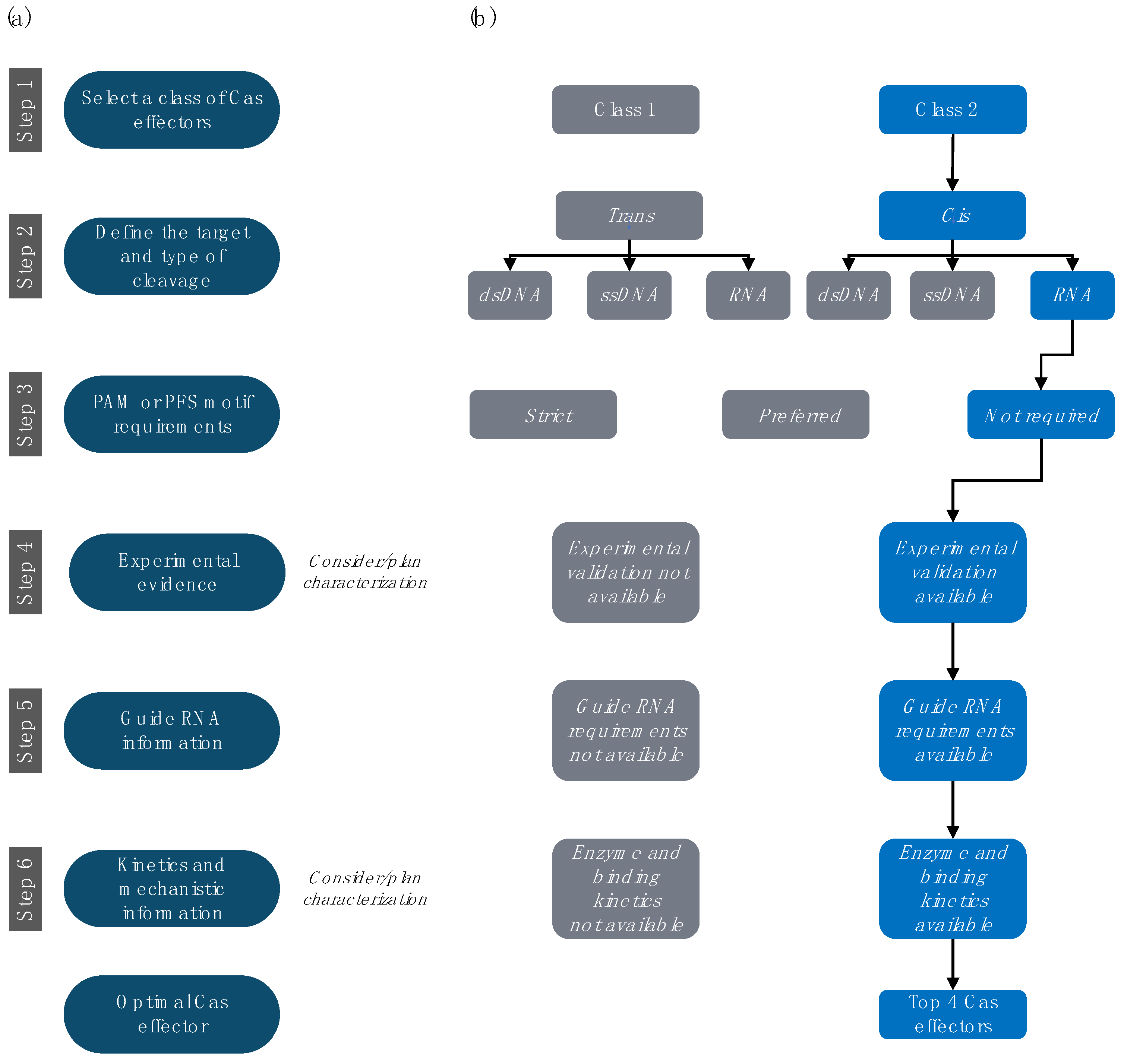
The requirements substantially differ between cellular and in vitro applications, with the latter emphasizing biochemical factors, such as Cas expression yields, solubility and stability in buffers, and binding affinities or catalytic efficiency. As most CRISPR development still focuses on cellular applications and DNA-targeting Cas proteins, such data are not widely available for RNA-targeting CRISPR systems. Many in vitro Cas features that are critical for designing new applications, such as their mechanism of action, kinetics, and cleavage specificity, remain unknown or poorly documented. Therefore, selecting and characterizing the appropriate CRISPR-Cas ortholog for a given purpose becomes an essential but challenging task. Based on their evolution and architecture, Makarova et al. classified CRISPR-Cas systems into two classes, six types, and thirty-three subtypes [25]. We propose a step-by-step guide for identifying and prioritizing Cas effectors, specifically for in vitro applications (Figure 1). We concentrate on the underexplored RNA-targeting CRISPR systems and their use for in vitro RNA biosensors. Nevertheless, the selection criteria, tools, and methods we discuss equally apply to most in vitro uses of CRISPR-Cas systems.
2. Current CRISPR-Cas–Based RNA Detection Methods
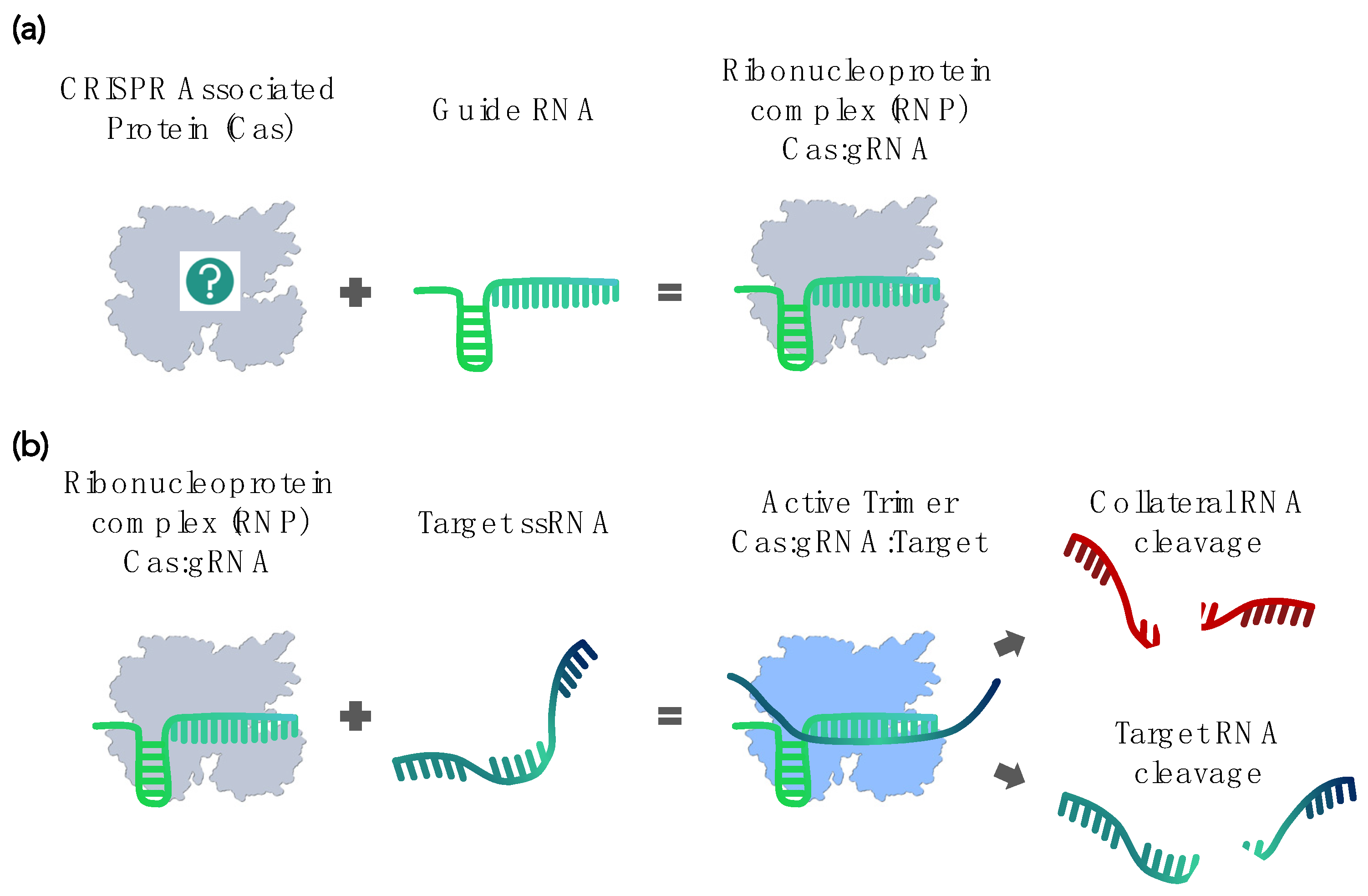
In 2016, Abudayyeh et al. demonstrated that the Cas ortholog LshCas13a specifically targets mRNA in vivo but also displays an in vitro “collateral” cleavage activity, meaning that it nonspecifically cleaves nearby RNA in trans [15] (Figure 2). This nuclease activity in trans nevertheless strictly depends on the specific binding of the RNP complex to its actual RNA target [15]. It is fast and exhibits multiple turnovers, unlike the target cleavage, which is normally a single turnover [26]. That same year, East-Seletsky et al. used the LbuCas13a activity for the Cas-based RNA sensing of endogenous ß-actin mRNA from HeLa cell RNA extracts [16]. The collateral cleavage of single-stranded reporter RNA (ssRNA) molecules amplified target recognition events by up to 10,000-fold and translated them into a sensitive fluorescence readout. Many of the subsequently developed Cas-based nucleic acid detection methods still use variations of this same principle.
Currently, the most prominent CRISPR-based detection assays are SHERLOCK (specific high-sensitivity enzymatic reporter unlocking) and DETECTR (DNA endonuclease-targeted CRISPR trans reporter) [23,24]. Both use Cas orthologs (LwaCas13a and LbCas12a, respectively) to detect RNA (and DNA) with high specificity. Both rely on additional sample processing steps, such as (reverse) transcription reactions (i.e., RT-RPA, T7 in vitro transcription) and enzymatic DNA amplification to reach attomolar (aM) sensitivity. For SHERLOCK, the target RNA is reverse-transcribed before amplification and subsequently transcribed back into RNA for Cas binding. The Cas ortholog used by the DETECTR system can only detect DNA. Therefore, the target RNA is first reverse-transcribed and then amplified as DNA, which is recognized by the Cas [23,24]. Both systems collaterally cleave a reporter molecule upon binding the Cas effector to its target.
The SHERLOCK, DETECTR, and similar Cas-based nucleic acid detection systems employ various approaches for converting reporter cleavage into a readout. The most common are fluorescence or colorimetry in solution or the capture of dyes on lateral flow strips. Fazouni et al. (2020) adapted LbuCas13a to the detection of the severe acute respiratory syndrome coronavirus 2 (SARS-Cov2) and used the camera of a mobile phone to sense and quantify RNA cleavage [27]. Another system, iSCAN (in vitro Specific CRISPR-based Assay for Nucleic acids detection), combines RT-LAMP (reverse transcription loop-mediated isothermal amplification) with LbCas12a, AacCas12b, or AapCas12b to detect SARS-Cov2 through a colorimetric readout on lateral flow cells [28]. SHERLOCKv2 multiplexes different Cas orthologs (PsmCas13b, AsCas12a, and CcaCas13b). Each Cas ortholog has a particular nucleotide-cleavage preference that facilitates the orthogonal detection of several targets in a one-pot reaction [29]. Similarly, HOLMES (one-hour low-cost multipurpose highly efficient system) and HOLMESv2 use LbCas12a and AacCas12b orthologs, respectively, to detect DNA and RNA with aM sensitivity [30,31].
Cas effectors have also been actively developed into more field-deployable platforms. For example, a CRISPR/Cas13a powered portable electrochemiluminescence chip (called PECL-CRISPR) detects miRNA by combining the sensitivity of LbuCas13a with the low background and large dynamic range of chemiluminescence and electrochemical detection. This system reports the detection of 1 femtomolar (fM) of miR-17 on a paper-based bipolar electrode [32]. A strategy inspired by digital PCR, SATORI (for CRISPR-based amplification-free digital RNA detection) combines LwaCas13a with the parallel observation of multiple reactions in femtoliter microchamber arrays and could detect 10 fM of ssRNA in less than 5 min [33].
In conclusion, different studies have combined various Cas orthologs with different signal amplification and readout strategies. In many cases, the rapid collateral cleavage activity of Cas13 systems is used to enzymatically amplify the response by unquenching the fluorescent signal from a reporter molecule (SHERLOCK, DETECTR [23,24]). Other methods use external reporting systems, such as electrochemical transistors, lateral flow paper strips, colorimetry, or electrochemiluminescence [27,28,32]. The requirements of the desired output system will be an important factor in the selection of a Cas ortholog. However, the reasons for selecting one ortholog over another with similar features are not always clear.
3. Computational Tools for CRISPR Identification
Computational protein prediction from sequences combined with experimental work has resulted in a vast and continuously increasing list of CRISPR-Cas systems. Many online tools help identifying new CRISPRs, spacers, and Cas proteins using sequence analysis. Several Cas system databases have been compiled. For example, Tang et al. constructed the Cas Protein Data Bank that catalogs Cas proteins from bacteria and archaea [34]. CRISPRminer attempts to describe the full biological picture of CRISPR genes, classification, spacers, and targets in several thousand bacterial and archaeal species [35]. CRISPRCasFinder [36] identifies CRISPR arrays, DRs, spacer sequences, and Cas genes along with their type and subtype in user-provided sequences. The associated CRISPRCasdb database catalogs this information for all known bacterial and archaeal genomes [37]. In order to facilitate the selection of RNA-targeting Cas effectors from the large number of theoretical candidates, we propose a step-by-step approach that is described in the following.
4. Step-by-Step Selection and Design of RNA-Targeting CRISPR-Cas Systems
4.1. Step 1. Select the Preferred Class of Cas Effectors
There are two main classes of Cas effectors (Table 1). Class 1 generally relies on multiple Cas subunit proteins to degrade foreign genetic elements (currently with only one exception [38]). Class 2 combines gRNA processing, target recognition, and cleavage activities into a single multidomain protein. Reducing the number of parts also reduces the complexity of engineering. Moreover, the published research on Cas applications generally utilizes Class 2 effectors. Therefore, we do not consider Class 1 CRISPR-Cas systems in our selection. Genomic and metagenomic data analyses identify more than 175,000 Class 2 Cas family members [39]. This number is constantly increasing due to computational predictions, protein screens, and new sequencing data analyses.
4.2. Step 2. Select the Preferred Target and Type of Cleavage
The three main targets for Cas endonucleases are double-stranded DNA (dsDNA), single-stranded DNA (ssDNA), and RNA, and we focus on the latter. The first identified RNA-targeting CRISPR-Cas systems belonged to the Class 1 Type III-B (Pyrococcus furiosus, Cmr) [40]. However, we excluded Class 1 enzymes in Step 1 because of their multiple-effector architectures. Despite possessing a single-effector nuclease, we also excluded the highly diverse Class 2 Type V effectors because their main cleavage target is dsDNA or ssDNA (i.e., Cas12 and Cas14), although some cleave “bystander” ssRNA in a trans/collateral way (i.e., Cas12). Only Cas effectors belonging to Class 2 Types II and VI are likely to recognize and cleave ssRNA and, therefore, were further considered.
While we did not specifically consider it in our example, collateral cleavage activity may often become an additional selection criterion. Efficient collateral RNA cleavage is a hallmark of Type VI (Cas13) Cas proteins but, as mentioned, can also be found in some Type V Cas orthologs [41]. Evidently, cleavage of “bystander” RNA may not be desirable for all applications but is often used for signal amplification and readout. Interestingly, certain Cas orthologs show a nucleotide or sequence bias also for trans-cleaved substrates. Such a property may add a secondary degree of programmability, which could improve a device’s fidelity [29].
4.3. Step 3. Investigate PAM or PFS Motif Requirements
Protospacer adjacent motifs (PAMs) and protospacer flanking sites (PFSs) are both short sequences (~1–6 nt) immediately following the target sequence (DNA or RNA, respectively). Both PAMs and PFSs are “hard-wired” into the enzyme’s specificity, and, while they need to be present in the target, they are not encoded in the gRNA or its encoding CRISPR array. Thus, they protect the host CRISPR locus from cleavage by the host’s own Cas systems. These elements are indispensable for target recognition in many Cas orthologs (i.e., SpCas9, AacCas12, and BthCas12). Consequently, on the one hand, the PAM or PFS sequence must be present in the target and cannot be reprogrammed. This constraint reduces the number of possible target sites on a given sequence. On the other hand, PAM or PFS often improve the target specificity of the system. Some Cas orthologs can target ssDNA independently of a PAM but require a PAM for dsDNA targets [42]. For ssDNA and RNA targeting, attempts have been made to circumvent the PAM constraint using protein engineering [43].
The PFS sequences are localized in the 3′ of target ssRNA and affect the efficacy of some obligatory ssRNA-targeting Cas proteins of Type VI but are not a strict requirement for RNA targeting in mammalian cells [17,44]. However, many RNA-targeting Cas effectors, such as LwaCas13a (Type VI), EsCas13d (Type VI), and SauCas9 (Type II), do not require any PFS sequences for their in vitro activity [45]. We opted to prioritize the Cas proteins from Type II and VI without any requirement for PAM/PFS (respectively) to maximize our flexibility in selecting targets. However, dsDNA-specific Cas systems with PAM requirements may be “tricked” into the recognition and cleavage of ssDNA or RNA by the design of so-called PAMers [26,46,47]. PAMers are synthetic oligonucleotides that hybridize with a single-stranded target RNA (or DNA) and create a short double-stranded segment, including the enzyme’s PAM. There is also emerging evidence for the successful use of PAMers for RNA-targeting Cas proteins (i.e., SaCas9, SpyCas9, NmeCas9, and others.) [46,48].
4.4. Step 4. Prioritize Effectors with Experimental Characterization
Even after limiting the search to RNA-targeting Class 2 Type II and VI without PSF requirements, the number of available orthologs is still very large. For instance, CRISPRminer reports about 4300 entries of Cas proteins from Class 2 Types II and VI [35]. Given that the success rate in bioengineering is dramatically increased if a certain biological part has already been characterized experimentally, we prioritize Cas effectors for which such data are available.
As we focus on in vitro applications, the primary criterion is the availability of experimental in vitro evidence for RNA target cleavage or binding. A widely used method is the electrophoretic mobility shift assay (EMSA) [57], in which the binding of the Cas complex delays the migration of the target RNA/DNA through a gel. Alternative methods include filter binding assays [26,58] or biochemical cleavage assays [26,30,42]. In addition, fluorescence-based assays can provide proof of protein functionality, the mechanism of action, and target specificity [58,59]. Quantitative data on binding affinity are scarce but much preferred over qualitative results. These are often derived from densitometry quantification of EMSA bands (i.e., for SauCas9 [26]) or, more accurately, by biophysical methods, such as isothermal titration calorimetry, microscale thermophoresis, or surface plasmon resonance [60].
The documentation of protein expression and purification protocols is also important for in vitro applications. Studies rarely report their quantitative protein yield. Where data are available, we used them to prioritize Cas effectors that required a smaller number of purification steps and for which high solubility and detailed purification protocols are reported (i.e., LwaCas13a) [42]. We excluded Cas effectors without purification reports or those lacking results in controlled environments. In this case, this step eliminated PguCas13b, PspCas13b, and RfxCas13d. We also excluded some Cas orthologs, such as CasRx, which perform well in cell cultures (mammalian, insect, plant, or bacterial) but whose activity has not been characterized in vitro.
4.5. Step 5. Investigate gRNA Information
In native CRISPR-Cas9 systems, the gRNA consists of two base-paired RNA molecules: (i) a CRISPR RNA (crRNA) that encodes a sequence (spacer) that is complementary to the target and (ii) a trans-activating CRISPR RNA (tracrRNA) that mediates the interaction of the Cas effector protein with the crRNA [61]. A major initial step for enabling the use of CRISPR-Cas as a programmable DNA editing tool was the design of a single gRNA (sgRNA) that directed the Cas protein (and its associated enzymatic activity) to a chosen DNA target as defined by the complementary gRNA sequence [12]. Some Class 2 effectors, such as Cas13, naturally use only a single crRNA molecule. This natural gRNA contains a direct repeat (DR) stem-loop that mediates the interaction with the Cas protein and a spacer sequence that determines the target selectivity [62]. The terms gRNA, sgRNA, crRNA, and spacer are sometimes used interchangeably in the literature.
The exact gRNA design rules for a given Cas ortholog must be known before adapting the system to any application. At a minimum, this includes the spacer length requirements and the sequence and structure details of the DR. More detailed information on spacer specificity is preferable. For example, spacers of Cas9 orthologs usually feature a seed region (usually 8–10 nt at the 3′ end of the spacer) with low or no mismatch tolerance [12]. This seed region participates in the first target interrogation step [12,63]. For generally well-characterized Cas systems, we find quantitative data on the mismatch tolerance or sensitivity across the full spacer sequence (a particularly good example is LbuCas13a [58]). Target recognition may depend on divalent ions, such as Mg2+ and Mn2+, which is an important consideration for the experimental design (i.e., EsCas13a [64]). Even if the gRNA architecture is well understood, the selection of the actual target-specific spacer sequence may still substantially affect the functioning of the detection system. Several bioinformatics tools aim to help select spacer sequences for Cas9 systems targeting DNA [65]. Tools for non-Cas9 systems have also been developed and often consider the secondary structure of both the gRNA and target RNA [66,67,68]. For example, Wessels et al. created an online tool that predicts Cas13d gRNAs for all protein-coding transcripts in the human genome (cas13design.nygenome.org) [62]. If not already considered by the primary tool, secondary structure predictors, such as RNAfold [69] and RNAxs [70], may be employed as a secondary filter. Nevertheless, a strong consensus exists in the literature that at least two to five spacer sequences must be tested experimentally to determine the most efficient guide for a particular target and working conditions (with other variables including the buffer, temperature, or target binding kinetics) [27].
In the example analysis, we prioritized Cas effectors with detailed gRNA characterization data and reduced mismatch tolerance within and outside the seed region. Examples of such systems are LbuCas13a and LwaCas13a [15,58]. Coincidentally, both of these Cas orthologs can process or maturate their crRNA from a tandem array. Depending on the application, this activity could offer an opportunity for efficient multiplexing. However, this feature was not considered in the prioritization.
4.6. Step 6. Review Available Kinetics and Mechanistic Information
The Cas proteins have markedly different sizes (currently ranging from ~800 to 1700 aa) [71,72] and domain compositions (i.e., RucV versus HEPN-nuclease motifs). The smaller size of single-effector proteins facilitates their transfection into cells, both directly using protein transfection methods or through viral vectors (i.e., Cas13bt, d, e, and f [71,72,73]). Conversely, larger Cas proteins may have additional (beneficial) functions, such as the RNase modules that process pre-crRNA. Type II and VI effectors usually adopt a bi-lobed architecture (the recognition (REC) lobe and nuclease (NUC) lobe) typically connected by an arginine-rich bridge helix and linker loop [26].
For most Cas effectors, the mechanism by which the gRNA interacts with the Cas protein (duplex complex) and how this binary complex interacts with the target (triple complex) is still not completely understood. Structural analyses have indicated that Cas catalytic activity and specificity often depend on large conformational changes when moving from duplex to triple complexes [41,74]. In general, three-dimensional (3D) structural information is an important tool for informed engineering and testifies to the successful recombinant expression and purification of a protein. Therefore, we prioritized the Cas effectors for which structures were published. Additionally, we kept LwaCas13a, where a detailed record of biochemical characterization compensates for the lack of a structure.
Where quantitative comparisons were available, we preferred Cas orthologs with the higher catalytic activity (e.g., LwaCas13a rather than LshCas13a) [44]. Although relevant, we found that the affinity of the interaction between the Cas effector and its gRNA is generally lacking. However, apparent dissociation constants (Kds) are sometimes reported for the binding of the Cas–gRNA complex to its targets [26]. The availability of these data was the final criterion that promoted SauCas9 and LwaCas13a to the list of top notable candidates. Moreover, specific applications or experiments (e.g., control reactions) may require catalytically dead Cas proteins. We prioritized Cas effectors for which (i) target binding had been reported with both active and catalytically dead versions and (ii) the full description and (if possible) structural analysis of these inactivating mutations were available [17].
5. Detailed Profiles of Four Cas Effectors for In Vitro RNA Detection
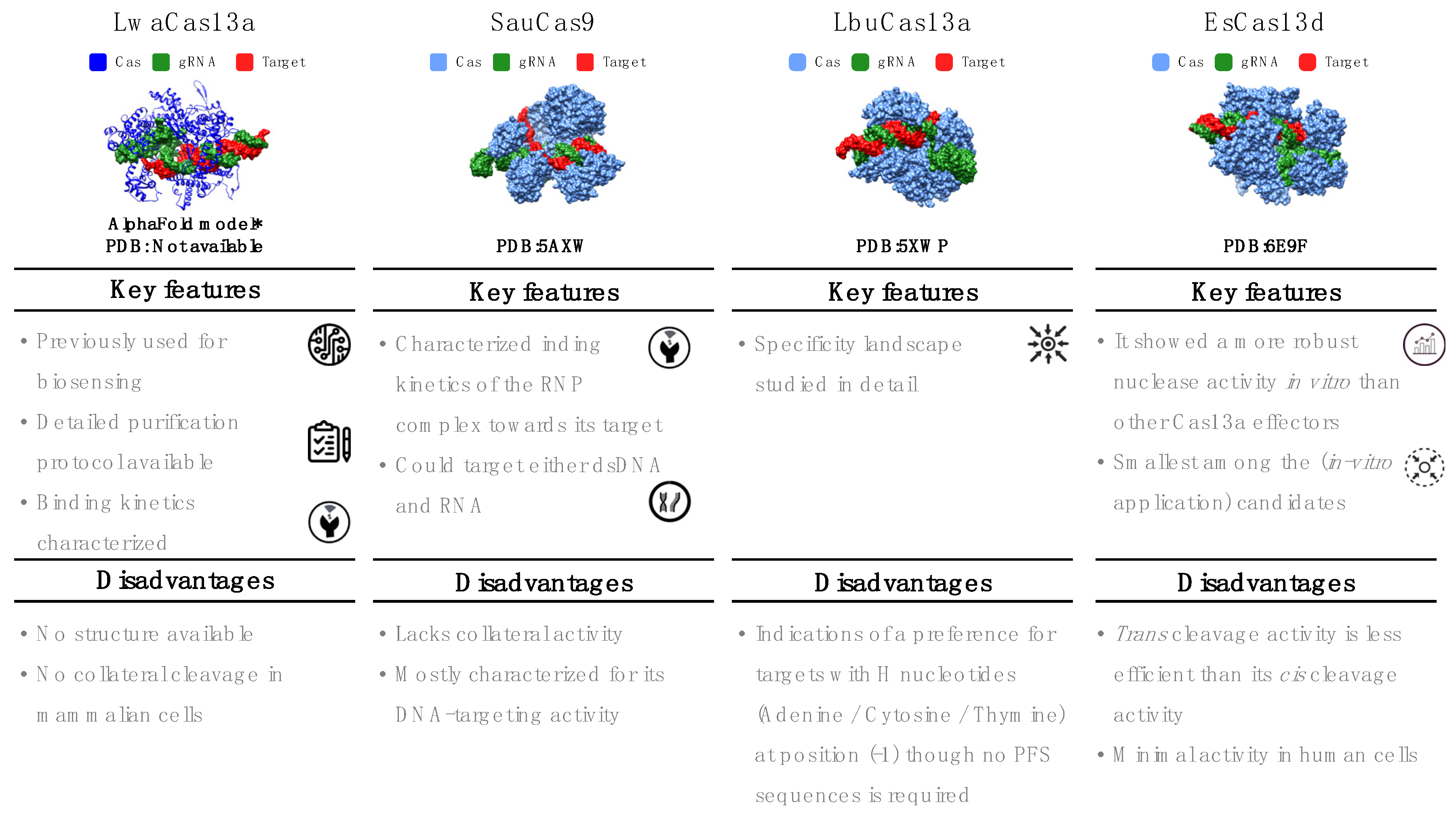
The six-step decision workflow led us to select four Cas systems that we consider top candidates for developing in vitro RNA detection systems (Figure 3). As we highlighted above, some of the selection criteria were strict constraints, such as the restriction to Class 2 enzymes or the availability of gRNA design rules. Other criteria resulted from the current scarcity of experimental data and sometimes had to be weighed against each other. These latter criteria can be reassessed when more experimental data are reported.
5.1. Leptotrichia wadei (LwaCas13a)
This single-effector nuclease is mostly known for its use in the SHERLOCK system. Its 3D structure has not yet been determined, but LwaCas13a has been extensively characterized for several in vitro and in vivo applications. LwaCas13a is an RNA-only targeting nuclease with cis and collateral/trans cleavage in vitro and in bacterial cells [16,42,44]. However, in mammalian cells, no collateral cleavage has been observed [44]. LwaCas13a does not require a PFS sequence. However, like other Cas13 orthologs, Cas:gRNA binding to the target can be hindered by strong secondary structure motifs in the target [16,44]. The catalytically inactive LwaCas13a version maintains binding to the target [44]. Its natural spacer length is 29–30 nt. Although LwaCas13a retains its cleavage activity with spacer lengths as short as 20 nt [44], most studies have used 28 nt spacers. The SHERLOCK assay successfully detects 20 fM of non-amplified ssRNA even after lyophilization and rehydration of LwaCas13a. When combined with nucleic acid amplification, the sensitivity of SHERLOCK reaches 2 aM [24]). LwaCas13a is incapable of cutting sequences with two or more mismatches. Consequently, spacers with one deliberate mismatch in the guide sequence allow the detection of targets with perfect sequence specificity. Single-nucleotide polymorphisms or closely related pathogen strains can be distinguished using this method [29,42]. Besides nucleic acid detection in clinical samples [24,29,75], LwaCas13a has been used to detect plant and food pathogens [76,77].
5.2. Staphylococcus aureus (SauCas9)
The single-effector Cas SauCas9 belongs to Class 2 Type II. With a molecular weight of 124 kDa, SauCas9 is markedly shorter than other Cas9 effectors (typically ~160 kDa) [78,79]. It possesses two nuclease domains (RuvC and HNH) and can cleave both dsDNA and RNA, but not simultaneously [26,78]. The RNA-cleavage activity has a strong preference for nonstructured/ssRNA targets. Unlike other RNA-targeting effectors, SauCas9 does not have collateral ribonuclease activity. It requires a relatively long PAM sequence (5′-NNGRRT-3′) for DNA targeting, but neither PAM nor PFS are required for RNA cleavage [26]. The literature on DNA targeting is extensive [78,80,81,82,83] and includes crystal structures with DNA targets (Protein Data Bank (PDB): 5AXW and 5CZZ) [78]. RNA targeting is less well studied but has been confirmed through in vitro cleavage assays, filter binding, and EMSA experiments [26]. Kinetic data suggest that SauCas9 is a multiple-turnover enzyme for DNA targets but a single-turnover enzyme for RNA targets [26,82]. The dissociation constant for the binding of SauCas9:gRNA to target RNA is about 1.8 nM [26,82]. SauCas9 requires divalent ions for cleavage. Its optimal spacer length is 23 nt, and the gRNA sequence is available [26,82]. A catalytically dead version (dSauCas9) and a split-protein version for DNA or RNA binding and recognition inside cells have been described [78].
5.3. Leptotrichia buccalis (LbuCas13a)
This Cas system is a single-effector protein of about 140 kDa. Similar to other effectors in Class 2 Type VI, it targets ssRNA and possesses both cis and trans cleavage activity in the presence of Mg2+ [16]. LbuCas13a can process its pre-crRNA, but the processing of the crRNA is not required to activate the effector protein [16,41,84,85]. Its specificity landscape has been studied in detail, concluding that this protein can distinguish between fully complementary and mismatched RNA transcripts (even those containing only a single mismatched nucleotide) before activating the RNP complex [58]. LbuCas13a is a sensitive RNA detector with observable reporter cleavage in the presence of only 10 fM of the activator target. Its structure has been determined (PDB:5XWP and 5XWY [84]), revealing conformational changes both after binding to the gRNA and after forming the triple complex with the target [84]. The target RNA must be at least 20 nt in length to activate the Cas effector [84]. The cleavage activity of LbuCas13a has been characterized in cell-free experiments through EMSA, filter binding, fluorescence assays, and fluorescence polarization [16,58]. LbuCas13a does not require a PFS sequence [58]. However, there are indications of a preference for targets with H nucleotides (adenine/cytosine/thymine) at position (-1) [58]. The sequence of the constant region of its gRNA has been published (Table 2) [16,41,58,74,84,85]. The seed region (gRNA nucleotides 9–14 nt) required for target binding is mismatch hypersensitive, whereas mismatches in the HEPN-nuclease switch region (gRNA nucleotides 5–8) can result in an inactive nuclease that remains tightly bound to the target [58].
5.4. Eubacterium siraeum (EsCas13d)
Derived from a gut-resident bacterium, EsCas13d (~105 kDa) was among the two first characterized single-effector Cas of Class 2, Type VI, Subtype d [71,74]. EsCas13d is an ssRNA-only nuclease [71]. It exhibits cis and trans cleavage activity, with trans less efficient than cis [71]. Structures for both apoprotein and binary (Cas:gRNA) or ternary (Cas:gRNA: target) complexes have been determined (Electron Microscopy Data Bank (EMDB): 9015, 9013, and 9014 and PDB: 6E9E and 6E9F [74]). Although EsCas13d exhibited minimal activity in human cells, in vitro experiments demonstrated more robust nuclease activity than Cas13a effectors [71]. EsCas13d does not require a PFS sequence. However, Mg2+ is necessary for target cleavage and can improve the efficiency of crRNA maturation. Additionally, evidence exists that two hydrated Mg2+ ions stabilize the conformation of the DR region [64]. EsCas13d can process its crRNA, a single-molecule guide comprising a DR region (of about 30 nt located, unlike others, at the 5′ end) and a spacer region [74].
Somewhat contradictory reports describe a solvent-exposed seed region, either in position 1–16 nt or 5–21 nt of the spacer, intolerant to two consecutive mismatches [62,74]. EsCas13d is considered a highly specific effector because its activation depends not only on binding the correct target sequence, but is in addition gated by a very profound conformational change [66]. Yan et al. found that an optimal cleavage activity is achieved with a 21–30 nt spacer, and the most commonly reported spacers are 23 nt long [86]. As other Cas proteins, EsCas13d prefers a low secondary structure content in the targeted RNA [66,86]. It has been experimentally characterized using biochemical cleavage assays and filter binding assays for which crRNA and target sequences were reported [66,74]. Both catalytically active and inactive Cas variants expressed well in Escherichia coli [71].
5.5. Other Notable Cas Candidates
Throughout the literature review, we encountered additional Cas effectors that, although they do not comply with all suggested criteria, possess features that may benefit particular applications. For example, Ruminococcus flavefaciens XPD3002 (CasRx or RfxCas13d) has been recognized and intensely studied for its high RNA knockdown efficacy with minimal off-target activity in human cells [62]. Another example is LbCas12a, a Cas12 ortholog used in the well-established diagnostic tool DETECTR, which is a strict DNA endonuclease (but was combined with reverse transcription for RNA detection [23,87]). Finally, while editing this review, Li et al. (2021) reported the cryo-EM structure of Cas12g. This subtype specifically recognizes and requires ssRNA for its activation but exhibits both collateral DNA and RNA cleaving capabilities [88].
6. Conclusions
We summarized recent technologies for RNA detection with Cas proteins and provided a detailed description of the most important features to search for when selecting a Cas effector. Computational screens have already identified thousands of Cas orthologs targeting both DNA and RNA, yet we have barely started exploring the rich CRISPR toolbox that nature offers. Experimental characterization of Cas systems has largely focused on cell applications, whereas in vitro characterization data, if available, are often buried in the supplementary information. However, the informed design of in vitro biosensors critically depends on this biochemical characterization, which often reveals important deviations from the efficacy and selectivity observed in cells [71]. Bioengineering studies must describe how and why a particular Cas ortholog was selected. While it may seem safer to base a new design on a Cas system that many others have already employed, the initial choice of this system may have been arbitrary. We urge the CRISPR bioengineering community to document such design choices and help expand the field of well-characterized candidates.
Author Contributions
Conceptualization, E.D.-G., R.G. and S.T.A.; methodology, E.D.-G., R.G. and S.T.A.; investigation, E.D.-G., R.G. and S.T.A.; writing—original draft preparation, E.D.-G., R.G. and S.T.A.; writing—review and editing, E.D.-G., R.G. and S.T.A.; supervision, R.G. and S.T.A.; project administration, R.G. and S.T.A.; funding acquisition, R.G. and S.T.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the King Abdullah University of Science and Technology (KAUST) through the baseline fund and Award No. REI/1/4204-01 from the Office of Sponsored Research (OSR).
Acknowledgments
We apologize to any author whose work was not cited due to space limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide Sequence of the Iap Gene, Responsible for Alkaline Phosphatase Isozyme Conversion in Escherichia Coli, and Identification of the Gene Product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.; Juez, G.; Rodríguez-Valera, F. Transcription at Different Salinities of Haloferax Mediterranei Sequences Adjacent to Partially Modified PstI Sites. Mol. Microbiol. 1993, 9, 613–621. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Díez-Villaseñor, C.; Soria, E.; Juez, G. Biological Significance of a Family of Regularly Spaced Repeats in the Genomes of Archaea, Bacteria and Mitochondria. Mol. Microbiol. 2000, 36, 244–246. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR Elements in Yersinia Pestis Acquire New Repeats by Preferential Uptake of Bacteriophage DNA, and Provide Additional Tools for Evolutionary Studies. Microbiology 2005, 151, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Ehrlich, S.D. Clustered Regularly Interspaced Short Palindrome Repeats (CRISPRs) Have Spacers of Extrachromosomal Origin. Microbiology 2005, 151, 2551–2561. [Google Scholar] [CrossRef] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.K.; Saikot, F.K.; Rahman, M.S.; Jamal, M.A.H.M.; Rahman, S.M.K.; Islam, S.M.R.; Kim, K.-H. Programmable Molecular Scissors: Applications of a New Tool for Genome Editing in Biotech. Mol. Ther.-Nucleic Acids 2019, 14, 212–238. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Almendros, C. Short Motif Sequences Determine the Targets of the Prokaryotic CRISPR Defence System. Microbiology 2009, 155, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-CrRNA Ribonucleoprotein Complex Mediates Specific DNA Cleavage for Adaptive Immunity in Bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 Is a Single-Component Programmable RNA-Guided RNA-Targeting CRISPR Effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- East-Seletsky, A.; O’Connell, M.R.; Knight, S.C.; Burstein, D.; Cate, J.H.D.; Tjian, R.; Doudna, J.A. Two Distinct RNase Activities of CRISPR-C2c2 Enable Guide-RNA Processing and RNA Detection. Nature 2016, 538, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA Editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Su, J.; Hu, X.; Zhou, C.; Li, H.; Chen, Z.; Xiao, Q.; Wang, B.; Wu, W.; Sun, Y.; et al. Glia-to-Neuron Conversion by CRISPR-CasRx Alleviates Symptoms of Neurological Disease in Mice. Cell 2020, 181, 590–603.e16. [Google Scholar] [CrossRef] [PubMed]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza. Cell 2020, 181, 865–876.e12. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhou, H.; Zhou, X.; Li, F. Conferring Resistance to Plant RNA Viruses with the CRISPR/CasRx System. Virol. Sin. 2021, 36, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sun, W.; Shi, T.; Lu, P.; Zhuang, M.; Liu, J.-L. Capturing RNA–Protein Interaction via CRUIS. Nucleic Acids Res. 2020, 48, e52. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Tokuyasu, T.A. CRISPR-Cas13-Based RNA-Interacting Protein Detection in Living Cells. Biochemistry 2020, 59, 1791–1792. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a Target Binding Unleashes Indiscriminate Single-Stranded DNase Activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic Acid Detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary Classification of CRISPR–Cas Systems: A Burst of Class 2 and Derived Variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Strutt, S.C.; Torrez, R.M.; Kaya, E.; Negrete, O.A.; Doudna, J.A. RNA-Dependent RNA Targeting by CRISPR-Cas9. eLife 2018, 7, e32724. [Google Scholar] [CrossRef] [PubMed]
- Fozouni, P.; Son, S.; de Derby, M.D.L.; Knott, G.J.; Gray, C.N.; D’Ambrosio, M.V.; Zhao, C.; Switz, N.A.; Kumar, G.R.; Stephens, S.I.; et al. Amplification-Free Detection of SARS-CoV-2 with CRISPR-Cas13a and Mobile Phone Microscopy. Cell 2021, 184, 323–333.e9. [Google Scholar] [CrossRef]
- Ali, Z.; Aman, R.; Mahas, A.; Rao, G.S.; Tehseen, M.; Marsic, T.; Salunke, R.; Subudhi, A.K.; Hala, S.M.; Hamdan, S.M.; et al. ISCAN: An RT-LAMP-Coupled CRISPR-Cas12 Module for Rapid, Sensitive Detection of SARS-CoV-2. Virus Res. 2020, 288, 198129. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Kellner, M.J.; Joung, J.; Collins, J.J.; Zhang, F. Multiplexed and Portable Nucleic Acid Detection Platform with Cas13, Cas12a, and Csm6. Science 2018, 360, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-Y.; Cheng, Q.-X.; Wang, J.-M.; Li, X.-Y.; Zhang, Z.-L.; Gao, S.; Cao, R.-B.; Zhao, G.-P.; Wang, J. CRISPR-Cas12a-Assisted Nucleic Acid Detection. Cell Discov. 2018, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, S.; Wu, N.; Wu, J.; Wang, G.; Zhao, G.; Wang, J. HOLMESv2: A CRISPR-Cas12b-Assisted Platform for Nucleic Acid Detection and DNA Methylation Quantitation. ACS Synth. Biol. 2019, 8, 2228–2237. [Google Scholar] [CrossRef]
- Zhou, T.; Huang, R.; Huang, M.; Shen, J.; Shan, Y.; Xing, D. CRISPR/Cas13a Powered Portable Electrochemiluminescence Chip for Ultrasensitive and Specific MiRNA Detection. Adv. Sci. 2020, 7, 1903661. [Google Scholar] [CrossRef]
- Shinoda, H.; Taguchi, Y.; Nakagawa, R.; Makino, A.; Okazaki, S.; Nakano, M.; Muramoto, Y.; Takahashi, C.; Takahashi, I.; Ando, J.; et al. Amplification-Free RNA Detection with CRISPR–Cas13. Commun. Biol. 2021, 4, 1–7. [Google Scholar] [CrossRef]
- Tang, Z.; Chen, S.; Chen, A.; He, B.; Zhou, Y.; Chai, G.; Guo, F.; Huang, J. CasPDB: An Integrated and Annotated Database for Cas Proteins from Bacteria and Archaea. Database 2019, 2019, baz093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Zhao, S.; Ren, C.; Zhu, Y.; Zhou, H.; Lai, Y.; Zhou, F.; Jia, Y.; Zheng, K.; Huang, Z. CRISPRminer Is a Knowledge Base for Exploring CRISPR-Cas Systems in Microbe and Phage Interactions. Commun. Biol. 2018, 1, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A Web Tool to Identify Clustered Regularly Interspaced Short Palindromic Repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CRISPR-CAS++. Available online: https://crisprcas.i2bc.paris-saclay.fr/ (accessed on 5 July 2020).
- Özcan, A.; Krajeski, R.; Ioannidi, E.; Lee, B.; Gardner, A.; Makarova, K.S.; Koonin, E.V.; Abudayyeh, O.O.; Gootenberg, J.S. Programmable RNA Targeting with the Single-Protein CRISPR Effector Cas7-11. Nature 2021, 597, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F. Development of CRISPR-Cas Systems for Genome Editing and Beyond. Q. Rev. Biophys. 2019, 52, E6. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Zhao, P.; Olson, S.; Duff, M.O.; Graveley, B.R.; Wells, L.; Terns, R.M.; Terns, M.P. RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex. Cell 2009, 139, 945–956. [Google Scholar] [CrossRef] [Green Version]
- East-Seletsky, A.; O’Connell, M.R.; Burstein, D.; Knott, G.J.; Doudna, J.A. RNA Targeting by Functionally Orthogonal Type VI-A CRISPR-Cas Enzymes. Mol. Cell 2017, 66, 373–383.e3. [Google Scholar] [CrossRef] [Green Version]
- Kellner, M.J.; Koob, J.G.; Gootenberg, J.S.; Abudayyeh, O.O.; Zhang, F. SHERLOCK: Nucleic Acid Detection with CRISPR Nucleases. Nat. Protoc. 2019, 14, 2986–3012. [Google Scholar] [CrossRef]
- Chatterjee, P.; Jakimo, N.; Lee, J.; Amrani, N.; Rodríguez, T.; Koseki, S.R.T.; Tysinger, E.; Qing, R.; Hao, S.; Sontheimer, E.J.; et al. An Engineered ScCas9 with Broad PAM Range and High Specificity and Activity. Nat. Biotechnol. 2020, 38, 1154–1158. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA Targeting with CRISPR–Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Tang, L. PAM-Less Is More. Nature Methods 2020, 17, 559. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, M.R.; Oakes, B.L.; Sternberg, S.H.; East-Seletsky, A.; Kaplan, M.; Doudna, J.A. Programmable RNA Recognition and Cleavage by CRISPR/Cas9. Nature 2014, 516, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Nelles, D.A.; Fang, M.Y.; O’Connell, M.R.; Xu, J.L.; Markmiller, S.J.; Doudna, J.A.; Yeo, G.W. Programmable RNA Tracking in Live Cells with CRISPR/Cas9. Cell 2016, 165, 488–496. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, B.A.; Hou, Z.; Gramelspacher, M.J.; Zhang, Y. Programmable RNA Cleavage and Recognition by a Natural CRISPR-Cas9 System from Neisseria Meningitidis. Molecular Cell 2018, 69, 906–914.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickar-Oliver, A.; Black, J.B.; Lewis, M.M.; Mutchnick, K.J.; Klann, T.S.; Gilcrest, K.A.; Sitton, M.J.; Nelson, C.E.; Barrera, A.; Bartelt, L.C.; et al. Targeted Transcriptional Modulation with Type I CRISPR-Cas Systems in Human Cells. Nat. Biotechnol. 2019, 37, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Classification and Nomenclature of CRISPR-Cas Systems: Where from Here? CRISPR J. 2018, 1, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Park, K.-H.; An, Y.; Woo, E.-J. In vitro Assembly of Thermostable Csm Complex in CRISPR-Cas Type III/A System. Methods Enzym. 2019, 616, 173–189. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An Updated Evolutionary Classification of CRISPR–Cas Systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Makarova, K.S. Origins and Evolution of CRISPR-Cas Systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180087. [Google Scholar] [CrossRef] [PubMed]
- Chylinski, K.; Makarova, K.S.; Charpentier, E.; Koonin, E.V. Classification and Evolution of Type II CRISPR-Cas Systems. Nucleic Acids Res. 2014, 42, 6091–6105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Gao, A.; Zhan, Q.; Wang, Y.; Feng, H.; Liu, S.; Gao, G.; Serganov, A.; Gao, P. Diverse Mechanisms of CRISPR-Cas9 Inhibition by Type IIC Anti-CRISPR Proteins. Mol. Cell 2019, 74, 296–309.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.X.; Hunnewell, P.; Alfonse, L.E.; Carte, J.M.; Keston-Smith, E.; Sothiselvam, S.; Garrity, A.J.; Chong, S.; Makarova, K.S.; Koonin, E.V.; et al. Functionally Diverse Type V CRISPR-Cas Systems. Science 2019, 363, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellman, L.M.; Fried, M.G. Electrophoretic Mobility Shift Assay (EMSA) for Detecting Protein–Nucleic Acid Interactions. Nat. Protoc. 2007, 2, 1849–1861. [Google Scholar] [CrossRef]
- Tambe, A.; East-Seletsky, A.; Knott, G.J.; Doudna, J.A.; O’Connell, M.R. RNA-Binding and HEPN-Nuclease Activation Are Decoupled in CRISPR-Cas13a. Cell Rep. 2018, 24, 1025–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wandera, K.G.; Beisel, C.L. Rapidly Characterizing CRISPR-Cas13 Nucleases Using Cell-Free Transcription-Translation Systems. In Post-Transcriptional Gene Regulation; Dassi, E., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2022; pp. 135–153. ISBN 978-1-07-161851-6. [Google Scholar]
- Cléry, A.; Gillioz, L.; Nguyen, C.K.X.; Allain, F.H.-T. A Step-by-Step Guide to Study Protein–RNA Interactions. CHIMIA Int. J. Chem. 2019, 73, 406–414. [Google Scholar] [CrossRef]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and Evolution of Class 2 CRISPR–Cas Systems. Nat. Rev. Microbiol. 2017, 15, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Wessels, H.-H.; Méndez-Mancilla, A.; Guo, X.; Legut, M.; Daniloski, Z.; Sanjana, N.E. Massively Parallel Cas13 Screens Reveal Principles for Guide RNA Design. Nat. Biotechnol. 2020, 38, 722–727. [Google Scholar] [CrossRef]
- Gorski, S.A.; Vogel, J.; Doudna, J.A. RNA-Based Recognition and Targeting: Sowing the Seeds of Specificity. Nat. Rev. Mol. Cell Biol. 2017, 18, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ye, Y.; Ye, W.; Perčulija, V.; Jiang, H.; Chen, Y.; Li, Y.; Chen, J.; Lin, J.; Wang, S.; et al. Two HEPN Domains Dictate CRISPR RNA Maturation and Target Cleavage in Cas13d. Nat. Commun. 2019, 10, 2544. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Corsi, G.I.; Anthon, C.; Qu, K.; Pan, X.; Liang, X.; Han, P.; Dong, Z.; Liu, L.; Zhong, J.; et al. Enhancing CRISPR-Cas9 GRNA Efficiency Prediction by Data Integration and Deep Learning. Nat. Commun. 2021, 12, 3238. [Google Scholar] [CrossRef] [PubMed]
- Bandaru, S.; Tsuji, M.H.; Shimizu, Y.; Usami, K.; Lee, S.; Takei, N.K.; Yoshitome, K.; Nishimura, Y.; Otsuki, T.; Ito, T. Structure-Based Design of GRNA for Cas13. Sci. Rep. 2020, 10, 11610. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Rahman, J.A.; Wessels, H.-H.; Méndez-Mancilla, A.; Haro, D.; Chen, X.; Sanjana, N.E. Transcriptome-Wide Cas13 Guide RNA Design for Model Organisms and Viral RNA Pathogens. Cell Genom. 2021, 1, 100001. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, J.; Wang, Q.; Wang, Y.; Kang, C. Rapid Design and Development of CRISPR-Cas13a Targeting SARS-CoV-2 Spike Protein. Theranostics 2021, 11, 649–664. [Google Scholar] [CrossRef]
- RNAfold Web Server. Available online: http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi (accessed on 21 December 2021).
- RNAxs Web Server. Available online: http://rna.tbi.univie.ac.at/cgi-bin/RNAxs/RNAxs.cgi (accessed on 21 December 2021).
- Konermann, S.; Lotfy, P.; Brideau, N.J.; Oki, J.; Shokhirev, M.N.; Hsu, P.D. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell 2018, 173, 665–676.e14. [Google Scholar] [CrossRef] [Green Version]
- Kannan, S.; Altae-Tran, H.; Jin, X.; Madigan, V.J.; Oshiro, R.; Makarova, K.S.; Koonin, E.V.; Zhang, F. Compact RNA Editors with Small Cas13 Proteins. Nat. Biotechnol. 2021, 1–4. [Google Scholar] [CrossRef]
- Xu, C.; Zhou, Y.; Xiao, Q.; He, B.; Geng, G.; Wang, Z.; Cao, B.; Wang, X.; Zhou, D.; Yuan, T.; et al. Novel Miniature CRISPR–Cas13 Systems from Uncultivated Microbes Effective in Degrading SARS-CoV-2 Sequences and Influenza Viruses. Res. Square 2020. [Google Scholar] [CrossRef]
- Zhang, C.; Konermann, S.; Brideau, N.J.; Lotfy, P.; Wu, X.; Novick, S.J.; Strutzenberg, T.; Griffin, P.R.; Hsu, P.D.; Lyumkis, D. Structural Basis for the RNA-Guided Ribonuclease Activity of CRISPR-Cas13d. Cell 2018, 175, 212–223.e17. [Google Scholar] [CrossRef] [Green Version]
- Freije, C.A.; Myhrvold, C.; Boehm, C.K.; Lin, A.E.; Welch, N.L.; Carter, A.; Metsky, H.C.; Luo, C.Y.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Programmable Inhibition and Detection of RNA Viruses Using Cas13. Mol. Cell 2019, 76, 826–837.e11. [Google Scholar] [CrossRef] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Kellner, M.J.; Zhang, F. Nucleic Acid Detection of Plant Genes Using CRISPR-Cas13. CRISPR J. 2019, 2, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Mahas, A.; Aman, R.; Mahfouz, M. CRISPR-Cas13d Mediates Robust RNA Virus Interference in Plants. Genome Biol. 2019, 20, 263. [Google Scholar] [CrossRef] [Green Version]
- Nishimasu, H.; Cong, L.; Yan, W.X.; Ran, F.A.; Zetsche, B.; Li, Y.; Kurabayashi, A.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal Structure of Staphylococcus Aureus Cas9. Cell 2015, 162, 1113–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, F.A.; Cong, L.; Yan, W.X.; Scott, D.A.; Gootenberg, J.S.; Kriz, A.J.; Zetsche, B.; Shalem, O.; Wu, X.; Makarova, K.S.; et al. In Vivo Genome Editing Using Staphylococcus Aureus Cas9. Nature 2015, 520, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Friedland, A.E.; Baral, R.; Singhal, P.; Loveluck, K.; Shen, S.; Sanchez, M.; Marco, E.; Gotta, G.M.; Maeder, M.L.; Kennedy, E.M.; et al. Characterization of Staphylococcus Aureus Cas9: A Smaller Cas9 for All-in-One Adeno-Associated Virus Delivery and Paired Nickase Applications. Genome Biol. 2015, 16, 257. [Google Scholar] [CrossRef] [Green Version]
- Batra, R.; Nelles, D.A.; Pirie, E.; Blue, S.M.; Marina, R.J.; Wang, H.; Chaim, I.A.; Thomas, J.D.; Zhang, N.; Nguyen, V.; et al. Elimination of Toxic Microsatellite Repeat Expansion RNA by RNA-Targeting Cas9. Cell 2017, 170, 899–912.e10. [Google Scholar] [CrossRef]
- Yourik, P.; Fuchs, R.T.; Mabuchi, M.; Curcuru, J.L.; Robb, G.B. Staphylococcus Aureus Cas9 Is a Multiple-Turnover Enzyme. RNA 2019, 25, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Mekler, V.; Kuznedelov, K.; Severinov, K. Quantification of the Affinities of CRISPR–Cas9 Nucleases for Cognate Protospacer Adjacent Motif (PAM) Sequences. J. Biol. Chem. 2020, 295, 6509–6517. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Li, X.; Wang, J.; Wang, M.; Chen, P.; Yin, M.; Li, J.; Sheng, G.; Wang, Y. Two Distant Catalytic Sites Are Responsible for C2c2 RNase Activities. Cell 2017, 168, 121–134.e12. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Li, X.; Ma, J.; Li, Z.; You, L.; Wang, J.; Wang, M.; Zhang, X.; Wang, Y. The Molecular Architecture for RNA-Guided RNA Cleavage by Cas13a. Cell 2017, 170, 714–726.e10. [Google Scholar] [CrossRef]
- Yan, W.X.; Chong, S.; Zhang, H.; Makarova, K.S.; Koonin, E.V.; Cheng, D.R.; Scott, D.A. Cas13d Is a Compact RNA-Targeting Type VI CRISPR Effector Positively Modulated by a WYL-Domain-Containing Accessory Protein. Mol. Cell 2018, 70, 327–339.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, A.; Huyke, D.A.; Sharma, E.; Sahoo, M.K.; Huang, C.; Banaei, N.; Pinsky, B.A.; Santiago, J.G. Electric Field-Driven Microfluidics for Rapid CRISPR-Based Diagnostics and Its Application to Detection of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 29518–29525. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, H.; Xiao, R.; Han, R.; Chang, L. Cryo-EM Structure of the RNA-Guided Ribonuclease Cas12g. Nat. Chem. Biol. 2021, 17, 387–393. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Selection criteria for RNA-targeting Cas. (a) General steps. (b) The choices leading to the four candidates described in greater detail are represented in blue.
Figure 1.
Selection criteria for RNA-targeting Cas. (a) General steps. (b) The choices leading to the four candidates described in greater detail are represented in blue.

Figure 2.
Schematic depiction of the RNA-targeting Cas system. (a) The Cas:gRNA complex comprises two parts: the Cas protein and guide RNA (gRNA). (b) The recognition and binding of target RNA activate the nuclease activity of the Cas:gRNA complex, resulting in the cleavage of target RNA. Some Cas orthologs, upon activation, also cleave nontarget, “collateral” RNA.
Figure 2.
Schematic depiction of the RNA-targeting Cas system. (a) The Cas:gRNA complex comprises two parts: the Cas protein and guide RNA (gRNA). (b) The recognition and binding of target RNA activate the nuclease activity of the Cas:gRNA complex, resulting in the cleavage of target RNA. Some Cas orthologs, upon activation, also cleave nontarget, “collateral” RNA.

Figure 3.
Key features of selected Cas effectors.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Features of CRISPR-Cas systems by type.
| Class | Features | Type | Features | Key Effectors | Target | Ref. |
|---|---|---|---|---|---|---|
| 1 | Effector, adaptation, and accessory functions distributed over multiple proteins | I |
| Cas1, Cas2, Cas4, Cas5, Cas6, Cas3, Cas8 | dsDNA | [25,49,50] |
| III |
| Cas1, Cas2, Cas5, Cas6, Cas7, Cas10 | dsDNA, RNA | [51,52] | ||
| IV |
| Cas5, Cas7, Csf1 | dsDNA | [52,53] | ||
| 2 | Single protein with multiple domains combines crRNA-binding, catalytic activity, and pre-crRNA processing | II |
| Cas9 | dsDNA, RNA | [12,25,54,55] |
| V |
| Cas12, Cas14 | dsDNA, ssDNA, RNA | [25,56] | ||
| VI |
| Cas13 | RNA | [15,25,41,54] |
Table 2.
Feature summary for the four selected Cas effector candidates.
| Identifier | Source Organism | Features | gRNA |
|---|---|---|---|
| LwaCas13a | Leptotrichia wadeii | Size: 1389 aa Structure: not available Nuclease domain: HEPN dCas mutations: D403G, R474A, and R1046A [44] PFS: not required Optimal spacer length: 20–28 nt Specificity: collateral cleavage in vitro but not in mammalian cells Turnover kinetics: not available Others: used for SHERLOCK diagnostics | 5′-GATTTAGACTACCCCAAAAACGAAGGGGACTAAAAC-SPACER |
| SauCas9 | Staphylococcus aureus | Size: 1053 aa Structure: 5AXW Nuclease domain: RuvC and HNH dCas mutations: D10A (RuvC), N580A (HNH) [26] PAM/PFS: PFS not required for ssRNA targeting; PAM required for dsDNA targeting (5’NNGRRT) Optimal spacer length: 23 nt Specificity: High DNA target specificity due to long PAM; mismatch tolerance characterization available for its RNA-targeting role; no collateral activity Turnover kinetics: DNA (multiple), RNA (single) Others: target secondary structure affects affinity | 5′-SPACER- GTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTT |
| LbuCas13a | Leptotrichia buccalis | Size: 1159 aa Structure: 5XWP Nuclease domain: HEPN dCas mutations: R472A, H477A, R1048A and H1053A [84] PFS: not required Optimal spacer length: 20–24 nt Activity: collateral activity with high turnover (104 turnovers per target RNA recognized) [41] Turnover kinetics: not available for target cleavage but multiple turnover for collateral cleavage | 5′- GGCCACCCCAAAAATGAAGGGGACTAAAACA-SPACER |
| EsCas13d | Eubacterium siraeum | Size: 954 aa Structure: 6E9F Nuclease domain: HEPN dCas mutations: R295A, H300A, R849A and H854A [71] PFS: not required Optimal spacer length: 20–30 nt Specificity: collateral activity Turnover kinetics: not available Others: robust expression in E. coli; limited activity in mammalian cells [71] | 5′ AACTACACCCGTGCAAAAATGCAGGGGTCTAAAAC-SPACER |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Díaz-Galicia, E.; Grünberg, R.; Arold, S.T. How to Find the Right RNA-Sensing CRISPR-Cas System for an In Vitro Application. Biosensors 2022, 12, 53. https://0-doi-org.brum.beds.ac.uk/10.3390/bios12020053
AMA Style
Díaz-Galicia E, Grünberg R, Arold ST. How to Find the Right RNA-Sensing CRISPR-Cas System for an In Vitro Application. Biosensors. 2022; 12(2):53. https://0-doi-org.brum.beds.ac.uk/10.3390/bios12020053
Chicago/Turabian StyleDíaz-Galicia, Escarlet, Raik Grünberg, and Stefan T. Arold. 2022. "How to Find the Right RNA-Sensing CRISPR-Cas System for an In Vitro Application" Biosensors 12, no. 2: 53. https://0-doi-org.brum.beds.ac.uk/10.3390/bios12020053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.