
Antimicrobial Susceptibility and Detection of Virulence-Associated Genes in Escherichia coli Strains Isolated from Commercial Broilers


Abstract
:1. Introduction
2. Results
2.1. Clermont’s Phylogenetic Typing
2.2. Detection of Iron-Uptake and Virulence-Associated Genes (VGs)
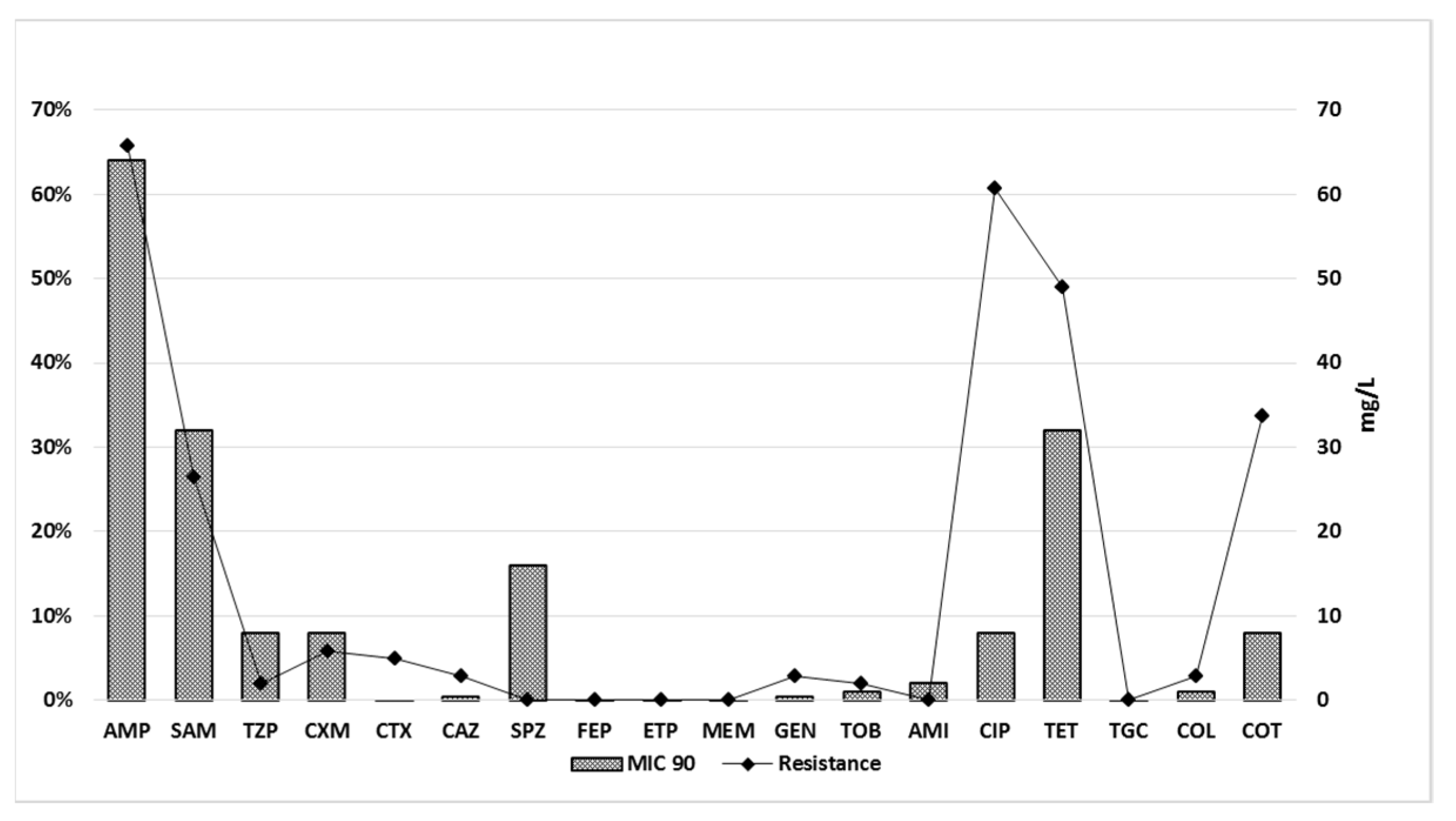
2.3. Antimicrobial Resistance
3. Discussion
4. Materials and Methods
4.1. Samples, Bacterial Isolation, and Identification
4.2. DNA Extraction, Clermont’s Phylogenetic Typing, and Detection of Iron Uptake and Virulence-Associated Genes
4.3. Antimicrobial Susceptibility Testing
4.4. Detection of Antimicrobial Resistance Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maluta, R.P.; Logue, C.M.; Casas, M.R.T.; Meng, T.; Guastalli, E.A.L.; Rojas, T.C.G.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped Sequence Types (STs) and Serogroups of Avian Pathogenic (APEC) and Human Extra-Intestinal Pathogenic (ExPEC) Escherichia coli Isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Lee, J.-Y.; Kang, M.-S.; Lee, H.-J.; Kang, S.-I.; Lee, O.-M.; Kwon, Y.-K.; Kim, J.-H. Comparative Characteristics and Zoonotic Potential of Avian Pathogenic Escherichia coli (APEC) Isolates from Chicken and Duck in South Korea. Microorganisms 2021, 9, 946. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, Z.; Shi, J.; Jia, Y.; Yang, K.; Wang, Z. Correlation between Exogenous Compounds and the Horizontal Transfer of Plasmid-Borne Antibiotic Resistance Genes. Microorganisms 2020, 8, 1211. [Google Scholar] [CrossRef]
- Kabir, S.M.L. Avian Colibacillosis and Salmonellosis: A Closer Look at Epidemiology, Pathogenesis, Diagnosis, Control and Public Health Concerns. Int. J. Environ. Res. Public Health 2010, 7, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messaili, C.; Messai, Y.; Bakour, R. Virulence Gene Profiles, Antimicrobial Resistance and Phylogenetic Groups of Fecal Escherichia coli Strains Isolated from Broiler Chickens in Algeria. Vet. Ital. 2019, 55, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and Molecular Characterization of Nalidixic Acid-Resistant Extraintestinal Pathogenic Escherichia coli from Retail Chicken Products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stromberg, Z.R.; Johnson, J.R.; Fairbrother, J.M.; Kilbourne, J.; Van Goor, A.; Curtiss, R.; Mellata, M. Evaluation of Escherichia coli Isolates from Healthy Chickens to Determine Their Potential Risk to Poultry and Human Health. PLoS ONE 2017, 12, e0180599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia Coli Isolates That Carry Vat, FyuA, ChuA, and YfcV Efficiently Colonize the Urinary Tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [Green Version]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, C.; Yu, X.; Xia, M.; Yue, H. Distribution of Serotypes and Virulence-Associated Genes in Pathogenic Escherichia coli Isolated from Ducks. Avian Pathol. 2010, 39, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, P.R.; Yadav, P.; Hemlata, H.; Tejavath, K.K.; Singh, A.P. Poultry-origin Extraintestinal Escherichia coli Strains Carrying the Traits Associated with Urinary Tract Infection, Sepsis, Meningitis and Avian Colibacillosis in India. J. Appl. Microbiol. 2021, 130, 2087–2101. [Google Scholar] [CrossRef]
- Dadi, B.R.; Abebe, T.; Zhang, L.; Mihret, A.; Abebe, W.; Amogne, W. Distribution of Virulence Genes and Phylogenetics of Uropathogenic Escherichia coli among Urinary Tract Infection Patients in Addis Ababa, Ethiopia. BMC Infect. Dis. 2020, 20, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdi, M.; Bouzari, M.; Ghaemi, E.A. Detection of Fim, Pap, Sfa and Afa Adhesin-Encoding Operons in Escherichia coli Strains Isolated from Urinary Tract Infections. Med. Lab. J. 2018, 12, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Köhler, C.-D.; Dobrindt, U. What Defines Extraintestinal Pathogenic Escherichia Coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Seyoum, Y.; Baye, K.; Humblot, C. Iron Homeostasis in Host and Gut Bacteria—A Complex Interrelationship. Gut Microbes 2021, 13, 1874855. [Google Scholar] [CrossRef] [PubMed]
- Fardeau, S.; Mullié, C.; Dassonville-Klimpt, A.; Audic, N.; Sonnet, P. Bacterial iron uptake: A promising solution against multidrug resistant bacteria. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Vilas, M., Ed.; Formatex Research Center: Badajoz, Spain, 2011; Volume 2, pp. 695–705. [Google Scholar]
- Tu, J.; Xue, T.; Qi, K.; Shao, Y.; Huang, B.; Wang, X.; Zhou, X. The Irp2 and FyuA Genes in High Pathogenicity Islands Are Involved in the Pathogenesis of Infections Caused by Avian Pathogenic Escherichia coli (APEC). Pol. J. Vet. Sci. 2016, 19, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handrova, L.; Kmet, V. Antibiotic Resistance and Virulence Factors of Escherichia coli from Eagles and Goshawks. J. Environ. Sci. Health Part B 2019, 54, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.K.Y.; Krewulak, K.D.; Vogel, H.J. Bacterial Ferrous Iron Transport: The Feo System. FEMS Microbiol. Rev. 2016, 40, 273–298. [Google Scholar] [CrossRef]
- Mosquito, S.; Pons, M.J.; Riveros, M.; Ruiz, J.; Ochoa, T.J. Diarrheagenic Escherichia coli Phylogroups Are Associated with Antibiotic Resistance and Duration of Diarrheal Episode. Sci. World J. 2015, 2015, 610403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jibril, A.H.; Okeke, I.N.; Dalsgaard, A.; Olsen, J.E. Association between Antimicrobial Usage and Resistance in Salmonella from Poultry Farms in Nigeria. BMC Vet. Res. 2021, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Joosten, P.; Sarrazin, S.; Van Gompel, L.; Luiken, R.E.C.; Mevius, D.J.; Wagenaar, J.A.; Heederik, D.J.J.; Dewulf, J.; EFFORT consortium; Graveland, H.; et al. Quantitative and Qualitative Analysis of Antimicrobial Usage at Farm and Flock Level on 181 Broiler Farms in Nine European Countries. J. Antimicrob. Chemother. 2019, 74, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Johar, A.; Al-Thani, N.; Al-Hadidi, S.H.; Dlissi, E.; Mahmoud, M.H.; Eltai, N.O. Antibiotic Resistance and Virulence Gene Patterns Associated with Avian Pathogenic Escherichia coli (APEC) from Broiler Chickens in Qatar. Antibiotics 2021, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The Application of Antibiotics in Broiler Production and the Resulting Antibiotic Resistance in Escherichia Coli: A Global Overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Sgariglia. Antibiotic Resistance Pattern and Virulence Genesin Avian Pathogenic Escherichia coli (APEC) from Different Breeding Systems. Vet. Ital. 2019, 55, 26–33. [Google Scholar] [CrossRef]
- Agyare, C.; Etsiapa Boamah, V.; Ngofi Zumbi, C.; Boateng Osei, F. Antibiotic Use in Poultry Production and Its Effects on Bacterial Resistance. In Antimicrobial Resistance—A Global Threat; Kumar, Y., Ed.; IntechO: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Adelowo, O.O.; Fagade, O.E.; Agersø, Y. Antibiotic Resistance and Resistance Genes in Escherichia coli from Poultry Farms, Southwest Nigeria. J. Infect. Dev. Ctries. 2014, 8, 1103–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bogaard, A.E. Antibiotic Resistance of Faecal Escherichia coli in Poultry, Poultry Farmers and Poultry Slaughterers. J. Antimicrob. Chemother. 2001, 47, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Khanal, T.; Raut, S.B.; Paneru, U. Study of Antibiotic Resistance on Escherichia coli in Commercial Poultry of Nepal. Nep. Vet. J. 2017, 34, 6–17. [Google Scholar] [CrossRef] [Green Version]
- Cassier, P.; Lallechère, S.; Aho, S.; Astruc, K.; Neuwirth, C.; Piroth, L.; Chavanet, P. Cephalosporin and Fluoroquinolone Combinations Are Highly Associated with CTX-M β-Lactamase-Producing Escherichia Coli: A Case–Control Study in a French Teaching Hospital. Clin. Microbiol. Infect. 2011, 17, 1746–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A. Resistance Plasmid Families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Baudry, P.J.; Nichol, K.; DeCorby, M.; Lagacé-Wiens, P.; Olivier, E.; Boyd, D.; Mulvey, M.R.; Hoban, D.J.; Zhanel, G.G. Mechanisms of Resistance and Mobility among Multidrug-Resistant CTX-M–Producing Escherichia coli from Canadian Intensive Care Units: The 1st Report of QepA in North America. Diagn. Microbiol. Infect. Dis. 2009, 63, 319–326. [Google Scholar] [CrossRef]
- Meletis, G. Carbapenem Resistance: Overview of the Problem and Future Perspectives. Ther. Adv. Infect. 2016, 3, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepas, V.; Soto, S.M. Relationship between Virulence and Resistance among Gram-Negative Bacteria. Antibiotics 2020, 9, 719. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Li, Y.; Hao, C. Association between Virulence Profile and Fluoroquinolone Resistance in Escherichia coli Isolated from Dogs and Cats in China. J. Infect. Dev. Ctries. 2017, 11, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Karam, M.R.A.; Habibi, M.; Bouzari, S. Relationships between Virulence Factors and Antimicrobial Resistance among Escherichia coli Isolated from Urinary Tract Infections and Commensal Isolates in Tehran, Iran. Osong Public Health Res. Perspect. 2018, 9, 217–224. [Google Scholar] [CrossRef]
- Jauréguy, F.; Carbonnelle, E.; Bonacorsi, S.; Clećh, C.; Casassus, P.; Bingen, E.; Picard, B.; Nassif, X.; Lortholary, O. Host and Bacterial Determinants of Initial Severity and Outcome of Escherichia coli Sepsis. Clin. Microbiol. Infect. 2007, 13, 854–862. [Google Scholar] [CrossRef]
- Bessède, E.; Angla-gre, M.; Delagarde, Y.; Sep Hieng, S.; Ménard, A.; Mégraud, F. Matrix-Assisted Laser-Desorption/Ionization BIOTYPER: Experience in the Routine of a University Hospital. Clin. Microbiol. Infect. 2011, 17, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli Phylo-Typing Method Revisited: Improvement of Specificity and Detection of New Phylo-Groups: A New E. Coli Phylo-Typing Method. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Lescat, M.; Clermont, O.; Woerther, P.L.; Glodt, J.; Dion, S.; Skurnik, D.; Djossou, F.; Dupont, C.; Perroz, G.; Picard, B.; et al. Commensal Escherichia coli Strains in Guiana Reveal a High Genetic Diversity with Host-Dependant Population Structure: Commensal Escherichia coli Population Structure. Environ. Microbiol. Rep. 2013, 5, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Russo, T.A.; Tarr, P.I.; Carlino, U.; Bilge, S.S.; Vary, J.C.; Stell, A.L. Molecular Epidemiological and Phylogenetic Associations of Two Novel Putative Virulence Genes, Iha and IroN E. coli, among Escherichia coli Isolates from Patients with Urosepsis. Infect. Immun. 2000, 68, 3040–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended Virulence Genotypes of Escherichia coli Strains from Patients with Urosepsis in Relation to Phylogeny and Host Compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Carlino, U.B.; Johnson, J.R. Identification of a New Iron-Regulated Virulence Gene, IreA, in an Extraintestinal Pathogenic Isolate of Escherichia. Coli. Infect. Immun. 2001, 69, 6209–6216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runyen-Janecky, L.J.; Reeves, S.A.; Gonzales, E.G.; Payne, S.M. Contribution of the Shigella Flexneri Sit, Iuc, and Feo Iron Acquisition Systems to Iron Acquisition In Vitro and in Cultured Cells. Infect. Immun. 2003, 71, 1919–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC Pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, C.; Li, G.; Wilking, H.; Kiebling, S.; Alt, K.; Antao, E.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T. Avian Pathogenic, Uropathogenic, and Newborn Meningitis-Causing Escherichia Coli: How Closely Related Are They? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Le Bouguenec, C.; Archambaud, M.; Labigne, A. Rapid and Specific Detection of the Pap, Afa, and Sfa Adhesin-Encoding Operons in Uropathogenic Escherichia coli Strains by Polymerase Chain Reaction. J. Clin. Microbiol. 1992, 30, 1189–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dozois, C.M.; Dho-Moulin, M.; Brée, A.; Fairbrother, J.M.; Desautels, C.; Curtiss, R. Relationship between the Tsh Autotransporter and Pathogenicity of Avian Escherichia coli and Localization and Analysis of the Tsh Genetic Region. Infect. Immun. 2000, 68, 4145–4154. [Google Scholar] [CrossRef] [Green Version]
- Germon, P.; Chen, Y.-H.; He, L.; Blanco, J.E.; Brée, A.; Schouler, C.; Huang, S.-H.; Moulin-Schouleur, M. IbeA, a Virulence Factor of Avian Pathogenic Escherichia Coli. Microbiology 2005, 151, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Robicsek, A.; Strahilevitz, J.; Sahm, D.F.; Jacoby, G.A.; Hooper, D.C. Qnr Prevalence in Ceftazidime-Resistant Enterobacteriaceae Isolates from the United States. Antimicrob. Agents Chemother. 2006, 50, 2872–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a Novel Plasmid-Mediated Colistin-Resistance Gene, Mcr-2, in Escherichia Coli, Belgium, June 2016. Eurosurveillance 2016, 21, 30280. [Google Scholar] [CrossRef]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for Rapid Detection of Genes Encoding CTX-M Extended-Spectrum β-Lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillaume, G.; Verbrugge, D.; Chasseur-Libotte, M.-L.; Moens, W.; Collard, J.-M. PCR Typing of Tetracycline Resistance Determinants (Tet A–E) in Salmonella Enterica Serotype Hadar and in the Microbial Community of Activated Sludges from Hospital and Urban Wastewater Treatment Facilities in Belgium. FEMS Microbiol. Ecol. 2000, 32, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Kerrn, M.B. Susceptibility of Danish Escherichia coli Strains Isolated from Urinary Tract Infections and Bacteraemia, and Distribution of Sul Genes Conferring Sulphonamide Resistance. J. Antimicrob. Chemother. 2002, 50, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingshead, S.; Vapnek, D. Nucleotide Sequence Analysis of a Gene Encoding a Streptomycin/Spectinomycin Adenyltransferase. Plasmid 1985, 13, 17–30. [Google Scholar] [CrossRef]
- Navia, M.M.; Ruiz, J.; Sanchez-Cespedes, J.; Vila, J. Detection of Dihydrofolate Reductase Genes by PCR and RFLP. Diagn. Microbiol. Infect. Dis. 2003, 46, 295–298. [Google Scholar] [CrossRef]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. Antibiotic Resistance in the ECOR Collection: Integrons and Identification of a Novel Aad Gene. Antimicrob. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weill, F.-X.; Demartin, M.; Fabre, L.; Grimont, P.A.D. Extended-Spectrum-β-Lactamase (TEM-52)-Producing Strains of Salmonella Enterica of Various Serotypes Isolated in France. J. Clin. Microbiol. 2004, 42, 3359–3362. [Google Scholar] [CrossRef] [Green Version]
- Gattringer, R.; Niks, M.; Ostertag, R.; Schwarz, K.; Medvedovic, H.; Graninger, W.; Georgopoulos, A. Evaluation of MIDITECH Automated Colorimetric MIC Reading for Antimicrobial Susceptibility Testing. J. Antimicrob. Chemother. 2002, 49, 651–659. [Google Scholar] [CrossRef] [PubMed]
- EUCAST (European Committee of Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 11.0, Valid from 1 January 2021. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_11.0_Breakpoint_Tables.pdf (accessed on 6 September 2021).


{kind=link}
| Pathotype | Number (%) of All Samples | A | B1 | B2 | D | F | Clade I | Unknown |
|---|---|---|---|---|---|---|---|---|
| ExPEC | 25 (21.7) | 9 (36.0) | 2 (8.0) | 0 (0) | 9 (36.0) | 4 (16.0) | 0 (0) | 1 (4.0) |
| UPEC | 8 (7.0) | 0 (0) | 0 (0) | 6 (75.0) | 2 (25.0) | 0 (0) | 0 (0) | 0 (0) |
| APEC | 7 (6.1) | 0 (0) | 2 (28.6) | 0 (0) | 0 (0) | 3 (42.9) | 1 (14.3) | 1 (14.3) |
| Phylogroup | Resistant Strains | |
|---|---|---|
| n | % | |
| A | 13 | 17.1 |
| B1 | 27 | 35.5 |
| B2 | 5 | 6.6 |
| C | 1 | 1.3 |
| D | 16 | 21.1 |
| F | 7 | 9.2 |
| Clade I | 2 | 2.6 |
| Unknown | 5 | 6.6 |
| Total | 76 | 100 |
| Strain Group | Mobilome | Virulence | Phenotypic Resistance | Genotypic Resistance | |
|---|---|---|---|---|---|
| Commensals | B-1 B1 | Int1, Tn3 | feoB | AMP, SAM, CXM, CAZ, TOB, CIP, COT | aac(6′)-Ib-cr, sul1, dfrA |
| B-2 A | Int1, Tn3 | feoB | AMP, SAM, CXM, CTX, CAZ, CIP, COT | aac(6′)-Ib-cr, sul1, dfrA | |
| B-13 B1 | Tn3 | feoB, sitA, iroN, iss | AMP, CXM, CTX, CIP, TET, COT | tetA, blaCTX-M-2 | |
| B-23 A | feoB, sitA, iutA, iroN, tsh, iss, cvaC | AMP, SAM, CIP | qnrS | ||
| UPEC | B-22 B2 | Int1, Tn3 | sitA, chuA, iutA, irp2, fyuA, papC, cvaC | AMP, CXM, CTX, CIP, TET, COT | aadA, sul2, tetB, dfrA, blaCTX-M-1 |
| B-42 B2 | feoB, sitA, chuA, iutA, iroN, ibeA, irp2, fyuA, tsh, iss, cvaC | Sensitive | |||
| B-21 B2 | Tn3 | feoB, sitA, chuA, iutA, irp2, fyuA, papC, cvaC | AMP, CIP, TET | aadA, sul2, tetB, dfrA | |
| B-32 D | Int1, Tn3 | feoB, sitA, chuA, iutA, irp2, fyuA, kpsMTII | AMP, SAM, CIP, COT | aadA | |
| APEC | B-31 F | Tn3 | feoB, sitA, chuA, iutA, iroN, irp2, tsh, iss, cvaC, kpsMTII | AMP, SAM, CIP, TET | qnrS, tetA |
| B-71 clade I | Int1, Tn3 | feoB, sitA, chuA, iutA, iroN, irp2, fyuA, papC, cvaC, kpsMTII | AMP, CIP, COT | aadA, sul2, tetB | |
| ExPEC | B-93 D | Int1, Tn3 | feoB, sitA, chuA, iutA, ibeA, iss, kpsMTII | AMP, GEN, TOB, CIP, TET, COT | aadA, sul1, sul2, tetA, dfrA |
| Gene | Primer Sequences (5′–3′) | Product (bp) | Annealing (°C) | References |
|---|---|---|---|---|
| arpA | AACGCTATTCGCCAGCTTGC TCTCCCCATACCGTACGCTA | 400 | 59 | [39] |
| chuA | ATGGTACCGGACGAACCAAC TGCCGCCAGTACCAAAGACA | 288 | 59 | [39] |
| yjaA | CAAACGTGAAGTGTCAGGAG AATGCGTTCCTCAACCTGTG | 211 | 59 | [39] |
| TspE4.C2 | CACTATTCGTAAGGTCATCC AGTTTATCGCTGCGGGTCGC | 152 | 59 | [39] |
| arpA (group C) | GATTCCATCTTGTCAAAATATGCC GAAAAGAAAAAGAATTCCCAAGAG | 301 | 57 | [40] |
| trpA (group E) | AGTTTTATGCCCAGTGCGAG TCTGCGCCGGTCACGCCC | 319 | 59 | [40] |
| iha | CTGGCGGAGGCTCTGAGATCA TCCTTAAGCTCCCGCGGCTGA | 827 | 60 | [41] |
| iroN | AAGTCAAAGCAGGGGTTGCCCG GACGCCGACATTAAGACGCAG | 655 | 60 | [41] |
| fyuA | TGATTAACCCCGCGACGGGAA CGCAGTAGGCACGATGTTGTA | 880 | 55 | [42] |
| ireA | TGGTCTTCAGCTATATGG ATCTATGATTGTGTTGGT | 415 | 55 | [43] |
| sitA | AGGGGGCACAACTGATTCTCG TACCGGGCCGTTTTCTGTGC | 608 | 59 | [44] |
| feoB | AATTGGCGTGCATGAAGATAACTG AGCTGGCGACCTGATAGAACAATG | 470 | 59 | [45] |
| irp2 | AAGGATTCGCGTGAC TCGTCGGGCAGCGTTTCTTCT | 287 | 59 | [45] |
| iss | ATCACATAGGATTCTGCCG ACAAAAAGTTCTATCGCTTCC | 700 | 61 | [46] |
| papC | GACGGCTGTACTGCAGGGTGTGGCG ATATCCTTTCTGCAGGGATGCAATA | 328 | 61 | [47] |
| iutA | GGCTGGACATGGGAACTGG CGTCGGGAACGGGTAGAATCG | 300 | 63 | [42] |
| cvaC | CACACACAAACGGGAGCTGTT CACACACAAACGGGAGCTGTT | 680 | 63 | [42] |
| tsh | GGTGGTGCACTGGAGTGG AGTCCAGCGTGATAGTGG | 620 | 55 | [48] |
| ibeA | TGAACGTTTCGGTTGTTTTG TGTTCAAATCCTGGCTGGAA | 814 | 55 | [49] |
| kps MTII | GCGCATTTGCTGATACTGTTG CATCCAGACGATAAGCATGAGCA | 272 | 63 | [42] |
| qnrA | ATTTCTCACGCCAGGATTTG GATCGGCAAAGGTTAGGTCA | 516 | 53 | [50] |
| qnrB | GATCGTGAAAGCCAGAAAGG ACGATGCCTGGTAGTTGTCC | 469 | 53 | [50] |
| qnrS | ACGACATTCGTCAACTGCAA TAAATTGGCACCCTGTAGGC | 417 | 53 | [50] |
| aac(6)-Ib-cr | GATCTCATATCGTCGAGTGGTGG GAACCATGTACACGGCTGGAC | 435 | 58 | [50] |
| mcr-1 | CGGTCAGTCCGTTTGTTC CTTGGTCGGTCTGTAGGG | 309 | 58 | [51] |
| mcr-2 | TGTTGCTTGTGCCGATTGGA AGATGGTATTGTTGGTTGCTG | 567 | 58 | [52] |
| blaCTX-M-1 | AAAAATCACTGCGCCAGTTC AGCTTATTCATCGCCACGTT | 415 | 52 | [53] |
| blaCTX-M-2 | CGACGCTACCCCTGCTATT CCAGCGTCAGATTTTTCAGG | 552 | 52 | [53] |
| tetA | GGCCTCAATTTCCTGACG AAGCAGGATGTAGCCTGTGC | 372 | 55 | [54] |
| tetB | GAGACGCAATCGAATTCGG TTTAGTGGCTATTCTTCCTGCC | 228 | 55 | [54] |
| sul1 | CGGCGTGGGCTACCTGAACG GCCGATCGCGTGAAGTTCCG | 433 | 69 | [55] |
| sul2 | GCGCTCAAGGCAGATGGCATT GCGTTTGATACCGGCACCCGT | 293 | 69 | [55] |
| aadA | TGATTTGCTGGTTACGGTGAC CGCTATGTTCTCTTGCTTTTG | 284 | 60 | [56] |
| dfrA | GTGAAACTATCACTAATGG TTAACCCTTTTGCCAGATTT | 474 | 55 | [57] |
| drfB | GATCGCCTGCGCAAGAAATC AAGCGCAGCCACAGGATAAAT | 141 | 60 | [57] |
| Int1 | GGGTCAAGGATCTGGATTTCG ACATGCGTGTAAATCATCGTCG | 483 | 62 | [58] |
| Tn3 | CACGAATGAGGGCCGACAGGA ACCCACTCGTGCACCCAACTG | 500 | 58 | [59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocúreková, T.; Karahutová, L.; Bujňáková, D. Antimicrobial Susceptibility and Detection of Virulence-Associated Genes in Escherichia coli Strains Isolated from Commercial Broilers. Antibiotics 2021, 10, 1303. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111303
Kocúreková T, Karahutová L, Bujňáková D. Antimicrobial Susceptibility and Detection of Virulence-Associated Genes in Escherichia coli Strains Isolated from Commercial Broilers. Antibiotics. 2021; 10(11):1303. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111303
Chicago/Turabian StyleKocúreková, Tímea, Lívia Karahutová, and Dobroslava Bujňáková. 2021. "Antimicrobial Susceptibility and Detection of Virulence-Associated Genes in Escherichia coli Strains Isolated from Commercial Broilers" Antibiotics 10, no. 11: 1303. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111303