
Sub-Inhibitory Concentrations of Ciprofloxacin Alone and Combinations with Plant-Derived Compounds against P. aeruginosa Biofilms and Their Effects on the Metabolomic Profile of P. aeruginosa Biofilms


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Antibiotics and Plant-Derived Compounds
2.3. MIC, MBIC, and MBEC Assay
2.4. Evaluation of the Antibiofilm Effects of Ciprofloxacin in Combination with the Compounds on Mono- and Triple-Species Biofilms of P. aeruginosa PAO1 and SEM Analysis
2.5. Alterations in Virulence Characteristics of P. aeruginosa Biofilm Cells
2.5.1. Swarming
2.5.2. Twitching
2.5.3. Quantification of Pyocyanin
2.6. Expression of the QS-Related Genes in Biofilms
2.7. Metabolomic Analysis
2.8. Statistical Analysis
3. Results
3.1. Effect of the Compounds and Ciprofloxacin on the Planktonic and Biofilm Cells of P. aeruginosa
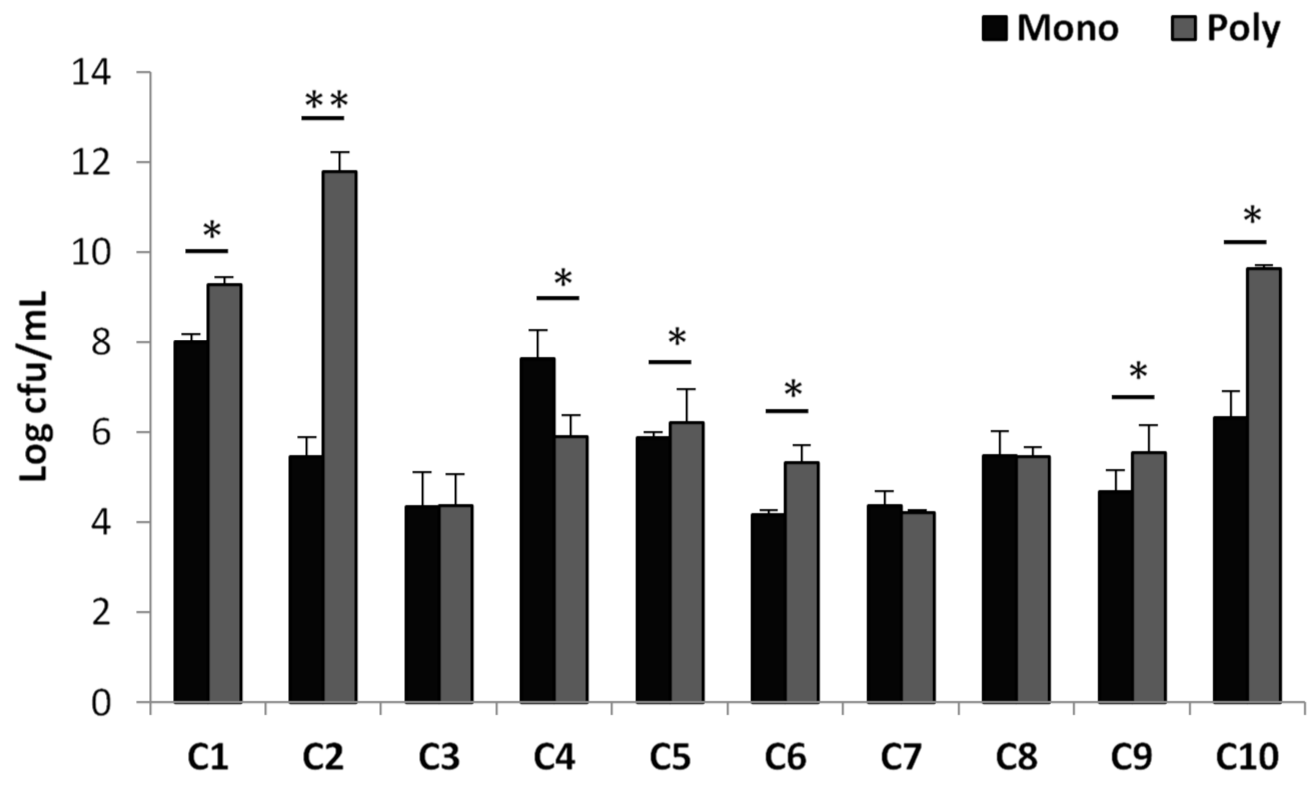
3.2. Comparison of the Effect of Compound–Ciprofloxacin Combinations with Sub-MBIC Values on Mono- and Triple-Species Biofilms of P. aeruginosa
3.3. Influence of the Compounds and Ciprofloxacin Combinations on Virulence Properties of P. aeruginosa
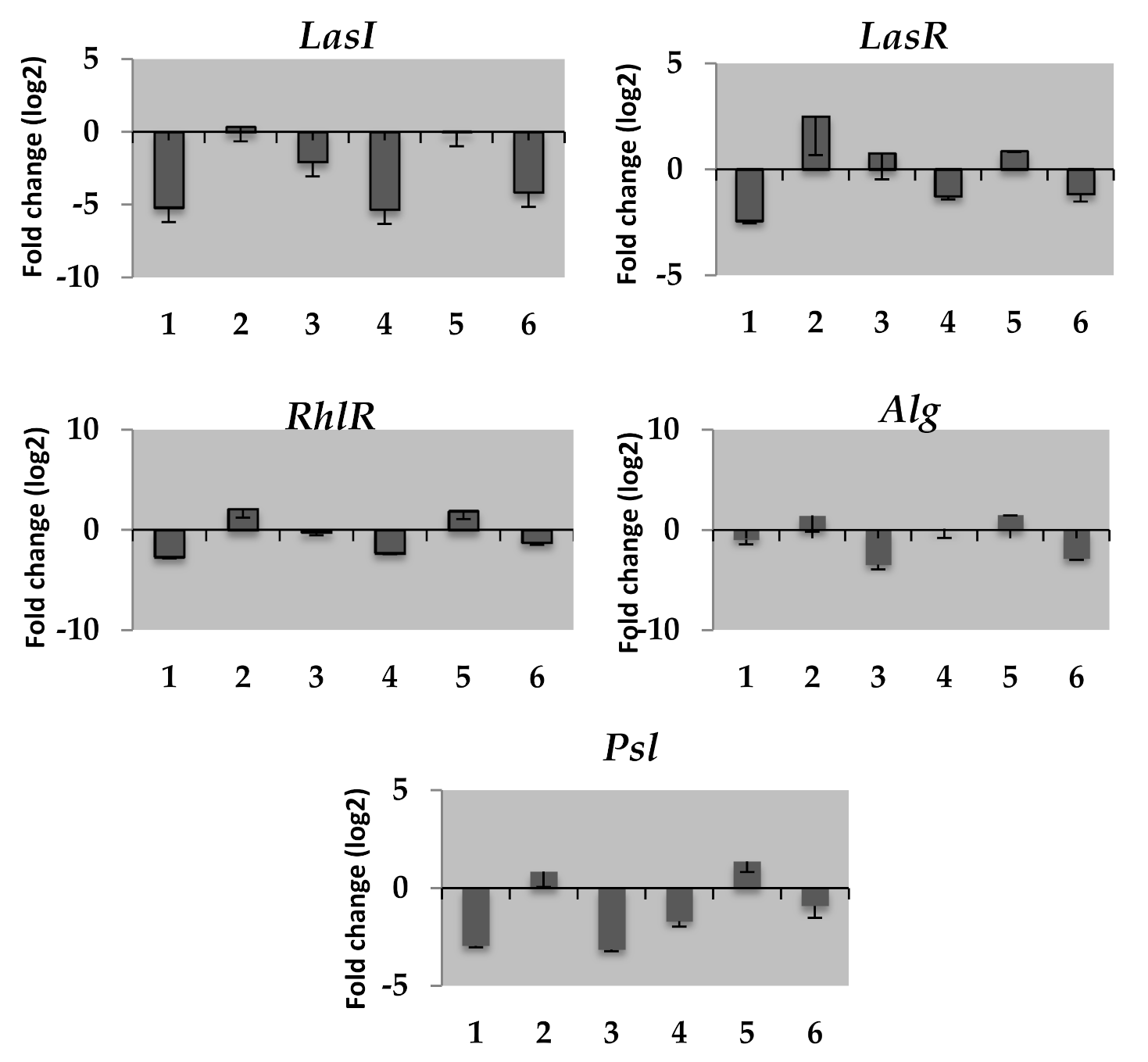
3.4. The Impact of Ciprofloxacin Combined with the Compounds on QS Gene Expressions of P. aeruginosa Biofilm Cells
3.5. Effect of Ciprofloxacin and Compounds Combinations on the Metabolic Profile of P. aeruginosa Biofilm Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tuon, F.F.; Gortz, L.W.; Rocha, J.L. Risk factors for pan-resistant Pseudomonas aeruginosa bacteremia and the adequacy of antibiotic therapy. Braz. J. Infect. Dis. 2012, 16, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Liu, Y.; Wu, H.; Hoiby, N.; Molin, S.; Song, Z.J. Current understanding of multi-species biofilms. Int. J. Oral Sci. 2011, 3, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Bayramov, D.F.; Neff, J.A. Beyond conventional antibiotics—New directions for combination products to combat biofilm. Adv. Drug Deliver Rev. 2017, 112, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Bassler, B.L. How bacteria talk to each other: Regulation of gene expression by quorum sensing. Curr. Opin. Microbiol. 1999, 2, 582–587. [Google Scholar] [CrossRef]
- Starkey, M.; Lepine, F.; Maura, D.; Bandyopadhaya, A.; Lesic, B.; He, J.X.; Kitao, T.; Righi, V.; Mi-lot, S.; Tzika, A.; et al. Identification of Anti-virulence Compounds That Disrupt Quorum-Sensing Regulated Acute and Persistent Pathogenicity. PLoS Pathog. 2014, 10, e1004321. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Hillaert, U.; Van Calenbergh, S.; Nelis, H.J.; Coenye, T. Use of quorum sensing inhibitors to interfere with biofilm formation and development in Burkholderia multivorans and Burkholderia cenocepacia. Res. Microbiol. 2009, 160, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazzini, R.A.B.; Skindersoe, M.E.; Bielecki, P.; Puchalka, J.; Givskov, M.; dos Santos, V.A.P.M. Protoanemonin: A natural quorum sensing inhibitor that selectively activates iron starvation response. Environ. Microbiol. 2013, 15, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Ulrey, R.K.; Barksdale, S.M.; Zhou, W.D.; van Hoek, M.L. Cranberry proanthocyanidins have anti-biofilm properties against Pseudomonas aeruginosa. BMC Complem. Altern. Med. 2014, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Kart, D.; Tavernier, S.; Van Acker, H.; Nelis, H.J.; Coenye, T. Activity of disinfectants against multi-species biofilms formed by Staphylococcus aureus, Candida albicans and Pseudomonas aeruginosa. Biofouling 2014, 30, 377–383. [Google Scholar] [CrossRef]
- Kart, D.; Ciftci, S.Y.; Nemutlu, E. Altered metabolomic profile of dual-species biofilm: Interactions between Proteus mirabilis and Candida albicans. Microbiol. Res. 2020, 230, 126346. [Google Scholar] [CrossRef]
- Reimmann, C.; Beyeler, M.; Latifi, A.; Winteler, H.; Foglino, M.; Lazdunski, A.; Haas, D. The global activator GacA of Pseudomonas aeruginosa PAO positively controls the production of the autoinducer N-butyryl-homoserine lactone and the formation of the virulence factors pyocyanin, cyanide, and lipase. Mol. Microbiol. 1997, 24, 309–319. [Google Scholar] [CrossRef]
- Cankirili, N.K.; Kart, D.; Celebi-Saltik, B. Evaluation of the biofilm formation of Staphylococcus aureus and Pseudomonas aeruginosa on human umbilical cord CD146+stem cells and stem cell-based decellularized matrix. Cell Tissue Bank. 2020, 21, 215–231. [Google Scholar] [CrossRef]
- Albano, M.; Crulhas, B.P.; Alves, F.C.B.; Pereira, A.F.M.; Andrade, B.F.M.T.; Barbosa, L.N.; Furlanetto, A.; Lyra, L.P.D.; Rall, V.L.M.; Fernandes, A. Antibacterial and anti-biofilm activities of cinnamaldehyde against S. epidermidis. Microb. Pathog. 2019, 126, 231–238. [Google Scholar] [CrossRef]
- Bahari, S.; Zeighami, H.; Mirshahabi, H.; Roudashti, S.; Haghi, F. Inhibition of Pseudomonas aeruginosa quorum sensing by subinhibitory concentrations of curcumin with gentamicin and azithromycin. J. Glob. Antimicrob. Resist. 2017, 10, 21–28. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafy, S.A.; Abd El Galil, K.H.; El-Messery, S.M.; Shaaban, M.I. Aspirin is an efficient inhibitor of quorum sensing, virulence and toxins in Pseudomonas aeruginosa. Microb. Pathog. 2014, 74, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Bag, A.; Chattopadhyay, R.R. Synergistic antibiofilm efficacy of a gallotannin 1,2,6-tri-O-galloyl-beta-D-glucopyranose from Terminalia chebula fruit in combination with gentamicin and trime-thoprim against multidrug resistant uropathogenic Escherichia coli biofilms. PLoS ONE 2017, 12, e0178712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.L.; Leonhard, M.; Moser, D.; Ma, S.; Schneider-Stickler, B. Antibiofilm efficacy of curcumin in combination with 2-aminobenzimidazole against single- and mixed-species biofilms of Candida albicans and Staphylococcus aureus. Colloid Surf. B 2019, 174, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Albano, M.; Alves, F.C.; Andrade, B.F.; Barbosa, L.N.; Pereira, A.F.; de Souza, M.D.; Rall, V.L.; Júnior, A.F. Antibacterial and anti-staphylococcal enterotoxin ac-tivities of phenolic compounds. Innov. Food Sci. Emerg. 2016, 38, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Lopez-Romero, J.C.; Oliveira, D.; Giaouris, E.; Simoes, M. Prevention, removal and inac-tivation of Escherichia coli and Staphylococcus aureus biofilms using selected monoterpenes of essential oils. J. Appl. Microbiol. 2017, 123, 104–115. [Google Scholar] [CrossRef]
- Miklasinska, M.; Kepa, M.; Wojtyczka, R.D.; Idzik, D.; Dziedzic, A.; Wasik, T.J. Catechin Hydrate Augments the Antibacterial Action of Selected Antibiotics against Staphylococcus aureus Clinical Strains. Molecules 2016, 21, 244. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rajaonson, S.; Diallo, B.; Mol, A.; El Jaziri, M.; Baucher, M. Identification of Catechin as One of the Flavonoids from Combretum albiflorum Bark Extract That Reduces the Production of Quorum-Sensing-Controlled Virulence Factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microb. 2010, 76, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, X.; Ling, J. Streptococcus gordonii LuxS/autoinducer-2 quorum-sensing system modulates the dual-species biofilm formation with Streptococcus mutans. J. Basic Microbiol. 2017, 57, 605–616. [Google Scholar] [CrossRef]
- Peters, B.M.; Ward, R.M.; Rane, H.S.; Lee, S.A.; Noverr, M.C. Efficacy of ethanol against Candida albicans and Staphylococcus aureus polymicrobial biofilms. Antimicrob. Agents Chemother. 2013, 57, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.Y.; Su, T.Y.; Wang, M.Y.; Yang, S.F.; Mar, K.; Hung, S.L. Inhibitory effects of tea catechin epigallocatechin-3-gallate against biofilms formed from Streptococcus mutans and a probiotic lactobacillus strain. Arch. Oral Biol. 2018, 94, 69–77. [Google Scholar] [CrossRef]
- Tan, Y.L.; Ma, S.; Leonhard, M.; Moser, D.; Haselmann, G.M.; Wang, J.; Eder, D.; Schneider-Stickler, B. Enhancing antibiofilm activity with functional chitosan nanoparticles targeting biofilm cells and biofilm matrix. Carbohyd. Polym. 2018, 200, 35–42. [Google Scholar] [CrossRef]
- Al Ahmar, R.; Kirby, B.D.; Hongwei, D.Y. Pyrimidine Biosynthesis Regulates the Small-Colony Variant and Mucoidy in Pseudomonas aeruginosa through Sigma Factor Competition. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Odonovan, G.A.; Neuhard, J. Pyrimidine Metabolism in Microorganisms. Bacteriol. Rev. 1970, 34, 278. [Google Scholar] [CrossRef]
- Beaume, M.; Kohler, T.; Fontana, T.; Tognon, M.; Renzoni, A.; van Delden, C. Metabolic pathways of Pseudomonas aeruginosa involved in competition with respiratory bacterial pathogens. Front. Microbiol. 2015, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Attila, C.; Whiteley, M.; Wood, T.K. Uracil influences quorum sensing and biofilm formation in Pseudomonas aeruginosa and fluorouracil is an antagonist. Microb. Biotechnol. 2009, 2, 62–74. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, D.M.; Wozniak, D.J. Understanding the control of Pseudomonas aeruginosa alginate synthesis and the prospects for management of chronic infections in cystic fibrosis. Mol. Microbiol. 2005, 56, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Trent, M.S. Biosynthesis and Export of Bacterial Lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Luengo, J.M.; Olivera, E.R. Catabolism of biogenic amines in Pseudomonas species. Environ. Microbiol. 2020, 22, 1174–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agents | MICs (mg/L) | MBICs (mg/L) | MBECs (mg/L) | ||
|---|---|---|---|---|---|
| Alone * | Alone ** | Combinations *** (CIP + Compounds) | Alone ** | Combinations *** (CIP + Compounds) | |
| Ciprofloxacin | 0.039 | 0.06 | - | 0.13 | - |
| Curcumin | 0.25 | 0.82 | 0.001 + 0.008 | >0.82 | 0.13 + 2 |
| Azathioprine | 0.25 | 0.82 | 0.008 + 0.06 | >0.82 | 0.13 + 2 |
| Resveratrol | 0.25 | 0.2 | 0.03 + 0.25 | >0.82 | 0.13 + 2 |
| Catechin hydrate | 0.51 | 0.82 | 0.03 + 0.26 | >0.82 | >0.13 + 2 |
| Baicalein | 0.13 | 0.2 | 0.001 + 0.008 | >0.82 | 0.13 + 2 |
| l-canavanine | 0.1 | 0.82 | 0.004 + 0.03 | >0.82 | 0.13 + 2 |
| 4-nitropyridine N-oxide | 0.02 | 0.41 | 0.008 + 0.06 | >0.82 | 0.13 + 2 |
| p-benzoquinone | 0.06 | 0.41 | 0.008 + 0.06 | >0.82 | 0.13 + 2 |
| Esculetin hydrate | 0.51 | 0.82 | 0.002 + 0.02 | >0.82 | 0.13 + 2 |
| Cinnamaldehyde | 0.26 | 0.41 | 0.002 + 0.02 | >0.82 | 0.13 + 2 |
| Curc. | Cinnam. | Curc. | Cinnam. | ||||
|---|---|---|---|---|---|---|---|
| Carbohydrate | Citric acid | 0.44↓ | - | Nitrogen | Pyroglutamic acid | 0.13↓ | 1.7↑ |
| Lactose | 0.32↓ | - | Aspartic acid | 0.03↓ | 2.08↑ | ||
| Glucose | 4.04↑ | - | Glutamic acid | 0.06↓ | - | ||
| Melibiose | 0.1↓ | 1.83↑ | N-acetyl-l-glutamic acid | 0.3↓ | - | ||
| Mannose | 1.78↑ | - | |||||
| Maltotriose | 0.02↓ | 2.92↑ | Lipid | Glyceric acid | 0.25↓ | - | |
| Maltotriitol | 0.01↓ | 2.61↑ | Glycine | 0.4↓ | - | ||
| Melezitose | 0.01↓ | 2.3↑ | Glycerol-phosphate | 0.35↓ | - | ||
| Glucuronic acid | 0.24↓ | - | Beta-glycerolphosphate | 0.42↓ | - | ||
| Amino Acid | Threonine | 0.43↓ | - | ||||
| Glyceraldehyde | 1.83↑ | - | |||||
| Proline | 0.24↓ | - | 3-phosphoglycerate | 0.04↓ | - | ||
| Methionine | 0.28↓ | - | |||||
| Lysine | 0.01↓ | 0.12↓ | Nucleotid | Uracil | 0.13↓ | 0.17↓ | |
| Phenylalanine | 0.49↓ | - | Uridine 5’-monophosphate | 1.5↑ | 0.02↓ | ||
| Tryptophane | 0.19↓ | Ribulose-5-phosphate | 0.23↓ | 0.06↓ | |||
| Amines | Phenylethylamine | 0.03↓ | 0.43↓ | ||||
| Pyrophosphate | 6.58↑ | - | |||||
| 2-amino-1-phenylethanol | 0.01↓ | 0.42↓ | Cytidine-5’-monophosphate | 0.35↓ | - | ||
| Tyramine | 0.14↓ | - | Porphine | 3.19↑ | - | ||
| Esculetin hyd | Baicalein hyd | ||
|---|---|---|---|
| Carbohydrate | Fumaric acid | - | 1.61↑ |
| Malic acid | - | 1.64↑ | |
| Melibiose | 1.51↑ | 1.71↑ | |
| Mannose | 1.47↑ | 1.83↑ | |
| Maltotriose | 2.08↑ | - | |
| Maltotriitol | 1.75↑ | - | |
| Melezitose | 1.56↑ | - | |
| Glucose-6-phosphate | - | 3.15↑ | |
| Amino Acid | Aspartic Acid | 2.8↑ | - |
| Lysine | 4.13↑ | - | |
| Tryptophane | 1.87↑ | 2.89↑ | |
| Nitrogen | Pyroglutamic Acid | 1.65↑ | - |
| Glutamic acid | 2.37↑ | - | |
| Amines | Phenylethylamine | 3.52↑ | 4.39↑ |
| 2-amino-1-phenylethanol | 12.83↑ | 1.47↑ | |
| Tyramine | 1.68↑ | 2.51↑ | |
| Lipid | Glycerol-phosphate | - | 1.79↑ |
| Beta-glycerolphosphate | - | 1.65↑ | |
| Nucleotid | Uracil | 6.11↑ | 11.01↑ |
| Uridine 5’-monophosphate | 0.41↓ | 3.81↑ | |
| Ribulose-5-phosphate | 1.38↑ | 9.27↑ | |
| Pyrophosphate | 1.49↑ | 5.89↑ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kart, D.; Reçber, T.; Nemutlu, E.; Sagiroglu, M. Sub-Inhibitory Concentrations of Ciprofloxacin Alone and Combinations with Plant-Derived Compounds against P. aeruginosa Biofilms and Their Effects on the Metabolomic Profile of P. aeruginosa Biofilms. Antibiotics 2021, 10, 414. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040414
Kart D, Reçber T, Nemutlu E, Sagiroglu M. Sub-Inhibitory Concentrations of Ciprofloxacin Alone and Combinations with Plant-Derived Compounds against P. aeruginosa Biofilms and Their Effects on the Metabolomic Profile of P. aeruginosa Biofilms. Antibiotics. 2021; 10(4):414. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040414
Chicago/Turabian StyleKart, Didem, Tuba Reçber, Emirhan Nemutlu, and Meral Sagiroglu. 2021. "Sub-Inhibitory Concentrations of Ciprofloxacin Alone and Combinations with Plant-Derived Compounds against P. aeruginosa Biofilms and Their Effects on the Metabolomic Profile of P. aeruginosa Biofilms" Antibiotics 10, no. 4: 414. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040414